Introduction
Interferons (IFNs) are produced by the innate immune
system via Toll-like receptor (TLR) stimulation and other signaling
cascades (1). According to the
primary structure of IFNs and their impact on three dimeric target
receptors, IFNs can be classified into several types and families.
There are three main classes of IFNs in humans: IFN-α, -β and -γ.
Among them, IFN-α and -β belong to the type I IFNs. IFN-α is
primarily secreted by monocytes/macrophages and can also be
synthesized by B cells and fibroblasts, whereas IFN-β is mainly
produced by fibroblasts. IFN-α and -β bind to the same receptor and
are widely distributed, including on monocytes/macrophages, B
cells, T cells, platelets, epithelial cells, endothelial cells and
cancer cells. Human IFN-α subtypes share ~50% sequence identity and
IFN-α2 is ~20% identical to IFN-β. IFN-α and -β have 186–190 amino
acids (aa) and have a cleavable signal peptide, which forms a
secreted protein of 165 or 166 aa (2,3).
IFN-α not only serves a vital role in modulating the
immune system and inducing antiviral innate immune responses, but
it also serves an important role in antitumor therapy (4–6).
Numerous mechanisms have been proposed for the anticancer effect of
IFN-α, including the induction of cell apoptosis, which is
initiated via the extrinsic signaling pathway, the intrinsic
mitochondrial signaling pathway or the stress kinase signaling
pathway (7). Due to its antitumor
properties, IFN-α has been widely used for the clinical treatment
of malignancies, such as renal cell cancer (RCC), hepatocellular
carcinoma (HCC), malignant melanoma and cervical cancer (8–10).
For a long time, most of the reviews on type I IFNs mainly focused
on IFN-β, and less attention was paid to IFN-α (11–15).
Therefore, in the present review, a brief overview of the
proapoptotic effects of IFN-α in various cancers will be provided
and the existing literature on the signaling pathways and molecular
mechanisms of IFN-α-induced cancer cell apoptosis will be explored
so as to supplement and improve IFN-α-related reviews.
Biological characteristics and subtypes of
IFN-α
IFN-α exhibits a wide variety of direct and/or
indirect biological properties, including antiproliferative and
antiviral properties, stimulating the cytotoxic activity of
different host-immune cells, inducing proapoptotic genes/proteins,
upregulating major histocompatibility complex class I antigens and
tumor-associated surface antigens, suppressing antiapoptotic genes,
inhibiting angiogenesis and modulating cell differentiation
(9,10,16–19).
It can therefore be hypothesized that IFN-α is an important agent
for treating various infectious diseases.
In total, there are 13 IFN-α subtypes expressed from
14 human IFN-α genes. The IFN-α subtype generated from the IFN-α13
gene is identical to that generated from the IFN-α1 gene.
Therefore, there are 12 different IFN-α subtypes in humans
(20–22). All IFN-α subtypes have a high
structural similarity, including the length of the protein and the
absence of introns. Out of the 12 IFN-α subtypes 11 are 166 aa in
length with a molecular weight of ~20 kDa (IFN-α2 is 165 aa due to
the deletion of D44). Their protein sequence is highly conserved
and the identity score among the IFN-α subtypes ranges from 76–96%
(23,24). Each IFN-α subtype exhibits
different activities, which include antiproliferative and antiviral
activities, as well as promoting the cytotoxic activities of T
cells and natural killer (NK) cells (25). For example, most subtypes of IFN-α
(IFN-αA, B, C, D, F, I and K) are capable of boosting NK cells.
However, IFN-αJ exhibits virtually no NK cell activity but has
potent antiviral and antiproliferative activities, which suggests
that it has an antagonist effect on NK cell activity via inhibiting
other IFN-α subtypes to stimulate NK cells.
IFN-α signaling and its regulation
Similar to other type I IFNs, IFN-α exerts
biological effects by binding to a specific receptor known as the
IFN-α/β receptor subunit (IFNAR)1/IFNAR2 heterodimer on the surface
of target cells (23,26). Upon binding, the downstream
molecules Janus kinase (JAK)1 and tyrosine kinase 2 are activated,
which results in the recruitment of STAT1 and STAT2 to the
cytoplasmic tail of the receptor and therefore forms STAT1/STAT2
heterodimers that can translocate into the nucleus. Subsequently,
the heterotrimeric transcriptional complex IFN-stimulated gene
(ISG) factor 3 (ISGF3) is formed by STAT1/STAT2 heterodimers
combining with IFN-regulatory factor (IRF)9. Upon binding to
specific DNA response elements, ISGs can be transactivated by ISGF3
(Fig. 1). IFN-α also stabilizes
other STAT homodimers or heterodimers, including the CRK-like
proto-oncogene adaptor protein/STAT5 heterodimer and NF-κB.
Moreover, IFN-α signaling can activate the PI3K signaling pathway.
IFN-α can also activate VAV guanine nucleotide exchange factor 1,
which elicits a broad response involving numerous transcription
factors, such as tumor protein p53, MYC, ETS transcription factor
ELK1 and the STAT1/STAT2 heterodimer (1,27,28).
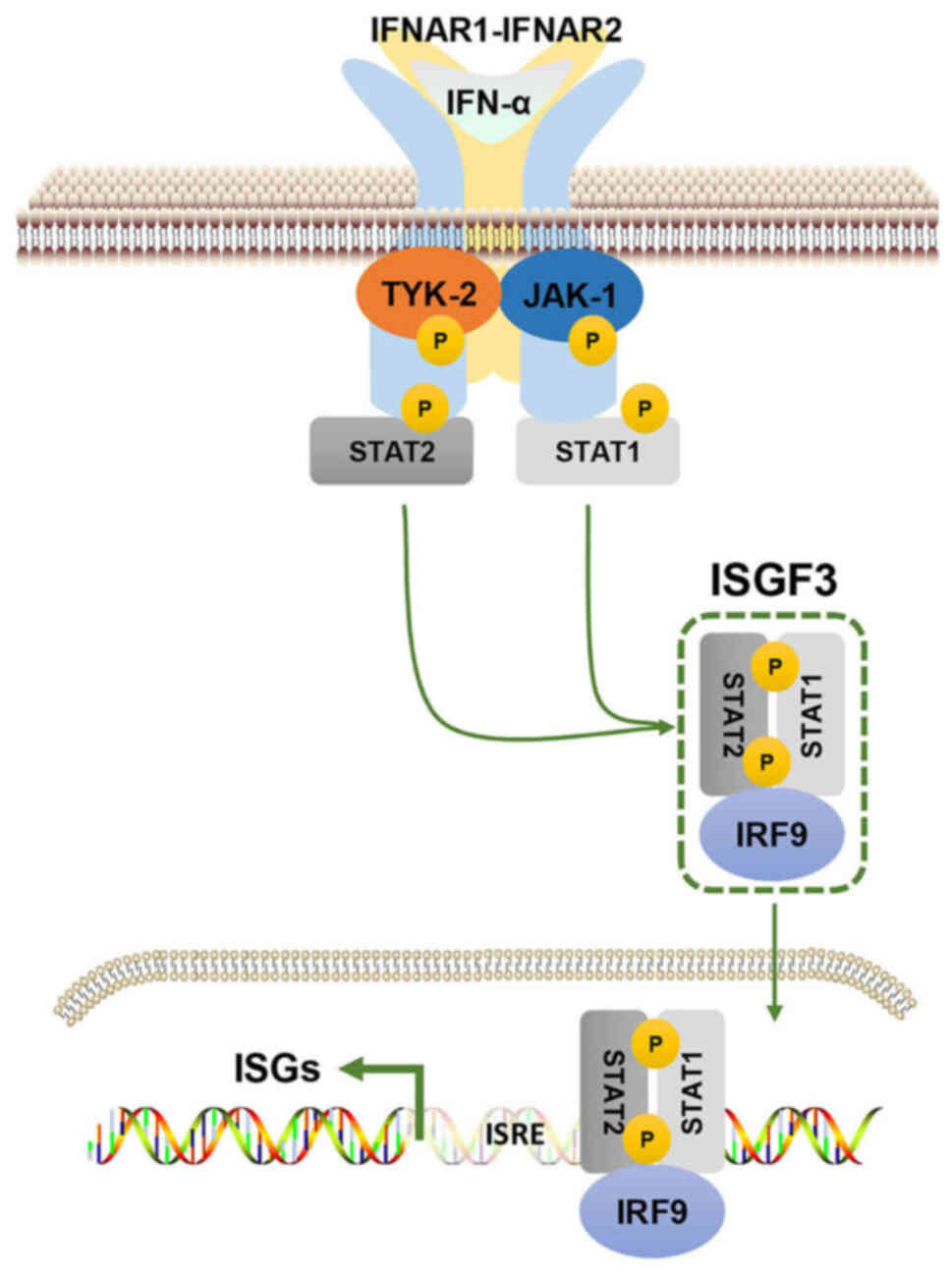 | Figure 1.Canonical IFN-α signaling pathway.
IFN-α binds to IFNAR1/IFNAR2 and subsequently activates JAK-1 and
TYK2, which are two members of the JAK family. This subsequently
leads to the phosphorylation of STAT1 and STAT2. The pSTAT1/pSTAT2
heterodimeric complex combines with IRF9 to form an ISGF3 complex.
ISGF3 binds to the homologous DNA sequence of ISRE to directly
activate the transcription of ISGs. IFN-α, interferon-α; IFNAR,
IFN-α/β receptor subunit; JAK, Janus kinase; TYK2, tyrosine kinase
2; p, phosphorylated; IRF9, IFN regulatory factor 9; ISG,
IFN-stimulated gene; ISGF3, ISG factor 3; ISRE, IFN-sensitive
response element. |
A number of compounds can affect the expression or
signaling of IFN-α, which therefore impacts its underlying
biological functions. RO8191 (CDM-3008), an orally administrable
low-molecular weight compound, is a potent IFN receptor agonist. It
mimics IFN-α via the direct binding of IFNAR2, which activates ISG
expression and JAK/STAT phosphorylation (29,30).
Small ubiquitin-related modifier (SUMO)ylation has been reported to
suppress type I IFN (IFN-α and -β) responses. However, TAK-981, a
selective small-molecule inhibitor of SUMOylation,
pharmacologically reactivates IFN-α and -β signaling. It was
previously demonstrated that in vivo treatment of wild-type
mice with TAK-981 upregulates the gene expression of IFN-α and -β
in blood cells and splenocytes (31). Tilorone dihydrochloride is the
first synthetic, orally active, low-molecular weight compound that
can significantly induce IFN-α in vivo within 24 h of
administration (32,33). It was previously reported that in
patients with Sézary syndrome, TLR7/8 agonists induce inflammatory
cytokines. In this study IFN-α, -β and -γ and a TLR9 agonist
efficiently induced IFN-α and IFN-β, even though this positive
association was not demonstrated for other cytokines (34). Oligo-deoxy-nucleotides with a CpG
motif and double/multi-stranded structure-forming sequences,
function as TLR9 agonists and increase the expression of IFN-α
(35). The small-molecule STAT3
inhibitor FLLL32 is hypothesized to selectively bind to JAK2 and
the STAT3 Src homology 2 domain, which serve vital roles in STAT3
dimerization and the signaling pathway. FLLL32 can downregulate
STAT3 phosphorylation via interactions with IL-6 and IFN-α
(36,37). IRF3 can regulate bacterial and
viral innate immune responses via the modulation of the secretion
of type I IFNs. Thymoquinone, a black cumin-derived compound,
suppresses the IRF3-mediated expression of IFN-α and -β by
suppressing TANK-binding kinase 1 (38). Moreover, abnormal IFN-α signaling
is associated with numerous immune diseases, such as chronic
infection, inflammation and autoimmune disease (39). Therefore, the integrated modulation
of the IFN-α response is important to maintain a balance between
IFN-α-mediated protective effects and cell toxicity due to
dysregulated IFN-α signaling.
Apoptosis
Apoptosis, a type of programmed cell death, is of
great significance for cell development and maintaining tissue
homeostasis. It is a complex and signal-regulated process involving
the participation of numerous molecules (40). Apoptotic events are mainly
performed by the caspase protease family
(cysteine-aspartic-specific proteases). According to their
functions, the caspases can be categorized into three groups: i)
Apoptotic executioner caspases; ii) apoptotic initiator caspases;
and iii) inflammatory caspases (41). Caspase-1, −4, −5, −11, −12, −13 and
−14 belong to the inflammatory caspases, which are associated with
inflammation. The apoptotic initiator caspases are important for
interactions with upstream adaptor molecules. All apoptotic
initiator caspases possess long pro-domains, which contain caspase
activation and recruitment domains (caspase-2 and −9) or death
effector domains (caspase-8 and −10) (42). The apoptotic executioner caspases
(caspase-3, −6 and −7) are typically processed and activated via
upstream caspases and perform apoptosis by cleaving cellular
components. Once the apoptotic signaling pathways are activated a
caspase cascade will occur (43).
It is currently considered that at least three signaling pathways
are related to the occurrence of apoptosis: i) The intrinsic
(mitochondrial) signaling pathway; ii) the extrinsic (death
receptor) signaling pathway; and iii) the endoplasmic reticulum
(ER) stress-related signaling pathway, of which the first two are
recognized as the main apoptotic signaling pathways in most cells
(44).
Extrinsic (death receptor) signaling
pathway
The extrinsic signaling pathway is associated with
the ligation of the tumor necrosis factor (TNF) receptor (TNFR)
superfamily (TNFRSF), including TNFRSF1a, TNFRSF21, TNFRSF25,
TNFRSF10a/b and TNFRSF6. This signaling pathway includes receptor
oligomerization and the recruitment of death domain (DD)-containing
adaptor proteins to the aggregated receptor domains via DD/DD
interactions. These adaptor proteins include a death-effector
domain module, which recruits procaspase-8 and −10 to induce a
death-inducing signaling complex that regulates oligomerization and
consequently activates caspase-8 and −10. The activated caspase-8
and −10 cleave additional downstream caspases, such as caspase-3,
−6 and −7, which triggers the morphological hallmarks of apoptosis,
such as apoptotic body formation, DNA fragmentation, cytoplasmic
condensation and cytoskeletal collapse (45–47).
Intrinsic (mitochondrial) signaling
pathway
The intrinsic apoptotic signaling pathway is
triggered in response to stress stimuli such as heat,
γ-irradiation, UV radiation, growth-factor deprivation, viral
virulence factors, certain oncogenic factors and DNA-damaging
agents (48). These stressors are
driven by different intracellular components that relay signals to
mitochondria, which result in a change in the mitochondrial
membrane permeability (MMP) that is primarily modulated by Bcl-2.
The MMP promotes the secretion of cytochrome c from the
mitochondria, which subsequently interacts with apoptotic
protease-activating factor-1 (Apaf-1) and induces nucleotide
exchange activity. This therefore results in the formation of the
homo-heptameric Apaf-1 complex, namely the apoptosome. Procaspase-9
is cleaved and activated by the apoptosome. The apoptosome complex
and caspase-9 can also form the holoenzyme, which activates the
downstream effectors caspase-3 and −7 (49–52).
ER stress-related signaling
pathway
The ER is important for protein modification,
folding and synthesis and it is also the main reservoir of
Ca2+. An increase in unfolded proteins or a calcium
imbalance leads to ER stress and the unfolded protein response
(UPR) to maintain normal cellular function. However, the prolonged
activation of the UPR may initiate apoptosis if ER protein
homeostasis is not restored (41).
Intracellularly the ER is the main store of Ca2+ ions.
Stress-induced apoptosis involves the release of Ca2+
from the ER into the cytosol. Moreover, ER stress specifically
activates mouse caspase-12 (that is equal to human caspase-4). The
activated caspase-12 translocates from the ER into the cytosol and
subsequently cleaves procaspase-9, which results in caspase-3
activation (53,54). Furthermore, the C/EBP homologous
protein is also responsible for ER stress-induced apoptosis
(55,56).
Mechanisms of IFN-α-induced cancer cell
apoptosis
Regardless of tissue histology or cell type,
apoptosis can be induced by almost all IFN subtypes, including
IFN-α. Furthermore, IFN-α is involved in Fas-associated via death
domain (FADD)/caspase-8 signaling, the disruption of the MMP, the
release of cytochrome c from mitochondria and the activation
of the caspase cascade, which suggests that diverse strategies can
be applied for cancer treatment (16,57).
Possible mechanisms of IFN-α alone or in combination with other
drugs to induce apoptosis in different cancer cell types will be
discussed in this section (Table
I; Fig. 2).
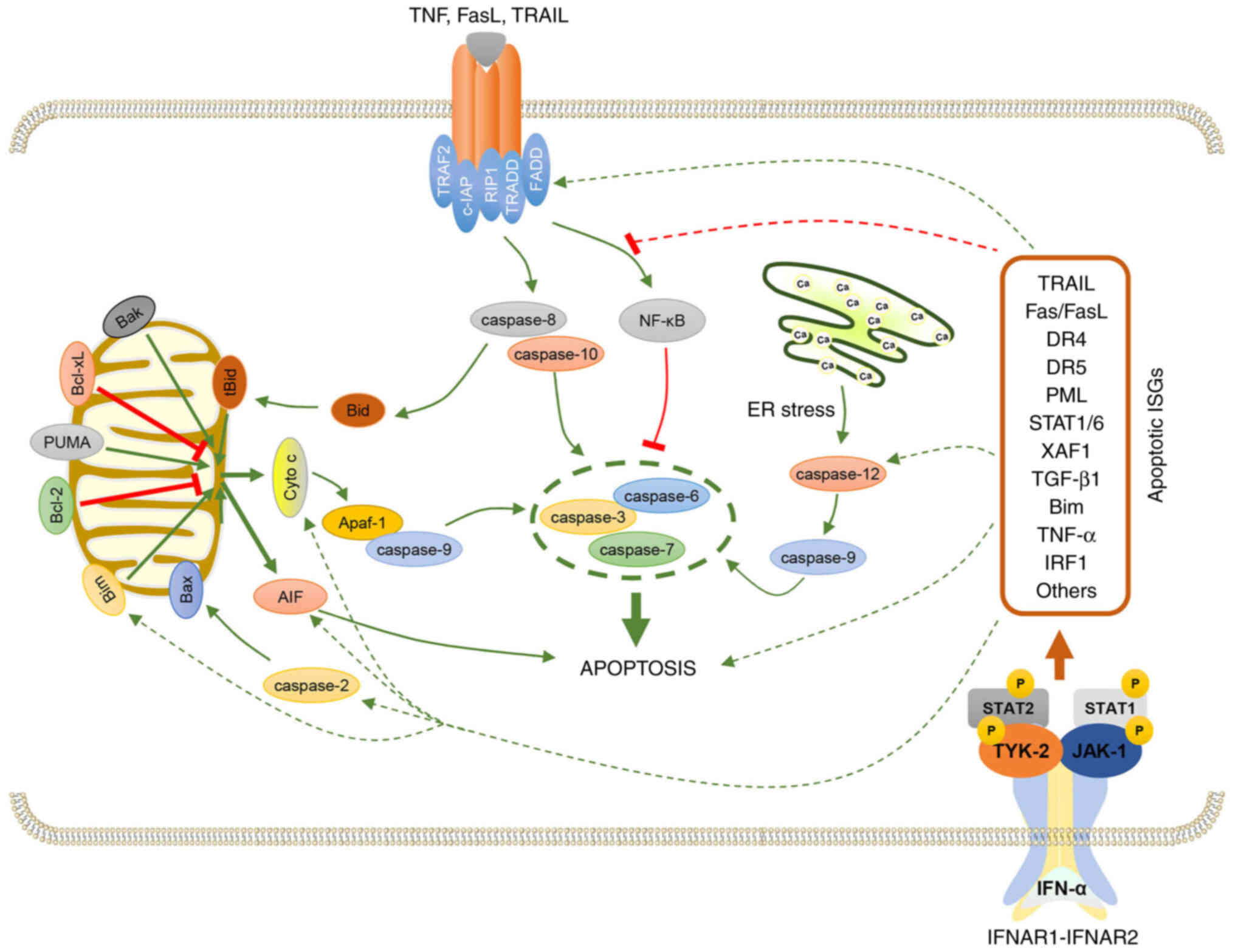 | Figure 2.Mechanism of IFN-α-induced apoptosis.
The apoptotic effects of IFN-α result from the induction of ISGs.
For example, IFN-α upregulates TRAIL, FasL and TNF-α, which bind to
the corresponding receptors and activate caspase-8 and −10. The
activated caspase-8 and −10 subsequently activate caspase-3, −6 and
−7, which results in cell apoptosis. IFN-α also induces other
proapoptotic proteins such as PML, STAT1, STAT6, XAF1, TGF-β1, Bim
and IRF1, which regulate apoptosis. These underlying regulatory
mechanisms are demonstrated in the figure by the green and red
dotted arrows. IFN-α, interferon-α; ISGs, IFN-stimulated genes;
TRAIL, TNF-related apoptosis-inducing ligand; FasL, Fas ligand;
TNF, tumor necrosis factor; PML, promyelocytic leukemia; XAF1,
XIAP-associated factor 1; Bim, Bcl-2-like protein 11; IRF1,
interferon regulatory factor 1. |
 | Table I.Main apoptotic signaling pathways and
mechanisms targeted by IFN-α in cancer. |
Table I.
Main apoptotic signaling pathways and
mechanisms targeted by IFN-α in cancer.
| Treatments | Apoptotic
pathways | Molecules
involved | Types of cancer
cells | (Refs.) |
|---|
| IFN-α2a | The extrinsic
(death receptor) pathway | TRAIL, DR5, NF-κB,
and caspase-8 | HuH-7 and
Hep3B | (58) |
| IFN-α | The extrinsic
(death receptor) pathway | TRAIL and PML | Hep3B, Huh7, Huh6,
HepG2, Chang and CEM | (59) |
|
IFN-α/celecoxib | The extrinsic
(death receptor) pathway | TRAIL, DR4, DR5,
PARP, caspase-3, and caspase-8 | SMMC-7721, HepG2,
and HLCZ01 | (60) |
| IFN-α/aspirin | The intrinsic
(mitochondrial) pathway | Caspase-3,
caspase-9, Bax, JAK1, STAT1, and XAF1 | Bel-7402 and
MHCC97L | (61) |
| IFN-α2b | The intrinsic
(mitochondrial) pathway | TGF-β1, ROS, JNK,
FoxO3a, PUMA, and cholesterol | Preneoplastic rat
hepatocytes | (62–64) |
| IFN-α | The intrinsic
(mitochondrial) pathway; the ER stress-related pathway | Caspase-3, Bim,
PARP, cytochrome c, and caspase-4 | HeLa | (67) |
| IFN-α2a | The intrinsic
(mitochondrial) pathway | Bid, Bak, and
AIF | OVCAR3 | (68) |
|
IFN-α2a/IFN-γ/IL-4-PE | Not mentioned | JAK, STAT1, STAT6,
PARP, caspase-3, and caspase-7 | OVCAR-5 | (69) |
| IFN-α2b | The extrinsic
(death receptor) pathway | ING4, caspase-3,
caspase-8, PARP, and Fas/FasL | A375 and
HT-144 | (53) |
|
IFN-α/bortezomib | The extrinsic
(death receptor) pathway | caspase-3,
caspase-7, caspase-8, caspase-9, PARP, Fas, and FADD | A375, HT-144,
B16F1, JB/MS, 1259 MEL, 18105 MEL, and MEL 39 | (70) |
| IFN-α | The intrinsic
(mitochondrial) pathway | Bak, Bim,
cytochrome c, caspase-2, caspase-3, caspase-8, caspase-9,
AIF, JAK1, and mTOR | NCI-H929 and
U266 | (57,73) |
| IFN-α/TRAIL | The extrinsic
(death receptor) pathway | Caspase-3,
caspase-8, PARP, and ERK | A-498, ACHN, and
786-O | (75) |
| IFN-α/Smac mimetic
BV6 | The extrinsic
(death receptor) pathway | RIP1, FADD,
caspase-8, caspase-9, and caspase-3 | CaKi1, CaKi2,
KTCTL2, KTCTL26, KTCTL30, A498, KTCTL26, KTCTL30, A498, 769P, and
786O | (76) |
| IFN-α/Smac mimetic
BV6 | The extrinsic
(death receptor) pathway | TNF-α, TNFR1, and
IRF1 | MV4-11, OCI-AML3,
Molm13 MonoMac6, and NB4 | (77) |
HCC
HCC is a commonly used cancer model to study the
mechanism of apoptosis caused by IFN-α. Previous studies have
reported the involvement of TNF-related apoptosis-inducing ligand
(TRAIL)-induced apoptosis following IFN-α stimulation in HCC. TRAIL
is a type of proapoptotic protein that can activate caspase-8 via
interacting with the TRAIL receptor, which consequently initiates
apoptosis. Shigeno et al (58) demonstrated that IFN-α pretreatment
could enhance the TRAIL-induction of Hep3B and HuH-7 cell
apoptosis, in which IFN-α increased the expression of TNFRSF10B.
However, this study also demonstrated that IFN-α pretreatment also
suppresses the TRAIL-regulated activation of NF-κB. In addition to
TRAIL, promyelocytic leukemia protein (PML) is also involved in
IFN-α-induced HCC apoptosis (59).
TRAIL functions as a downstream target of PML and both TRAIL and
PML serve essential roles in IFN-α-regulated HCC apoptosis.
Compared with IFN-α stimulation alone, IFN-α in conjunction with
other compounds can enhance cell apoptosis. For example, IFN-α and
celecoxib, a cyclooxygenase-2 inhibitor, synergistically increase
TRAIL-induced HCC apoptosis, which suggests that this combination
may serve as a new therapeutic option for TRAIL-resistant cancer
(60). Moreover, STAT1 can
regulate the proapoptotic effect of IFN-α. Aspirin may increase the
antitumor efficacy of IFN-α on hepatoma cells via activating the
JAK1/STAT1 signaling pathway, which improves IFN-α gene and protein
therapy (61).
Furthermore, in addition to promoting apoptosis in
multiple types of HCC cells, IFN-α can also cause apoptotic events
in rats in early-stage hepatocarcinogenesis. In a model described
in a previous study, it was demonstrated that IFN-α2b initiates the
intrinsic apoptotic cascade by inducing hepatocytes to produce
reactive oxygen species (ROS) and TGF-β1, which ultimately leads to
cell death (62). This previous
study also demonstrated that the endogenous production of ROS
caused by IFN-α2b-activated JNK in rat preneoplastic liver was
responsible for the transcriptional activity and nuclear
translocation of FoxO3a. FoxO3a positively modulates the expression
of proapoptotic Bcl-2 protein family members, such as p53
upregulated modulator of apoptosis (PUMA), which triggers the
mitochondrial apoptotic signaling pathway (63). There is also a correlation between
IFN-induced apoptosis and lipid metabolism (64). Treatment with IFN-α2b, decreases
the synthesis of liver cholesterol and increases its secretion,
which is required for IFN-α2b to promote cell apoptosis. These
aforementioned data have demonstrated the complicated role of
IFN-α2b in the early development of HCC.
Cervical and ovarian cancers
Cervical and ovarian cancers are the two most common
types of female malignant tumors, which severely affect the mental
and physical health of women. In the last decade, antitumor
research based on IFN-induced apoptosis of these two types of
cancer has been ongoing (65,66).
IFN-α promotes HeLa cell apoptosis via the activation of both ER
stress-induced and intrinsic mitochondrial signaling pathways
(67). The activation of
caspase-3, the secretion of cytochrome c from mitochondria,
the downregulation of Bcl-extra-large (Bcl-xL) and the upregulation
of Bcl-2-like protein 11 (Bim) and cleaved poly(ADP-ribose)
polymerase (PARP) are observed following IFN-α treatment, which
suggests that the intrinsic apoptotic signaling pathway is
activated. Furthermore, caspase-4, which is responsible for ER
stress-induced apoptosis, is activated following treatment with
IFN-α. In ovarian cancer OVCAR3 cells, IFN-α2a-induced apoptosis is
regulated by apoptosis-inducing factor (AIF) signaling. IFN-α2a
treatment results in the cleavage of BH3 interacting domain death
agonist that activates mitochondrial Bcl-2 homologous
antagonist/killer to impair the integrity of the mitochondrial
membrane, which leads to AIF secretion. AIF induces nuclear
fragmentation and cell apoptosis after being translocated from the
mitochondria to the nucleus, which indicates a novel
mitochondria-associated apoptotic signaling pathway (68). In a previous study, the combination
of IL-4-Pseudomonas exotoxin, IFN-γ and IFN-α resulted in
increased apoptotic cell death in ovarian cancer. This mechanism of
the synergistic anticancer effect is dependent on IFN-mediated
JAK/STAT signaling and the consequent activation of
apoptosis-related molecules, including caspase-3, −7 and PARP
(69). These aforementioned
studies have provided a theoretical basis for the immunotherapy of
cervical cancer and ovarian cancer based on IFN-α.
Melanoma
Melanoma is the most severe type of skin cancer and
is resistant to existing therapies. The combination of IFN-α and
other drugs has been proven to significantly enhance cell apoptosis
in melanoma. Cai et al (70) reported that inhibitor of growth
family member 4 (ING4) overexpression potentially improves the
effects of IFN-α2b and induces melanoma cell apoptosis. This study
also demonstrated that ING4 overexpression reduces the expression
levels of caspase-3, −8 and PARP and increases the expression
levels of cleaved caspase-3, −8, cleaved PARP and Fas/Fas ligand
(FasL), which indicates the involvement of the Fas/FasL-mediated
death receptor apoptotic signaling pathway. Similar, the
combination of bortezomib and IFN-α leads to enhanced apoptotic
cell death in melanoma cell lines (71). Moreover, decreased levels of the
apoptosis-antagonizing proteins myeloid leukemia-1 and Bcl-2 are
detected following treatment with IFN-α and bortezomib, which
suggests that the intrinsic apoptotic signaling pathway is promoted
via the modulation of protein targets in the mitochondria. However,
bortezomib in combination with IFN-α stimulates the extrinsic
signaling pathway of apoptosis via the activation of FADD-induced
caspase-8. Therefore, a combination of IFN-α and other drugs may be
effective against apoptosis in melanoma cells.
Multiple myeloma
IFN-α has been used in the treatment of several
hematological neoplasia, including multiple myeloma (72). In human myeloma H929 and U266 cell
lines, apoptosis induced by IFN-α results in phosphatidylserine
exposure, MMP loss, Bak conformational change, Bim upregulation,
reduced levels of cytochrome c release from the mitochondria
and a low rate of caspase activation, as well as AIF release.
Moreover, PUMA levels increase following IFN-α treatment, whereas
PUMA knockdown has no effect on IFN-α-induced apoptosis, which
suggests that PUMA is not required for IFN-α triggered apoptosis.
Furthermore, IFN-α-induced apoptosis is completely inhibited by
JAK1, whereas rapamycin, an mTOR inhibitor, mitigates apoptosis in
U266 cells but potentiates it in H929 cells. The potentiating
action of rapamycin on H929 cell apoptosis is related to the
upregulation of Bim levels induced by IFN-α (73). A previous study reported that
IFN-α-induced U266 cell apoptosis is related to the activation of
caspase-2, −3, −8 and −9. The activation of caspase-3 relies on the
activities of caspases-8 and −9 and caspase-8 lies upstream of
IFN-α-related caspase cascades. The interaction between the
Fas-receptor and its ligand is independent of IFN-α-induced
apoptosis (57). These data have
demonstrated that IFN-α induces apoptosis in myeloma cells via the
activation of the mitochondrial pathway and that inhibitors of mTOR
or JAK1 may facilitate IFN-α maintenance therapy in patients with
multiple myeloma.
RCC
RCC is the third most common urological cancer and
has a poor prognosis. Researchers have long been committed to the
study of treatment strategies against RCC, including the
application of IFNs (74).
Although IFN-α can directly promote apoptosis in various cancer
cell lines, as demonstrated in the aforementioned sections, it is
currently used to treat RCC mainly in combination with other
antineoplastic agents. Clark et al (75) demonstrated that TRAIL and IFN-α act
synergistically to induce RCC cell death. IFN-α on its own does not
cause RCC cell apoptosis as there is no effect on the expression of
TRAIL or death receptors and other known mediators of the intrinsic
and extrinsic apoptotic signaling cascades, including caspase-3,
−8, PARP and Bcl-2 family proteins. However, the extracellular
signal-regulated kinases (ERKs) are prominently activated upon
IFN-α treatment alone or in combination with TRAIL. The apoptotic
synergy between TRAIL and IFN-α is due at least in part to the
activation of ERK mediated by IFN-α.
IFN-α together with BV6, which antagonizes inhibitor
of apoptosis proteins, displays cooperative antitumor activity in
different cancer cell lines. In RCC cells, BV6/IFN-α have a
significant antitumor effect, including in reducing cell viability
and inducing apoptosis (76).
Molecular studies have reported that the scaffold function of
receptor-interacting protein 1 (RIP1) is important for
BV6/IFN-α-induced apoptosis. BV6 and IFN-α work together to induce
caspase activation by forming a cytosolic cell death complex
(caspase-8, FADD and RIP1). The synergistic effect of IFN-α and BV6
in acute myeloid leukemia cell death has also been identified
(77). BV6 and IFN-α cooperate to
enhance the expression of TNF-α. As they are secreted into the
supernatant they initiate a TNFR1 loop that triggers cell
apoptosis. IFN-α/BV6-induced cell apoptosis is also dependent on
IRF1. This combination approach of IFN-α and BV6 may serve as a
potential strategy to induce apoptosis in cancer cells.
The activation of effector caspases can be achieved
by the convergence of the extrinsic and intrinsic apoptotic
signaling pathways. Crosstalk between these two signaling pathways
has previously been reported. For example, caspase-3, −6 and −7 are
involved in the execution phase of apoptosis via both the intrinsic
and extrinsic signaling pathways (41). In the intrinsic pathway, caspase-3
and −7 are proteolytically activated by caspase-6. Subsequently,
caspase-8 is cleaved or translocated into the nucleus to cleave its
target substrates, which results in cell death. Therefore,
caspase-8 cleavage and apoptosis are markedly attenuated via the
inhibition of caspase-6 activity in cells, which indicates that
caspase-8 is mainly activated by caspase-6 in vivo (78–80).
The association between these two signaling pathways demonstrates
that stress-inducers or chemotherapeutic agents may sensitize cells
to death ligand-induced apoptosis. This information is important to
determine the proapoptotic and antitumor mechanisms of IFN-α.
Conclusions and future prospects
The aim of the present review was to assess the
scientific advances made concerning IFN-α-induced cancer cell
apoptosis. In most cases, IFN-α needs to be used in combination
with other drugs or molecules in order to have an improved
antitumor effect. This information will provide a focus area for
future research into the clinical application of IFN-α in cancer
treatment.
Over the past decade, numerous clinical trials
involving IFN-α have been implemented worldwide for use in
different types of cancer (81–85).
However, the mechanisms of IFN-α antitumor activity do not only
include the proapoptotic effects mentioned in the present review,
but also consist of various other functions, including
antiproliferation, immunological and regulatory effects. These
other areas still require further research. Furthermore, it is
necessary to clarify the mechanisms of IFN-α toxicity so that IFN-α
can be safely used as an antitumor agent either alone or in
combination with other anticancer drugs (86,87).
In-depth consideration of these aspects may help establish
eligibility criteria for cancer therapy.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation of Hebei Province (grant no. H2019208216), the
Scientific Research Foundation for PhD (grant no. 81/1181286), the
Science and Technology Research Program for Colleges and
Universities in Hebei Province (grant no. ZD2022011) and the
Natural Science Foundation of Hebei Province (grant no.
H2020208002).
Availability of data and materials
Not applicable.
Authors' contributions
WS and YW designed the framework and theme of the
review. WS retrieved the literature and wrote the first draft. XY
and YF participated in writing the manuscript. Data authentication
is not applicable. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cheon H, Borden EC and Stark GR:
Interferons and their stimulated genes in the tumor
microenvironment. Semin Oncol. 41:156–173. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bekisz J, Schmeisser H, Hernandez J,
Goldman ND and Zoon KC: Human interferons alpha, beta and omega.
Growth Factors. 22:243–251. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pestka S: The human interferon-alpha
species and hybrid proteins. Semin Oncol. 24 (Suppl 9):S9-4-S9-17.
1997.
|
|
4
|
El-Baky NA and Redwan EM: Therapeutic
alpha-interferons protein: Structure, production, and biosimilar.
Prep Biochem Biotechnol. 45:109–127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lazear HM, Schoggins JW and Diamond MS:
Shared and distinct functions of type I and type III interferons.
Immunity. 50:907–923. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Blaauboer A, Sideras K, van Eijck CHJ and
Hofland LJ: Type I interferons in pancreatic cancer and development
of new therapeutic approaches. Crit Rev Oncol Hematol.
159:1032042021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Grilo AL and Mantalaris A: Apoptosis: A
mammalian cell bioprocessing perspective. Biotechnol Adv.
37:459–475. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
McNab F, Mayer-Barber K, Sher A, Wack A
and O'Garra A: Type I interferons in infectious disease. Nat Rev
Immunol. 15:87–103. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zitvogel L, Galluzzi L, Kepp O, Smyth MJ
and Kroemer G: Type I interferons in anticancer immunity. Nat Rev
Immunol. 15:405–414. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bekisz J, Baron S, Balinsky C, Morrow A
and Zoon KC: Antiproliferative properties of type I and type II
interferon. Pharmaceuticals (Basel). 3:994–1015. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Haji Abdolvahab M, Mofrad MR and
Schellekens H: Interferon beta: From molecular level to therapeutic
effects. Int Rev Cell Mol Biol. 326:343–372. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sin WX, Li P, Yeong JP and Chin KC:
Activation and regulation of interferon-β in immune responses.
Immunol Res. 53:25–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Markowitz CE: Interferon-beta: Mechanism
of action and dosing issues. Neurology. 68 (Suppl 4):S8–S11. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kali SK, Dröge P and Murugan P: Interferon
β, an enhancer of the innate immune response against SARS-CoV-2
infection. Microb Pathog. 158:1051052021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jakimovski D, Kolb C, Ramanathan M,
Zivadinov R and Weinstock-Guttman B: Interferon β for multiple
sclerosis. Cold Spring Harb Perspect Med. 8:a0320032018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chawla-Sarkar M, Lindner DJ, Liu YF,
Williams BR, Sen GC, Silverman RH and Borden EC: Apoptosis and
interferons: Role of interferon-stimulated genes as mediators of
apoptosis. Apoptosis. 8:237–249. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
De Groof A, Ducreux J, Aleva F, Long AJ,
Ferster A, van der Ven A, van de Veerdonk F, Houssiau FA and
Lauwerys BR: STAT3 phosphorylation mediates the stimulatory effects
of interferon alpha on B cell differentiation and activation in
SLE. Rheumatology (Oxford). 59:668–677. 2020.PubMed/NCBI
|
|
18
|
Indraccolo S: Interferon-alpha as
angiogenesis inhibitor: Learning from tumor models. Autoimmunity.
43:244–247. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kotredes KP and Gamero AM: Interferons as
inducers of apoptosis in malignant cells. J Interferon Cytokine
Res. 33:162–170. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pestka S, Krause CD and Walter MR:
Interferons, interferon-like cytokines, and their receptors.
Immunol Rev. 202:8–32. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pestka S: Purification and cloning of
interferon alpha. Curr Top Microbiol Immunol. 316:23–37.
2007.PubMed/NCBI
|
|
22
|
Pestka S: The human interferon alpha
species and receptors. Biopolymers. 55:254–287. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wittling MC, Cahalan SR, Levenson EA and
Rabin RL: Shared and unique features of human interferon-beta and
interferon-alpha subtypes. Front Immunol. 11:6056732021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gibbert K, Schlaak JF, Yang D and Dittmer
U: IFN-α subtypes: Distinct biological activities in anti-viral
therapy. Br J Pharmacol. 168:1048–1058. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ortaldo JR, Herberman RB, Harvey C,
Osheroff P, Pan YC, Kelder B and Pestka S: A species of human alpha
interferon that lacks the ability to boost human natural killer
activity. Proc Natl Acad Sci USA. 81:4926–4929. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schreiber G: The molecular basis for
differential type I interferon signaling. J Biol Chem.
292:7285–7294. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schreiber G and Piehler J: The molecular
basis for functional plasticity in type I interferon signaling.
Trends Immunol. 36:139–149. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schneider WM, Chevillotte MD and Rice CM:
Interferon-stimulated genes: A complex web of host defenses. Annu
Rev Immunol. 32:513–545. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Furutani Y, Toguchi M, Shiozaki-Sato Y,
Qin XY, Ebisui E, Higuchi S, Sudoh M, Suzuki H, Takahashi N,
Watashi K, et al: An interferon-like small chemical compound
CDM-3008 suppresses hepatitis B virus through induction of
interferon-stimulated genes. PLoS One. 14:e02161392019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Konishi H, Okamoto K, Ohmori Y, Yoshino H,
Ohmori H, Ashihara M, Hirata Y, Ohta A, Sakamoto H, Hada N, et al:
An orally available, small-molecule interferon inhibits viral
replication. Sci Rep. 2:2592012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lightcap ES, Yu P, Grossman S, Song K,
Khattar M, Xega K, He X, Gavin JM, Imaichi H, Garnsey JJ, et al: A
small-molecule SUMOylation inhibitor activates antitumor immune
responses and potentiates immune therapies in preclinical models.
Sci Transl Med. 13:eaba77912021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Krueger RE and Mayer GD: Tilorone
hydrochloride: An orally active antiviral agent. Science.
169:1213–1214. 1970. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang J, Yao Q and Liu Z: An effective
synthesis method for tilorone dihydrochloride with obvious IFN-α
Inducing Activity. Molecules. 20:21458–21463. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Manfrere KC, Torrealba MP, Miyashiro DR,
Oliveira LM, de Carvalho GC, Lima JF, Branco AC, Pereira NZ,
Pereira J, Sanches JA Jr and Sato MN: Toll-like receptor agonists
partially restore the production of pro-inflammatory cytokines and
type I interferon in Sézary syndrome. Oncotarget. 7:74592–74601.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu D, Putta MR, Bhagat L, Dai M, Wang D,
Trombino AF, Sullivan T, Kandimalla ER and Agrawal S: Impact of
secondary structure of toll-like receptor 9 agonists on interferon
alpha induction. Antimicrob Agents Chemother. 52:4320–4325. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin L, Hutzen B, Zuo M, Ball S, Deangelis
S, Foust E, Pandit B, Ihnat MA, Shenoy SS, Kulp S, et al: Novel
STAT3 phosphorylation inhibitors exhibit potent growth-suppressive
activity in pancreatic and breast cancer cells. Cancer Res.
70:2445–2454. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lin L, Deangelis S, Foust E, Fuchs J, Li
C, Li PK, Schwartz EB, Lesinski GB, Benson D, Lü J, et al: A novel
small molecule inhibits STAT3 phosphorylation and DNA binding
activity and exhibits potent growth suppressive activity in human
cancer cells. Mol Cancer. 9:2172010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Aziz N, Son YJ and Cho JY: Thymoquinone
suppresses IRF-3-mediated expression of type I interferons via
suppression of TBK1. Int J Mol Sci. 19:13552018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen K, Liu J and Cao X: Regulation of
type I interferon signaling in immunity and inflammation: A
comprehensive review. J Autoimmun. 83:1–11. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Carneiro BA and El-Deiry WS: Targeting
apoptosis in cancer therapy. Nat Rev Clin Oncol. 17:395–417. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jin Z and El-Deiry WS: Overview of cell
death signaling pathways. Cancer Biol Ther. 4:139–163. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vigneswara V and Ahmed Z: The role of
caspase-2 in regulating cell fate. Cells. 9:12592020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Obeng E: Apoptosis (programmed cell death)
and its signals-A review. Braz J Biol. 81:1133–1143. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kashyap D, Garg VK and Goel N: Intrinsic
and extrinsic pathways of apoptosis: Role in cancer development and
prognosis. Adv Protein Chem Struct Biol. 125:73–120. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tummers B and Green DR: Caspase-8:
Regulating life and death. Immunol Rev. 277:76–89. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sayers TJ: Targeting the extrinsic
apoptosis signaling pathway for cancer therapy. Cancer Immunol
Immunother. 8:1173–1180. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ashkenazi A: Targeting the extrinsic
apoptotic pathway in cancer: Lessons learned and future directions.
J Clin Invest. 125:487–489. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gibson CJ and Davids MS: BCL-2 antagonism
to target the intrinsic mitochondrial pathway of apoptosis. Clin
Cancer Res. 22:5021–5029. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Luo X, Budihardjo I, Zou H, Slaughter C
and Wang X: Bid, a Bcl2 interacting protein, mediates cytochrome c
release from mitochondria in response to activation of cell surface
death receptors. Cell. 94:481–490. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bock FJ and Tait SWG: Mitochondria as
multifaceted regulators of cell death. Nat Rev Mol Cell Biol.
21:85–100. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Burke PJ: Mitochondria, bioenergetics and
apoptosis in cancer. Trends Cancer. 3:857–870. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xiong S, Mu T, Wang G and Jiang X:
Mitochondria-mediated apoptosis in mammals. Protein Cell.
5:737–749. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sano R and Reed JC: ER stress-induced cell
death mechanisms. Biochim Biophys Acta. 1833:3460–3470. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Obeng EA and Boise LH: Caspase-12 and
caspase-4 are not required for caspase-dependent endoplasmic
reticulum stress-induced apoptosis. J Biol Chem. 280:29578–29587.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hu H, Tian M, Ding C and Yu S: The C/EBP
homologous protein (CHOP) transcription factor functions in
endoplasmic reticulum stress-induced apoptosis and microbial
infection. Front Immunol. 9:30832019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Rozpedek W, Pytel D, Mucha B, Leszczynska
H, Diehl JA and Majsterek I: The role of the PERK/eIF2α/ATF4/CHOP
signaling pathway in tumor progression during endoplasmic reticulum
stress. Curr Mol Med. 6:533–544. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Thyrell L, Erickson S, Zhivotovsky B,
Pokrovskaja K, Sangfelt O, Castro J, Einhorn S and Grandér D:
Mechanisms of Interferon-alpha induced apoptosis in malignant
cells. Oncogene. 21:1251–1262. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Shigeno M, Nakao K, Ichikawa T, Suzuki K,
Kawakami A, Abiru S, Miyazoe S, Nakagawa Y, Ishikawa H, Hamasaki K,
et al: Interferon-alpha sensitizes human hepatoma cells to
TRAIL-induced apoptosis through DR5 upregulation and NF-kappa B
inactivation. Oncogene. 22:1653–1662. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Herzer K, Hofmann TG, Teufel A, Schimanski
CC, Moehler M, Kanzler S, Schulze-Bergkamen H and Galle PR:
IFN-alpha-induced apoptosis in hepatocellular carcinoma involves
promyelocytic leukemia protein and TRAIL independently of p53.
Cancer Res. 69:855–862. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zuo C, Qiu X, Liu N, Yang D, Xia M, Liu J,
Wang X, Zhu H, Xie H, Dan H, et al: Interferon-α and
cyclooxygenase-2 inhibitor cooperatively mediates TRAIL-induced
apoptosis in hepatocellular carcinoma. Exp Cell Res. 333:316–326.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Li T, Dong ZR, Guo ZY, Wang CH, Tang ZY,
Qu SF, Chen ZT, Li XW and Zhi XT: Aspirin enhances IFN-α-induced
growth inhibition and apoptosis of hepatocellular carcinoma via
JAK1/STAT1 pathway. Cancer Gene Ther. 20:366–374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Quiroga AD, Alvarez Mde L, Parody JP,
Ronco MT, Francés DE, Pisani GB, Carnovale CE and Carrillo MC:
Involvement of reactive oxygen species on the apoptotic mechanism
induced by IFN-alpha2b in rat preneoplastic liver. Biochem
Pharmacol. 73:1776–1785. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Parody JP, Ceballos MP, Quiroga AD,
Frances DE, Carnovale CE, Pisani GB, Alvarez ML and Carrillo MC:
FoxO3a modulation and promotion of apoptosis by interferon-α2b in
rat preneoplastic liver. Liver Int. 34:1566–1577. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Quiroga AD, Vera MC, Ferretti AC, Lucci A,
Comanzo CG, Lambertucci F, Ceballos MP and Carrillo MC: IFN-α-2b
induces apoptosis by decreasing cellular cholesterol levels in rat
preneoplastic hepatocytes. Cytokine. 133:1551722020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lee SJ, Yang A, Wu TC and Hung CF:
Immunotherapy for human papillomavirus-associated disease and
cervical cancer: Review of clinical and translational research. J
Gynecol Oncol. 27:e512016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Stewart C, Ralyea C and Lockwood S:
Ovarian cancer: An integrated review. Semin Oncol Nurs. 35:151–156.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Shi WY, Cao C and Liu L: Interferon α
induces the apoptosis of cervical cancer HeLa cells by activating
both the intrinsic mitochondrial pathway and endoplasmic reticulum
stress-induced pathway. Int J Mol Sci. 17:18322016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Miyake K, Bekisz J, Zhao T, Clark CR and
Zoon KC: Apoptosis-inducing factor (AIF) is targeted in
IFN-α2a-induced Bid-mediated apoptosis through Bak activation in
ovarian cancer cells. Biochim Biophys Acta. 1823:1378–1388. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Green DS, Husain SR, Johnson CL, Sato Y,
Han J, Joshi B, Hewitt SM, Puri RK and Zoon KC: Combination
immunotherapy with IL-4 Pseudomonas exotoxin and IFN-α and IFN-γ
mediate antitumor effects in vitro and in a mouse model of human
ovarian cancer. Immunotherapy. 11:483–496. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Cai L, Liu J and Wang Y, Chen H, Ma Y and
Wang Y and Wang Y: Enhanced anti-melanoma efficacy of interferon
α-2b via overexpression of ING4 by enhanced Fas/FasL-mediated
apoptosis. Oncol Lett. 15:9577–9583. 2018.PubMed/NCBI
|
|
71
|
Lesinski GB, Raig ET, Guenterberg K, Brown
L, Go MR, Shah NN, Lewis A, Quimper M, Hade E, Young G, et al:
IFN-alpha and bortezomib overcome Bcl-2 and Mcl-1 overexpression in
melanoma cells by stimulating the extrinsic pathway of apoptosis.
Cancer Res. 68:8351–8360. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Minnie SA and Hill GR: Immunotherapy of
multiple myeloma. J Clin Invest. 130:1565–1575. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Gómez-Benito M, Balsas P, Carvajal-Vergara
X, Pandiella A, Anel A, Marzo I and Naval J: Mechanism of apoptosis
induced by IFN-alpha in human myeloma cells: Role of Jak1 and Bim
and potentiation by rapamycin. Cell Signal. 19:844–854. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Deleuze A, Saout J, Dugay F, Peyronnet B,
Mathieu R, Verhoest G, Bensalah K, Crouzet L, Laguerre B,
Belaud-Rotureau MA, et al: Immunotherapy in renal cell carcinoma:
The future is now. Int J Mol Sci. 21:25322020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Clark PE, Polosukhina DA, Gyabaah K, Moses
HL, Thorburn A and Zent R: TRAIL and interferon-alpha act
synergistically to induce renal cell carcinoma apoptosis. J Urol.
184:1166–1174. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Reiter M, Eckhardt I, Haferkamp A and
Fulda S: Smac mimetic sensitizes renal cell carcinoma cells to
interferon-α-induced apoptosis. Cancer Lett. 375:1–8. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Bake V, Roesler S, Eckhardt I, Belz K and
Fulda S: Synergistic interaction of Smac mimetic and IFNα to
trigger apoptosis in acute myeloid leukemia cells. Cancer Lett.
355:224–231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Cowling V and Downward J: Caspase-6 is the
direct activator of caspase-8 in the cytochrome c-induced apoptosis
pathway: Absolute requirement for removal of caspase-6 prodomain.
Cell Death Diff. 9:1046–1056. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Inoue S, Browne G, Melino G and Cohen GM:
Ordering of caspases in cells undergoing apoptosis by the intrinsic
pathway. Cell Death Differ. 16:1053–1061. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Slee EA, Harte MT, Kluck RM, Wolf BB,
Casiano CA, Newmeyer DD, Wang HG, Reed JC, Nicholson DW, Alnemri
ES, et al: Ordering the cytochrome c-initiated caspase cascade:
Hierarchical activation of caspases-2, −3, −6, −7, −8, and −10 in a
caspase-9-dependent manner. J Cell Biol. 144:281–292. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Aricò E, Castiello L, Capone I, Gabriele L
and Belardelli F: Type I interferons and cancer: An evolving story
demanding novel clinical applications. Cancers (Basel).
11:19432019. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Muñoz de Escalona Rojas JE, García Serrano
JL, Cantero Hinojosa J, Padilla Torres JF and Bellido Muñoz RM:
Application of interferon alpha 2b in conjunctival intraepithelial
neoplasia: Predictors and prognostic factors. J Ocul Pharmacol
Ther. 30:489–494. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Yoon SY and Won JH: The clinical role of
interferon alpha in Philadelphia-negative myeloproliferative
neoplasms. Blood Res. 56:S44–S50. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Ghosh D, Ghosh D and Parida P:
Physiological proteins in therapeutics: A current review on
interferons. Mini Rev Med Chem. 12:947–952. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Di Trolio R, Simeone E, Di Lorenzo G,
Buonerba C and Ascierto PA: The use of interferon in melanoma
patients: A systematic review. Cytokine Growth Factor Rev.
2:203–312. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Hauschild A, Kähler KC, Schäfer M and
Fluck M: Interdisciplinary management recommendations for toxicity
associated with interferon-alfa therapy. J Dtsch Dermatol Ges.
6:829–838. 2008.(In English, German). PubMed/NCBI
|
|
87
|
Conlon KC, Miljkovic MD and Waldmann TA:
Cytokines in the treatment of cancer. J Interferon Cytokine Res.
39:6–21. 2019. View Article : Google Scholar : PubMed/NCBI
|














