Introduction
There is increasing interest in the use of herbal
medicines for the treatment and/or prevention of a variety of
diseases (1,2). Musa basjoo (MB) has been used
as a traditional herb, grown for the treatment of inflammatory
conditions accompanying high fever (3,4). MB
is classified as one of the 50 species belonging to the genus
Musa that includes edible banana species, such as Musa
paradisiaca and Musa sapientum (5). In addition, the fiber of Musa
balbisiana has been used as a material for clothing termed
Bashofu from ancient times in Japan (3).
Previous findings showed that the compounds isolated
from herbs have a broad range of biological activities including
anticancer and/or cancer preventive properties (6,7). In
fact, the dietary administration of powdered leaves of
Peucedanum japonicum and Terminalia catappa reduce
the occurrence of azoxymethane-induced aberrant crypt foci,
preneoplastic lesions in rat colon carcinogenesis (8,9). In
addition, growth inhibitory activities of crude extracts obtained
from herbal plants in the Ryukyu Islands on several human
colorectal cancer cell lines have been identified (10).
Although the above-mentioned findings obtained with
herbal plants in experimental studies are promising, the molecular
mechanisms by which the crude extracts of MB exert anticancer
effects have yet to be clarified. Even if such herbal products are
often perceived as being of natural origin and therefore harmless,
investigation of their molecular mechanisms of action and their
specific cellular targets is crucial to predict possible side
effects. Furthermore, this information is useful for developing and
designing new drugs for more effective treatment and prevention of
cancer and other diseases. Based on the findings of a previous
study (11), authors of the
present study were interested in examining the crude extracts of MB
in the p53 wild-type and p53 mutant human colorectal cancer cell
lines. Thus, the aim of the present study was to determine whether
crude extracts of MB contain compounds that may inhibit the growth
of human colorectal and other types of cancer cell lines.
Materials and methods
Crude dried leaf extracts of MB
Samples of MB in the current study were collected
from the Herbal Garden, Gifu Pharmaceutical University, Gifu,
Japan. In brief, the dried leaf with blade and stalk was extracted
with acetone or methanol, filtered and evaporated under reduced
pressure to remove the acetone or methanol, as described in a
previous study (10). Remaining
crude extracts were dissolved in dimethylsulfoxide (DMSO)
(Sigma-Aldrich) and stored at −20°C until use. Each extract
(acetone extract sample and methanol extract sample) was used for
the biological assays described below.
Thin layer chromatography
Thin layer chromatography (TLC) was performed with
silica gel 60 F254 (1.05715.0001; Merck KGaA) to analyze
crude extracts of MB. The extracts were developed with ethyl
acetate (AcOEt):n-hexane at 1:3 for 9 cm. Then, the
resulting chromatogram was visualized with a handheld UV (254/365
nm) lamp (UVGL-58; Analytik Jena AG) equipped with Chromato-Vue
Cabinet (C-10; Analytik Jena AG) followed by treatment with 5%
phosphomolybdic acid hydrate in ethanol solution and then heating
to enhance visualization [phosphomolybdic acid staining at 200°C
for enough time to distinguish coloration (~1 min)].
Cell lines and treatment with the
crude extracts of MB
HT29 (HTB38) and HCT116 (CCL247) human colorectal
cancer, HepG2 (HB8065) liver cancer/hepatoma, MCF-7 (HTB22) human
breast cancer, and PC-3 (CRL1435) human prostate cancer cell lines
[American Type Culture Collection (ATCC)] were maintained in
Dulbecco's modified Eagle's medium (DMEM) (FUJIFILM Wako Pure
Chemical Corp.) supplemented with 5% (v/v) fetal bovine serum (FBS)
(Biowest) in an incubator with humidified air at 37°C with 5%
CO2. Cells were plated in 10-cm culture dishes, treated
with the indicated concentrations of the crude acetone or methanol
extracts, and harvested at the indicated times. As an untreated
solvent control, the cells were treated with DMSO at a final
concentration <0.5%.
Cell proliferation assays
Cell proliferation was measured by both the colony
formation assay (HT29, HCT116, HepG2, MCF-7 and PC-3 cell lines)
and MTT assay (HT29 and HCT116 cell lines) as described elsewhere
(12,13). The MTT assay was performed to
confirm the results of the colony formation assay. Experiments were
performed at an optimal timing point, at which cells were dividing
and did not reach a plateau. Cells were plated into 6-well 35-mm
diameter culture plates (5×102 cells per well) and
treated with different concentrations (12.5-200 µg/ml) of crude
acetone or methanol extracts of MB at 37°C for 7 days in DMEM plus
5% FBS. After washing with phosphate-buffered saline (PBS),
colonies were stained with Giemsa solution (Sigma-Aldrich) and then
counted. The clonogenicity of cancer cells was tested using the
colony formation assay. Results were expressed as the percentage of
the control untreated culture. Each concentration of the extracts
was tested at least in duplicate. The relative surviving fraction,
when compared with cells treated with the vehicle, was plotted on
the dose-response curve. A viability of 100% corresponded to the
control cells.
Cells were plated onto 96-well plates
(5×103 cells per well) and cultured overnight to allow
for cell attachment. Subsequently, these cells were treated with
increasing concentrations (25–400 µg/ml) of crude acetone or
methanol extracts of MB, grown in DMEM containing 5% FBS at 37°C
for 96 h, and assayed using an MTT assay kit (Cytiva). The growth
inhibitory activity of the extracts of MB was examined by the MTT
assay. Each concentration of the extracts was tested in duplicate.
Solubilization buffer (10% SDS in 0.01 M HCl) was used to dissolve
the formazan. The quantity of formazan product was measured using a
spectrophotometric microplate reader (Bio-Rad Laboratories Inc.) at
595 nm wavelength. A viability of 100% corresponded to the control
cells.
Flow cytometry
Flow cytometric analysis was performed as described
previously (13,14). HT29 and HCT116 cells were plated
onto 10-cm dishes (5×105 cells per dish) in DMEM
containing 5% FBS and grown at 37°C overnight to allow for cell
attachment. Cells were then treated with DMSO (<0.5%) or 100
µg/ml crude acetone extract of MB at 37°C for 96 h, harvested,
fixed with 70% ethanol, centrifuged (750 × g, at room temperature
for 5 min), resuspended in 400 µl of PBS containing 2 mg/ml RNase
(Nacalai Tesque Inc.), and stained with 400 µl of 0.1 mg/ml
propidium iodide (Nacalai Tesque). The cell suspension was filtered
through a 40 µm nylon filter (Ikemoto Scientific Technology Co.
Ltd.). Samples of 20,000 cells suspended in BD FACSFlow™ Sheath
Fluid at room temperature were then analyzed for DNA histograms
(<1 min/run) and cell cycle phase distributions by flow
cytometry using a FACSCalibur instrument (BD Biosciences), and the
data were analyzed by a CELLQuest computer program (BD
Biosciences), as previously described (13–15).
Each assay was repeated in triplicate to confirm the results.
Western blot analysis
The assay was carried out as described previously
(13,16). HT29 and HCT116 cells were treated
with DMSO (<0.5%), 50 or 100 µg/ml acetone extract of MB at 37°C
for 96 h and harvested. Then, the cells were lysed with modified
radioimmunoprecipitation assay (RIPA) buffer [150 mM NaCl, 1% NP40,
0.1% SDS, 50 mM Tris-HCl (pH 8.0), 0.5% deoxycholic acid, 1 mM
EDTA, 2 mM EGTA, 1 mM DTT, and 25% glycerol]. Cell lysates (40 µg
per lane) were separated by SDS-PAGE (12.0-13.5% gel) and
transferred onto an Immobilon-P transfer membrane (Merck KGaA).
Immunoblots using monoclonal or polyclonal antibodies were then
prepared by established methods (12). Primary antibodies used in this
study included cyclin D1 (06–137; Merck KGaA; 1:1,000), cyclin E
(sc-377100; Santa Cruz Biotechnology Inc.; 1:100),
p21CIP1 (sc-817; Santa Cruz Biotechnology Inc.; 1:200),
p27KIP1 (610242; BD Biosciences; 1:2,500), p53 (sc-126;
Santa Cruz Biotechnology Inc.; 1:200), cdk2 (sc-163; Santa Cruz
Biotechnology Inc.; 1:200), cdk4 (sc-260; Santa Cruz Biotechnology
Inc.; 1:200), poly (ADP-ribose) polymerase (PARP; 9542; Cell
Signaling Technology, Inc.; 1:1,000), and β-actin (sc-1616-R; Santa
Cruz Biotechnology Inc.; 1:200). Anti-mouse IgG, Horseradish
peroxidase-linked species-specific F(ab')2 fragment
(NA9310; Cytiva; 1:4,000) or anti-rabbit IgG, horseradish
peroxidase-linked species-specific whole antibody (NA934; Cytiva;
1:3,000) antibodies were used as the secondary antibodies. The
membranes were then incubated at room temperature for 1 h. Each
membrane was developed using an ImmunoStar Long Detection System
(FUJIFILM Wako Pure Chemical Corp.) and visualized with
Light-CaptureII imaging analyzer (ATTO Corp.). Each assay was
repeated to confirm the results. Densitometry was performed to
obtain relative band intensity using ImageJ computer program (Image
J software, National Institute of Health) as described earlier
(11,13,15).
Tumor xenograft assay
A total of 8 female BALB/cSlc-nu/nu mice aged 6
weeks and weighing 18.8±0.22 g were purchased from Japan SLC, Inc.
Dose level (2 mg/kg) was determined based on the results of our
preliminary experiments (unpublished data). All 8 mice were
quarantined for 1 week and housed in plastic cages (4 mice/cage)
with free access to tap water and basal diet (MF diet; Oriental
Yeast Co., Ltd.) under controlled conditions of humidity (50±10%),
temperature (23±2°C) and lighting (12 h light/12 h dark cycle; 8 am
light on, 8 pm light off). All the animal experiments were
performed with the approval of the Animal Ethics Committee of the
Nagoya City University (approval no. H28M-03) and according to the
guidelines of the committee.
Viable HT29 human colorectal cancer cells
(2.5×106 cells/200 µl DMEM without L-glutamine and
phenol red) were subcutaneously injected into the flank of all 8
mice. After confirming the visible tumor mass, the mice were
assigned into two experimental groups (4 mice in each group). Crude
acetone extract was dissolved into saline containing 20% ethanol.
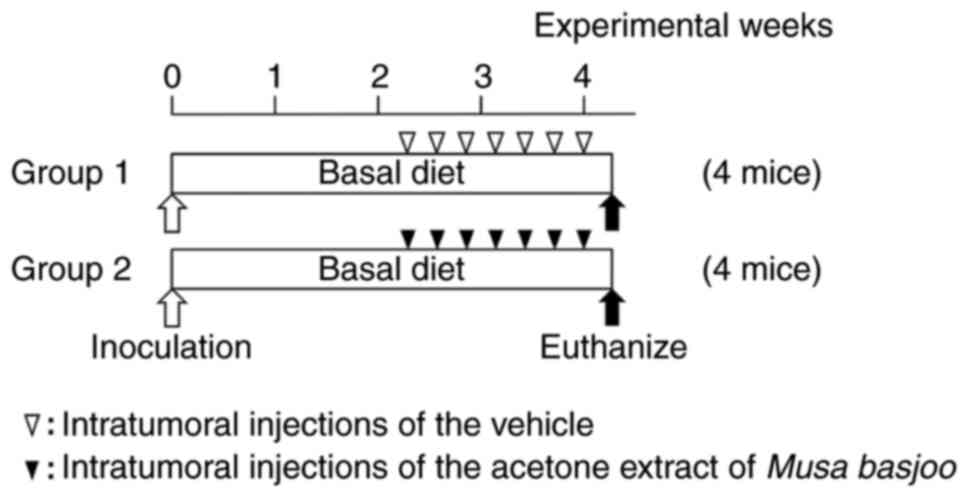
As shown in Fig. 1, mice in group
1 (control) and group 2 (treatment) received intratumoral
injections of the vehicle (saline containing 20% ethanol) and crude
acetone extract of MB (2 mg/kg), respectively, every other day (a
total of 7 times). The mice were observed on a daily basis for
tumor growth, body weight, and symptomatic adverse side effects.
Tumors were measured three times a week. The mean volume per tumor
was calculated using the formula: V
(mm3)=axb2/2 where V is the volume, and a is
the maximum and b is the minimum diameter of the tumor (17). At the 4th experimental week, all 8
mice were euthanized by decapitation following anesthesia with 3%
isoflurane and complete autopsy was performed. Tumors and main
organs (heart, lung, liver, and kidney) were carefully removed,
fixed with 10% buffered formalin and processed for
histopathological examination and [hematoxylin and eosin (HE)
staining]. Furthermore, we measured the necrosis area in the HE
tumor sections using an imaging system (Digital microscope
VHX-5000; Keyence Corp.). The experiment was terminated when a
tumor reached 20 mm or 2 cm in dimension, or when the study period
finished.
Statistical analysis
Comparisons between the vehicle-treated control
group and the acetone extract-treated group were made using one-way
ANOVA (western blot analysis) or two-way ANOVA (xenograft assay),
and Tukey's multiple comparison test was applied to evaluate
statistical significance. IBM SPSS Statistics version 24 was used
to evaluate the data. Student's unpaired t-test was used to compare
two groups with normal data distribution and homogenous variances.
If variances were heterogenous, Welch's t-test was used.
Mann-Whitney U test was used to compare two groups with non-normal
data distribution. Results were expressed as means ± SEs.
Differences between groups of P<0.05 were considered
statistically significant.
Results
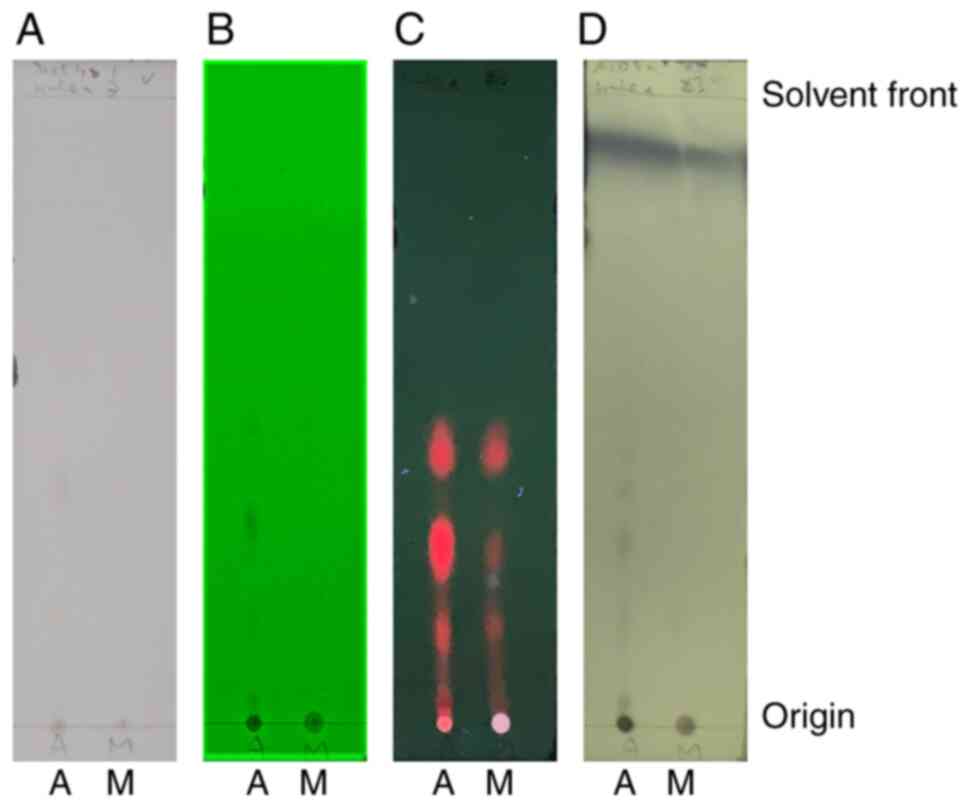
Crude extracts of MB exhibited the
difference in migration of spots on TLC
To examine dissimilar contents of the possible
active components contained in both acetone and methanol extracts,
TLC was performed. Although both extracts had the same several
fluorescent spots, visualized with 365 nm UV lamp irradiation, the
acetone extract had specific spots migrated marginally from its
origin [Retardation factor (Rf): 0.03] (Fig. 2). These unique spots were
visualized with both 254 nm UV lamp irradiation and phosphomolybdic
acid staining, suggesting that they are aromatic compounds.
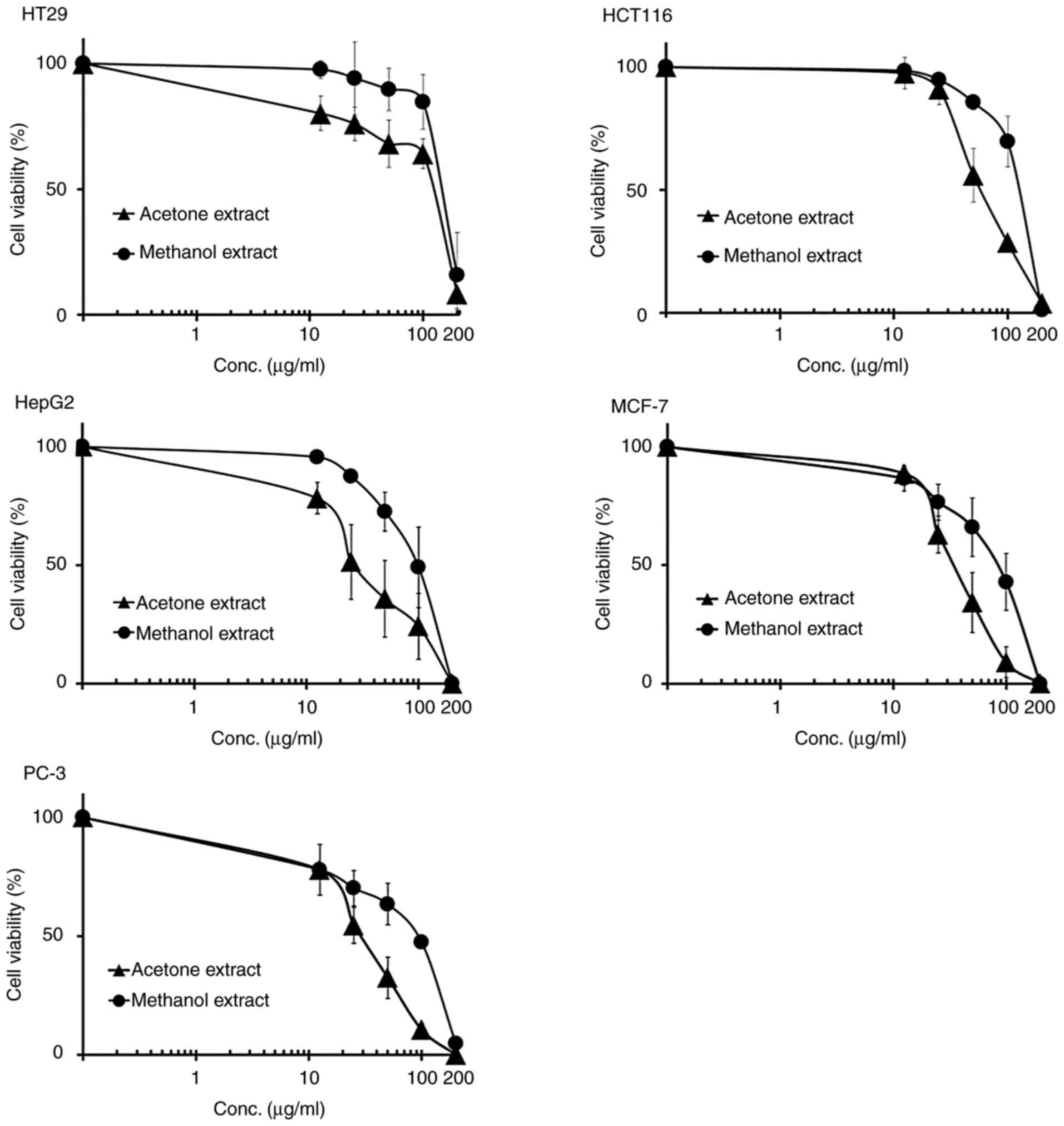
Crude extracts of MB inhibited the
proliferation of five human cancer cell lines
To examine the antiproliferative activity of acetone
or methanol extracts in a variety of human cancer cell lines,
colorectal (HT29 and HCT116) and other types (HepG2, MCF-7 and
PC-3) of human cancer cell lines were examined. Exponentially
dividing cells were treated with increasing doses of the MB
extracts (12.5-200 µg/ml for colony assays, 25-400 µg/ml for MTT
assays). In these five cell lines, both acetone and methanol
extracts inhibited cell proliferation, in a dose-dependent manner,
with IC50 values in the range of 29-136 µg/ml (acetone
extract) and 85-175 µg/ml (methanol extract) (Table I and Fig. 3). Representative images of culture
plates (HCT116 and MCF-7 cell lines) of colony formation assay are
shown in Figure S1. Additional
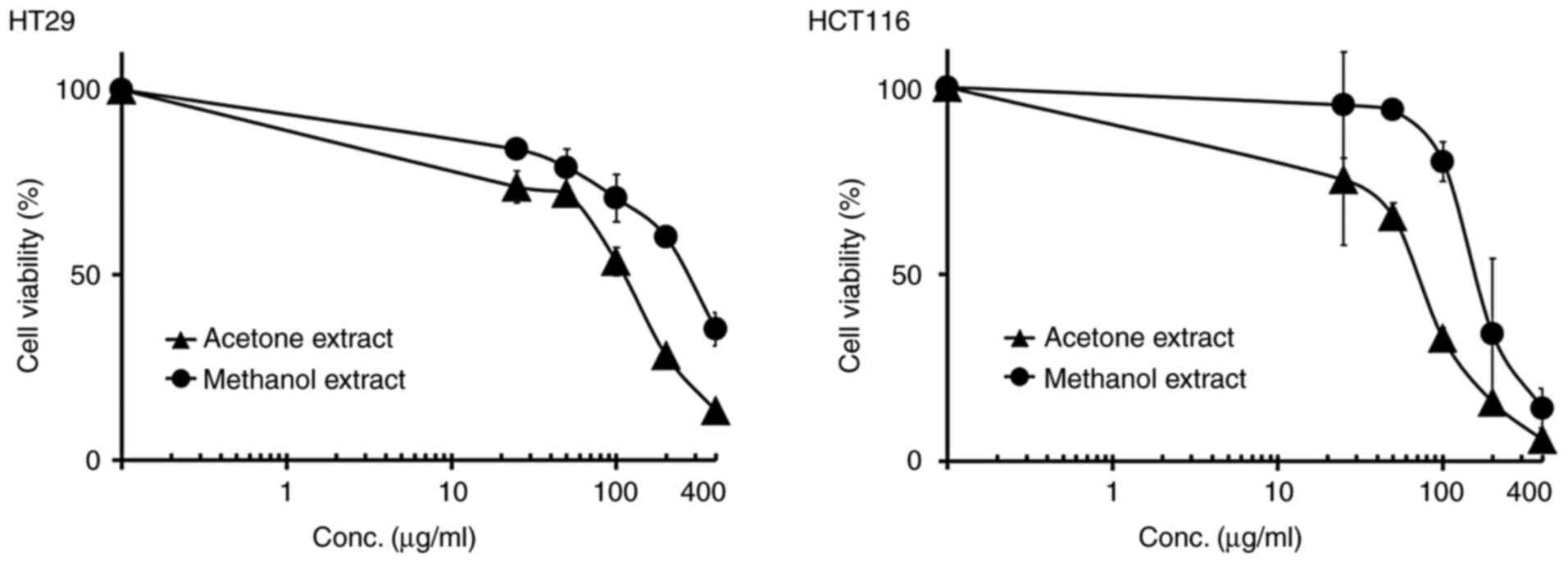
studies were performed with the HT29 and HCT116 cell lines to
confirm the growth inhibition by crude extracts of MB using the MTT
assays. In the HT29 cell line, both acetone and methanol extracts
of MB showed marked inhibition of the growth of these cells, with
IC50 values of ~126 and 260 µg/ml, respectively
(Table II and Fig. 4). In the HCT116 cell line, both
acetone and methanol extracts decreased the growth of cells in a
dose-dependent manner, with IC50 values of about 68 and
216 µg/ml, respectively (Table II
and Fig. 4). As a result of the
colony formation assay and MTT assay, the acetone extract exhibited
stronger antiproliferative activity than the methanol extract
(Tables I and II).
 | Table I.Effects of the extracts of Musa
basjoo. |
Table I.
Effects of the extracts of Musa
basjoo.
|
| IC50
(µg/ml) |
|---|
|
|
|
|---|
| Cell line | Acetone
extract | Methanol
extract |
|---|
| HT29 | 136±9.5 | 175±17.0 |
| HCT116 |
51±10.8a | 137±10.3 |
| HepG2 | 45±16.2 | 102±17.0 |
| MCF-7 | 40±12.7 | 85±12.3 |
| PC-3 | 29±10.7 | 85±11.3 |
| Table II.Effects of the extracts of Musa
basjoo. |
Table II.
Effects of the extracts of Musa
basjoo.
|
| IC50
(µg/ml) |
|---|
|
|
|
|---|
| Cell line | Acetone
extract | Methanol
extract |
|---|
| HT29 |
126±4.3a | 260±6.4 |
| HCT116 | 68±17.4 | 216±20.3 |
Acetone extract of MB caused an
increase in cells of G1-phase in the colorectal cancer cell
line
In view of the above-mentioned growth inhibitory
effects, it was of interest to examine the effects of the acetone
extract on cell cycle progression in exponentially dividing
cultures of the HT29 and HCT116 cell lines. Cells were treated with
either DMSO (control) or 100 µg/ml acetone extract at 37°C for 96
h. Since we were interested in simultaneously demonstrating cell
cycle arrest and changes in protein expression in each cell line, a
single concentration of 100 µg/ml was selected for flow cytometry
or two different concentrations of 50 and 100 µg/ml for western
blot analysis. The dose 100 µg/ml is almost the IC50
value of these two cell lines. A representative histogram for the
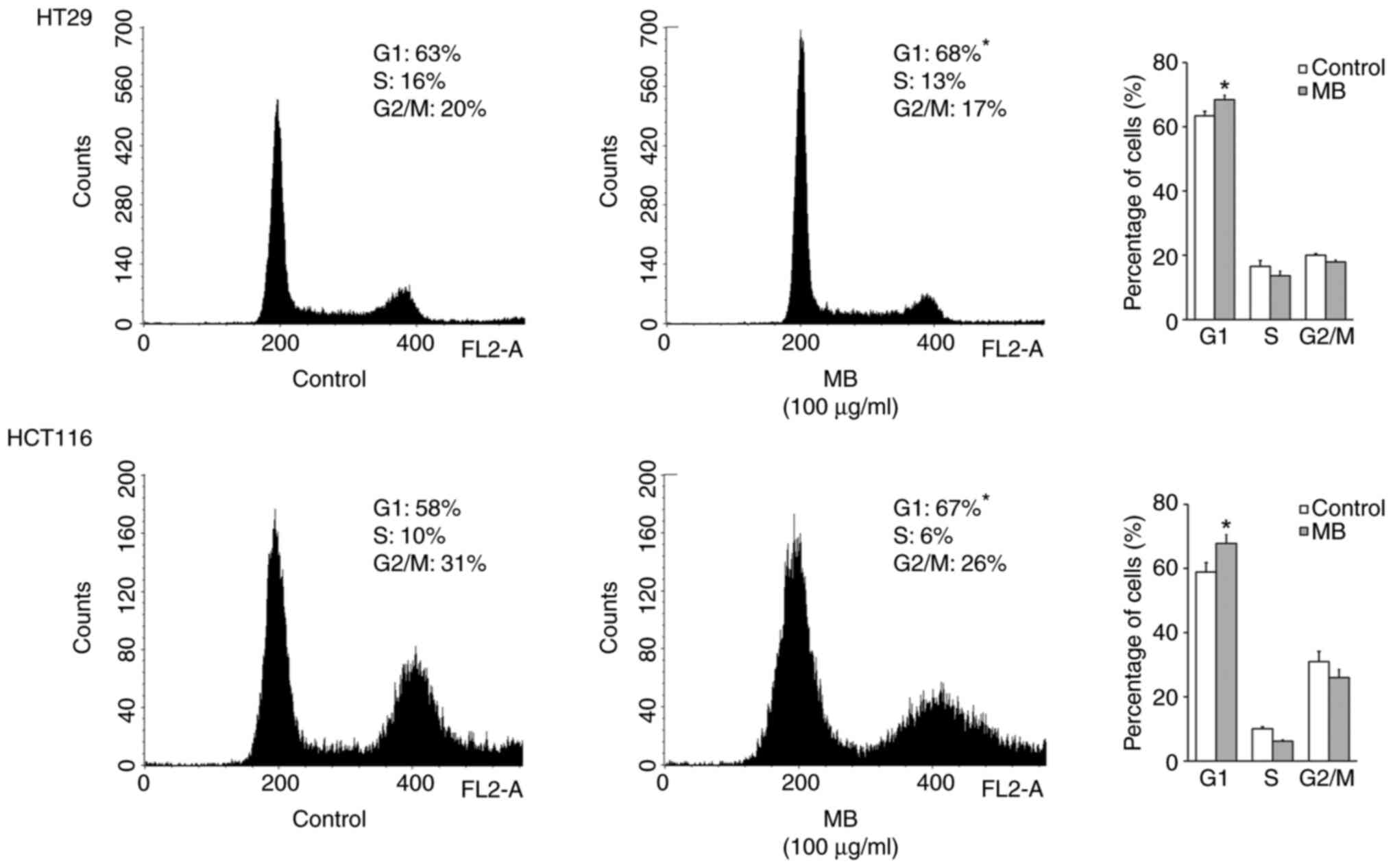
HT29 and HCT116 cell lines is shown in Fig. 5. Flow cytometric analysis indicated
that when cells were treated with the indicated concentration of
acetone extract of MB, the percentage of the HT29 and HCT116 cell
lines in G1 significantly increased by 5 and 9%, respectively,
after 96 h of treatment, and this was associated with a concomitant
decrease of cells in the S and/or G2-M phases of the cell cycle.
There was no evidence of apoptosis by an increase in the sub-G1
population of DNA when cancer cells were treated with 100 µg/ml of
acetone extract of MB.
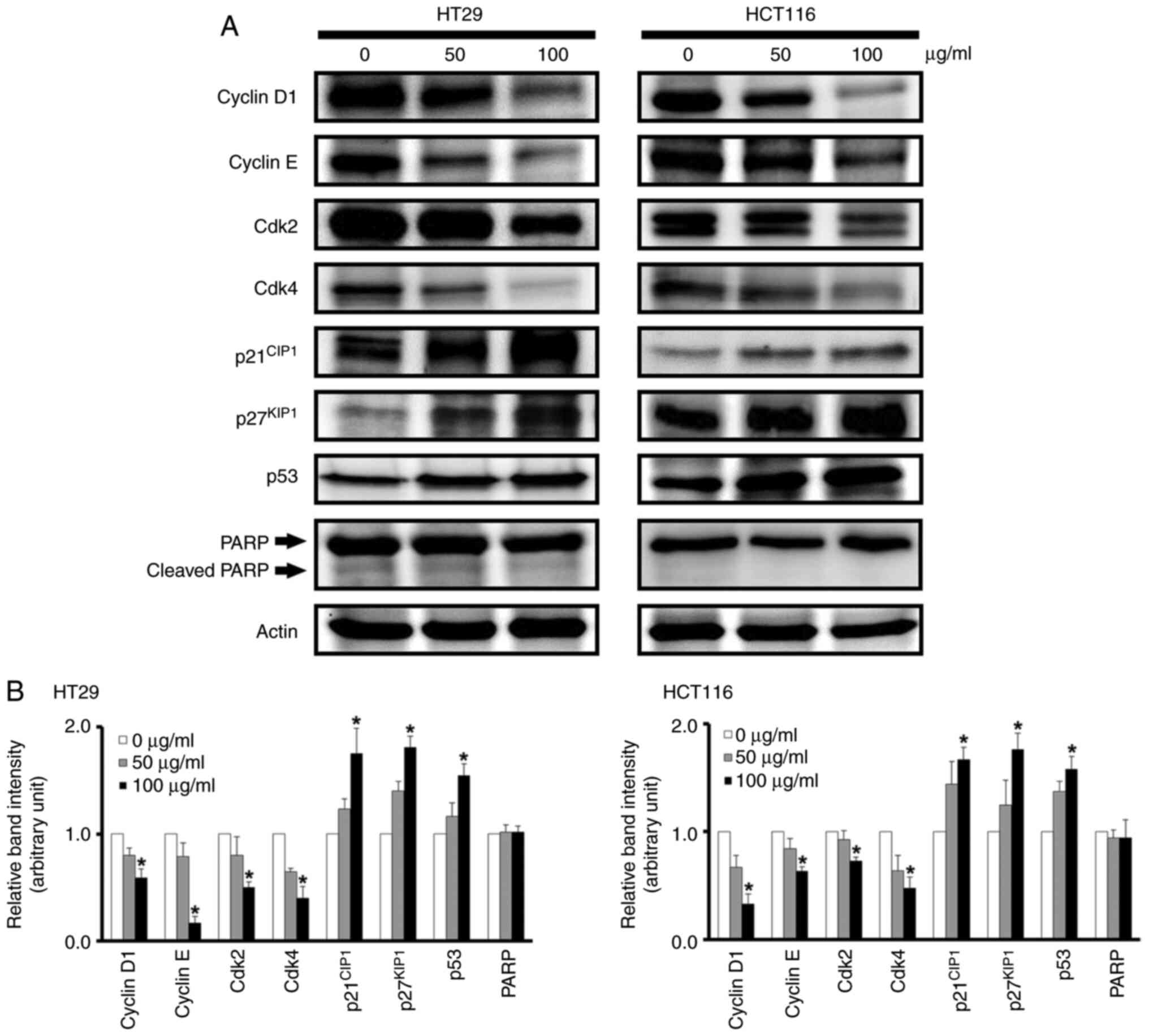
Acetone extract of MB causes an
increase in the protein expression levels of p21CIP1,
p27KIP1, and p53 and a decrease in those of cyclin D1,
cyclin E, cdk2, and cdk4
As the acetone extract induced a G1 arrest in the
cell cycle (Fig. 5), western blot
analysis was performed to determine whether the treatment of HT29
and HCT116 cells with the acetone extract of MB alters the cellular
expression levels of the G1 cell cycle control proteins, cyclin
D1/cdk4 (early G1 phase) and cyclin E/cdk2 (late G1 phase), and the
cell cycle inhibitor proteins p21CIP1,
p27KIP1, and p53. Investigation of the acetone extract
of MB in the p53 wild-type HCT116 and p53 mutant HT29 cell lines
was also conducted, as presented in a previous study (11). Thus, these two colorectal cancer
cell lines were chosen for the analysis. The results revealed that
when these two cell lines were treated with the indicated
concentrations (50 and 100 µg/ml) of the acetone extract of MB at
37°C for 96 h, there was a marked increase in the
p21CIP1, p27KIP1, and p53 proteins (Fig. 6). There was also a marked
inhibition in expression levels of the cyclin D1, cyclin E, cdk2,
and cdk4 proteins (Fig. 6). The
sub-G1 population of DNA was not detected in the flow cytometric
analysis (Fig. 5). Next, the
expression level of apoptosis-associated molecule PARP was
examined. Western blot analysis demonstrated that the expression
level of PARP did not change and cleaved PARP was not detected
after HT29 and HCT116 cells were treated with 50 and 100 µg/ml MB
extract.
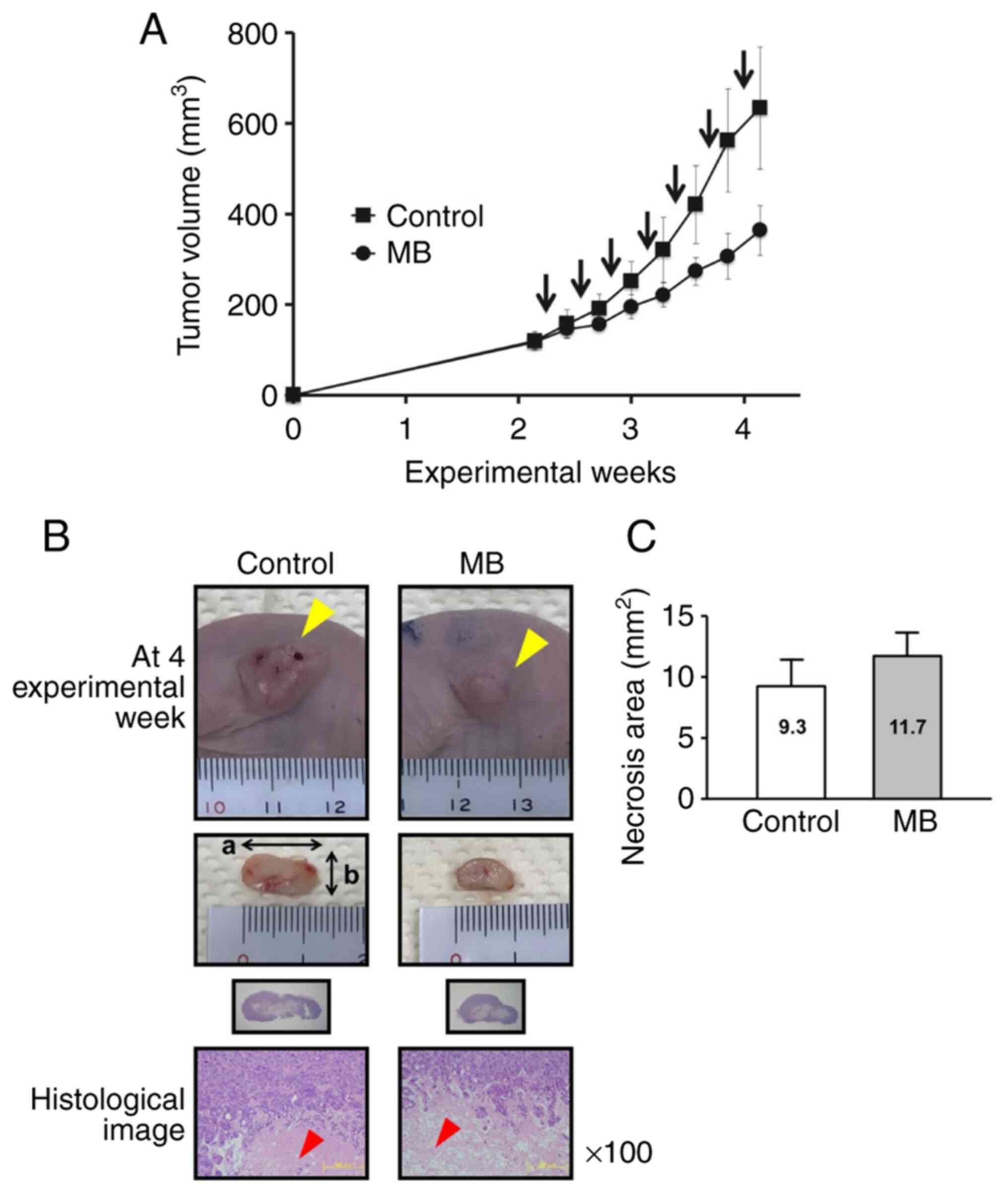
Tendency for acetone extract of MB to
inhibit xenograft tumor growth in BALB/cSlc-nu/nu mice
At the end of the experiment, the treatment of mice
with acetone extract of MB caused a decrease by 42% in tumor
volume, but this effect was not statistically significant (Fig. 7A). There was a tendency of an
increase in the necrosis area (mm2) of the MB treatment
group when compared to that of the control group (Fig. 7B and C). At the end of the
experiment, there was no statistically significant difference in
body, liver, kidney, relative liver, and relative kidney weights
between control and MB treatment groups (Table III). Additionally, there were no
symptomatic side effects evident between the control and MB
treatment groups during the experimental period. No histologically
significant adverse side effects were observed in heart, lung,
liver, and kidney (Fig. S2).
| Table III.Body, organ, and relative organ
weights of mice at the end of the experiment. |
Table III.
Body, organ, and relative organ
weights of mice at the end of the experiment.
| Group no. | Effective no. of
mice | Treatment | Body weight
(g) | Liver weight
(g) | Relative liver
weighta | Kidney weight
(g) | Relative kidney
weightb |
|---|
| 1 | 4 | None | 19.15±0.56 | 0.95±0.04 | 0.05±0.0004 | 0.36±0.01 | 0.03±0.0004 |
| 2 | 4 | Acetone extract of
MB | 19.33±0.33 | 1.00±0.04 | 0.06±0.001 | 0.35±0.01 | 0.02±0.001 |
Discussion
Although MB has previously been shown to have
biological activities (3,4), there are no experimental data
characterizing its anticancer properties during human consumption.
Current studies have provided further evidence to support the
empirical use of MB and identify a potential active component of MB
for evaluating its in vivo treatment and/or prevention
efficacy. The present study demonstrated the broad anticancer
properties of MB in a wide variety of human cancer cell lines. Both
the acetone and methanol extracts of MB caused a dose-dependent
cancer cell growth inhibition, indicating that MB contains
component(s) that have antiproliferative activity in these cell
lines. In the human colorectal cancer cell lines, this
antiproliferative activity appears to be due to its ability to
induce G1-phase arrest by causing a decrease in the cellular levels
of the cyclin D1, cyclin E, cdk2, and cdk4 proteins. In the current
study, acetone and methanol extracts were examined for their growth
inhibitory activity using cell proliferation assays and the results
showed that the acetone extract exerted stronger growth inhibition
than the methanol extract in the cell lines tested. The differences
observed between the two extracts are presumably due to the
dissimilar contents of the active components contained in the
extracts. The finding that the acetone extract may possibly contain
components that inhibit the growth of cancer cells more effectively
than the methanol extract requires further investigation.
In addition, HCT116 cells were found to be more
sensitive to growth inhibition in the acetone/methanol extracts
than HT29 cells. As similar results concerning different
sensitivity in several human colorectal cancer cell lines were
previously identified (10), the
above-mentioned findings may provide clues as to why susceptibility
to growth inhibition differs depending on the cell lines.
p21CIP1 is a tumor suppressor protein and acts as an
inhibitor of cell cycle progression by negatively regulating the
activity of cyclin D1-cyclin-dependent kinase (CDK) complex via a
p53-dependent or -independent mechanism (18). In the current study, the levels of
expression of p21CIP1 and p53 proteins in the p53
wild-type HCT116 and p53 mutant HT29 cell lines were examined. The
acetone extract of MB caused an increase in the cellular levels of
the p21CIP1 and p53 proteins. The results presumably
indicate that p53 status, at least in part, is critical to the
antiproliferative activity in HCT116 cells, as seen in a previous
study (11). The reason for which
the p53 mutant cell line HT29 is sensitive to the treatment of the
MB extract requires further studies. It is known that the p53
family consists of three proteins, p53, p63 and p73 that are
homologous at the amino acid level (19). Both p63 and p73 use two different
promoters, resulting in the expression of two isoforms with
different N-terminal domains, including transactivation domain (TA)
and ΔN (amino-truncated) isoforms (20). Despite the high frequency of p53
mutations, p63 and p73 genes are rarely mutated in cancer (20). In vivo models indicated that
the TA isoforms of p63 and p73 are tumor suppressors since the loss
of these isoforms causes spontaneous tumor formation (21,22).
In addition, p63 isoforms induced apoptosis in BHK cells
transfected with wild-type p53, mutant p53 or p63 isoforms
(23).
There are more than 50 species in genus Musa
(3). The constituents of a plant
may depend on the phytogeographical factors including plant
sources, specific location of plant distribution, and seasonal time
periods of the harvest (24–28).
These factors may influence the biological activities and amount of
active component present in the plant. Thus, it is of interest to
investigate whether the chemical composition of various Musa
species differs depending on the above-mentioned factors. Related
to this fact, we had previously reported such discrepancy in the
growth inhibition of cancer cells via investigation of the ethanol
extracts between Brazilian and Chinese propolis (11). Furthermore, to evaluate the
possibility of the clinical application of MB, it is necessary to
identify the specific component present in the extracts of MB that
cause growth inhibitory activity in human cancer cells, and to
determine the precise chemical structure of this compound. As for
active components obtained from rhizomes of MB, three known
compounds 4-(4′-hydroxyphenyl)-2-methoxyphenalen-1-one,
2-phenyl-naphthalic anhydride, and 1,7-bis(4-hydroxyphenyl)
hepta-4(E), 6(E)-dien-3-one exhibited significant
cytotoxicities with IC50 values of 23-28, 6.5-18, and
3.7-11 µM against human cancer cell lines, respectively (29). In the present study, the dried leaf
with a leaf blade and a stalk was extracted with acetone or
methanol, a TLC-based simple identification method was performed,
and the results showed that the acetone extract had specific spots
visualized with UV lamp irradiation (254 and 365 nm) and
phosphomolybdic acid staining. The results indicated that the crude
extract of MB may contain aromatic compounds with a certain number
of conjugated double bond (30–35)
and/or an antioxidant compound with a hydroxy group (36,37),
suggesting polyphenols and/or flavonoids as potential active
components. The extract of Chinese propolis contains polyphenols
and flavonoids (38–40). Previous findings demonstrated that
the extract of Chinese propolis caused a marked growth inhibition
on the HCT116 and HT29 human colorectal cancer cell lines but only
a marginal growth inhibition on the FHC normal human colonic
epithelial cell line (11).
Therefore, cancer cell-specific effects may be caused by the MB
treatment; however, this aspect should be further investigated.
Limitations of the present study include the lack of data
elucidating cell proliferation in the tumor tissue (e.g., Ki-68
immunohistochemical staining) and expression levels of
apoptosis-associated proteins other than PARP (e.g., caspase-3).
Fractionation followed by chromatographic analysis of the extract
of MB is also needed to obtain information on structure-activity
relationships (41–43).
There is limited information on the toxicity of the
acetone or methanol extracts in experimental animal models, and
there are, apparently, no clinical toxicity data on the use of the
extracts of MB in humans. In the present study, in a mouse
xenograft bioassay system no significant adverse side effects were
caused by the administration of the acetone extract of MB into the
tumor (Fig. S2), although a
decrease in tumor volume was not statistically significant. The
possible applications of the present study may depend on whether
the acetone extract of MB can be given safely to humans at amounts
appropriate enough to achieve pharmacologically active conditions.
The important consideration is whether sufficient blood and tissue
levels of the active component can be achieved and whether the
extracts exert significant adverse side effects. To answer these
questions, further investigations are in progress to identify the
active component of the crude dried leaf extracts and its
underlying mechanism of action.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Sachi Sri Kantha
(Center for General Education, Gifu University, Gifu Pharmaceutical
University, Gifu, Japan) for valuable discussions, comments and
editing the manuscript. The authors also acknowledge the assistance
of the Research Equipment Sharing Center at the Nagoya City
University.
Funding
This study was supported in part by grants from the Research
Foundation for Oriental Medicine, Nagoya, Japan (grant nos. H27-2
and H30-6) and the Ministry of Education, Culture, Sports, Science
and Technology of Japan (grant nos. 22501050, 25430154 and
17K07223).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HM, MI and MS designed the study. HM, SA, EY, TN,
NS, MI, KO and MS performed the experiments and acquired the data.
HM and MS confirm the authenticity of all the raw data. HM, KF, TN,
MI, KO, KK and MS analyzed and interpreted the data. HM, KF, MI,
KO, KK and MS wrote, reviewed and/or revised the manuscript. MI and
MS contributed to the material management. MS supervised the study.
All of the authors are fully aware of the contents of this paper.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were performed with the
approval of the Animal Ethics Committee of the Nagoya City
University (approval no. H28M-03) and according to the guidelines
of the committee. Samples of Musa basjoo used in the current
study were identified by MI, and located at the Herbal Garden, Gifu
Pharmaceutical University, Japan. Voucher specimens can be made
available for researchers, when requested via authentic
certification.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Surh YJ: Cancer chemoprevention with
dietary phytochemicals. Nat Rev Cancer. 3:768–780. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee KW, Bode AM and Dong Z: Molecular
targets of phytochemicals for cancer prevention. Nat Rev Cancer.
11:211–218. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kennedy J: Bananas and people in the
homeland of genus Musa: Not just pretty fruit. Ethnobot Res
Appl. 7:179–197. 2009. View Article : Google Scholar
|
|
4
|
Zhang XW, Park YJ, Choe YH and Kim BS:
Effect of anti-inflamentation extracts from Korean traditional
medicinal herb. Int J Pharm Res. 4:122–125. 2014.
|
|
5
|
Imam MZ and Akter S: Musa
paradisiaca L. and Musa sapientum L.: A phytochemical
and pharmacological review. J Appl Pharm Sci. 1:14–20. 2011.
|
|
6
|
Nishino H, Tokuda H, Satomi Y, Masuda M,
Onozuka M, Yamaguchi S, Takayasu J, Tsuruta J, Takemura M, Ii T, et
al: Cancer chemoprevention by phytochemicals and their related
compounds. Asian Pac J Cancer Prev. 1:49–55. 2000.PubMed/NCBI
|
|
7
|
Jiao L, Bi L, Lu Y, Wang Q, Gong Y, Shi J
and Xu L: Cancer chemoprevention and therapy using Chinese herbal
medicine. Biol Proced Online. 20:12018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Morioka T, Suzui M, Nabandith V, Inamine
M, Aniya Y, Nakayama T, Ichiba T, Mori H and Yoshimi N: The
modifying effect of Peucedanum japonicum, a herb in the
Ryukyu Islands, on azoxymethane-induced colon preneoplastic lesions
in male F344 rats. Cancer Lett. 205:133–141. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Morioka T, Suzui M, Nabandith V, Inamine
M, Aniya Y, Nakayama T, Ichiba T and Yoshimi N: Modifying effects
of Terminalia catappa on azoxymethane-induced colon
carcinogenesis in male F344 rats. Eur J Cancer Prev. 14:101–105.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kaneshiro T, Suzui M, Takamatsu R,
Murakami A, Ohigashi H, Fujino T and Yoshimi N: Growth inhibitory
activities of crude extracts obtained from herbal plants in the
Ryukyu Islands on several human colon carcinoma cell lines. Asian
Pac J Cancer Prev. 6:353–358. 2005.PubMed/NCBI
|
|
11
|
Ishihara M, Naoi K, Hashita M, Itoh Y and
Suzui M: Growth inhibitory activity of ethanol extracts of Chinese
and Brazilian propolis in four human colon carcinoma cell lines.
Oncol Rep. 22:349–354. 2009.PubMed/NCBI
|
|
12
|
Suzui M, Masuda M, Lim JTE, Albanese C,
Pestell RG and Weinstein IB: Growth inhibition of human hepatoma
cells by acyclic retinoid is associated with induction of p21CIP1
and inhibition of expression of cyclin D1. Cancer Res.
62:3997–4006. 2002.PubMed/NCBI
|
|
13
|
Ando S, Fukamachi K, Yoshimoto E,
Matsumoto H, Iinuma M and Suzui M: Palmitoyl piperidinopiperidine,
a novel derivative of 10-hydroxy-2-decenoic acid, as a potent and
selective anticancer agent against human colon carcinoma cell
lines. Int J Oncol. 58:251–265. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Suzui M, Inamine M, Kaneshiro T, Morioka
T, Yoshimi N, Suzuki R, Kohno H and Tanaka T: Indole-3-carbinol
inhibits the growth of human colon carcinoma cells but enhances the
tumor multiplicity and volume of azoxymethane-induced rat colon
carcinogenesis. Int J Oncol. 27:1391–1399. 2005.PubMed/NCBI
|
|
15
|
Suzui M, Sunagawa N, Chiba I, Moriwaki H
and Yoshimi N: Acyclic retinoid, a novel synthetic retinoid,
induces growth inhibition, apoptosis, and changes in mRNA
expression of cell cycle- and differentiation-related molecules in
human colon carcinoma cells. Int J Oncol. 28:1193–1199.
2006.PubMed/NCBI
|
|
16
|
Suzui M, Shimizu M, Masuda M, Lim JTE,
Yoshimi N and Weinstein IB: Acyclic retinoid activates retinoic
acid receptor β and induces transcriptional activation of p21CIP1
in HepG2 human hepatoma cells. Mol Cancer Ther. 3:309–316.
2004.PubMed/NCBI
|
|
17
|
Suzui M, Okuno M, Tanaka T, Nakagama H and
Moriwaki H: Enhanced colon carcinogenesis induced by azoxymethane
in min mice occurs via a mechanism independent of beta-catenin
mutation. Cancer Lett. 183:31–41. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Weinstein IB: Disorders in cell circuitry
during multistage carcinogenesis: The role of homeostasis.
Carcinogenesis. 21:857–864. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Freed-Pastor WA and Prives C: Mutant p53:
One name, many proteins. Genes Dev. 26:1268–1286. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Amelio I and Melino G: The p53 family and
the hypoxia-inducible factors (HIFs): Determinants of cancer
progression. Trends Biochem Sci. 40:425–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tomasini R, Tsuchihara K, Wilhelm M,
Fujitani M, Rufini A, Cheung CC, Khan F, Itie-Youten A, Wakeham A,
Tsao MS, et al: TAp73 knockout shows genomic instability with
infertility and tumor suppressor functions. Genes Dev.
22:2677–2691. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guo X, Keyes WM, Papazoglu C, Zuber J, Li
W, Lowe SW, Vogel H and Mills AA: TAp63 induces senescence and
suppresses tumorigenesis in vivo. Nat Cell Biol. 11:1451–1457.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang A, Kaghad M, Wang Y, Gillett E,
Fleming MD, Dötsch V, Andrews NC, Caput D and McKeon F: p63, a
p53 homolog at 3q27-29, encodes multiple products with
transactivating, death-inducing, and dominant-negative activities.
Mol Cell. 2:305–316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang B, Ban X, He J, Tong J, Tian J and
Wang Y: Comparative analysis of essential oil components and
antioxidant activity of extracts of Nelumbo nucifera from
various areas of China. J Agric Food Chem. 58:441–448. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Q, Yang Y, Zhao X, Zhu B, Nan P, Zhao
J, Wang L, Chen F, Liu Z and Zhong Y: Chemical variation in the
essential oil of Ephedra sinica from Northeastern China.
Food Chem. 98:52–58. 2006. View Article : Google Scholar
|
|
26
|
Sefidkon F, Jamzad Z and Mirza M: Chemical
variation in the essential oil of Satureja sahendica from
Iran. Food Chem. 88:325–328. 2004. View Article : Google Scholar
|
|
27
|
Wang YH and Zhang YR: Variations in
compositions and antioxidant activities of essential oils from
leaves of Luodian Blumea balsamifera from different harvest
times in China. PLoS One. 15:e02346612020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tan XJ, Li Q, Chen XH, Wang ZW, Shi ZY, Bi
KS and Jia Y: Simultaneous determination of 13 bioactive compounds
in Herba Artemisiae Scopariae (Yin Chen) from different harvest
seasons by HPLC-DAD. J Pharm Biomed Anal. 47:847–853. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang L, Zhang B, Wang Y, Sun J, Ma X,
Wang G, Fu S, Lin C and Li Y: Three new acenaphthene derivatives
from rhizomes of Musa basjoo and their cytotoxic activity.
Nat Prod Res. 35:1307–1312. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Uchikura T, Sugiwaki H, Yoshimura M,
Mitsuhashi H, Fuchino H, Kawahara N, Hakamatsuka T and Amakura Y:
Characterization of UV-sensitive marker constituents of Polygala
root for TLC: Applications in quality control of single crude drug
extract preparations. Chem Pharm Bull (Tokyo). 66:1174–1180. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Patil S, Nivsarkar M and Anandajiwala S:
Isolation and TLC densitometric quantification of Lysergol from the
seeds of Ipomoea muricata (Linn.) Jacq. ISRN Chromatogr.
2013:1345862013. View Article : Google Scholar
|
|
32
|
Kaya B, Menemen Y and Saltan FZ: Flavonoid
compounds identified in Alchemilla L. species collected in
the north-eastern Black Sea region of Turkey. Afr J Tradit
Complement Altern Med. 9:418–425. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hemmalakshmi S, Priyanga S and Devaki K:
Phytochemical screening and HPTLC fingerprinting analysis of
ethanolic extract of Erythrina variegate L. flowers. Int J
Pharm Pharm Sci. 8:210–217. 2016.
|
|
34
|
Bhat A and Raveesha KA: Antifungal
activity of Pimenta dioica (L.) merril an aromatic medicinal
tree. Int J Pharm Pharm Sci. 8:92–95. 2016. View Article : Google Scholar
|
|
35
|
Wang X, Wang D, Huo Y, Dai D, Li C and Liu
G: Identification of isoliquiritigenin as an activator that
stimulates the enzymatic production of glycyrrhetinic acid
monoglucuronide. Sci Rep. 7:125032017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lv GP, Aoli M, Zhou B and Zhao J:
Development of a rapid and simple non-derivatization method to
determine constituents and antioxidative capacity of camellia oils
by HPTLC. Food Nutr Sci. 4:204–210. 2013.
|
|
37
|
Sobstyl E, Szopa A, Ekiert H, Gnat S,
Typek R and Choma IM: Effect directed analysis and TLC screening of
Schisandra chinensis fruits. J Chromatogr A.
1618:4609422020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Usia T, Banskota AH, Tezuka Y, Midorikawa
K, Matsushige K and Kadota S: Constituents of Chinese propolis and
their antiproliferative activities. J Nat Prod. 65:673–676. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sun LP, Xu X, Hwang HH, Wang X, Su KY and
Chen YL: Dichloromethane extracts of propolis protect cell from
oxygen-glucose deprivation-induced oxidative stress via reducing
apoptosis. Food Nutr Res. 60:300812016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xuan H, Wang Y, Li A, Fu C, Wang Y and
Peng W: Bioactive components of Chinese propolis water extract on
antitumor activity and quality control. Evid Based Complement
Alternat Med. 2016:96419652016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ito T, Furusawa M, Tanaka T, Ali Z, Iliya
I, Nakaya K, Murata J, Darnaedi D and Iinuma M: Resveratrol
derivatives from Upuna borneensis. Chem Pharm Bull (Tokyo).
53:219–224. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hattori H, Okuda K, Murase T, Shigetsura
Y, Narise K, Semenza GL and Nagasawa H: Isolation, identification,
and biological evaluation of HIF-1-modulating compounds from
Brazilian green propolis. Bioorg Med Chem. 19:5392–5401. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Robles AJ, McCowen S, Cai S, Glassman M,
Ruiz F II, Cichewicz RH, McHardy SF and Mooberry SL:
Structure-activity relationships of new natural product-based
diaryloxazoles with selective activity against androgen
receptor-positive breast cancer cells. J Med Chem. 60:9275–9289.
2017. View Article : Google Scholar : PubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBIPubMed/NCBI
|