Introduction
Malignant melanoma has an age-standardized mortality
rate of 0.3 per 100,000 in China (1), and it is considered one of the most
challenging cancers to be diagnosed owing to the limited medical
knowledge regarding the early-stage development of its lesions
(2). It has been proposed that
melanoma is produced by the transformation of melanocytes (3,4).
According to its general clinical and pathological characteristics,
melanoma can be divided into four subtypes, including superficial
diffuse, nodular, malignant and acromegaly melanoma (5–7).
Despite the fact that numerous primary melanomas can be
successfully treated by surgery, the treatment of patients with
advanced metastatic melanoma remains challenging (8,9). The
exact reasons responsible for the higher incidence of melanoma
remain unclear (10). In view of
this evidence, the present study was conducted to explore the
mechanism of malignant melanoma and to identify optimal therapeutic
strategies for its therapy.
Tepotinib (Tepmetko™) is a selective tyrosine kinase
inhibitor of MET proto-oncogene, receptor tyrosine kinase (MET)
developed by Sigma-Aldrich; Merck KGaA, which is often used to
treat solid tumors (11). Due to
its high retention in tumors, tepotinib can block MET as well as
its downstream pathways and serve as a potent drug for the
treatment of various types of cancer, such as gastric and non-small
cell lung cancer (12,13). MET is a receptor tyrosine kinase,
which exhibits abnormal activity in human cancer. The activity of
this enzyme is associated with aggressive cancer phenotypes,
metastatic dissemination, and poor disease prognosis (14–16).
It has been reported that c-MET is overexpressed in melanoma and
contributes to its malignant progression (17). The PI3K/AKT pathway plays a
critical role in cell cycle progression, which involves cellular
proliferation and cancer formation (18). An increasing number of studies have
revealed that the PI3K/AKT pathway can be a therapeutic target for
the treatment of various types of cancer, including triple-negative
breast cancer and melanoma (19,20).
The purpose of the present study was to investigate
the effects of tepotinib on melanoma cells and to examine its
specific mechanism of action. The data aimed to provide new
insights into the potential treatment of malignant melanoma.
Materials and methods
Cell culture and treatment
The human malignant melanoma cell line WM451 was
purchased from the Cell Bank of Central South University. The cells
were incubated in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C in a humidified incubator with 5%
CO2. Subsequently, different concentrations of tepotinib
(1, 2, 5, 10, 20 and 50 ng/ml; Selleck Chemicals) were used to
treat WM451 cells for 48 h (13).
For tepotinib + hepatocyte growth factor (HGF; R&D Systems,
Inc.)-treated group, 50 ng/ml of HGF (a natural agonist of MET) and
10 ng/ml tepotinib were concurrently administered to WM451 cells
for 48 h.
MTT assay
The proliferative capacity of WM451 cells was
evaluated using the MTT assay. WM451 cells were incubated in
96-well plates at a density of 5×104 cells/ml/well for
48 h at 37°C. Subsequently, tepotinib was used at different
concentrations (1, 2, 5, 10, 20 and 50 ng/ml) to treat WM451 cells.
Following exposure to a 10 µl MTT solution for 4 h, the cells were
incubated with formazan lysis solution (DMSO) until the purple
crystals were completely dissolved. Finally, the absorbance was
detected at 570 nm with a spectrophotometer.
Wound healing assay
WM451 cells were seeded in six-well plates
(6×104 cells/well) and cultured to 90–100% confluence. A
straight scratch was made in the cell monolayer with a 200-µl
pipette tip, followed by PBS washing three times. During the wound
healing assay, WM451 cells were cultured in serum-free DMEM in an
incubator at 37°C with 5% CO2 for 48 h. The width of the
wound was recorded and images were captured at 0 and 48 h time
points by using an inverted light microscope (Olympus Corporation).
Finally, the area occupied by the migratory cells in the linear
scratch was detected by ImageJ Software (version 6.0; National
Institutes of Health).
Transwell assay
The invasive ability of WM451 cells was detected
using a Transwell plate with an 8-µm pore insert precoated with
Matrigel (BD Biosciences) at room temperature for 1 h. The cells
were plated at a density of 4×104 cells into the upper
compartment of the Transwell chamber with serum-free DMEM.
Subsequently, the lower compartment of the Transwell chamber was
filled with 500 µl DMEM supplemented with 10% FBS; the cells were
incubated for 24 h at 37°C. Subsequently, the invaded cells were
fixed with 4% paraformaldehyde for 15 min at room temperature, and
stained with 0.1% crystal violet at room temperature for 5 min.
Finally, the visualization of the invasive cells was observed with
an inverted light microscope (Olympus Corporation).
TUNEL assay
The effects of tepotinib (10 ng/ml) on the induction
of WM451 cell apoptosis of were assessed with a TUNEL assay kit
(Invitrogen; Thermo Fisher Scientific, Inc.) in accordance with the
manufacturer's protocol. A total of 2×105 cells/well in
a 24-well plate were treated with tepotinib for 48 h and
subsequently fixed with 4% paraformaldehyde for 15 min at room
temperature and then permeabilized with 0.25% Triton X-100 for 2
min at 4°C. Subsequently, the cells were labeled with TUNEL at 37°C
for 60 min and counterstained with 0.01 mg/ml DAPI for 10 min in
the dark. Slides were mounted using glycerol, and images of the
TUNEL-positive apoptotic cells in six randomly selected fields of
view were captured with a fluorescence microscope (Nikon
Corporation) and quantified by ImageJ Software (version 6.0;
National Institutes of Health).
Western blot analysis
Total protein from WM451 cells was isolated using
RIPA lysis buffer (Beyotime Institute of Biotechnology) and
quantified using the bicinchoninic acid method (Beyotime Institute
of Biotechnology). The proteins (40 µg per lane) were separated
with 10% SDS-PAGE and transferred on polyvinylidene fluoride
membranes. Following blocking with 5% skimmed milk for 2 h at room
temperature, specific primary antibodies were incubated with the
membranes at 4°C overnight. Following the primary incubation, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibody (1:3,000; cat. no. 7074S; Cell Signaling
Technology, Inc.) for 2 h at room temperature. Finally, an enhanced
chemiluminescence kit (Beyotime Institute of Biotechnology) was
applied to determine the expression levels of the protein bands.
The intensities of the bands were detected by ImageJ Software
(version 6.0; National Institutes of Health). GAPDH was used as an
internal control. The following primary antibodies were diluted to
1:1,000 in TBS + 0.1% Tween-20 buffer and used: E-cadherin (cat.
no. 3195T), N-cadherin (cat. no. 13116T), vimentin (cat. no.
5741T), Bcl-2 (cat. no. 4223T), Bax (cat. no. 5023T), cleaved
caspase 3 (cat. no. 9664T), caspase 3 (cat. no. 14220T), phospho
(p)-MET (cat. no. 3077T), MET (cat. no. 4560S), PI3K (cat. no.
4249T), p-AKT (cat. no. 4060T), AKT (cat. no 4685S) GAPDH (cat. no.
5174T; all from Cell Signaling Technology, Inc.) and p-PI3K (cat.
no. ab278545; Abcam).
Statistical analysis
All data collected from the experiments are
displayed as the mean ± standard deviation and assessed with
GraphPad Prism 8.0 (GraphPad Software, Inc.). The comparisons
between two groups were conducted with the unpaired Student's
t-test. One-way analysis of variance followed by Tukey's post hoc
test was applied to determine the statistical significance among
multiple groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Tepotinib inhibits the proliferation,
migration and invasion of melanoma cells
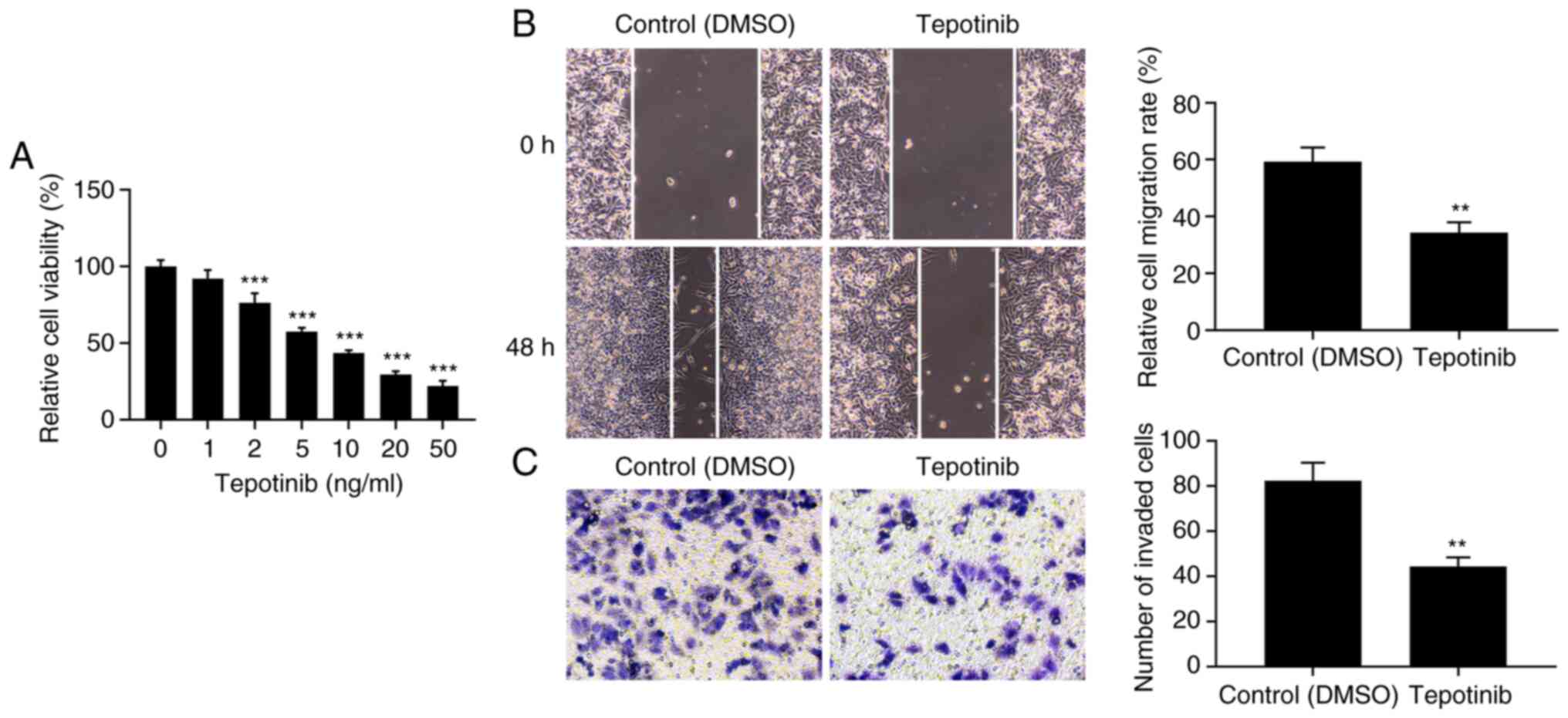
Initially, the effects of tepotinib were
investigated on the cell proliferation of WM451 cells. The
proliferation of WM451 cells was significantly decreased by
tepotinib treatment in comparison with the control group (Fig. 1A). In addition, the inhibitory
effects of tepotinib on the proliferation of WM451 cells occurred
in a concentration-dependent manner. A total of 10 ng/ml tepotinib
presented ~50% inhibitory effect to WM451 cell proliferation,
therefore, this concentration was selected to perform the
subsequent experiments. The migratory and invasive activities of
tepotinib-treated WM451 cells were detected with the application of
wound healing and Transwell assays, respectively. The results from
Fig. 1B demonstrated that
treatment with 10 ng/ml tepotinib significantly diminished the
migration of WM451 cells compared with that of the control group.
Similarly, the invasive activity of WM451 cells was also reduced by
tepotinib (Fig. 1C). These results
suggested that tepotinib could suppress the proliferation,
migration and invasion of melanoma cells.
Tepotinib suppresses the
epithelial-mesenchymal transition (EMT) process of melanoma
cells
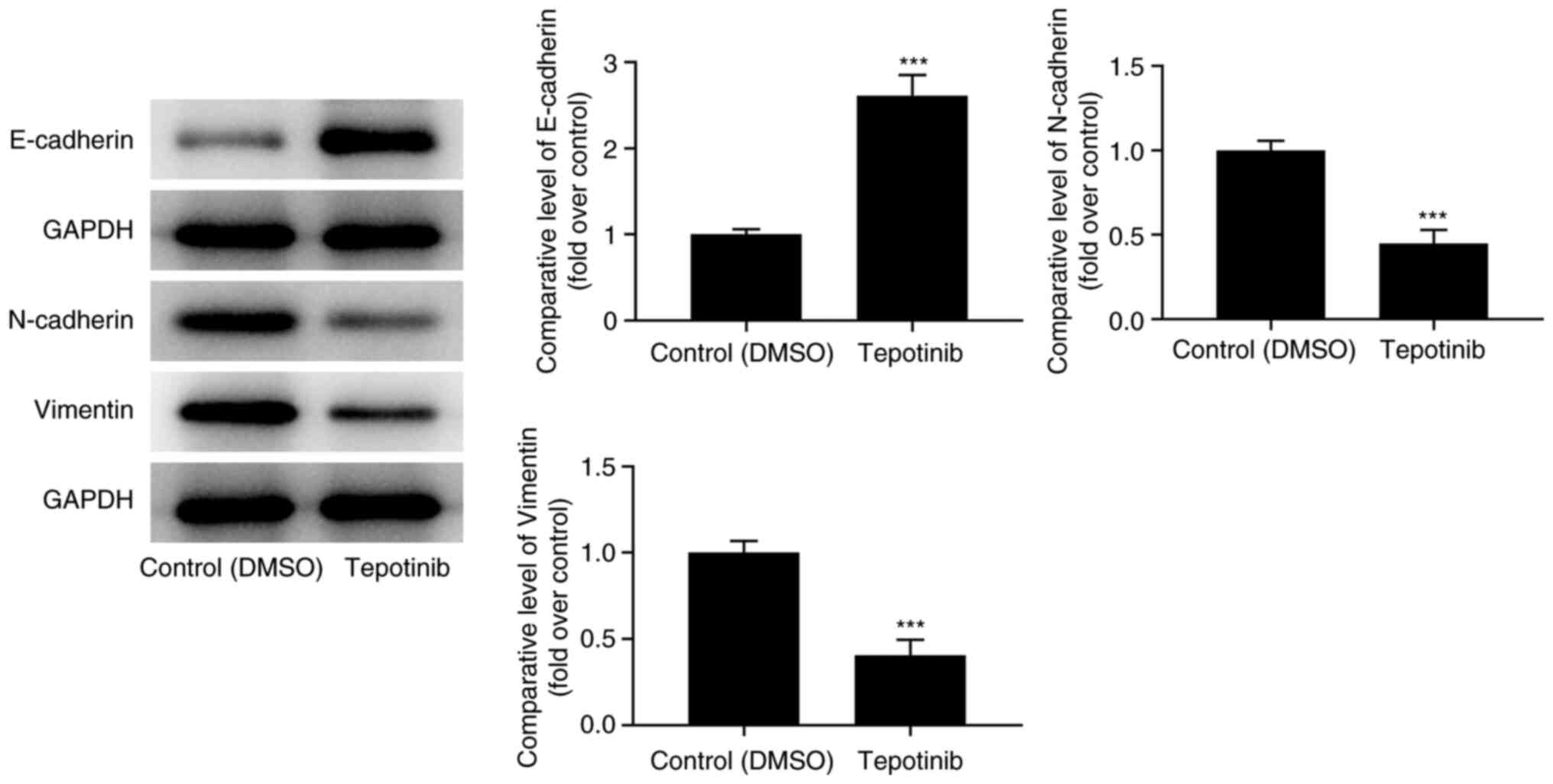
The expression levels of EMT-related proteins were
assessed with western blot analysis. Tepotinib treatment caused an
apparent upregulation of E-cadherin expression levels, while
downregulating N-cadherin and vimentin expression levels compared
with those of the control group (Fig.
2). In summary, the aforementioned results suggested that
tepotinib exerted inhibitory effects on the EMT process of
malignant melanoma cells.
Tepotinib promotes the apoptosis of
melanoma cells
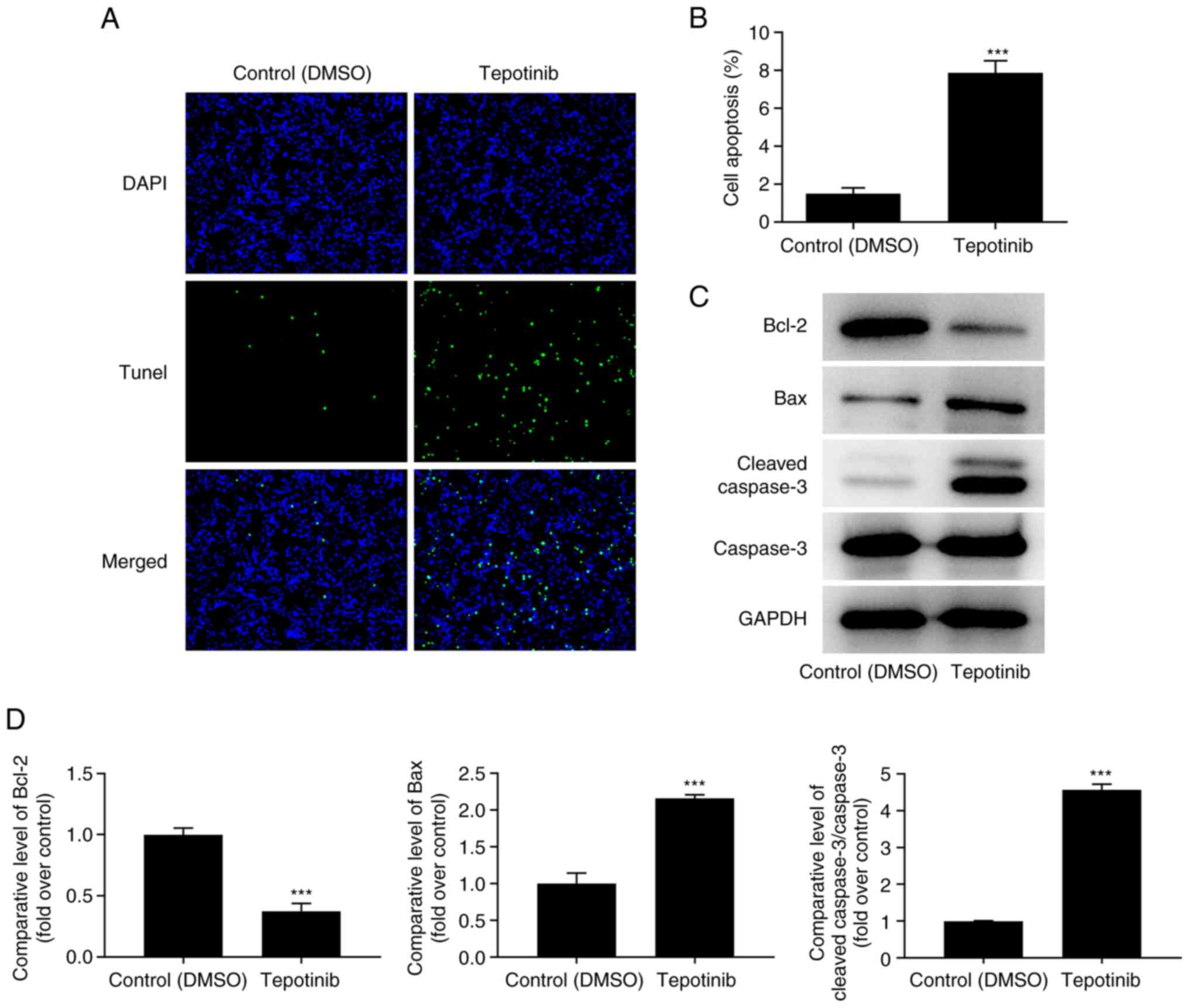
The induction of apoptosis of tepotinib-treated
WM451 cells was evaluated using TUNEL staining. The induction of
apoptosis of WM451 cells was significantly increased following
tepotinib (10 ng/ml) treatment compared with that of the control
group, revealing that this compound could induce apoptosis of WM451
cells (Fig. 3A and B). In
addition, the expression levels of the apoptosis-related proteins
were detected by western blot analysis and the results indicated
that tepotinib significantly downregulated Bcl-2 expression, while
upregulating the expression levels of Bax and cleaved caspase 3 in
comparison with the corresponding effects noted in the control
group (Fig. 3C and D). In
conclusion, the data indicated that tepotinib induced apoptosis of
melanoma cells.
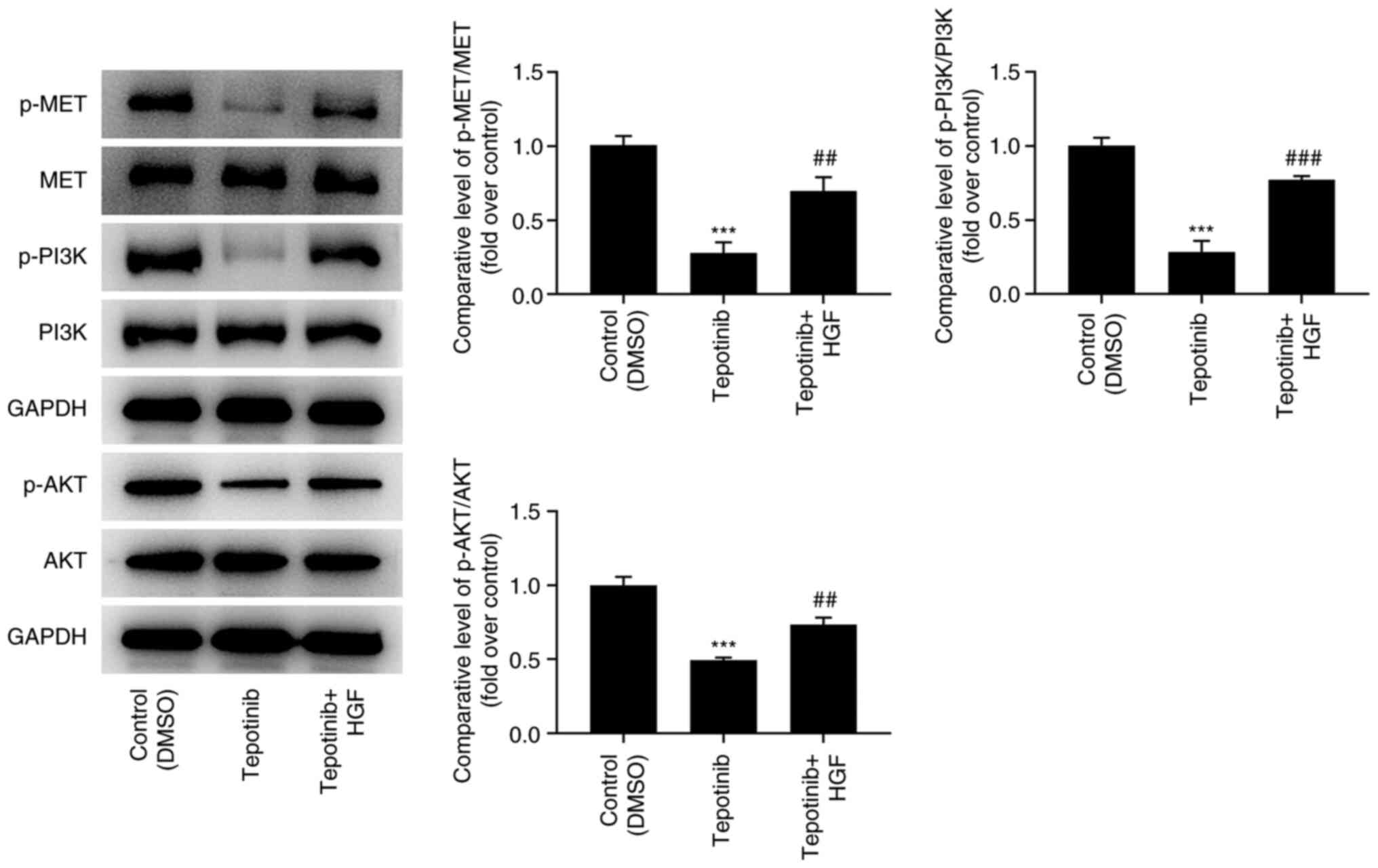
Tepotinib reduces activation of MET
and PI3K/AKT signaling pathways
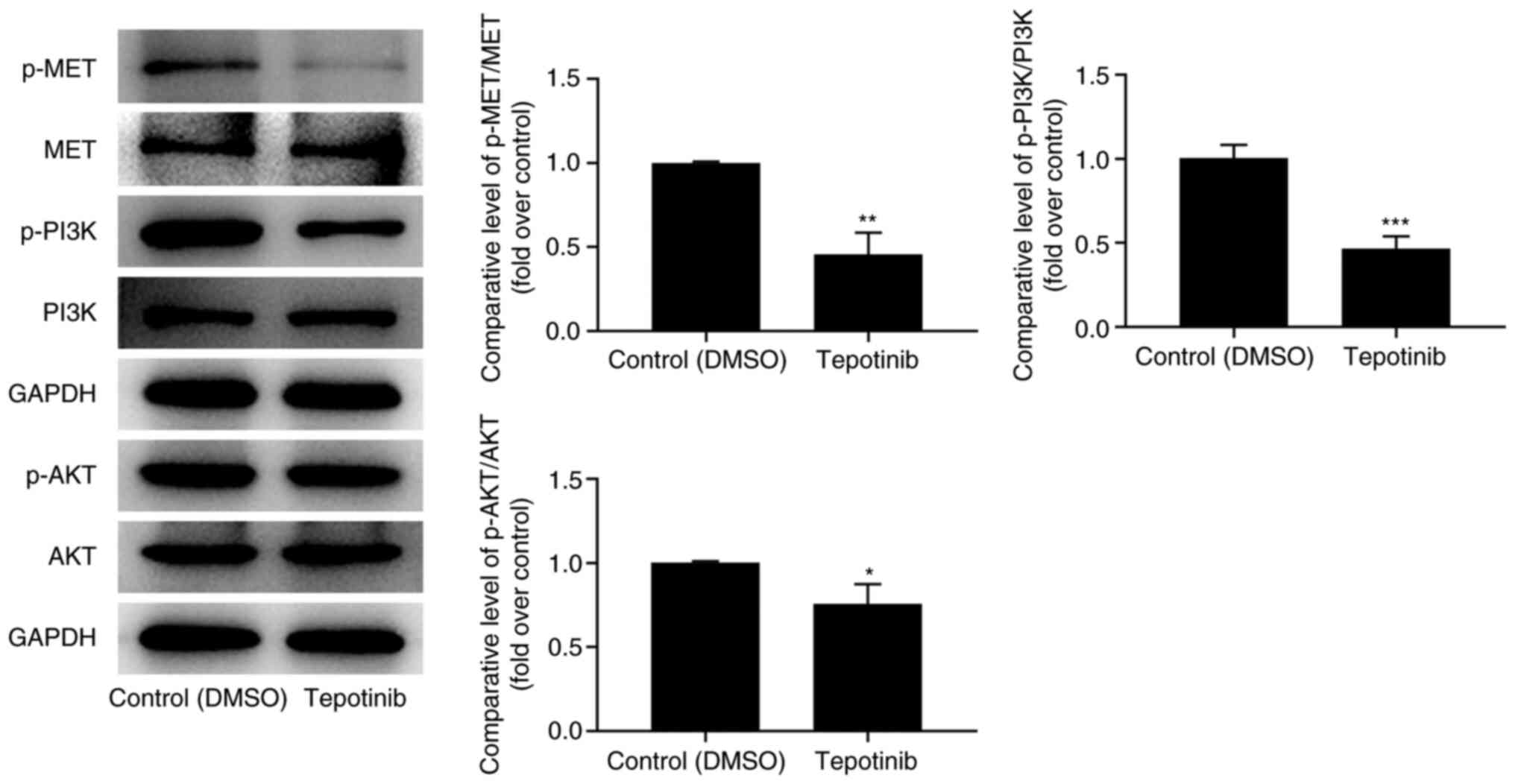
To investigate the effects of tepotinib on MET and
PI3K/AKT signaling pathways, western blot analysis was utilized to
measure the expression levels of MET, phosphorylated (p)-MET, PI3K,
p-PI3K, AKT and p-AKT. Tepotinib diminished the expression levels
of MET, p-MET, p-PI3K and p-AKT compared with those noted in the
control group, suggesting that it exhibited inhibitory effects on
activation of MET and PI3K/AKT signaling pathways (Fig. 4).
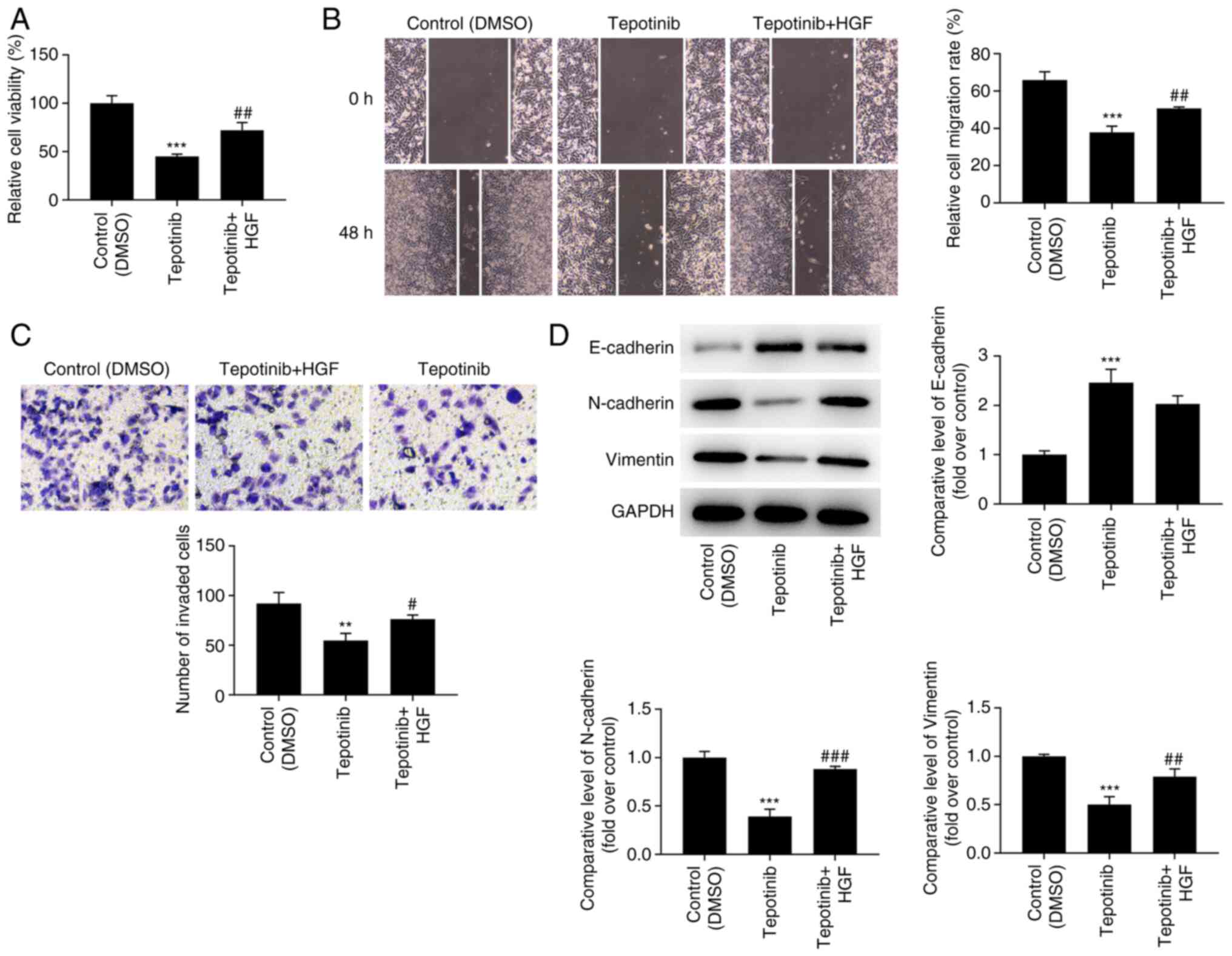
Tepotinib inhibits the proliferation,
migration and invasion of WM451 cells by reducing activation of MET
and PI3K/AKT signaling pathways
In order to assess the detailed mechanism of action
of tepotinib, HGF (50 ng/ml), which is a natural agonist of MET,
was administered to WM451 cells for 48 h. As exhibited in Fig. 5, the addition of HGF significantly
upregulated the expression of p-MET, p-PI3K and p-AKT when compared
with the tepotinib group. Moreover, tepotinib inhibited the
proliferative ability of WM451 cells, which was partially reversed
by HGF, indicating the beneficial effects of this factor on the
proliferation of tepotinib-treated WM451 cells (Fig. 6A). Tepotinib considerably
diminished the relative migration rate of WM451 cells, while HGF
administration partially abolished the suppressive effects of
tepotinib, as demonstrated by the increased migration of the
tepotinib + HGF cell group in comparison with that noted in the
tepotinib cell group (Fig. 6B).
Similarly, the declined invasive activity of tepotinib-treated
WM451 cells was subsequently enhanced following HGF administration
(Fig. 6C). Besides, tepotinib
increased E-cadherin expression, while reducing the expression
levels of N-cadherin and vimentin, whereas HGF administration
exhibited the opposite effects, as determined by the decreased
expression levels of E-cadherin and the increased expression levels
of N-cadherin and vimentin in the tepotinib + HGF group (Fig. 6D).
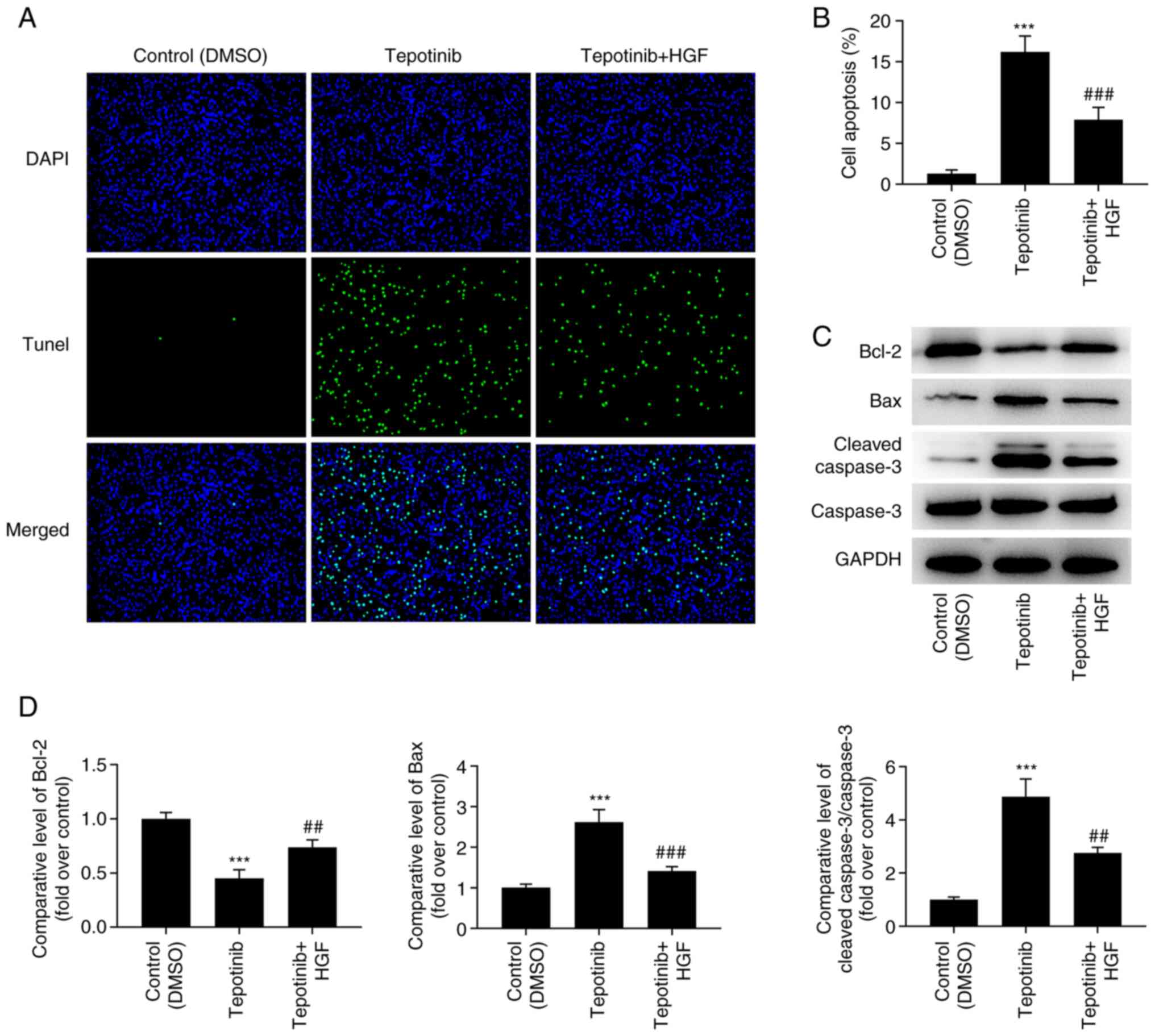
Tepotinib promotes the induction of
apoptosis of WM451 cells by reducing the activation of the MET
pathway and inhibiting the PI3K/AKT signaling pathway
According to the results obtained from the TUNEL
assay, the increased apoptotic levels caused by tepotinib treatment
were significantly diminished by HGF administration (Fig. 7A and B). In addition, the
expression levels of the apoptosis-related proteins were measured
by western blot analysis and the results revealed that tepotinib
significantly downregulated Bcl-2 expression levels, while causing
a significant upregulation in the expression levels of Bax and
cleaved caspase 3. Nevertheless, HGF administration exerted the
opposite effects on these proteins, as determined by the increased
Bcl-2 expression levels as well as the decreased expression levels
of Bax and cleaved caspase 3 in the tepotinib + HGF group (Fig. 7C and D).
Discussion
To the best of our knowledge, the present study is
the first to explore the effects of tepotinib on melanoma. The
effects of this compound on the proliferation of WM451 cells were
assessed and the results indicated that it may suppress the
proliferative ability of WM451 cells in a concentration-dependent
manner. Subsequently, a series of cellular experiments were
conducted to examine the effects of tepotinib on melanoma cells.
The data indicated that tepotinib exerted suppressive effects on
the migration, invasion, and the EMT, while promoting the induction
of apoptosis of WM451 cells. It was also shown that tepotinib
diminished the expression levels of p-MET, p-PI3K and p-AKT,
indicating the inhibitory effects of this compound on the
activation of MET and PI3K/AKT signaling pathways. To further
assess the mechanism of action of tepotinib, HGF, an agonist of
MET, was applied to treat WM451 cells. The data indicated that the
effects of tepotinib on the proliferation, migration, invasion and
apoptosis of WM451 cells were partially reversed by the activation
of MET and PI3K/AKT signaling pathways.
Melanoma is the most devastating form of skin
cancer, contributing to the largest number of skin cancer-related
deaths globally which was estimated as 57,000 deaths in 2020
(21). One of the main
characteristics of cancer cells is the abnormal and uncontrolled
proliferation (22). In addition,
migration and invasion are considered to contribute to
cancer-related death (22).
Moreover, the alteration of the rate of apoptosis participates in
the advancement and progression of tumors (23). Based on this finding, it can be
concluded that the underlying mechanism of melanoma involves
proliferation, migration, invasion and apoptosis. Since tepotinib
is a highly selective oral MET inhibitor, it has shown promising
prospect in the treatment of MET-driven tumors (24). In the present study, it was found
that tepotinib suppressed the proliferation of WM451 cells in a
concentration-dependent manner. In addition, it was also revealed
that tepotinib suppressed migration and invasion and promoted the
apoptosis of WM451 cells. Furthermore, tepotinib reduced the
expression levels of N-cadherin and vimentin in WM451 cells,
indicating its suppressive effects on the EMT of melanoma
cells.
It has been shown that the expression of the HGF
receptor MET is associated with the malignant stage of melanoma
(25). In preclinical studies, the
increase in HGF/c-MET activity was shown to activate the
proliferation of melanoma cells (26), increase their invasive ability
(27), and protect them from the
induction of apoptosis (28). The
protein kinase signaling pathways are involved in promoting tumor
proliferation, migration and invasion. The abnormal activation of
the PI3K/AKT signaling is related to the occurrence and development
of a variety of cancer types and plays crucial roles in regulating
cell migration, invasion and other biological processes (29,30).
Previous studies have demonstrated that the inhibition of the
PI3K/AKT pathway contributes to the suppression of melanoma
progression (31–33). In addition, the activation of MET
can promote the proliferation, migration and invasion of tumor
cells by regulating intracellular signaling pathways, such as the
PI3K/AKT signaling pathway (34).
In the present study, it was shown that tepotinib suppressed the
activation of MET and PI3K/AKT signaling.
To further investigate the underlying mechanism of
action of MET and PI3K/AKT signaling on the proliferation,
invasion, migration and apoptosis of melanoma cells, WM451 cells
were treated with tepotinib and the activation of MET and PI3K/AKT
signaling was assessed by using HGF, which is an agonist of MET.
The present study indicated that the diminished proliferation,
migration and invasion of WM451 cells caused by tepotinib treatment
was improved by HGF, implying that the activation of MET and
PI3K/AKT signaling abolished the protective effects of tepotinib on
melanoma cells. Moreover, the increased levels of apoptosis and the
expression levels of the pro-apoptotic proteins Bax and cleaved
caspase 3 were decreased in tepotinib-treated WM451 cells following
HGF administration. These findings indicated that tepotinib
protected against malignant melanoma cell proliferation by blocking
the activation of MET and PI3K/AKT signaling pathways.
In summary, the present study indicated that
tepotinib inhibited proliferation, invasion, and migration of
melanoma cells, while inducing apoptosis of melanoma cells by
blocking the MET and the PI3K/AKT signaling pathways. However, the
side effects of tepotinib on melanoma cells were not explored in
the present study. Therefore, additional studies need to be carried
out. Additionally, the usage of only one melanoma cell line to
explore the effects of tepotinib on the malignant biological
behaviors of melanoma is another limitation of the present study;
more melanoma cell lines will be recruited in the future
experiments to support the present conclusions.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GJ and FY designed the study and analyzed the data.
HX performed the experiments. GJ and FY drafted the manuscript and
interpreted the data. FY and HX confirm the authenticity of all the
raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bai R, Huang H, Li M and Chu M: Temporal
trends in the incidence and mortality of skin malignant melanoma in
China from 1990 to 2019. J Oncol. 24:99898242021.PubMed/NCBI
|
|
2
|
Magro CM, Crowson AN and Mihm MC: Unusual
variants of malignant melanoma. Mod Pathol. 19 (Suppl 2):S41–S70.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Koh HK: Cutaneous melanoma. N Engl J Med.
325:171–182. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Miller AJ and Mihm MC Jr: Melanoma. N Engl
J Med. 355:51–65. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arrington JH III, Reed RJ, Ichinose H and
Krementz ET: Plantar lentiginous melanoma: A distinctive variant of
human cutaneous malignant melanoma. Am J Surg Pathol. 1:131–143.
1977. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Clark WH Jr, From L, Bernardino EA and
Mihm MC: The histogenesis and biologic behavior of primary human
malignant melanomas of the skin. Cancer Res. 29:705–727.
1969.PubMed/NCBI
|
|
7
|
McGovern VJ: The classification of
melanoma and its relationship with prognosis. Pathology. 2:85–98.
1970. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
de Souza CF, Morais AS and Jasiulionis MG:
Biomarkers as key contributors in treating malignant melanoma
metastases. Dermatol Res Pract. 2012:1560682012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fecher LA, Cummings SD, Keefe MJ and Alani
RM: Toward a molecular classification of melanoma. J Clin Oncol.
25:1606–1620. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Somasundaram R, Villanueva J and Herlyn M:
Intratumoral heterogeneity as a therapy resistance mechanism: Role
of melanoma subpopulations. Adv Pharmacol. 65:335–359. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Falchook GS, Kurzrock R, Amin HM, Xiong W,
Fu S, Piha-Paul SA, Janku F, Eskandari G, Catenacci DV, Klevesath
M, et al: First-in-man phase I trial of the selective MET inhibitor
tepotinib in patients with advanced solid tumors. Clin Cancer Res.
26:1237–1246. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Friese-Hamim M, Bladt F, Locatelli G,
Stammberger U and Blaukat A: The selective c-Met inhibitor
tepotinib can overcome epidermal growth factor receptor inhibitor
resistance mediated by aberrant c-Met activation in NSCLC models.
Am J Cancer Res. 7:962–972. 2017.PubMed/NCBI
|
|
13
|
Sohn SH, Sul HJ, Kim B, Kim BJ, Kim HS and
Zang DY: Tepotinib inhibits the epithelial-mesenchymal transition
and tumor growth of gastric cancers by increasing GSK3beta,
E-cadherin, and mucin 5AC and 6 levels. Int J Mol Sci. 21:60272020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bladt F, Faden B, Friese-Hamim M, Knuehl
C, Wilm C, Fittschen C, Grädler G, Meyring M, Dorsch D, Jaehrling
F, et al: EMD 1214063 and EMD 1204831 constitute a new class of
potent and highly selective c-Met inhibitors. Clin Cancer Res.
19:2941–2951. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bladt F, Friese-Hamim M, Ihling C, Wilm C
and Blaukat A: The c-met inhibitor MSC2156119J effectively inhibits
tumor growth in liver cancer models. Cancers (Basel). 6:1736–1752.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
De Silva DM, Roy A, Kato T, Cecchi F, Lee
YH, Matsumoto K and Bottaro DP: Targeting the hepatocyte growth
factor/Met pathway in cancer. Biochem Soc Trans. 45:855–870. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cao HH, Cheng CY, Su T, Fu XQ, Guo H, Li
T, Tse AKW, Kwan HY, Yu H and Yu ZL: Quercetin inhibits HGF/c-Met
signaling and HGF-stimulated melanoma cell migration and invasion.
Mol Cancer. 14:1032015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie Y, Shi X, Sheng K, Han G, Li W, Zhao
Q, Jiang B, Feng J, Li J and Gu Y: PI3K/Akt signaling transduction
pathway, erythropoiesis and glycolysis in hypoxia (Review). Mol Med
Rep. 19:783–791. 2019.PubMed/NCBI
|
|
19
|
Tan AC: Targeting the PI3K/Akt/mTOR
pathway in non-small cell lung cancer (NSCLC). Thorac Cancer.
11:511–518. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Davies MA: The role of the PI3K-AKT
pathway in melanoma. Cancer J. 18:142–147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ferlay J, Colombet M, Soerjomataram I,
Parkin DM, Piñeros M, Znaor A and Bray F: Cancer statistics for the
year 2020: An overview. Int J Cancer. Apr 5–2021.(Epub ahead of
print). View Article : Google Scholar
|
|
22
|
Lu W, Zhang H, Niu Y, Wu Y, Sun W, Li H,
Kong J, Ding K, Shen HM, Wu H, et al: Long non-coding RNA linc00673
regulated non-small cell lung cancer proliferation, migration,
invasion and epithelial mesenchymal transition by sponging
miR-150-5p. Mol Cancer. 16:1182017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'Orazi G: Apoptosis as anticancer mechanism: function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Paik PK, Felip E, Veillon R, Sakai H,
Cortot AB, Garassino MC, Mazieres J, Viteri S, Senellart H,
Meerbeeck JV, et al: Tepotinib in non-small-cell lung cancer with
met exon 14 skipping mutations. N Engl J Med. 383:931–943. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee YJ, Kim DH, Lee SH, Kim DW, Nam HS and
Cho MK: Expression of the c-Met proteins in malignant skin cancers.
Ann Dermatol. 23:33–38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Recio JA and Merlino G: Hepatocyte growth
factor/scatter factor activates proliferation in melanoma cells
through p38 MAPK, ATF-2 and cyclin D1. Oncogene. 21:1000–1008.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Otsuka T, Takayama H, Sharp R, Celli G,
LaRochelle WJ, Bottaro DP, Ellmore N, Vieira W, Owens JW, Anver M
and Merlino G: c-Met autocrine activation induces development of
malignant melanoma and acquisition of the metastatic phenotype.
Cancer Res. 58:5157–5167. 1998.PubMed/NCBI
|
|
28
|
Beuret L, Flori E, Denoyelle C, Bille K,
Busca R, Picardo M, Bertolotto C and Ballotti R: Up-regulation of
MET expression by alpha-melanocyte-stimulating hormone and MITF
allows hepatocyte growth factor to protect melanocytes and melanoma
cells from apoptosis. J Biol Chem. 282:14140–14147. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cheng JC, Chou CH, Kuo ML and Hsieh CY:
Radiation-enhanced hepatocellular carcinoma cell invasion with
MMP-9 expression through PI3K/Akt/NF-kappaB signal transduction
pathway. Oncogene. 25:7009–7018. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen WY, Xu YY and Zhang XY: Targeting
GOLM1 by microRNA-200a in melanoma suppresses cell proliferation,
invasion and migration via regulating PI3K/Akt signaling pathway
and epithelial-mesenchymal transition. Eur Rev Med Pharmacol Sci.
23:6997–7007. 2019.PubMed/NCBI
|
|
31
|
Dang N, Meng X, Ma S, Zhang Q, Sun X, Wei
J and Huang S: MDA-19 suppresses progression of melanoma via
inhibiting the PI3K/Akt pathway. Open Med (Wars). 13:416–424. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang J, Li L, Liu S, Zhao Y, Wang L and Du
G: FOXC1 promotes melanoma by activating MST1R/PI3K/AKT.
Oncotarget. 7:84375–84387. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Choi EO, Cho EJ, Jeong JW, Park C, Hong
SH, Hwang HJ, Moon SK, Son CG, Kim WJ and Choi YH: Baicalein
inhibits the migration and invasion of B16F10 mouse melanoma cells
through inactivation of the PI3K/Akt signaling pathway. Biomol Ther
(Seoul). 25:213–221. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sharma N and Adjei AA: In the clinic:
Ongoing clinical trials evaluating c-MET-inhibiting drugs. Ther Adv
Med Oncol. 3 (Suppl 1):S37–S50. 2011. View Article : Google Scholar : PubMed/NCBI
|