Introduction
Artemether, a derivative of artemisinin, is widely
used as an antimalarial drug (1–3),
including in cerebral malaria (4),
which is caused by the parasite Plasmodium falciparum
(5). Increasing evidence indicates
that artemether also has significant antineoplastic effects. A
recent study by Chen et al (6) reported that artemether alleviates the
progression of non-small cell lung cancer by inducing apoptosis and
cell cycle arrest and promoting cell senescence. Wu et al
(7) demonstrated that the heat
shock protein 90/Akt signaling pathway mediates artemether-induced
apoptosis of Cal27 cells in tongue squamous cell carcinoma. Another
study indicated that artemether inhibits proliferation and induces
apoptosis in diffuse large B-cell lymphoma (8). However, the mechanism of action of
artemether in hepatocellular carcinoma (HCC) has remained to be
fully elucidated. Therefore, the present study aimed to evaluate
the antitumor effect and mechanism of action of artemether in
HCC.
The cytochrome P450 (CYP450) cyclooxygenases are a
subgroup of heme-containing enzymes capable of epoxidation of
polyunsaturated fatty acids and the metabolism of exogenous drugs.
These enzymes therefore serve an important role in metabolism and
organ-specific toxicity. Studies have indicated that CYP family 2
subfamily J member 2 (CYP2J2) is not only expressed in the heart,
but also in the liver, kidney and lung (9–12).
The expression of human CYP enzyme in human placenta, fetal liver
and adult liver was characterized from macroscopic and microscopic
perspectives. Compared with fetal tissue, CYPs were mainly
expressed in the adult liver (9).
Bièche et al (13) analyzed
the mRNA levels of each CYP subtype in total RNA from human organ
specimens and also proved that CYP2J2 is expressed in liver tissue.
In recent years, CYP2J2 has been demonstrated to be upregulated in
a variety of tumor types and has therefore attracted increasing
attention (11,14). By analyzing CYP2J2 mRNA and protein
expression levels, researchers have indicated that, compared with
healthy human cell lines or tissues, CYP2J2 expression levels are
increased in human-derived cancer cell lines and human cancer
tissues, indicating that CYP2J2 is a positive regulator of
cancer-cell proliferation (15). A
previous study demonstrated that CYP2J2 gene knockdown is able to
inhibit the release of 11,12-epoxyeicosatrienoic acid, thereby
inhibiting the angiogenic function of M2 microglia and
significantly enhancing the antitumor effect of the cannabinoid
receptor 2 agonist JWH133 on glioma (16). Furthermore, it has been
demonstrated that 6,8-diphenylpropylene glycol induces apoptosis of
human HCC cells by activating forkhead box O3 and inhibiting CYP2J2
(10). Gui et al (17) reported that increased expression of
CYP2J2 in HCC cells upregulated the expression of
14,15-epoxyeicosatrienoic acid, which improved the stability of
hypoxia-inducible factor-1α and finally promoted the malignant
development of HCC. Therefore, CYP2J2 has the potential to be a
therapeutic target for HCC.
According to the STITCH database (http://stitch.embl.de/), artemether may target CYP2J2.
It may therefore be hypothesized that artemether is able to inhibit
the malignant process of HCC via targeting of CYP2J2. In the
present study, the effects of artemether and CYP2J2 on the
proliferation, invasion and migration of HCC cells and the
underlying mechanisms were investigated, in the hope that the
results will provide a basis for the understanding of the antitumor
effects of artemether and indicate a novel target and strategy for
the clinical treatment of patients with HCC.
Materials and methods
Cell culture
The human HCC cell line Hep3B2.1-7 and the human
embryonic hepatocyte cell line HHL-5 were obtained from The Cell
Bank of Type Culture Collection of The Chinese Academy of Sciences.
The cells were cultured in DMEM (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin (Invitrogen;
Thermo Fisher Scientific, Inc.) at 37°C in a humidified atmosphere
with 5% CO2.
Bioinformatics analysis
The STITCH database (version 5.0; http://stitch.embl.de/) is a database that may be used
to predict interactions between artemether and CYP2J2 (confidence,
0.400). In the STITCH database, each protein-protein interaction is
annotated with one or more ‘scores’. Of note, these scores do not
indicate the strength or the specificity of the interaction.
Instead, they are indicators of confidence, i.e. how likely STITCH
judges an interaction to be true, given the available evidence. The
minimum required interaction score is divided into the following
categories: Highest confidence (0.900); high confidence (0.700);
intermediate confidence (0.400) and low confidence (0.150). The
ENCORI website (https://starbase.sysu.edu.cn/panGeneDiffExp.php) is
used to predict CYP2J2 expression in HCC and noncancerous liver
cells. The expression data of cancers were downloaded from The
Cancer Genome Atlas project via the Genomic Data Commons Data
Portal (https://gdc.cancer.gov). The expression
values of genes from RNA-sequencing data were scaled as log2(FPKM +
0.01).
Cell Counting Kit-8 (CCK-8) assay
The CCK-8 assay was performed to assess cell
viability. In brief, cells were inoculated in 96-well plates at a
density of 8×103 cells/well, with different
concentrations (20, 40, 80 and 160 µM) of artemether (Chemical
Abstracts Service no. 71963-77-4; Toronto Research Chemicals) and
incubated at 37°C for 24, 48 and 72 h, control without artemether.
Following incubation, 10 µl CCK-8 reagent was added into each well
and cells were cultured for another 2 h at 37°C. The absorbance in
each well was measured at a wavelength of 450 nm using a microplate
reader (BioTek Instruments, Inc.).
Colony-formation assay
Hep3B2.1-7 cells (1×103 cells/well) were
suspended in DMEM supplemented with 10% FBS and were seeded into
6-well plates and cultured in a 5% CO2 incubator at 37°C
for 14 days with different concentrations (20, 40 and 80 µM) of
artemether, while cells without artemether were used as a control.
Subsequently, the cells were fixed using 4% paraformaldehyde at
room temperature for 20 min and stained with 0.05% crystal violet
solution for 25 min at 37°C. Finally, the colonies (>50
cells/colony) were counted using an Olympus BX40 light microscope
(Olympus Corporation).
Wound-healing assay
Hep3B2.1-7 cells were inoculated in 6-well plates
and incubated in a 5% CO2 incubator at 37°C until
reaching 70–80% confluency. A 200-µl pipette tip was used to make
linear scratches on the cell monolayer. PBS was used to wash the
monolayer three times for 2 min each to remove any cell debris.
Subsequently, cells were cultured with different concentrations
(20, 40 and 80 µM) of artemether in an incubator containing 5%
CO2 at 37°C and were imaged at 0 and 24 h using an EVOS™
M7000 imaging system (Thermo Fisher Scientific, Inc; magnification,
×100). ImageJ software (version 1.8.0; National Institutes of
Health) was used to quantify the area occupied by cells migrating
into the linear scratches.
Transwell assay
The invasion ability of the cells was assessed using
the Transwell assay. In brief, Hep3B2.1-7 cells (density,
5×104 cells) were suspended in plasma-free DMEM. The
upper chamber was precoated with Matrigel (MilliporeSigma) and was
subsequently inoculated with Hep3B2.1-7 cells (0.1 ml cell
suspension/well). The lower compartment was filled with DMEM
containing 20% FBS. After 24 h of incubation, the upper chamber was
collected and cleaned and the cells were stained with 0.3% crystal
violet (MilliporeSigma) at room temperature for 20 min. Images of
cell invasion were observed under an EVOS™ M7000 imaging system
(Thermo Fisher Scientific, Inc; magnification, ×100).
Western blot analysis
Cells from each group were collected and the total
protein was extracted using RIPA lysis buffer (Beijing Solarbio
Science & Technology Co., Ltd.). Protein concentrations were
determined using the BCA Protein Detection Kit (Beyotime Institute
of Biotechnology) according to the manufacturer's protocol
(18). The amount of total protein
loaded for each group was 40 µg, which was separated by 10%
SDS-PAGE and transferred to PVDF membranes (MilliporeSigma).
Membranes were blocked in 5% non-fat milk (Phygene Scientific) at
room temperature for 4 h. After washing for 3 times with 1X
Tris-buffered saline for 5 min each, the membranes were incubated
with the following primary antibodies (all purchased from Abcam)
were overnight at 4°C: Anti-Bcl-2 (1:1,000 dilution; cat. no.
Ab32124), anti-Bax (1:1,000 dilution; cat. no. Ab32503),
anti-E-cadherin (1:10,000 dilution; cat. no. Ab40772),
anti-N-cadherin (1:5,000 dilution; cat. no. Ab76011), anti-vimentin
(1:1,000 dilution; cat. no. Ab92547), anti-CYP2J2 (1:1,000
dilution; cat. no. Ab151996) and anti-GAPDH (1:1,000 dilution; cat.
no. Ab8245). Following the primary incubation, the membranes were
incubated with HRP-conjugated secondary antibodies (1:5,000
dilution; Santa Cruz Biotechnology, Inc.) at room temperature for 2
h. Protein bands were visualized using ECL solution (Absin) and
were imaged using a gel imager (C150; Azure Biosystems, Inc.). The
gray value of the protein bands was analyzed using ImageJ (version
1.51; National Institutes of Health) with GAPDH as the loading
control.
Terminal deoxynucleotidyl transferase
deoxyuridine triphosphate nick-end labeling (TUNEL) assay
The effects of artemether on apoptosis of Hep3B2.1-7
cells were detected by TUNEL staining in accordance with the
manufacturer's protocol. In brief, cells (1×105
cells/well) were collected and washed three times with PBS for 2
min each. Following fixing with 4% paraformaldehyde at room
temperature for 5 min, the cells were gently washed with PBS twice
for 2 min each time. A DAPI staining solution (cat. no. C1005;
Beyotime Institute of Biotechnology) was added to just cover the
cells and they were incubated at room temperature for 3–5 min.
Following washing with PBS 2–3 times for 3–5 min each time, the
cells were incubated with 0.3% Triton-X-100 at room temperature for
5 min. Subsequently, 50 µl TUNEL assay solution (cat. no. C1086;
Beyotime Institute of Biotechnology) was added to the cells,
followed by incubation at 37°C in the dark for 60 min. The
detection solution was discarded and cells were washed three times
with PBS. Subsequently, three fields of view were selected at
random and cells were sealed with Antifade Mounting Medium
(Beyotime Institute of Biotechnology) for observation under a
fluorescence microscope (magnification, ×200; Zeiss GmbH).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from cells using
TRIzol® reagent according to the manufacturer's
protocol. The RNA concentration and quality were assessed using a
NanoDrop spectrophotometer (Thermo Fisher Scientific, Inc.).
Following addition of 10 U/µl DNase I digestion solution (Roche)
for 10 min at 37°C, total RNA was reverse-transcribed into
complementary DNA (cDNA) using a cDNA Synthesis Kit (Invitrogen;
Thermo Fisher Scientific, Inc.). qPCR was performed using SYBR
Premix Ex Taq reagents (Takara Bio, Inc.) with an ABI 7500 qPCR
instrument (Applied Biosystems; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. The following
thermocycling conditions were used for qPCR: 95°C for 10 min;
followed by 40 cycles of 95°C for 10 sec and 60°C for 60 sec. The
following primers (GenScript) were used for qPCR: CYP2J2 forward,
5′-GCCACCCCTGACACATTCAA-3′ and reverse, 5′-GGCATGCCCGCTTTCCTATT-3′;
and GAPDH forward, 5′-AGCCACATCGCTCAGACAC-3′ and reverse,
5′-GCCCAATACGACCAAATCC-3′. GAPDH served as the internal reference
gene and relative gene expression was determined using the
2−ΔΔCq method (19).
Cell transfection
The CYP2J2-overexpression vector [overexpressed
(Ov)-CYP2J2] and the corresponding negative control (NC; Ov-NC)
were synthesized by Shanghai GeneChem Co., Ltd. The cells were
inoculated in 12-well plates at a density of 3×105
cells/well and cultured in a 5% CO2 incubator at 37°C
for 24 h. Following incubation, cells were transfected with the
aforementioned plasmids at a concentration of 20 nM using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Cells
in the blank control group (Control) remained untreated. Following
transfection for 48 h, the protein expression levels were evaluated
via RT-qPCR.
Statistical analysis
All data were analyzed using GraphPad Prism 7
software (GraphPad Software, Inc.). Values are expressed as the
mean ± standard deviation from ≥3 independent experimental repeats.
Significant differences between two groups were determined using an
unpaired Student's t-test, whereas those among more than two groups
were assessed using one-way ANOVA followed by Tukey's post-hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
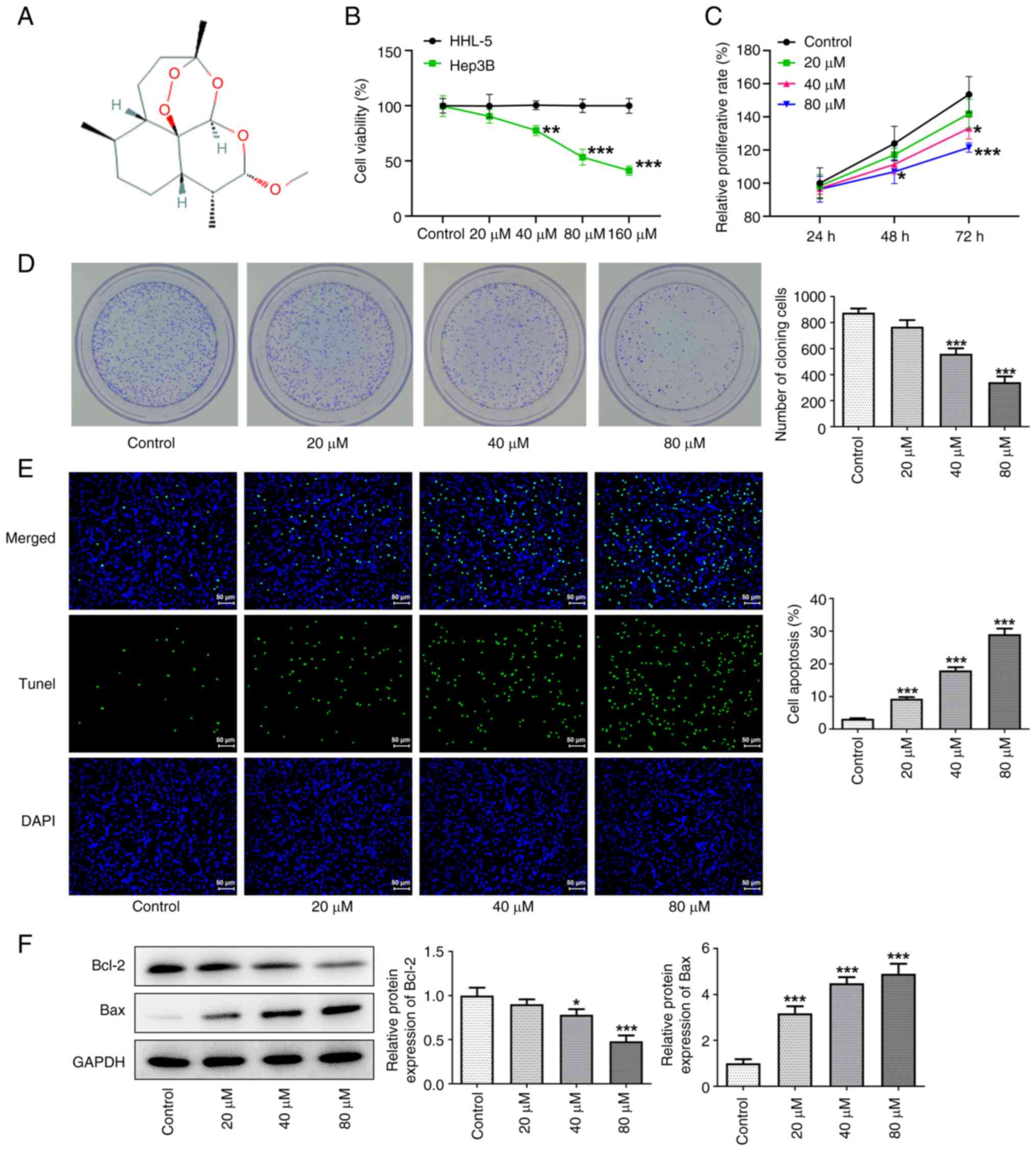
Artemether inhibits HCC cell
proliferation
The effects of different concentrations of
artemether on cell viability were detected using a CCK-8 assay
(Fig. 1A). The results indicated
that, following incubation with cells for 24, 48 and 72 h,
artemether significantly inhibited the proliferation of HCC cells
in a dose- and time-dependent manner (Fig. 1B and C). When the concentration of
artemether was 160 µM, the viability of Hep3B2.1-7 cells decreased
to 40%. Furthermore, at the same concentration, artemether exerted
no significant inhibitory effect on the proliferation of HHL-5
cells, which indicated that artemether may not be harmful to
healthy cells while inhibiting the proliferation of HCC cells. In
addition, as presented in Fig. 1D,
the colony formation assay further confirmed that artemether had a
significant inhibitory effect on HCC cell proliferation. TUNEL
staining indicated that artemether promoted apoptosis of HCC cells
in a concentration-dependent manner (Fig. 1E). Subsequently, the expression
levels of apoptosis-related proteins (Bcl-2 and Bax) were detected
via western blot analysis (Fig.
1F). The results also indicated that artemether obviously
promoted apoptosis of HCC cells.
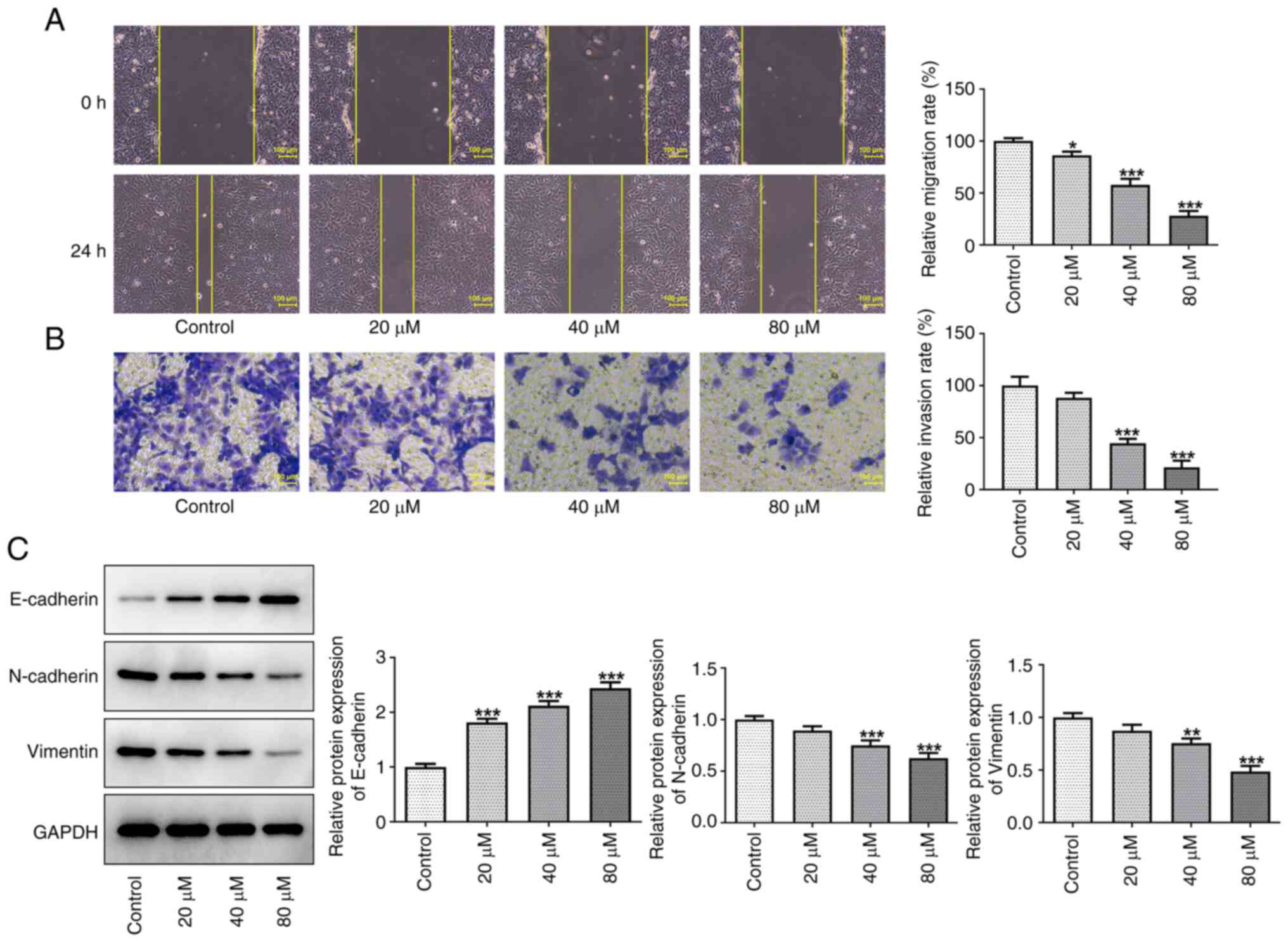
Artemether inhibits HCC cell invasion
and migration
Wound-healing and Transwell assays were performed to
assess the effects of artemether on the invasion and migration of
Hep3B2.1-7 cells after 24 h of treatment. Compared with those in
the control group, the cell migration and invasion ability were
decreased by artemether in a dose-dependent manner (Fig. 2A and B). Furthermore, as
epithelial-mesenchymal transition (EMT) is an important internal
mechanism of tumor invasion and metastasis (20), the expression levels of the
EMT-related proteins E-cadherin, N-cadherin and vimentin were
detected by western blot analysis. The results presented in
Fig. 2C demonstrated that,
compared with the control group, E-cadherin protein expression
levels increased, whereas the protein expression levels of
N-cadherin and vimentin decreased in a dose-dependent manner with
increasing artemether concentrations. These results therefore
indicated that artemether may effectively inhibit the migration and
invasion of HCC cells to surrounding and distant tissues.
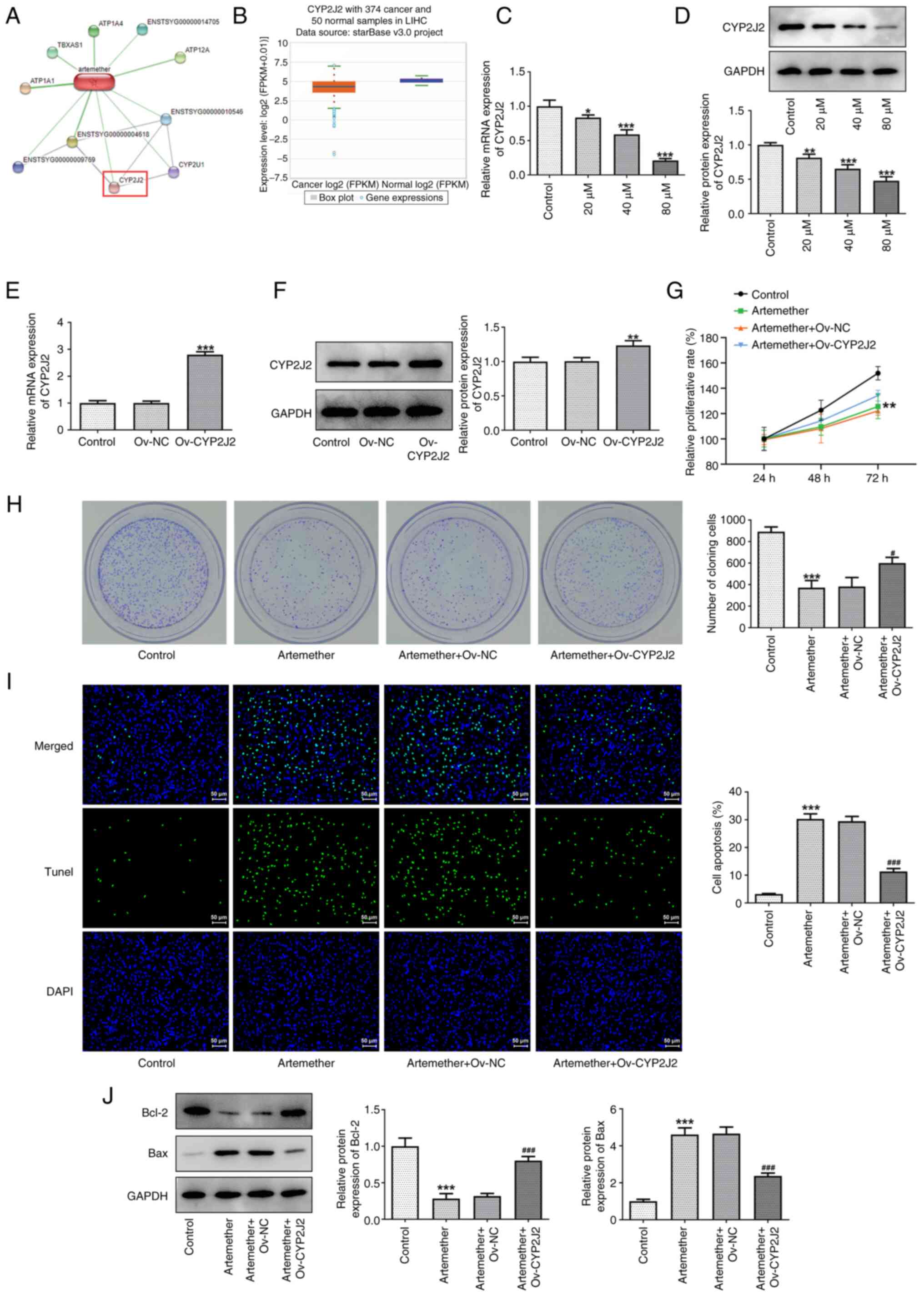
CYP2J2 overexpression reverses the
inhibitory effect of artemether on HCC cell proliferation
It was determined that artemether may target CYP2J2
based on a prediction made with the STITCH database (Fig. 3A). In addition, the ENCORI website
(https://starbase.sysu.edu.cn/) predicted
no abnormal expression of CYP2J2 in HCC (Fig. 3B). Subsequently, CYP2J2 mRNA and
protein expression levels in Hep3B2.1-7 cells treated with
artemether were detected using RT-qPCR and western blot analysis,
respectively. As presented in Fig. 3C
and D, the CYP2J2 mRNA and protein expression levels decreased
in a dose-dependent manner with increasing artemether
concentrations. When the concentration of artemether was 80 µM,
CYP2J2 mRNA and protein expression levels were at their lowest,
indicating that artemether effectively inhibited CYP2J2 expression.
Consequently, 80 µM artemether was selected for subsequent
experiments. The transfection efficiency of the CYP2J2
overexpression plasmid in Hep3B2.1-7 cells was also detected using
RT-qPCR and western blot analysis (Fig. 3E and F). The results demonstrated
that the CYP2J2 mRNA and protein expression levels in the Ov-CYP2J2
group were higher compared with those in the Ov-NC group.
Furthermore, CCK-8 (Fig. 3G) and
colony formation assays (Fig. 3H)
were applied to examine Hep3B2.1-7 cell proliferation. The results
demonstrated that artemether significantly inhibited the CYP2J2
expression levels and that Ov-CYP2J2 reversed the inhibitory effect
of artemether on the proliferation of HCC cells. In addition,
Ov-CYP2J2 also reversed the apoptotic effect of artemether on HCC
cells (Fig. 3I and J).
CYP2J2 overexpression reverses the
inhibitory effect of artemether on HCC cell invasion and
migration
Whether artemether affected the invasion and
migration of HCC cells by targeting CYP2J2 was subsequently
investigated. The results of the wound-healing and Transwell assays
demonstrated that CYP2J2 overexpression significantly reversed the
inhibitory effect of artemether on Hep3B2.1-7 cell invasion and
migration (Fig. 4A and B).
Furthermore, the elevated EMT marker E-cadherin, and the reduced
N-cadherin and vimentin levels indicated that CYP2J2 overexpression
counteracted the suppressive impact of artemether on the EMT
process in Hep3B2.1-7 cells (Fig.
4C). These results collectively suggested that artemether may
inhibit the proliferation, invasion and migration of HCC cells by
targeting CYP2J2 expression.
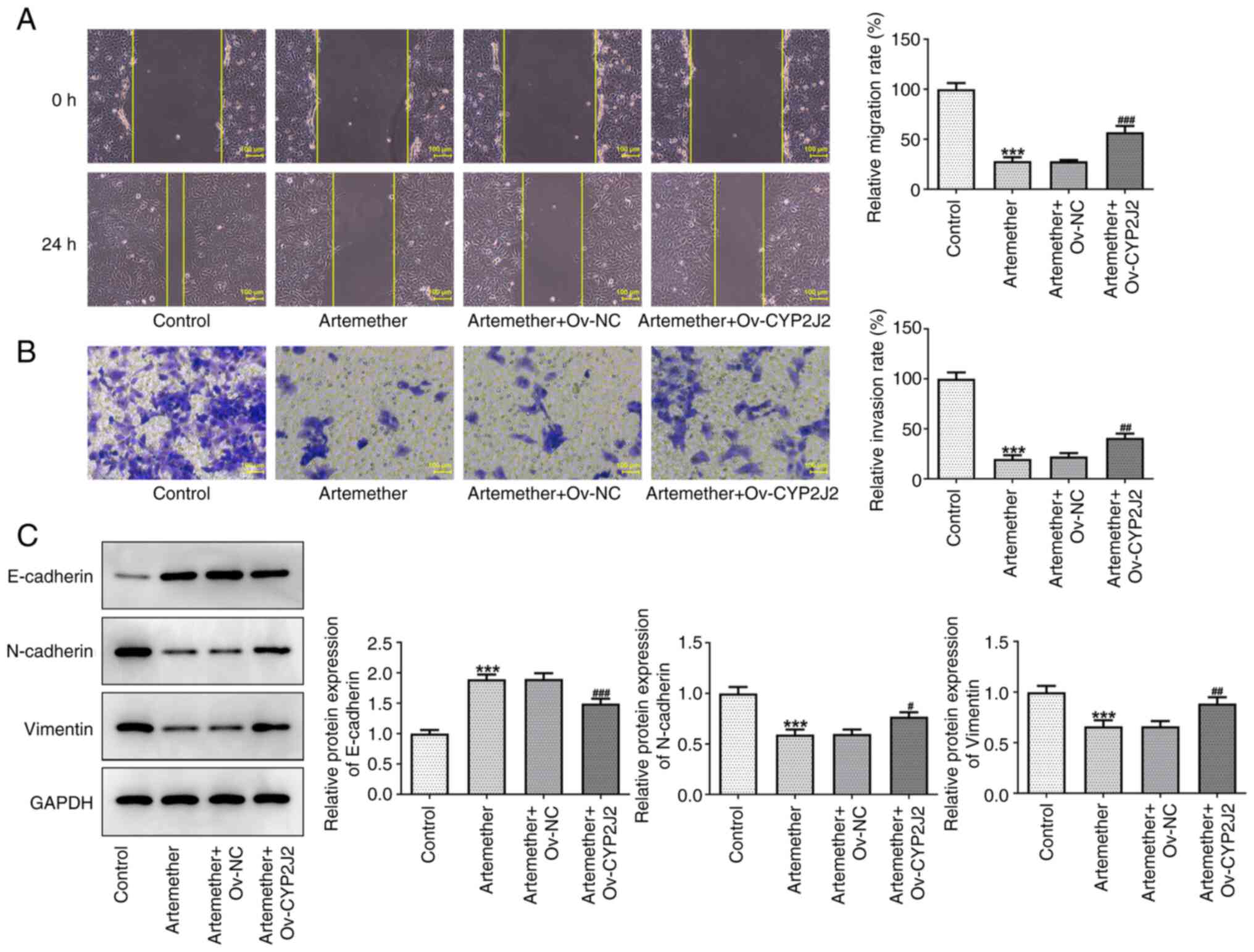 | Figure 4.Overexpression of CYP2J2 reverses the
inhibitory effect of artemether on the invasion and migration of
hepatocellular carcinoma cells. (A) Representative images of the
wound-healing assay (magnification, ×100; scale bar, 100 µm) and
quantified results. (B) Representative images of the Transwell
assay (magnification, ×200; scale bar, 100 µm) and quantified
results. (C) E-cadherin, N-cadherin and vimentin protein expression
levels were assessed by western blot analysis. ***P<0.001 vs.
Control; #P<0.05, ##P<0.01 and
###P<0.001 vs. Artemether + Ov-NC. NC, negative
control; Ov, overexpression vector; CYP2J2, cytochrome P450 family
2 subfamily J member 2; HCC, hepatocellular carcinoma. |
Discussion
HCC is associated with chronic liver disease and is
one of the most common and fatal cancer types worldwide (21). According to Global Cancer
Statistics in 2018, HCC ranked seventh in incidence among other
malignant tumors and is still the fifty-second highest cause of
cancer-associated mortality worldwide (22). The mean age at diagnosis for HCC is
between 50 and 60 years. Early HCC is usually treated with surgical
resection, liver transplantation, ablation or radiotherapy.
However, the 5-year overall survival rate following diagnosis is
only 50–70% (23). Despite decades
of research on cytotoxicity, targeted drugs and immunotherapy, only
a limited number of effective treatment options are available for
advanced HCC (24). Therefore, the
development of novel therapeutics is of great significance for
improving the prognosis of patients with HCC. In the present study,
the results demonstrated that artemether inhibited the
proliferation and induced apoptosis of Hep3B2.1-7 cells in a
dose-dependent manner, which may be related to the ability of
artemether to induce cell cycle arrest/growth inhibition (25). This is consistent with the results
of the study by Hou et al (25), which examined the effect of
artemisinin on the cell cycle of liver cancer cells, indicating
that after artemisinin treatment of HepG2 cells, the proportion of
cells in G1 phase was significantly increased, with higher
concentrations inducing more significant G1-phase arrest.
Artemisinin and its derivatives have long been
recognized as the most effective anti-malarial drugs worldwide
(2,26,27).
With the further development of artemisinin and its derivatives,
studies have reported that they possess good antitumor activity in
the treatment of human cancer (28–30).
Artemether, as a natural derivative of artemisinin, may also be
more effective than artemisinin (31). It was demonstrated that when using
artemether and triglyceride docosahexaenoate as a lipid core,
nanoemulsion, nanostructure lipid carrier and poly (lactic
acid)-poly (ethylene glycol), nanocapsules were successfully
prepared to reduce the activity, proliferation and migration of
MDA-MB-231 and MCF-7 breast cancer cells in a dose-dependent manner
(32). Furthermore, artemether was
able to regulate the sensitivity of B7-H3 human neuroblastoma cells
to adriamycin (33). Interference
with vascular cell adhesion protein-1 in combination with short
hairpin-RNA significantly inhibited the malignant behavior of human
glioma cells (34). These
properties make artemether an attractive potential candidate for
chemotherapy. However, to the best of our knowledge, the efficacy
of artemether in HCC has remained elusive. In the present study,
artemether significantly reduced the activity of Hep3B2.1-7 cells
and promoted apoptosis in a dose-dependent manner. These results
verified the antitumor effect of artemether in HCC.
To further explore the mechanism of artemether in
HCC, the STITCH database was used to predict that artemether is
able to target CYP2J2. As previously mentioned, CYP2J2 is a
cyclooxygenase that is able to metabolize numerous unsaturated
fatty acids and serves a variety of biological roles in the
cardiovascular system and a number of solid human cancer types
(12,35,36).
For instance, Park et al (37) demonstrated that CYPs inhibit the
proliferation of human HCC cells and lead to apoptosis via
non-competitive inhibition of the CYP2J2 enzyme. Furthermore,
Allison et al (38)
demonstrated that paclitaxel, a chemotherapeutic drug, induces
apoptosis in breast cancer cells via mediating lipid peroxidation
and generating reactive oxygen species, but CYP2J2 overexpression
was able to activate aldehyde dehydrogenase 1 family member A1,
which reversed the inhibitory effect of paclitaxel on tumor growth.
Another study also reported that acetylshikonin exerted inhibitory
effects against CYP2J2 and anti-cancer activity in human liver
cancer HepG2 cells (37). In the
present study, it was confirmed that artemether inhibited the
expression of CYP2J2 in a concentration-dependent manner and
further overexpression of CYP2J2 revealed that Ov-CYP2J2 partially
reversed the inhibitory effects of artemether on the proliferation,
migration and invasion of HCC cells.
The present study had several limitations. First,
the mechanism of the effect of artemether on Hep3B2.1-7 cells was
assessed by overexpression of CYP2J2. However, other methods, such
as CYP2J2 antagonists or inhibitors, may be useful to confirm the
present observations. Furthermore, in vitro studies using
only Hep3B2.1-7 cells were performed in the present study and other
HCC cell lines or animal experiments may provide additional
information to complement the current findings. In addition,
further evaluation of the effect of artemether on the cell cycle of
HCC cells and verification of the upregulation of CYP2J2 expression
in HCC compared with normal hepatocytes will be endeavored in a
future study.
In conclusion, artemether inhibited the activity of
Hep3B2.1-7 cells in a dose-dependent manner. The results of the
present study suggested that artemether suppressed the
proliferation and activity of HCC by targeting CYP2J2. Therefore,
the development of effective CYP2J2 inhibitors may be of potential
therapeutic value. In future work, the specific anticancer
mechanism of artemether will continue to be investigated and in
vivo experiments in mice will also be performed.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XZ and MY conceptualized and designed the current
study. GY, ZS and QS acquired, analyzed and interpreted data. QS,
XZ and MY drafted the manuscript and revised it critically for
important intellectual content. All authors agreed to be held
accountable for the current study in ensuring questions related to
the integrity of any part of the work are appropriately
investigated and resolved. XZ and QS confirm the authenticity of
the raw data. All authors read approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Esu EB, Effa EE, Opie ON and Meremikwu MM:
Artemether for severe malaria. Cochrane Database Syst Rev.
6:CD0106782019.PubMed/NCBI
|
|
2
|
Visser BJ, Bierhoff M, van Gool T, van
Hattem JM, Grobusch MP and van Vugt M: The treatment of malaria.
Ned Tijdschr Geneeskd. 163:D33212019.(In Dutch).
|
|
3
|
Cheong DHJ, Tan DWS, Wong FWS and Tran T:
Anti-malarial drug, artemisinin and its derivatives for the
treatment of respiratory diseases. Pharmacol Res. 158:1049012020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Prabhu P, Suryavanshi S, Pathak S, Patra
A, Sharma S and Patravale V: Nanostructured lipid carriers of
artemether-lumefantrine combination for intravenous therapy of
cerebral malaria. Int J Pharm. 513:504–517. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ouji M, Barnoin G, Fernandez Alvarez A,
Augereau JM, Hemmert C, Benoit-Vical F and Gornitzka H: Hybrid
Gold(I) NHC-artemether complexes to target falciparum malaria
parasites. Molecules. 25:28172020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen J, Huang X, Tao C, Xiao T, Li X, Zeng
Q, Ma M and Wu Z: Artemether attenuates the progression of
non-small cell lung cancer by inducing apoptosis, cell cycle arrest
and promoting cellular senescence. Biol Pharm Bull. 42:1720–1725.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu J, Li L, Wang Y, Ren X, Lin K and He Y:
The HSP90/Akt pathway may mediate artemether-induced apoptosis of
Cal27 cells. FEBS Open Bio. 9:1726–1733. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao X, Guo X, Yue W, Wang J, Yang J and
Chen J: Artemether suppresses cell proliferation and induces
apoptosis in diffuse large B cell lymphoma cells. Exp Ther Med.
14:4083–4090. 2017.PubMed/NCBI
|
|
9
|
Robinson JF, Hamilton EG, Lam J, Chen H
and Woodruff TJ: Differences in cytochrome p450 enzyme expression
and activity in fetal and adult tissues. Placenta. 100:35–44. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee CM, Lee J, Jang SN, Shon JC, Wu Z,
Park K, Liu KH and Park SH: 6,8-diprenylorobol induces apoptosis in
human hepatocellular carcinoma cells via activation of FOXO3 and
inhibition of CYP2J2. Oxid Med Cell Longev. 2020:88872512020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zou X and Mo Z: CYP2J2 is a diagnostic and
prognostic biomarker associated with immune infiltration in kidney
renal clear cell carcinoma. Biomed Res Int. 2021:37718662021.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Evangelista EA, Aliwarga T, Sotoodehnia N,
Jensen PN, McKnight B, Lemaitre RN, Totah RA and Gharib SA: CYP2J2
modulates diverse transcriptional programs in adult human
cardiomyocytes. Sci Rep. 10:53292020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bièche I, Narjoz C, Asselah T, Vacher S,
Marcellin P, Lidereau R, Beaune P and de Waziers I: Reverse
transcriptase-PCR quantification of mRNA levels from cytochrome
(CYP)1, CYP2 and CYP3 families in 22 different human tissues.
Pharmacogenet Genomics. 17:731–742. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tao P, Jiang Y, Wang H and Gao G:
CYP2J2-produced epoxyeicosatrienoic acids contribute to the
ferroptosis resistance of pancreatic ductal adenocarcinoma in a
PPARү-dependent manner. Zhong Nan Da Xue Xue Bao Yi Xue Ban.
46:932–941. 2021.(In Chinese). PubMed/NCBI
|
|
15
|
Jiang JG, Chen CL, Card JW, Yang S, Chen
JX, Fu XN, Ning YG, Xiao X, Zeldin DC and Wang DW: Cytochrome P450
2J2 promotes the neoplastic phenotype of carcinoma cells and is
up-regulated in human tumors. Cancer Res. 65:4707–4715. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lei X, Chen X, Quan Y, Tao Y and Li J:
Targeting CYP2J2 to enhance the anti-glioma efficacy of cannabinoid
receptor 2 stimulation by inhibiting the pro-angiogenesis function
of M2 microglia. Front Oncol. 10:5742772020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gui L, Xu Q, Huang J, Wu G, Tang H, Hui L,
Hua P, Zhang L and Zhu Y: CYP2J2 promotes the development of
hepatocellular carcinoma by increasing the EETs production to
improve HIF-1α stability. Am J Transl Res. 12:7923–7937.
2020.PubMed/NCBI
|
|
18
|
Khramtsov P, Kalashnikova T, Bochkova M,
Kropaneva M, Timganova V, Zamorina S and Rayev M: Measuring the
concentration of protein nanoparticles synthesized by desolvation
method: Comparison of Bradford assay, BCA assay, hydrolysis/UV
spectroscopy and gravimetric analysis. Int J Pharm. 599:1204222021.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hirschfeld M, Ge I, Rucker G, Waldschmidt
J, Mayer S, Jager M, Voigt M, Kammerer B, Nöthling C, Berner K, et
al: Mutually distinguishing microRNA signatures of breast, ovarian
and endometrial cancers in vitro. Mol Med Rep. 22:4048–4060.
2020.PubMed/NCBI
|
|
20
|
Zhang Y and Weinberg RA:
Epithelial-to-mesenchymal transition in cancer: Complexity and
opportunities. Front Med. 12:361–373. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hartke J, Johnson M and Ghabril M: The
diagnosis and treatment of hepatocellular carcinoma. Semin Diagn
Pathol. 34:153–159. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
European Association for the Study of the
Liver. Electronic address, . simpleEasloffice@easloffice.eu;
European Association for the Study of the Liver: ‘EASL Clinical
Practice Guidelines: Management of hepatocellular carcinoma’. J
Hepatol. 69:182–236. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
An Y, Jiang J, Zhou L, Shi J, Jin P, Li L,
Peng L, He S, Zhang W, Huang H, et al: Peroxiredoxin 1 is essential
for natamycin-triggered apoptosis and protective autophagy in
hepatocellular carcinoma. Cancer Lett. 521:210–223. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hou J, Wang D, Zhang R and Wang H:
Experimental therapy of hepatoma with artemisinin and its
derivatives: In vitro and in vivo activity, chemosensitization, and
mechanisms of action. Clin Cancer Res. 14:5519–5530. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Talman AM, Clain J, Duval R, Menard R and
Ariey F: Artemisinin bioactivity and resistance in malaria
parasites. Trends Parasitol. 35:953–963. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Menard D and Dondorp A: Antimalarial drug
resistance: A threat to malaria elimination. Cold Spring Harb
Perspect Med. 7:a0256192017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao Y, Feng YH, Gao LW, Li XY, Jin QX,
Wang YY, Xu YY, Jin F, Lu SL and Wei MJ: Artemisinin enhances the
anti-tumor immune response in 4T1 breast cancer cells in vitro and
in vivo. Int Immunopharmacol. 70:110–116. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang F, Zhou JY, Zhang D, Liu MH and Chen
YG: Artesunate induces apoptosis and autophagy in HCT116 colon
cancer cells, and autophagy inhibition enhances the
artesunate-induced apoptosis. Int J Mol Med. 42:1295–1304.
2018.PubMed/NCBI
|
|
30
|
Zhao F, Vakhrusheva O, Markowitsch SD,
Slade KS, Tsaur I, Cinatl J Jr, Michaelis M, Efferth T, Haferkamp A
and Juengel E: Artesunate impairs growth in cisplatin-resistant
bladder cancer cells by cell cycle arrest, apoptosis and autophagy
induction. Cells. 9:26432020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang Y, Xu G, Zhang S, Wang D, Saravana
Prabha P and Zuo Z: Antitumor research on artemisinin and its
bioactive derivatives. Nat Prod Bioprospect. 8:303–319. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lanna EG, Siqueira RP, Machado MGC, de
Souza A, Trindade IC, Branquinho RT and Mosqueira VCF: Lipid-based
nanocarriers co-loaded with artemether and triglycerides of
docosahexaenoic acid: Effects on human breast cancer cells. Biomed
Pharmacother. 134:1111142021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tan WQ, Chen G, Ye M and Jia B: Artemether
regulates chemosensitivity to doxorubicin via regulation of B7-H3
in human neuroblastoma cells. Med Sci Monit. 23:4252–4259. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang YB, Hu Y, Li Z, Wang P, Xue YX, Yao
YL, Yu B and Liu YH: Artemether combined with shRNA interference of
vascular cell adhesion molecule-1 significantly inhibited the
malignant biological behavior of human glioma cells. PLoS One.
8:e608342013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Aliwarga T, Evangelista EA, Sotoodehnia N,
Lemaitre RN and Totah RA: Regulation of CYP2J2 and EET levels in
cardiac disease and diabetes. Int J Mol Sci. 19:19162018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Das A, Weigle AT, Arnold WR, Kim JS,
Carnevale LN and Huff HC: CYP2J2 molecular recognition: A new axis
for therapeutic design. Pharmacol Ther. 215:1076012020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Park SH, Phuc NM, Lee J, Wu Z, Kim J, Kim
H, Kim ND, Lee T, Song KS and Liu KH: Identification of
acetylshikonin as the novel CYP2J2 inhibitor with anti-cancer
activity in HepG2 cells. Phytomedicine. 24:134–140. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Allison SE, Chen Y, Petrovic N, Zhang J,
Bourget K, Mackenzie PI and Murray M: Activation of ALDH1A1 in
MDA-MB-468 breast cancer cells that over-express CYP2J2 protects
against paclitaxel-dependent cell death mediated by reactive oxygen
species. Biochem Pharmacol. 143:79–89. 2017. View Article : Google Scholar : PubMed/NCBI
|