Introduction
In 2020, 905,677 patients were diagnosed with liver
cancer and this malignancy was accountable for 830,180
cancer-associated mortalities worldwide (1). Hepatocellular carcinoma (HCC), which
is a multigene disease with heterogeneous pathological mechanisms
and clinical manifestation, accounts for 75–85% of primary liver
cancer cases (1) and is a major
health problem worldwide. It has been observed that the use of
ultrasound monitoring every 6 months (with or without α-fetoprotein
for its treatment) is associated with improved early detection as
well as improved overall survival (OS); however, in clinical
practice, implementation-related limitations frequently result in a
high proportion of HCC cases only detected at late stages (2). Furthermore, despite the improvements
associated with the use of antiangiogenic drugs and immunotherapy
over the past decade, HCC prognosis is limited (3). The major unmet challenges related to
HCC treatments include advancements in the treatment at earlier
stages of the disease, applying the treatment to patients with
liver dysfunction, the discovery and validation of predictive
biomarkers and the development of more effective combinatorial or
sequential treatment approaches (3,4).
Therefore, the mechanisms of HCC require to be explored and the
identification of valuable biomarkers is urgently required.
Genome-wide expression profiling has enabled the
analysis of patient heterogeneity within a short period. It has
been proposed that the expression of numerous genes, including
forkhead box protein M1 (FOXM1) (5,6) and
polo-like kinase 1 (PLK1) (7,8), may
serve as a putative prognostic biomarker for HCC. Although several
studies have indicated that FOXM1 and PLK1 overexpression are
associated with poor cancer prognosis (9–11),
the underlying mechanisms have remained to be fully elucidated.
Furthermore, to the best of our knowledge, the effect of the
association between FOXM1 and PLK1 on the development and prognosis
of HCC has not been reported in any previous study.
FOXM1 belongs to a large family of Fox transcription
factors, all of which have a conserved domain attached to DNA
(winged helix) (12). Furthermore,
it has an important role in regulating cell cycle progression via
the stimulation of the genes that are critical for G1-S and G2-M
transition, including S-phase kinase-associated protein 2, PLK1,
centromeric protein A and survivin (13,14),
and is itself regulated during the cell cycle process.
Transcriptional activation of FOXM1 depends on cyclin-dependent
kinase and PLK1 kinase mediates its phosphorylation (15–17).
Furthermore, FOXM1 is frequently expressed at a higher level than
normal in a variety of human cancers (18–20).
Furthermore, several studies have demonstrated that FOXM1 is a key
transcription factor that is associated with HCC (21,22).
It has also been reported that elevated FOXM1 expression is
associated with a poor prognosis of the disease (23,24).
Cell cycle disorders are essential for tumor
development and protein kinases, which have important roles in
regulating the cell cycle, are valuable targets for cancer therapy.
Specifically, PLK1, a member of the serine/threonine kinase family,
promotes cell mitosis in mammalian cells (25,26).
It has also been identified as an essential mitotic kinase that
controls mitotic entry, spindle assembly, centrosome maturation and
cytokinesis (27,28). PLK1 overexpression was reported to
cause cell cycle overrides in tumor cells, resulting in the
survival, enhanced proliferation and immune evasion of cancer cells
(29–31). It has also been observed that its
expression is increased in numerous cancer types, including lung,
bladder, breast and liver cancers (32–34).
In addition, several studies have indicated that PLK1
overexpression may serve as an important prognostic factor for HCC
(35,36); however, the underlying associated
mechanisms have remained elusive. Selective inhibitors of PLK1
potently cause mitotic arrest and induce tumor cell apoptosis
(37–39), indicating that PLK1 is a potential
target for antitumor treatment.
PLK1, a target of FOXM1, is required for FOXM1
transcription activation and the formation of a feedback loop
(14,16). In addition, PLK1 and FOXM1 are
essential for the cell cycle process and are related to HCC
prognosis. However, the association between this feedback loop and
HCC has remained to be investigated. Therefore, in the present
study, the prognostic values of PLK1 and FOXM1 overexpression in
HCC were estimated using data from The Cancer Genome Atlas (TCGA)
and International Cancer Genome Consortium Japan (ICGC JP) HCC
cohorts. Furthermore, molecular analyses and cell proliferation
assays were performed to examine the role of PLK1 and FOXM1 in Huh7
cells and the results suggested that the feedback loop is required
for the proliferation of Huh7 cells.
Materials and methods
Clinical cohorts
Sequencing and clinical data of patients with HCC
were obtained from two public cohorts, namely TCGA (http://xena.ucsc.edu) and ICGC (https://dcc.icgc.org). A total of 373 and 243 patients
from the TCGA and ICGC JP cohorts, respectively, were included in
the analysis. Patients with incomplete OS or disease-free survival
(DFS) information were excluded. OS was defined as the time from
the date of initial pathological diagnosis to the time of death or
last follow-up, while DFS was defined as the time from first
treatment to the time of tumor recurrence or death. The TCGA cohort
was used as the exploration cohort (clinicopathological information
is provided in Table I), while the
ICGC JP cohort was used for validation.
 | Table I.Baseline of characteristics of the
patients (n=373). |
Table I.
Baseline of characteristics of the
patients (n=373).
| Characteristic | N (%) |
|---|
| Age, years |
|
|
<50 | 70
(18.8) |
|
50-59 | 99
(26.5) |
|
60-69 | 121 (32.4) |
|
≥70 | 83
(22.3) |
| Sex |
|
|
Male | 252 (67.6) |
|
Female | 121 (32.4) |
| Stage |
|
| I | 173 (49.6) |
| II | 86
(24.6) |
|
III–IV | 90
(25.8) |
| Virus status |
|
|
None | 199 (56.2) |
|
HBV | 99
(28.0) |
|
HCV | 48
(13.8) |
|
HBV+HCV | 7
(2.0) |
Cell lines and cell culture
The human Huh7 cell line (American Type Culture
Collection) was cultured in a growth medium consisting of
Dulbecco's Modified Eagle's Medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 4.5 g/l glucose, 10% fetal
bovine serum (FBS; Shanghai ExCell Biology, Inc.) and 1%
penicillin/streptomycin at 37°C in a humidified atmosphere with 5%
CO2.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. Complementary DNA (cDNA) was synthesized
using the ReverTra Ace qPCR RT Master Mix with a gDNA Remover kit
(Toyobo Life Science) according to the manufacturer's instructions.
qPCR was then performed in a 25-µl volume reaction mixture
containing 12.5 µl AceQ Universal SYBR qPCR Master Mix (Vazyme
Biotech Co., Ltd.), 2 µl template cDNA (100 ng/µl) and 1 µM of
primers in a LightCycler 96 (Roche Diagnostics Co., Ltd.). The
thermocycling conditions were as follows: 95°C for 10 min, followed
by 40 cycles of 94°C for 15 sec, 60°C for 30 sec and 72°C for 30
sec. GAPDH was used as a normalization control. Each experiment was
performed independently at least three times and the fold change in
the expression of each gene was calculated using the
2−ΔΔCq method (40).
Primers for qPCR were obtained from BioSune Biotechnology Co., Ltd.
All primers were designed to cross an intron-exon junction sequence
to minimize genomic DNA contamination. The primer sequences were as
follows: qPCR-PLK1-forward (F), 5′-AAGAGATCCCGGAGGTCCTA-3′ and
qPCR-PLK1-reverse (R), 5′-GCTGCGGTGAATGGATATTT-3′; qPCR-FOXM1-F,
5′-CGTGGATTGAGGACCACTTT-3′ and qPCR-FOXM1-R,
5′-TCTGCTGTGATTCCAAGTGC-3′; qPCR-GAPDH-F,
5′-ACAACTTTGGTATCGTGGAAGG-3′ and qPCR-GAPDH-R,
5′-GCCATCACGCCACAGTTTC-3′.
Immunoblotting
Cells were lysed in radioimmunoprecipitation assay
cell lysis buffer (Beyotime Institute of Biotechnology) containing
protease inhibitors (cat. no. HY-K0010; MedChem Express) and
phosphatase inhibitor cocktail (cat. no. 78427; Thermo Fisher
Scientific, Inc.). Protein concentrations were determined via a
bicinchoninic acid protein assay (Pierce; Thermo Fisher Scientific,
Inc.). Proteins were separated using 10% SDS-PAGE [gels using 1X
running buffer and transferred to polyvinylidene difluoride
membranes (MilliporeSigma)]. Afterwards, the membranes were blocked
by 5% skimmed milk (Anchor; Fonterra Co-operative Group) at room
temperature for 1 h and incubated with primary antibodies against
FOXM1 (rabbit; cat. no. A2493; dilution, 1:1,000; Abclonal), PLK1
(rabbit; cat. no. 208G4; dilution, 1:1,000; Cell Signaling
Technology, Inc.) and GAPDH (mouse; cat. no. AC002; dilution,
1:10,000; Abclonal) at 4°C overnight. Subsequently, the membranes
were cleaned twice with TBS with 0.1% Tween-20 and incubated with
secondary goat anti-rabbit IgG (H+L) Cross-Adsorbed Secondary
Antibody, HRP (dilution, 1:5,000; cat. no. G21234; Thermo Fisher
Scientific, Inc.) or goat anti-mouse IgG (H+L) Cross-Adsorbed
Secondary Antibody, HRP (dilution, 1:5,000; cat. no. G21040; Thermo
Fisher Scientific, Inc.) at room temperature for 1 h. Proteins were
detected via enhanced chemiluminescence (Vazyme Biotech Co., Ltd.)
with a digital luminescent image analyzer (Tanon-4200; Tanon
Science and Technology Co., Ltd.). The intensities of protein bands
were semi-quantified using ImageJ software (ImageJ bundled with
64-bit Java 1.8.0_172; National Institutes of Health).
RNA interference
Human FOXM1 small interfering RNA (siRNA), human
PLK1 siRNA and control siRNA were obtained from Shanghai
GenePharma, Co., Ltd. The siRNA oligonucleotides were transfected
into Huh7 cells (at 70% confluence) using Lipofectamine RNAiMAX
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. siRNAs were mixed with Opti-MEM reduced
serum medium (Gibco; Thermo Fisher Scientific, Inc.) and
Lipofectamine RNAiMAX and incubated for 20 min at room temperature.
Subsequently, siRNA/Lipofectamine in Opti-MEM was diluted 1:5
(corresponding to a final concentration of 50 nM) in
differentiation medium, and added to the cells. Thereafter, cells
were incubated for 48 h at 37°C with 5% CO2. The
following siRNAs were used for the experiments, which had the
following sequences: siFOXM1-1, 5′-GCUGGGAUCAAGAUUAUUATT-3′;
siFOXM1-2, 5′-GGCUGCACUAUCAACAAUATT-3′; siPLK1-1,
5′-CCCUCACAGUCCUCAAUAATT-3′; siPLK1-2, 5′-GGCAACCAAAGUCGAAUAUTT-3′;
si negative control (NC), 5′-ACGUGACACGUUCGGAGAATT-3′.
Cell counting kit 8 (CCK-8) assay
A CCK-8 kit (Vazyme Biotech Co., Ltd.) was used to
measure the proliferation of Huh7 cells. A total of 1,000 cells in
a volume of 100 µl per well were cultured in six replicate wells in
a 96-well plate in medium containing 10% FBS and 1%
penicillin-streptomycin at 37°C with 5% CO2 for 6 h.
When cells had adhered, CCK-8 reagent (10 µl) was added to 90 µl
DMEM to generate the working solution, of which 100 µl was added
per well and incubated for 2 h. This assay was performed at 0, 24,
48, 72 and 96 h. The optical densities were measured at a spectral
wavelength of 450 nm using a microplate reader (Thermo Fisher
Scientific, Inc.). Six replicates were analyzed for each time
point.
5-Ethynyl-2′-deoxyuridine (EdU)-DNA
synthesis assay
To measure the DNA replication activity of the Huh7
cells, a Cell-Light EdU Apollo488 in Vitro Kit (cat. no.
C10310-3; Guangzhou RiboBio Co., Ltd.) was used. Huh7 cells were
seeded in 96-well plates at a density of 8×103 cells per
well. After 24 h, the cell culture medium was replaced with 50 µM
EdU solution diluted with growth culture medium, followed by
incubation for 2 h. The cells were then processed using the
Cell-Light EdU Apollo488 in Vitro Kit according to the
manufacturer's protocol. Images were captured with an Olympus
fluorescence microscope (BX53; Olympus Corporation).
Cell-cycle analysis
A total of 1×106 cells were washed twice
with PBS and fixed overnight with 1 ml pre-cooled 75% ethanol at
4°C. Thereafter, cells were collected by centrifugation (500 × g; 5
min; 4°C), washed twice in PBS and incubated with propidium iodide
(5 ug/ml, Sigma) and RNase A (0.1 mg/ml; Thermo Fisher Scientific,
Inc.) for 30 min at 4°C in the dark. A 40-µm screen filter was then
used to filter the cell suspension and remove any adhesive cells.
This was followed by flow cytometry to analyze the DNA content
using the BD LSRFortessa system (BD Biosciences). FlowJo_v10
software (Tree Star, Inc.) was used to estimate the proportion of
cells in the G0/G1, S and G2/M phases.
Pharmacological inhibitor of PLK1
The pharmacological PLK1 inhibitor BI 2536
(HY-50698) was purchased from MedChemExpress. The drugs were
reconstituted in DMSO and aliquots were stored at −20°C. An
equivalent amount of DMSO was used for each experiment as a vehicle
control. For cell proliferation and cell cycle assays, Huh7 cells
were incubated with 10 nM BI 2536 for 24 h.
Statistical analysis
Statistical analysis was performed using R software
(version 4.0.3) for survival analysis and Cox analysis, and
GraphPad Prism (version 8.0; GraphPad Software, Inc.) for others.
Kaplan-Meier curves were used to estimate OS and DFS. The log-rank
test was used to compare patient survival times between high and
low gene expression groups and hazard ratios (HRs) were calculated
using the Cox proportional hazards model. The combined expression
of FOXM1 and PLK1 was dependent on FOXM1 expression and PLK1
expression, thus they were analyzed respectively in Multivariable
Cox analysis. Due to the crossing of survival curves, the ‘TSHRC’
package (v0.1-6; http://CRAN.R-project.org/package=TSHRC) of R
software, which is a two-stage procedure for comparing HR
functions, particularly suited for situations where HR functions
cross, was used to perform a two-stage test rather than the
log-rank test (41,42). Pearson's correlation coefficient
was calculated to analyze the correlation between FOXM1 and PLK1
expression. Differences between 2 groups were analyzed using the
unpaired Student's t-test with or without Welch's correction.
One-way ANOVA followed by Tukey's post-hoc test was used for
comparisons between multiple groups. Values are expressed as the
mean ± standard deviation. P<0.05 was considered to indicate a
statistically significant difference.
Results
Baseline characteristics
The TCGA cohort was used as the exploration cohort
and the ICGC JP cohort was used for validation. The baseline
characteristics of the TCGA data set analyzed in the present study
are summarized in Table I. The
mean age of TCGA cohort was 59.5 years and the percentage of men
was 67.6%. The most prevalent hepatitis virus was HBV. The ICGC JP
cohort contained sequencing and clinical data of 243 patients with
HCC from Japan. The mean age of the ICGC cohort was 67.5 years and
the percentage of men was 74.9%. Most patients presented with
primary tumors (98.8%) and approximately half of the patients had
stage II tumors (45.3%).
Overexpression of FOXM1 and PLK1 is
associated with poor prognosis for HCC
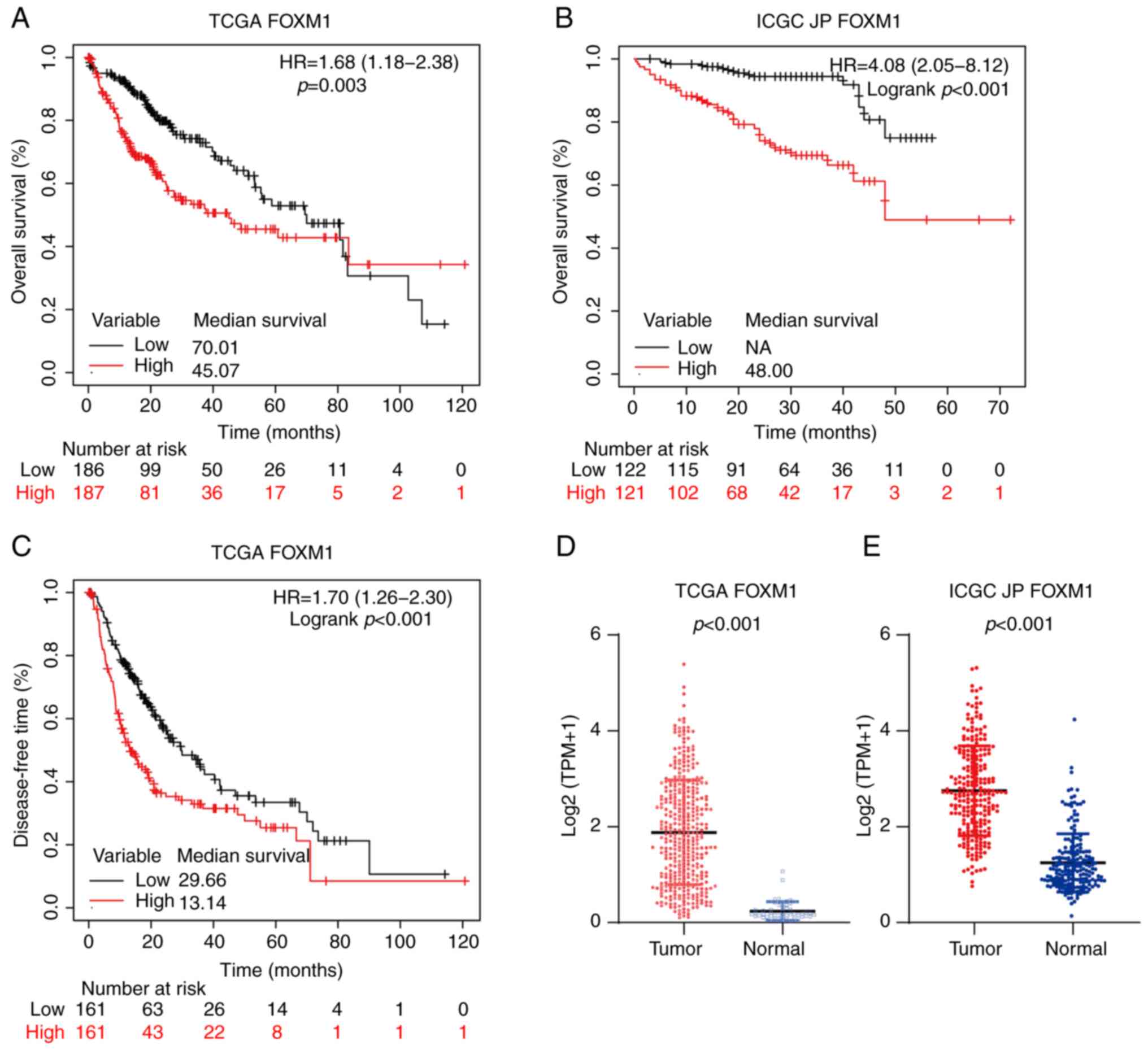
The expression of FOXM1 and PLK1 in HCC and their
effect on survival were first analyzed (Fig. 1). Compared with non-tumor liver
tissues, the tumor tissues exhibited significantly higher FOXM1
expression in the TCGA (P<0.001; Fig. 1D) and ICGC JP (P<0.001; Fig. 1E) cohorts. To investigate the
prognostic value of FOXM1 and PLK1 expression in HCC, the patients
with HCC were divided into low and high expression groups at the
50th percentile. In addition, in the TCGA cohort, patients with
high FOXM1 expression levels (FH) had shorter OS [HR,
1.68; 95% confidence interval (CI), 1.18-2.38; P=0.003; Fig. 1A] and DFS (HR, 1.70; 95% CI,
1.26-2.30; P<0.001; Fig. 1C).
Consistently, FH was associated with poor prognosis in
the ICGC JP cohort (HR, 4.08; 95% CI, 2.05-8.12; P<0.001;
Fig. 1B).
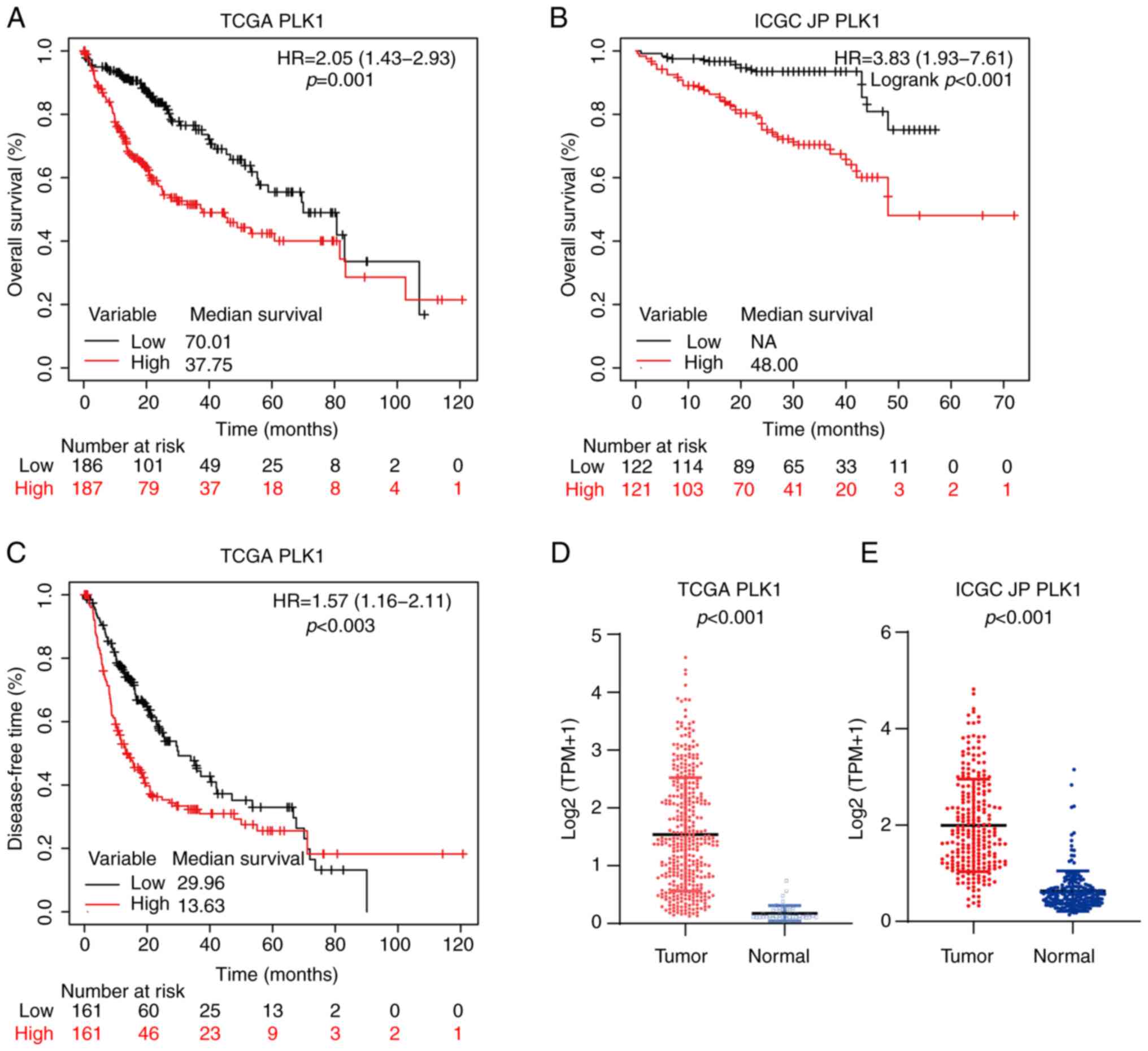
As presented in Fig.
2, similar to FOXM1, PLK1 was highly expressed in HCC tumor
tissues in both the TCGA (P<0.001; Fig. 2D) and ICGC JP (P<0.001; Fig. 2E) cohorts. In addition, in the TCGA
cohort, patients with high PLK1 expression level (PH)
had shorter OS (HR, 2.05; 95% CI, 1.43-2.93; P<0.001; Fig. 2A) and DFS (HR, 1.57; 95% CI,
1.16-2.11; P=0.003; Fig. 2C). In
the ICGC JP cohort, patients with PH also had
significantly poorer OS (HR, 3.83; 95% CI, 1.93-7.61; P<0.001;
Fig. 2B).
To assess the independent predictive value of
FH and PH, logistic regression with a
multivariate Cox proportional hazards model was utilized. After
adjusting for age, sex, stage and virus status, stage and virus
status were identified as independent prognostic factors for OS.
However, neither FH (HR, 1.14; 95% CI, 0.67-1.95;
P=0.634; Table II), nor
PH (HR, 1.73; 95% CI, 1.00-2.99; P=0.052; Table II) were significantly and
independently associated with a shorter OS.
| Table II.Cox regression proportional hazards
model for the analysis of the prognostic values of FOXM1 expression
and PLK1 expression for overall survival in patients with
hepatocellular carcinoma. |
Table II.
Cox regression proportional hazards
model for the analysis of the prognostic values of FOXM1 expression
and PLK1 expression for overall survival in patients with
hepatocellular carcinoma.
|
| Univariate | Multivariate |
|---|
|
|
|
|
|---|
| Characteristic | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Age
(continuous) | 1.01
(1.00-1.03) | 0.089 | 1.01
(0.99-1.03) | 0.292 |
| Sex (female vs.
male) | 1.26
(0.88-1.80) | 0.200 | 1.08
(0.72-1.63) | 0.712 |
| Stage |
|
|
|
|
| II vs.
I | 1.42
(0.87-2.31) | 0.164 | 1.11
(0.66-1.88) | 0.691 |
| III–IV
vs. I | 2.82
(1.86-4.28) | <0.001 | 2.00
(1.26-3.18) | 0.003 |
| Virus status (vs.
none) |
|
|
|
|
|
HBV | 0.34
(0.20-0.56) | <0.001 | 0.43
(0.24-0.78) | 0.005 |
|
HCV | 0.95
(0.56-1.61) | 0.847 | 1.15
(0.64-2.05) | 0.646 |
|
HBV+HCV | 0.42
(0.10-1.74) | 0.233 | 0.37
(0.08-1.61) | 0.184 |
| FOXM1 (high vs.
low) | 1.68
(1.18-2.38) | 0.004 | 1.14
(0.67-1.95) | 0.634 |
| PLK1 (high vs.
low) | 2.05
(1.43-2.93) | <0.001 | 1.73
(1.00-2.99) | 0.052 |
Combined overexpression of FOXM1 and
PLK1 is associated with poor HCC prognosis
FOXM1 and PLK1 are essential for cell cycle
progression. PLK1 itself is a target of FOXM1 and phosphorylation
of FOXM1 mediated by PLK1 is required for FOXM1 transcription
activation (14,16). In both the TCGA
(r2=0.793, P<0.001; Fig.
3A) and ICGC JP (r2=0.714, P<0.001; Fig. 3B) cohorts, a positive linear
correlation between FOXM1 and PLK1 expression was observed,
consistent with the presence of a feedback loop between them in HCC
tissues. In the TCGA cohort, patients with combined high expression
of FOXM1 and PLK1 (FH-PH) exhibited
significantly shorter OS (HR, 2.04; 95% CI, 1.40-2.97; P<0.001;
Fig. 3C) and DFS (HR, 1.73; 95%
CI, 1.26-2.38, P<0.001; Fig.
3E). The median OS corresponding to the
FH-PH (33.02; Fig. 3C) group was shorter than that
corresponding to the FH (45.07; Fig. 1A) and PH (37.75;
Fig. 2A) groups. These
observations were validated in the ICGC JP cohort (Fig. 3B). Furthermore, the prognostic
value of FH-PH expression in patients with
HCC was confirmed using logistic regression with a multivariate Cox
proportional hazards model. The results indicated that
FH-PH expression was the most significant
independent predictor of OS (HR, 1.94; 95% CI, 1.31-2.89; P=0.001;
Table III).
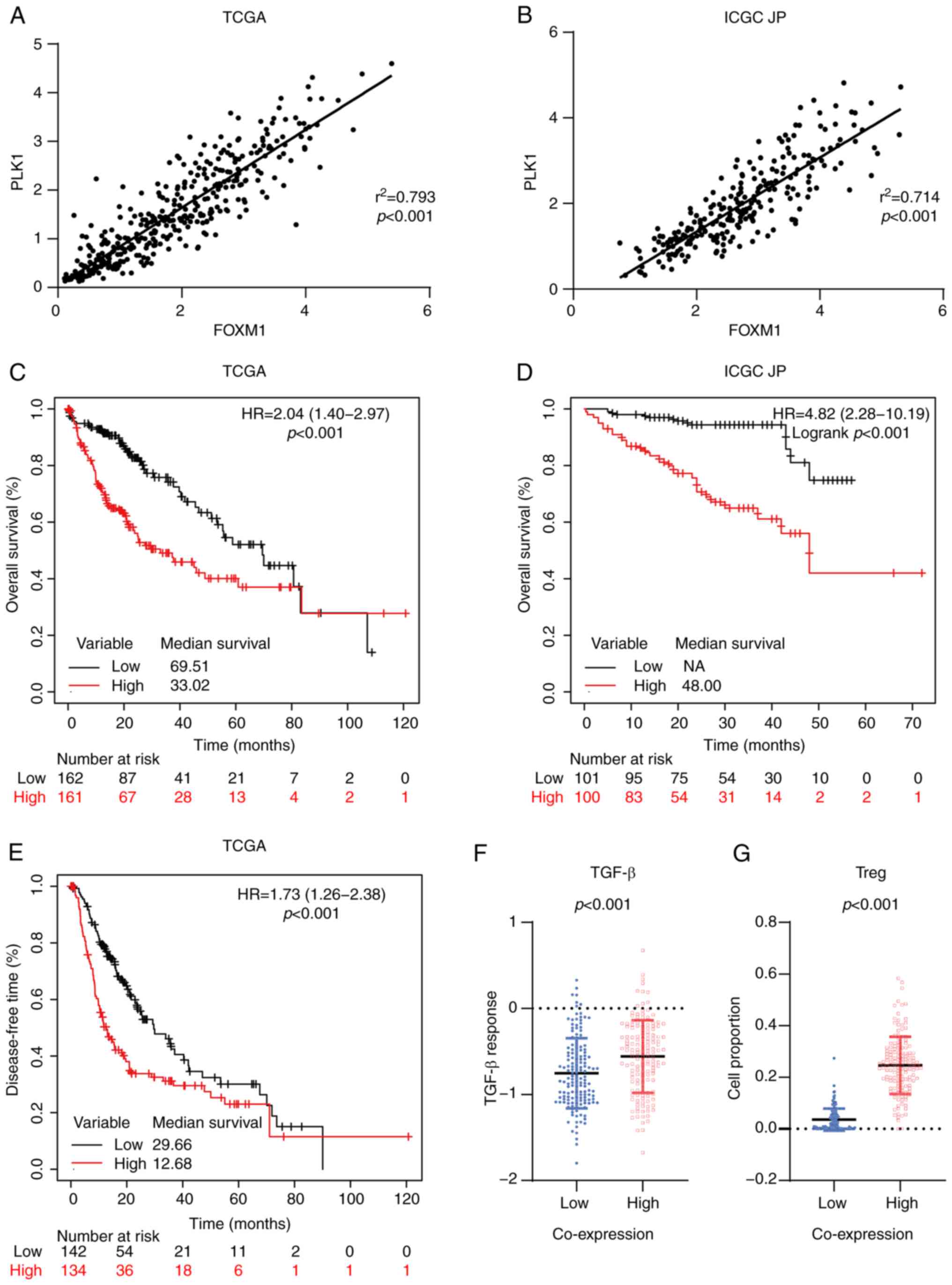 | Figure 3.Coordinated overexpression of FOXM1
and PLK1 is associated with poor prognosis in HCC. Correlation
between FOXM1 and PLK1 expression in HCC in the (A) TCGA and (B)
ICGC JP cohorts based on Pearson's correlation analysis.
Kaplan-Meier survival curve estimates of overall survival in the
(C) TCGA (n=323) and (D) ICGC JP (n=201) cohorts. The patients were
divided into two subgroups: High FOXM1+PLK1 and low FOXM1+PLK1. (E)
Kaplan-Meier survival curve estimates of disease-free survival in
the TCGA cohort (n=276). (F) Transforming growth factor-β response
score of HCC in the TCGA cohort, stratified by the co-expression
status of FOXM1 and PLK1. (G) Tumor-infiltrating Tregs in HCC in
the TCGA cohort, stratified based on the co-expression status of
FOXM1 and PLK1. FOXM1, forkhead box protein M1; PLK1, polo-like
kinase 1; HR, hazard ratio; TCGA, The Cancer Genome Atlas; ICGC JP,
International Cancer Genome Consortium Japan; HCC, hepatocellular
carcinoma; NA, not available; Tregs, regulatory T cells. |
| Table III.Cox regression proportional hazards
model for the analysis of the prognostic value of combined
expression of FOXM1 and PLK1 for overall survival in patients with
hepatocellular carcinoma. |
Table III.
Cox regression proportional hazards
model for the analysis of the prognostic value of combined
expression of FOXM1 and PLK1 for overall survival in patients with
hepatocellular carcinoma.
|
| Univariate | Multivariate |
|---|
|
|
|
|
|---|
| Characteristic | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Age
(continuous) | 1.01
(1.00-1.03) | 0.089 | 1.01
(1.01-1.03) | 0.150 |
| Sex (female vs.
male) | 1.26
(0.88-1.80) | 0.200 | 1.22
(0.79-1.89) | 0.366 |
| Stage |
|
|
|
|
| II vs.
I | 1.42
(0.87-2.31) | 0.164 | 1.03
(0.59-1.81) | 0.905 |
| III–IV
vs. I | 2.82
(1.86-4.28) | <0.001 | 1.97
(1.20-3.26) | 0.008 |
| Virus status (vs.
none) |
|
|
|
|
|
HBV | 0.34
(0.20-0.56) | <0.001 | 0.49
(0.25-0.96) | 0.038 |
|
HCV | 0.95
(0.56-1.61) | 0.847 | 1.07
(0.58-1.98) | 0.821 |
|
HBV+HCV | 0.42
(0.10-1.74) | 0.233 | 0.44
(0.10-2.01) | 0.289 |
| Combined FOXM1+PLK1
(high vs. low) | 2.02
(1.39-2.95) | <0.001 | 1.94
(1.31-2.89) | 0.001 |
Cancer immunotherapy has revolutionized cancer
treatment. Antibodies against programmed death 1/ligand 1 and
cytotoxic T-lymphocyte-associated protein 4 are effective for the
treatment of HCC (43–45). Given that the immune
microenvironment has an important role in the response to
immunotherapy (46,47), the impact of
FH-PH expression on the immune
microenvironment in patients with HCC was evaluated. Low activation
of the transforming growth factor (TGF)-β pathway is associated
with better clinical outcomes for patients with cancer (32). Thus, the TGF-β response score
(48) were applied in patients
with HCC and it was observed that the FH-PH
group had a relatively higher score (P<0.001; Fig. 3F). Furthermore, CIBERSORT (49) has been used to analyze immune cell
infiltration. Its application in the present study suggested that
regulatory T (Treg) cells were significantly enriched in the
FH-PH group (P<0.001; Fig. 3G). Of note, Treg cells are able to
inhibit T-cell proliferation and cytokine production and have a
critical role in preventing tumor immune response (50,51).
These results suggested that FH-PH expression
is associated with a suppressive immune microenvironment and may
hamper immunotherapy treatment in HCC.
FOXM1 and PLK1 are required for HCC
cell proliferation
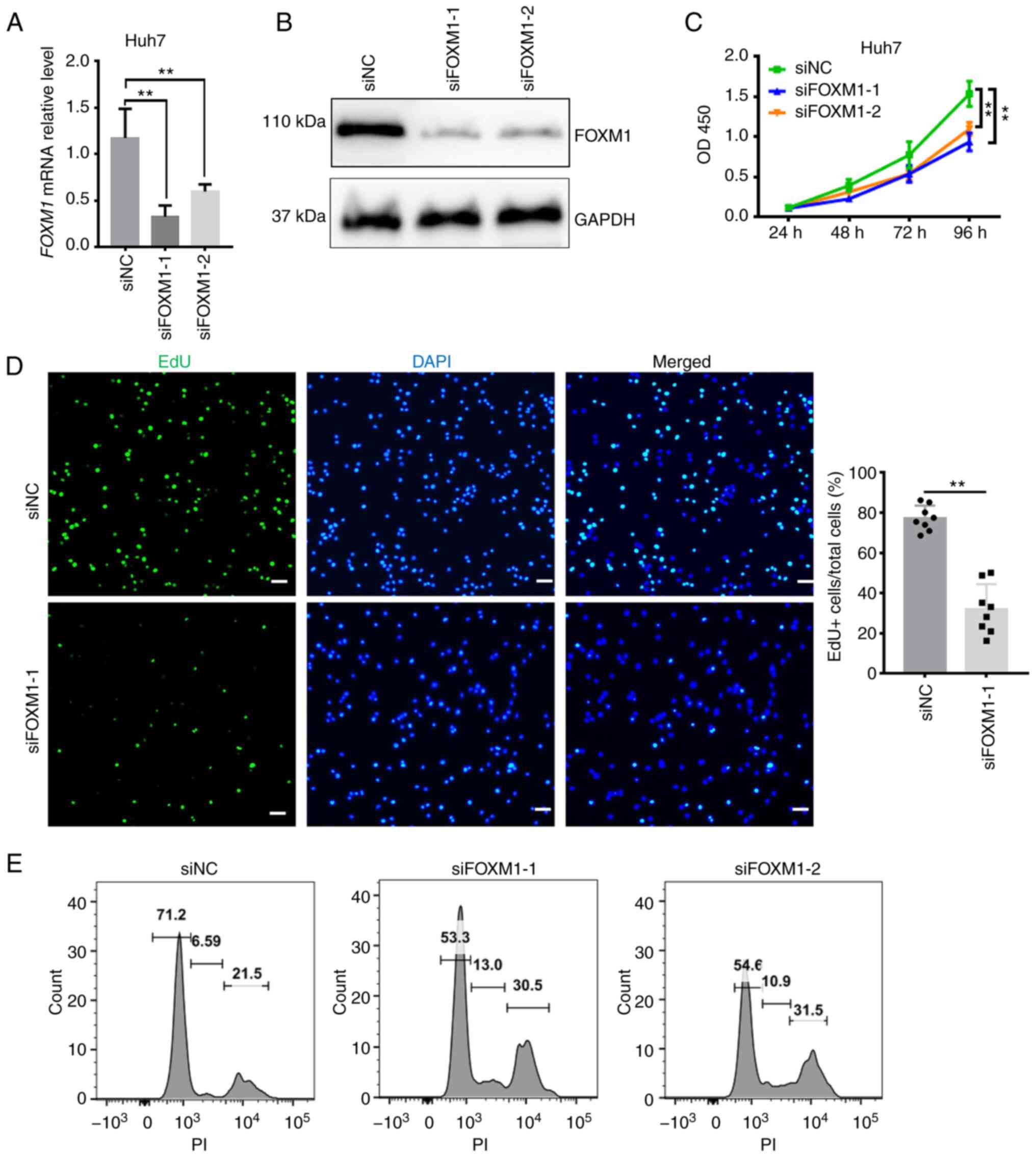
Given that the overexpression of FOXM1 and PLK1 was
associated with poor outcomes of HCC and based on their key
regulatory roles in cell cycle progression, it was investigated
whether FOXM1 and PLK1 influence HCC progression (Figs. 4 and 5). siRNA targeting FOXM1 and PLK1 was
synthesized and the knockdown efficiency was measured in Huh7 cells
(Figs. 4A and B and 5A and B). The cell proliferation rate was
estimated using CCK-8 and EdU assays. FOXM1 knockdown decreased
Huh7 cell viability (Fig. 4C), as
well as the percentage of EdU-positive Huh7 cells (Fig. 4D), indicating a lower proportion of
cells entering the DNA replication phase of the cell cycle. PLK1
knockdown exerted a similar effect on Huh7 cells (Fig. 5C and E). To confirm the requirement
of PLK1 for Huh7 cell proliferation, a pharmacological inhibitor of
PLK1, BI 2536 (38), was applied
during the CCK-8 assay. In the presence of the inhibitor, a
significant decrease in the proliferation of Huh7 cells was
observed (Fig. 5D). Furthermore,
flow cytometry suggested that knockdown of FOXM1 (Fig. 4E) or PLK1 (Fig. 5F) resulted in a marked increase in
the proportion of cells in S and G2/M phase, as well as a decrease
in the proportion of cells in G1 phase at 24 h after transfection.
It appears that, as more cells progress away from G0/G1 phase, the
cells are trying to proliferate but they exhibit cell cycle arrest
in S and G2/M phase. Consistent with this observation, Huh7 cells
treated with BI 2536 also exhibited a marked increase in the
proportion of cells in S and G2/M phase (Fig. 5G). These results suggested that
FOXM1 and PLK1 are required for HCC cell proliferation.
Insufficient FOXM1 or PLK1 were thus indicated to hamper cell cycle
progression as well as the proliferation of HCC cells, providing
additional evidence that the coordinated overexpression of FOXM1
and PLK1 is an independent prognostic factor for HCC.
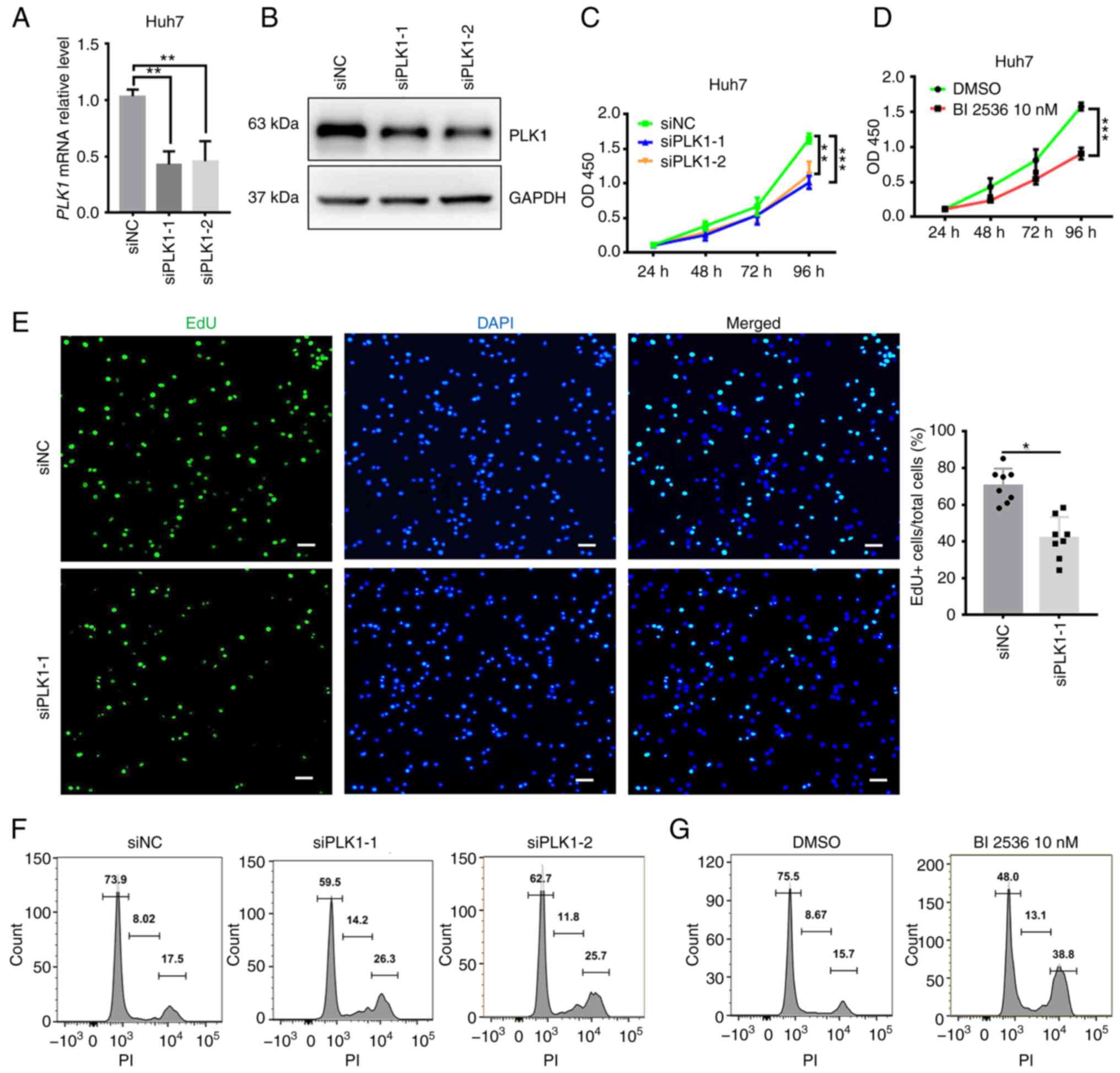 | Figure 5.PLK1 is required for Huh7 cell
proliferation. Knockdown efficiency of PLK1 confirmed using (A)
reverse transcription-quantitative PCR and (B) western blot
analysis. Cell proliferation rate analyzed using a Cell Counting
Kit-8 assay after (C) transfection with siPLK1 or (D) treatment
with 10 nM BI 2536 in Huh7 cells. (E) Decrease in the proliferation
rate of Huh7 cells following PLK1 knockdown as indicated by EdU
staining (scale bars, 50 µm). DNA content of Huh7 cell analyzed via
flow cytometry 24 h after (F) transfection with siPLK1 or (G)
treatment with 10 nM BI 2536. Results are provided in histograms.
Quantified values are expressed as the mean ± standard deviation of
three independent experiments. *P<0.05, **P<0.01,
***P<0.001 vs. siNC. PLK1, polo-like kinase 1; siRNA, small
interfering RNA; siNC, negative control siRNA; siPLK1, siRNA
targeting PLK1; PI, propidium iodide; EdU,
5-ethynyl-2′-deoxyuridine; OD450, optical density at 450 nm. |
Discussion
HCC is the primary malignancy of the liver and
complete surgical resection is the only curative approach. However,
most patients with HCC are only diagnosed when the disease is
already in the advanced stage, which is unsuitable for surgery. In
addition, the prognosis associated with the systemic treatment of
HCC is poor (3) and valuable
prognostic biomarkers are urgently required.
Cell cycle dysfunction is a marked feature of tumor
cells (52) and several regulators
of the cell cycle have been proposed as putative prognostic
biomarkers for cancer (53). In
this regard, FOXM1 and PLK1, which are essential cell cycle
regulators with prognostic value in HCC, have been extensively
studied. Consistent with previous reports (23,35),
the present results suggested that both FOXM1 and PLK1 were
overexpressed in tumor tissues and associated with poor prognosis
in patients with HCC. However, after adjusting for age, sex, stage
and virus status, a multivariate Cox proportional hazards model
indicated that neither FOXM1 nor PLK1 is able to independently
serve as a prognostic factor for HCC.
PLK1 is a target of FOXM1 and is required for FOXM1
transcriptional activation. A positive linear correlation between
FOXM1 and PLK1 expression was observed in HCC tissues. Insufficient
FOXM1 or PLK1 may hamper the cell cycle and it may be speculated
that the coordinated overexpression of FOXM1 and PLK1 may serve as
an independent prognostic factor for HCC. The present results
indicated that FH-PH expression was
associated with significantly shorter OS and DFS in patients with
HCC. In addition, after adjusting for age, sex, stage and virus
status, an FH-PH expression status was
indicated to be the most significant prognostic factor for patients
with HCC. Furthermore, an FH-PH expression
status was associated with lower TGF-β response scores and a higher
number of infiltrating Treg cells, suggesting that patients with
HCC with FH-PH expression status harbor a
suppressed immune microenvironment that leads to treatment
failure.
In the present study, the requirement for FOXM1 and
PLK1 expression in HCC cells was investigated via in vitro
knockdown of FOXM1 and PLK1 in Huh7 cells. It was observed that
either FOXM1 or PLK1 knockdown was able to hamper cell cycle
progression as well as the proliferation of Huh7 cells. The
antitumor activity of pharmacological inhibitors of PLK1 has been
evaluated in clinical trials, but the efficacy was limited
(54–56). The limited antitumor activity may
be attributed to the low PLK1 or FOXM1 expression levels in these
patients. For these patients, PLK1 was not the essential
tumor-growth driving factor. Thus, the present results suggested
that, to improve therapeutic efficacy in the future, it may be
necessary to perform clinical trials involving patients with
coordinated overexpression of FOXM1 and PLK1.
In conclusion, the present results indicated that
FOXM1 and PLK1 were overexpressed in HCC tumor tissues and
exhibited a positive linear correlation. FOXM1 and PLK1 are
required for HCC cell proliferation. The present results also
indicated that FH and PH expression were
associated with poor prognosis for HCC; however, only the
coordinated overexpression of FOXM1 and PLK1 was able to serve as
an independent prognostic factor for HCC. Therefore, targeting
FOXM1 or PLK1 is a potential treatment for improving HCC
prognosis.
Acknowledgements
Not applicable.
Funding
This work was supported by the Shandong Province Key Research
Program (grant no. 2019GSF108012).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BJ conceived and designed the study and also
provided administrative support. WF and HM provided the study
materials, performed the experiments and collected the public data.
All the authors performed data analysis and interpretation,
participated in writing the manuscript and all authors read and
approved the final version of the manuscript. WF and HM confirm the
authenticity of all the raw data. The authors are accountable for
all aspects of the work in ensuring that questions related to the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Singal AG, Lampertico P and Nahon P:
Epidemiology and surveillance for hepatocellular carcinoma: New
trends. J Hepatol. 72:250–261. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Faivre S, Rimassa L and Finn RS: Molecular
therapies for HCC: Looking outside the box. J Hepatol. 72:342–352.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sangro B, Sarobe P, Hervás-Stubbs S and
Melero I: Advances in immunotherapy for hepatocellular carcinoma.
Nat Rev Gastroenterol Hepatol. 18:525–543. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Song BN and Chu IS: A gene expression
signature of FOXM1 predicts the prognosis of hepatocellular
carcinoma. Exp Mol Med. 50:e4182018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Teng L, Wang K, Liu Y, Ma Y, Chen W and Bi
L: Based on integrated bioinformatics analysis identification of
biomarkers in hepatocellular carcinoma patients from different
regions. Biomed Res Int. 2019:17423412019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Agarwal R, Narayan J, Bhattacharyya A,
Saraswat M and Tomar AK: Gene expression profiling, pathway
analysis and subtype classification reveal molecular heterogeneity
in hepatocellular carcinoma and suggest subtype specific
therapeutic targets. Cancer Genet. 216–217. 37–51. 2017.PubMed/NCBI
|
|
8
|
Sun W, Su Q, Cao X, Shang B, Chen A, Yin H
and Liu B: High expression of polo-like kinase 1 is associated with
early development of hepatocellular carcinoma. Int J Genomics.
2014:3121302014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Seyedabadi S, Saidijam M, Najafi R,
Mousavi-Bahar SH, Jafari M, MohammadGanji S and Mahdavinezhad A:
Assessment of CEP55, PLK1 and FOXM1 expression in patients with
bladder cancer in comparison with healthy individuals. Cancer
Invest. 36:407–414. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dibb M, Han N, Choudhury J, Hayes S,
Valentine H, West C, Sharrocks AD and Ang YS: FOXM1 and polo-like
kinase 1 are co-ordinately overexpressed in patients with gastric
adenocarcinomas. BMC Res Notes. 8:6762015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang R, Song Y, Xu X, Wu Q and Liu C: The
expression of Nek7, FoxM1, and Plk1 in gallbladder cancer and their
relationships to clinicopathologic features and survival. Clin
Transl Oncol. 15:626–632. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lam EW, Brosens JJ, Gomes AR and Koo CY:
Forkhead box proteins: Tuning forks for transcriptional harmony.
Nat Rev Cancer. 13:482–495. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang IC, Chen YJ, Hughes D, Petrovic V,
Major ML, Park HJ, Tan Y, Ackerson T and Costa RH: Forkhead box M1
regulates the transcriptional network of genes essential for
mitotic progression and genes encoding the SCF (Skp2-Cks1)
ubiquitin ligase. Mol Cell Biol. 25:10875–10894. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Laoukili J, Kooistra MR, Brás A, Kauw J,
Kerkhoven RM, Morrison A, Clevers H and Medema RH: FoxM1 is
required for execution of the mitotic programme and chromosome
stability. Nat Cell Biol. 7:126–136. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen YJ, Dominguez-Brauer C, Wang Z, Asara
JM, Costa RH, Tyner AL, Lau LF and Raychaudhuri P: A conserved
phosphorylation site within the forkhead domain of FoxM1B is
required for its activation by cyclin-CDK1. J Biol Chem.
284:30695–30707. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fu Z, Malureanu L, Huang J, Wang W, Li H,
van Deursen JM, Tindall DJ and Chen J: Plk1-dependent
phosphorylation of FoxM1 regulates a transcriptional programme
required for mitotic progression. Nat Cell Biol. 10:1076–1082.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang J, Yuan C, Wu J, Elsayed Z and Fu Z:
Polo-like kinase 1-mediated phosphorylation of forkhead box protein
M1b antagonizes its SUMOylation and facilitates its mitotic
function. J Biol Chem. 290:3708–3719. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pilarsky C, Wenzig M, Specht T, Saeger HD
and Grützmann R: Identification and validation of commonly
overexpressed genes in solid tumors by comparison of microarray
data. Neoplasia. 6:744–750. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim IM, Ackerson T, Ramakrishna S,
Tretiakova M, Wang IC, Kalin TV, Major ML, Gusarova GA, Yoder HM,
Costa RH and Kalinichenko VV: The Forkhead Box m1 transcription
factor stimulates the proliferation of tumor cells during
development of lung cancer. Cancer Res. 66:2153–2161. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Z, Park HJ, Carr JR, Chen YJ, Zheng
Y, Li J, Tyner AL, Costa RH, Bagchi S and Raychaudhuri P: FoxM1 in
tumorigenicity of the neuroblastoma cells and renewal of the neural
progenitors. Cancer Res. 71:4292–4302. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu G, Yan Z, Zhang C, Cheng M, Yan Y, Wang
Y, Deng L, Lu Q and Luo S: FOXM1 promotes hepatocellular carcinoma
progression by regulating KIF4A expression. J Exp Clin Cancer Res.
38:1882019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chai N, Xie HH, Yin JP, Sa KD, Guo Y, Wang
M, Liu J, Zhang XF, Zhang X, Yin H, et al: FOXM1 promotes
proliferation in human hepatocellular carcinoma cells by
transcriptional activation of CCNB1. Biochem Biophys Res Commun.
500:924–929. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Egawa M, Yoshida Y, Ogura S, Kurahashi T,
Kizu T, Furuta K, Kamada Y, Chatani N, Hamano M, Kiso S, et al:
Increased expression of Forkhead box M1 transcription factor is
associated with clinicopathological features and confers a poor
prognosis in human hepatocellular carcinoma. Hepatol Res.
47:1196–1205. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun HC, Li M, Lu JL, Yan DW, Zhou CZ, Fan
JW, Qin XB, Tang HM and Peng ZH: Overexpression of Forkhead box M1
protein associates with aggressive tumor features and poor
prognosis of hepatocellular carcinoma. Oncol Rep. 25:1533–1539.
2011.PubMed/NCBI
|
|
25
|
Combes G, Alharbi I, Braga LG and Elowe S:
Playing polo during mitosis: PLK1 takes the lead. Oncogene.
36:4819–4827. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Colicino EG and Hehnly H: Regulating a key
mitotic regulator, polo-like kinase 1 (PLK1). Cytoskeleton
(Hoboken). 75:481–494. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Barr FA, Silljé HH and Nigg EA: Polo-like
kinases and the orchestration of cell division. Nat Rev Mol Cell
Biol. 5:429–440. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xie S, Xie B, Lee MY and Dai W: Regulation
of cell cycle checkpoints by polo-like kinases. Oncogene.
24:277–286. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He Z, Wu J, Dang H, Lin H, Zheng H and
Zhong D: Polo-like kinase 1 contributes to the tumorigenicity of
BEL-7402 hepatoma cells via regulation of Survivin expression.
Cancer Lett. 303:92–98. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Park JS, Sohn HJ, Park GS, Chung YJ and
Kim TG: Induction of antitumor immunity using dendritic cells
electroporated with Polo-like kinase 1 (Plk1) mRNA in murine tumor
models. Cancer Sci. 102:1448–1454. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jang HR, Shin SB, Kim CH, Won JY, Xu R,
Kim DE and Yim H: PLK1/vimentin signaling facilitates immune escape
by recruiting Smad2/3 to PD-L1 promoter in metastatic lung
adenocarcinoma. Cell Death Differ. 28:2745–2764. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fristrup N, Ulhøi BP, Birkenkamp-Demtröder
K, Mansilla F, Sanchez-Carbayo M, Segersten U, Malmström PU,
Hartmann A, Palou J, Alvarez-Múgica M, et al: Cathepsin E, maspin,
Plk1, and survivin are promising prognostic protein markers for
progression in non-muscle invasive bladder cancer. Am J Pathol.
180:1824–1834. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Takai N, Hamanaka R, Yoshimatsu J and
Miyakawa I: Polo-like kinases (Plks) and cancer. Oncogene.
24:287–291. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheng L, Wang C and Jing J: Polo-like
kinase 1 as a potential therapeutic target for osteosarcoma. Curr
Pharm Des. 21:1347–1350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He ZL, Zheng H, Lin H, Miao XY and Zhong
DW: Overexpression of polo-like kinase1 predicts a poor prognosis
in hepatocellular carcinoma patients. World J Gastroenterol.
15:4177–4182. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tian L, Yao K, Liu K, Han B, Dong H, Zhao
W, Jiang W, Qiu F, Qu L, Wu Z, et al: PLK1/NF-κB feedforward
circuit antagonizes the mono-ADP-ribosyltransferase activity of
PARP10 and facilitates HCC progression. Oncogene. 39:3145–3162.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gumireddy K, Reddy MV, Cosenza SC,
Boominathan R, Baker SJ, Papathi N, Jiang J, Holland J and Reddy
EP: ON01910, a non-ATP-competitive small molecule inhibitor of
Plk1, is a potent anticancer agent. Cancer Cell. 7:275–286. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Steegmaier M, Hoffmann M, Baum A, Lénárt
P, Petronczki M, Krssák M, Gürtler U, Garin-Chesa P, Lieb S, Quant
J, et al: BI 2536, a potent and selective inhibitor of polo-like
kinase 1, inhibits tumor growth in vivo. Curr Biol. 17:316–322.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rudolph D, Steegmaier M, Hoffmann M,
Grauert M, Baum A, Quant J, Haslinger C, Garin-Chesa P and Adolf
GR: BI 6727, a Polo-like kinase inhibitor with improved
pharmacokinetic profile and broad antitumor activity. Clin Cancer
Res. 15:3094–3102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Qiu P and Sheng J: A two-stage procedure
for comparing hazard rate functions. J R Stat Soc Series B (Stat
Methodol). 70:191–208. 2008.PubMed/NCBI
|
|
42
|
Li H, Han D, Hou Y, Chen H and Chen Z:
Statistical inference methods for two crossing survival curves: A
comparison of methods. PLoS One. 10:e01167742015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Duffy AG, Ulahannan SV, Makorova-Rusher O,
Rahma O, Wedemeyer H, Pratt D, Davis JL, Hughes MS, Heller T,
ElGindi M, et al: Tremelimumab in combination with ablation in
patients with advanced hepatocellular carcinoma. J Hepatol.
66:545–551. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Finn RS, Qin S, Ikeda M, Galle PR, Ducreux
M, Kim TY, Kudo M, Breder V, Merle P, Kaseb AO, et al: Atezolizumab
plus bevacizumab in unresectable hepatocellular carcinoma. N Engl J
Med. 382:1894–1905. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ren Z, Xu J, Bai Y, Xu A, Cang S, Du C, Li
Q, Lu Y, Chen Y, Guo Y, et al: Sintilimab plus a bevacizumab
biosimilar (IBI305) versus sorafenib in unresectable hepatocellular
carcinoma (ORIENT-32): A randomised, open-label, phase 2–3 study.
Lancet Oncol. 22:977–990. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cariani E and Missale G: Immune landscape
of hepatocellular carcinoma microenvironment: Implications for
prognosis and therapeutic applications. Liver Int. 39:1608–1621.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Oura K, Morishita A, Tani J and Masaki T:
Tumor Immune microenvironment and immunosuppressive therapy in
hepatocellular carcinoma: A review. Int J Mol Sci. 22:58012021.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Thorsson V, Gibbs DL, Brown SD, Wolf D,
Bortone DS, Ou Yang TH, Porta-Pardo E, Gao GF, Plaisier CL, Eddy
JA, et al: The immune landscape of cancer. Immunity.
48:812–830.e14. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Newman AM, Liu CL, Green MR, Gentles AJ,
Feng W, Xu Y, Hoang CD, Diehn M and Alizadeh AA: Robust enumeration
of cell subsets from tissue expression profiles. Nat Methods.
12:453–457. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kondĕlková K, Vokurková D, Krejsek J,
Borská L, Fiala Z and Ctirad A: Regulatory T cells (TREG) and their
roles in immune system with respect to immunopathological
disorders. Acta Medica (Hradec Kralove). 53:73–77. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hatzioannou A, Boumpas A, Papadopoulou M,
Papafragkos I, Varveri A, Alissafi T and Verginis P: Regulatory T
cells in autoimmunity and cancer: A duplicitous lifestyle. Front
Immunol. 12:7319472021. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Matthews HK, Bertoli C and de Bruin RAM:
Cell cycle control in cancer. Nat Rev Mol Cell Biol. 23:74–88.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kashyap D, Garg VK, Sandberg EN, Goel N
and Bishayee A: Oncogenic and tumor suppressive components of the
cell cycle in breast cancer progression and prognosis.
Pharmaceutics. 13:5692021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Mross K, Dittrich C, Aulitzky WE,
Strumberg D, Schutte J, Schmid RM, Hollerbach S, Merger M, Munzert
G, Fleischer F and Scheulen ME: A randomised phase II trial of the
Polo-like kinase inhibitor BI 2536 in chemo-naïve patients with
unresectable exocrine adenocarcinoma of the pancreas-a study within
the Central European society anticancer drug research (CESAR)
collaborative network. Br J Cancer. 107:280–286. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Garcia-Manero G, Fenaux P, Al-Kali A, Baer
MR, Sekeres MA, Roboz GJ, Gaidano G, Scott BL, Greenberg P,
Platzbecker U, et al: Rigosertib versus best supportive care for
patients with high-risk myelodysplastic syndromes after failure of
hypomethylating drugs (ONTIME): A randomised, controlled, phase 3
trial. Lancet Oncol. 17:496–508. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pujade-Lauraine E, Selle F, Weber B,
Ray-Coquard IL, Vergote I, Sufliarsky J, Del Campo JM, Lortholary
A, Lesoin A, Follana P, et al: Volasertib versus chemotherapy in
platinum-resistant or -refractory ovarian cancer: A randomized
phase II groupe des investigateurs nationaux pour l'Etude des
cancers de l'Ovaire study. J Clin Oncol. 34:706–713. 2016.
View Article : Google Scholar : PubMed/NCBI
|