Introduction
Distant metastases of cancer are responsible for
most cancer-associated deaths. Breast cancer (BC), which is the
most common cancer type in women worldwide, has enormous
socio-economic and public health impacts. The progression of BC
occurs through a series of gradually abnormal stages, beginning
with ductal carcinoma in situ (DCIS), which, if untreated,
might progress to invasive ductal carcinoma (IDC) (1). IDC, the most common form of BC
(2), is a histologically
heterogeneous group of breast lesions with the potential for
progression to metastatic BC (3).
Ovarian cancer (OC) is less common than BC but, due
to the absence of signs or symptoms associated with early-stage
disease, it is responsible for more deaths than other cancer types
in women. High-grade serous OC (HGSOCs), which account for 68% of
OC cases, are aggressive neoplasms commonly diagnosed at an
advanced stage and have the worst prognosis (4). Complications associated with OC
progression that have an ultimate fatal outcome occur in ~75% of
patients despite good initial responses to chemotherapy (5).
BC type 1 gene (BRCA1) is a DNA repair and
cancer suppressor gene that plays an essential role in maintaining
genome integrity (6). Cells
lacking BRCA1 protein are inclined to repair DNA damage by an
error-prone mechanism resulting in gross chromosomal rearrangements
and the generation of mutations that lead to carcinogenesis
(7). Thus, female carriers of
germline BRCA1 pathogenic mutations are at an increased risk
of developing aggressive BC and OC at an early age. The
inactivation of BRCA1 by epigenetic alteration is an
alternative mechanism during sporadic BC and OC carcinogenesis
(8,9). BC tumors, harboring hypermethylated
BRCA1 promoter, display pathological features similar to
BRCA1-mutated hereditary BC (8,10).
Both types of tumors occur at an early age and are associated with
the triple-negative (TNG) BC subtype (11,12).
Moreover, the methylated BRCA1 promoter occurs in all
histological types of epithelial OC, including serous, endometrioid
and clear cell carcinomas (8,13,14).
In 2008, Snell et al (15) made a breakthrough, finding that
methylated BRCA1 promoter is observed in peripheral white
blood cells (WBC) of patients with mutation-negative familial BC
and cancer-free controls. Since then, several studies have shown
the association of WBC BRCA1 promoter methylation with the
risk of developing early onset BC and high-grade serous OC, with
pathological features similar to those of patients with germline
mutated BRCA1 (15–24). The detection of BRCA1
promoter methylation in the WBCs of cancer-free (CF) females has
raised the question of whether those individuals are at risk of
developing breast and ovarian cancer (15,16,20,21,24,25).
Our previous study demonstrated a strong association between
BRCA1 promoter methylation and cancer-associated molecular
changes in WBCs of CF BRCA1 methylation carriers (21). However, further studies are still
needed to confirm the cancer risk of those individuals.
MicroRNA (miRNA/miR) is a type of cancer-associated
molecule that regulates various cellular mechanisms, such as
proliferation, differentiation and oncogenesis (26). These miRNAs are small 18–22
base-pair non-coding RNA molecules that play a crucial role in
regulating gene expression by binding to the 3′-untranslated region
of mRNA of the target gene. Long-lasting exposure to carcinogens
results in miRNA alterations that activate carcinogenic mechanisms
(27). The activated carcinogenic
process, such as chromosome deletion and silencing of miRNA host
genes, results in the irreversible loss of miRNA. Thus, miRNAs are
a sensitive tool in detecting carcinogenic exposure and the
pathological consequences induced by that exposure (27). Amongst the identified miRNAs is
miR-126. The expression of miR-126 is increased as a defense
mechanism to asbestos exposure. The subsequent loss of miR-126, due
to the accumulation of DNA damage and chromosome deletion, leads to
malignant mesothelioma (27).
miR-126 is one of several miRNAs that play critical
roles in several human cancer types. miR-126 is located within the
7th intron of the epidermal growth factor-like protein 7 gene, and
acts as a suppressor of metastasis in several cancer types. Loss of
miR-126 expression in tumor tissue is associated with poor distal
metastasis-free survival, and restoration of miR-126 reduces
overall tumor growth and proliferation (28). Studies have shown that miR-126
differentiates malignant BC from benign BC (29,30).
The patients with DCIS BC have a lower level of tissue and
circulating miR-126 compared with normal adjacent tissue and
healthy controls, respectively (31). Furthermore, downregulation of
miR-126 is associated with aggressive OC with a poor prognosis
(32,33). However, another study has observed
upregulation of miR-126 in OC (34).
In contrast to the use of plasma miRNA, few studies
have evaluated the use of miRNA in peripheral blood WBCs as a
biomarker of cancer risk. In the present study, miR-126 in WBCs and
plasma is investigated as a potential biomarker for the early
prediction of BC and OC in CF BRCA1-methylated female
carriers.
Materials and methods
Patient population
Fresh blood samples (10 ml) were collected from 502
patients with BC and 187 patients with OC who visited the
Department of Oncology in King Faisal Specialist Hospital and
Research Centre (Riyadh, Saudi Arabia) between November 2017 and
November 2021. The age of the patients ranged from 20–83 years
(median, 48 years) for BC and 18–88 years (median, 53 years) for
OC. Age, histological grade, estrogen receptor status and
progesterone receptor status were provided by the Department of
Pathology. For the CF female group, 10 ml fresh blood was collected
from 425 CF female volunteers with an age range from 15–50 years.
For newborn females, 20 leftover WBC RNA samples from our previous
study (24) were used. Ethical
approval (approval no. RAC #2170017) was obtained from the Human
Research Ethics Committee of King Faisal Specialist Hospital and
Research Centre. All participants provided written informed
consent. The guardian of the volunteers provided written informed
consent for participants <18 years old.
DNA and RNA isolation from WBC
Each fresh blood sample was collected in two BD
Vacutainer EDTA (Becton, Dickinson and Company) blood collection
tubes. The tubes were centrifuged immediately at 4°C for 10 min at
1,962 × g. The supernatants were frozen at −80°C in Eppendorf tubes
for subsequent circulating RNA extraction using the QIAamp
Circulating Nucleic Acid Kit (Qiagen GmbH). The WBC layers were
collected and transferred into two 2-ml Eppendorf tubes. One tube
contained 900 ml RBC lysis solution for subsequent DNA extraction,
and the other tube contained 1.2 ml RNALater solution for
subsequent RNA extraction using the Gentra Puregene Blood Kit and
RiboPure Blood Kit (Ambion; Thermo Fisher Scientific, Inc.),
respectively (24).
Methylation-specific polymerase chain
reaction (PCR)
Next, 2 µg WBC DNA was treated with sodium bisulfate
and purified using the EpiTect Bisulfite kit (Qiagen GmbH)
following the manufacturer's recommendations. The treated DNA was
amplified using BRCA1 PCR primers that distinguish between
methylated and unmethylated DNA (Table
I) (8). The PCR conditions
used were an initial cycle at 95°C for 1 min, then 40 cycles of
65°C for 30 sec, 72°C for 30 sec and a final extension at 72°C for
7 min. All reactions were repeated at least twice.
 | Table I.Methylation-specific PCR and
RT-quantitative PCR primers. |
Table I.
Methylation-specific PCR and
RT-quantitative PCR primers.
| Primer | Sequence
(5′-3′) | Annealing
temperature, °C |
|---|
| M BRCA1 |
| 65 |
|
Forward |
GGTTAATTTAGAGTTTCGAGAGACG |
|
|
Reverse |
TCAACGAACTCACGCCGCGCAATCG |
|
| U BRCA1 |
| 65 |
|
Forward |
GGTTAATTTAGAGTTTTGAGAGATG |
|
|
Reverse |
TCAACAAACTCACACCACACAATCA |
|
| RT
BRCA1 |
| 59 |
|
Forward |
TGTAGGCTCCTTTTGGTTATATCATTC |
|
|
Reverse |
CATGCTGAAACTTCTCAACCAGAA |
|
| β-actin |
| 59 |
|
Forward |
TCCCTGGAGAAGAGCTACGA |
|
|
Reverse |
TGAAGGTAGTTTCGTGGATGC |
|
| miR-126
Stem-loop |
CGGCCCAUUAUUACUUUUGGUACGCGCUAUGC | 60 |
|
|
CACUCUCAACUCGUACCGUGAGUAAUAAUGC |
|
| U6 |
GTGCTCGCTTCGGCAGCACATATACTAAAATTGGAA | 60 |
|
|
CGATACAGAGAAGATTAGCATGGCCCCTGCGCAAG |
|
|
|
GATGACACGCAAATTCGTGAAGCGTTCCATATTTT |
|
Stem-loop PCR assay
Reverse transcription-quantitative PCR (RT-qPCR) for
miR-126 was performed using a stem-loop RT primer and TaqMan miRNA
RT kit (catalog no. 4427975; Applied Biosystems; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol (Table I) and using the thermocycling
conditions stated below. The small nuclear RNA U6 (U6; assay ID:
001973) was used for normalization, and all primers are stated in
Table I. The expression level was
calculated based on the threshold cycle value using the
2−ΔΔCq method (35).
The fold-change of miR-126 expression in patients and carriers was
performed relative to controls.
RT-qPCR
cDNA was synthesized from WBC RNA using Superscript
III, reverse transcriptase and random hexamers (High-Capacity cDNA
Reverse Transcription Kit; cat. no. 4368814; Applied Biosystems;
Thermo Fisher Scientific, Inc.). qPCR using specific primers for
the BRCA1 transcript (Table I) was
performed as described previously (24). β-actin was used as a housekeeping
gene, and the primers for this and BRCA1 are stated in Table I. PCR was performed using the CFX96
Real-Time System (Bio-Rad Laboratories, Inc.) with SYBR Green
(RT2 SYBR Green Fluor qPCR Mastermix; cat. no. 330513;
Qiagen GmbH). The qPCR thermocycling conditions were an initial
cycle at 95°C for 30 sec, followed by 44 cycles at 95°C for 15 sec
and 60°C for 30 sec. The 2−ΔΔCq method was used to
calculate the relative BRCA1 expression. The fold-changes of mRNA
expression were assessed relative to the unmethylated CF females,
for patients with BC and OC, and CF female carriers.
Statistical analysis
Fisher's exact test was performed to determine the
associations between BRCA1 promoter methylation and age,
miR-126 expression and clinicopathological features of BC and OC.
The unpaired t-test was performed to determine the statistical
significance between two groups for gene expression (adult CF
carriers vs. controls and newborns carriers vs. newborn controls).
One-way ANOVA with Dunnett's multiple comparison tests were
performed for comparing multiple groups. GraphPad version 9.1.0
(GraphPad Software, Inc.) was used for all analyses. P<0.05 was
considered to indicate a statistically significant difference.
Results
WBC BRCA1 promoter methylation
Among the 502 patients with BC, 284 were aged <50
years and 218 were aged ≥50 years. There were 57 patients with
methylated BRCA1 in their WBCs, of which 45 were aged <50
years (15.8%) and 12 were aged ≥50 years (5.5%). There was a
significant association between BRCA1 methylation and the
early onset of BC according to Fisher's exact test (P=0.0003;
Table II). The
clinicopathological characteristics of the screened patients with
BC are shown in Table III. For
the BRCA1 methylation–positive patients, the
clinicopathological parameters, other than age, were known for only
49 cases. Notably, 34.7% (17/49) of the methylated cases were TNG
BC, compared to 17.4% (63/363) of unmethylated cases. There was a
significant association between WBC BRCA1 methylation and
TNG BC according to Fisher's exact test (P=0.0066; Table III).
| Table II.Association between BC type 1 gene
promoter methylation in the white blood cells of patients with OC
or early onset BC. |
Table II.
Association between BC type 1 gene
promoter methylation in the white blood cells of patients with OC
or early onset BC.
| Group | Patients with
methylation, n (%) | Patients without
methylation, n (%) | P-value |
|---|
| Control
(n=425) | 40 (9.4) | 385 (90.6) |
|
| BC
(n=502)a | 57 (11.4) | 443 (88.2) | 0.3890 |
| Age,
years |
|
| 0.0003b |
|
<50
(n=284) | 45 (15.8) | 238 (83.8) |
|
|
≥50 (n=218) | 12 (5.5) | 205 (94.0) |
|
| OC (n=187) |
|
| 0.0266b |
| Age,
years |
|
| 0.3000 |
|
<50 (n=70) | 14 (20.0) | 56 (80.0) |
|
|
≥50 (n=117) | 16 (13.7) | 101 (86.3) |
|
| Table III.Clinicopathological features of
screened BC cases. |
Table III.
Clinicopathological features of
screened BC cases.
| Cancer subtype | Patients with
methylation, n (%) | Patients without
methylation, n (%) | ND, n (%) | P-value |
|---|
| BC (n=502) | 57 (11.4) | 443 (88.2) | 2 (0.4) |
|
| TNG
(n=80) | 17 (34.7) | 63 (17.4) |
| 0.0066a |
| IDC
(n=264) | 26 (53.1) | 238 (65.5) |
|
|
| DCIS
(n=19) | 3 (6) | 16 (4.4) |
|
|
|
Other | 3 | 46 |
|
|
| ND | 8 | 80 |
|
|
Among the 187 patients with OC, 70 were aged <50
years and 117 were aged ≥50 years. There were 30 patients with OC
(16%) who tested positive for BRCA1 methylation in their
WBCs, with a significant association between WBC BRCA1
methylation and the incidence of ovarian cancer according to
Fisher's exact test (P=0.0266; Table
II). However, unlike BC, there was no association between
BRCA1 methylation status and the onset of OC [20.0% aged
<50 years (14/70) and 13.7% aged ≥50 years (16/117); P=0.3000].
The clinicopathological characteristics of the screened patients
with OC are shown in Table IV.
Most OC cases were of the serous OC subtype. Notably, 2 patients
were positive for both BC and OC (Table IV).
| Table IV.Clinicopathological features of
screened OC cases. |
Table IV.
Clinicopathological features of
screened OC cases.
| Cancer type | Patients with
methylation, n (%) | Patients without
methylation, n (%) |
|---|
| OC (n=187) | 30 (16.0) | 157 (84.0) |
|
HGSOC | 24 (80.0) | 93 (59.2) |
| OC +
BC | 2 (6.7) | 5 (3.2) |
|
Other | 4 (13.3) | 64 (40.8) |
In addition, among the 425 CF females who were
screened, 9.4% (40/425) tested positive for the methylated
BRCA1 promoter (Table
II).
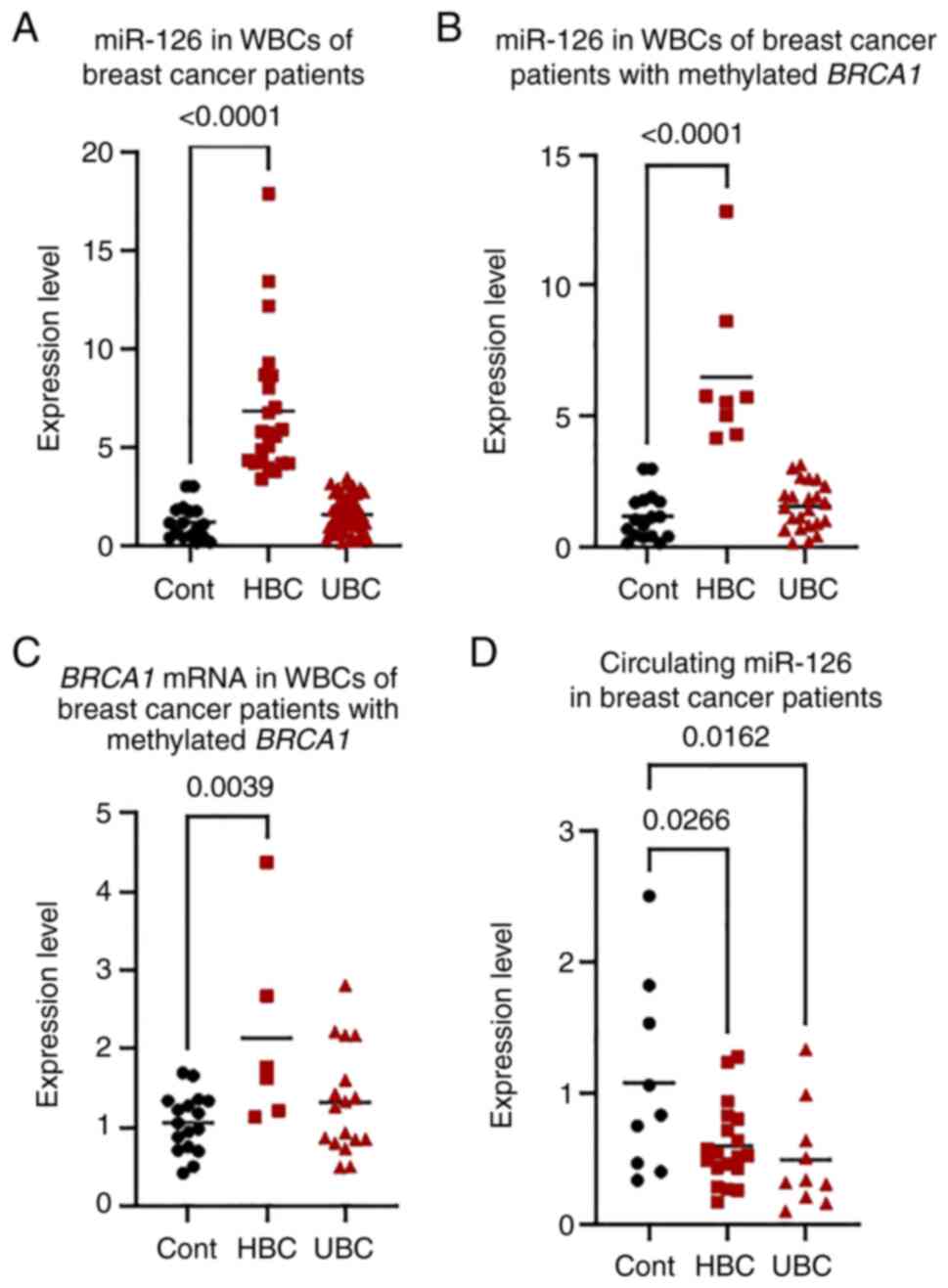
miR-126 in WBCs of patients with
BC
miR-126 was measured in 74 randomly selected
patients with BC (age range, 29–82 years; median age, 47 years), of
which 32 patients were positive for methylated BRCA1 (median
age, 44 years) (Table V). Based on
a cut-off value of +3-fold relative to 17 age-matched female
controls, there were 26 patients (35.1%) who had higher miR-126
expression, up to 18-fold, compared with the control (HBC group),
and 48 patients (64.9%) who had unchanged miR-126 expression (UBC
group) (Fig. 1A). The
clinicopathological features, which were known for only 72
patients, showed that in the HBC group only 1 patient (3.8%) had
distant metastasis compared to 11 patients (24%) in the UBC group.
There was a significant negative association between miR-126
expression in WBC and the risk of distant BC metastasis according
to Fisher's exact test (P=0.0452) (Table V). When the 32 patients with
BRCA1-methylated BC were separately analyzed, similar
results were observed where 9 patients (28%) had higher miR-126
expression, up to 13-fold, compared with the control (HBC group),
and 23 patients (71.9%) had unchanged miR-126 expression (UBC
group) (Fig. 1B). However, the
negative association between miR-126 expression and the risk of
distant BC metastasis was not statistically significant according
to Fisher's exact test (P=0.3742) (Table VI).
| Table V.MicroRNA-126 expression in the white
blood cells of patients with BC (n=74). |
Table V.
MicroRNA-126 expression in the white
blood cells of patients with BC (n=74).
| Cancer subtype | HBC (n=26) | UBC (n=48) | P-value |
|---|
| Methylated
BRCA1 | 9 (34.6) | 23 (47.9) |
|
| TNG | 5 (19.2) | 11 (23.9) |
|
| IDC | 20 (76.9) | 39 (84.8) |
|
| DCIS | 4 (15.4) | 3 (6.5) |
|
| Metastasis | 1 (3.8) | 11 (23.9) | 0.0452a |
| ND | 0 (0.0) | 2 (4.2) |
|
| Table VI.MicroRNA-126 expression in the white
blood cells of patients with BC type 1 gene methylated BC
(n=32). |
Table VI.
MicroRNA-126 expression in the white
blood cells of patients with BC type 1 gene methylated BC
(n=32).
| Cancer subtype | HBC (n=9) | UBC (n=23) | P-value |
|---|
| TNG | 3 (33.3) | 9 (42.9) |
|
| IDC | 8 (88.9) | 19 (90.5) |
|
| DCIS | 1 (11.1) | 1 (4.7) |
|
| Metastasis | 1 (11.1) | 7 (33) | 0.3742 |
| ND | 0 (0.0) | 2 (4.3) |
|
mRNA in WBCs of patients with BC
When BRCA1 mRNA was measured in the WBCs of
the patients with BRCA1-methylated BC, expression was
significantly higher in the HBC group by up to 4-fold compared with
the control group (P=0.0039). However, the UBC group did not
significantly differ from the control group (P=0.4400) (Fig. 1C).
Circulating miR-126 in patients with
BC
When miR-126 expression was measured in the plasma
of patients with BC, there was less circulating miR-126 in the HBC
(P=0.0260) and UBC (P=0.0160) groups compared with that in
age-matched controls (Fig.
1D).
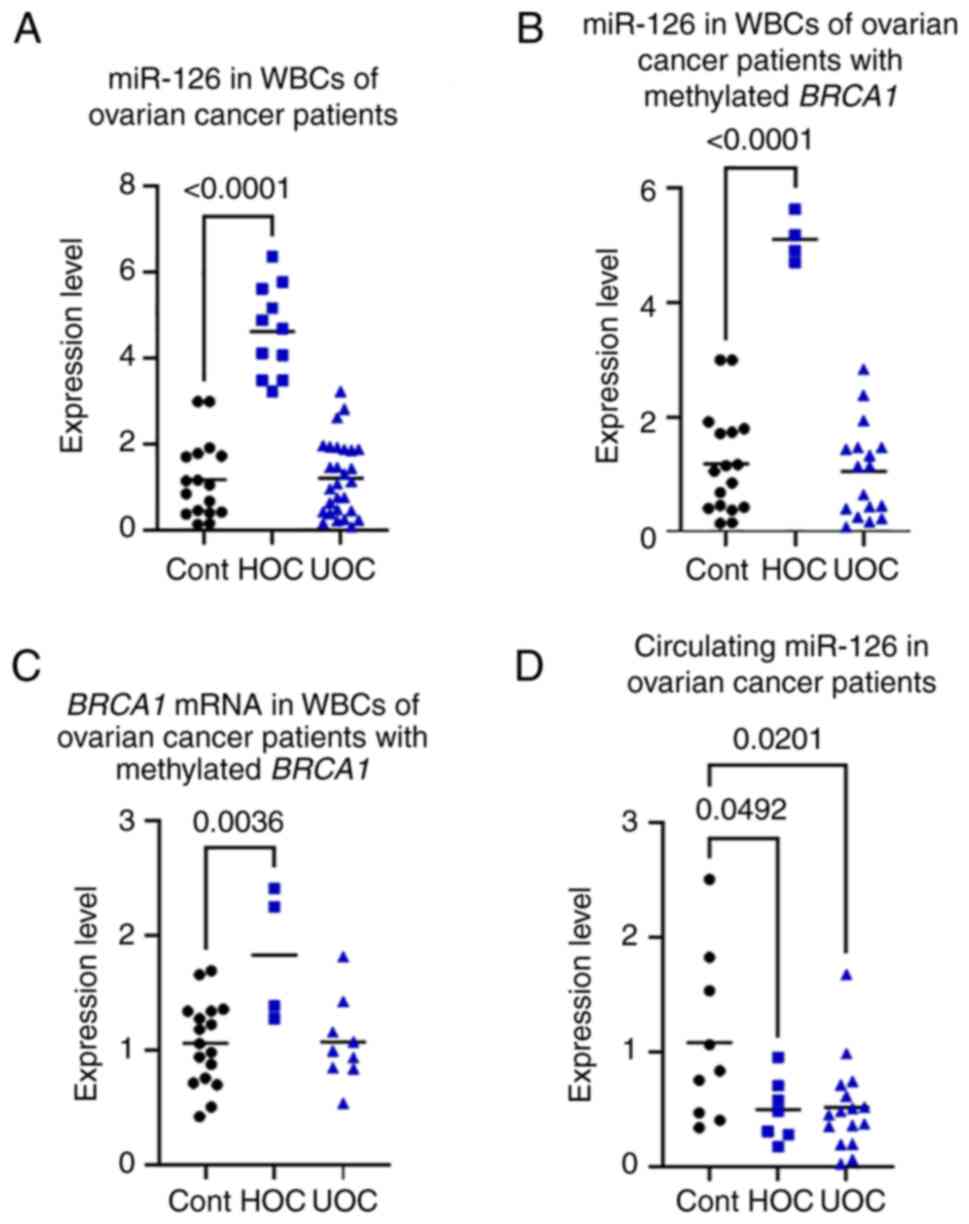
miR-126 in WBCs of patients with
OC
miR-126 expression in WBCs was measured in 46
randomly selected patients with OC (age range, 19–88 years; median
age, 51.5 years), of which 21 were positive for methylated
BRCA1 (median age, 50 years) (Table VII). Similar to patients with BC,
based on a cut-off value of +3-fold relative to 17 age-matched
female controls, there were 11 patients (23.9%) who had higher
miR-126 expression (HOC group), up to 7-fold, compared with the
control, and 35 patients (76.1%) who had unchanged ImiR-126
expression (UOC group) (Fig. 2A;
Table VII). In the HOC group, 8
patients (72.7%) had disease progression and 6 (54.5%) died,
compared with the UOC group, where 10 patients (28.6%) had disease
progression and 3 (8.57%) died. There was a significant positive
association between the miR-126 expression in WBCs and the risk of
OC disease progression and death according to Fisher's exact test
(P=0.0029) (Table VII).
| Table VII.MicroRNA-126 expression in the white
blood cells of patients with OC (n=46). |
Table VII.
MicroRNA-126 expression in the white
blood cells of patients with OC (n=46).
| Cancer outcome | HOC (n=11) | UOC (n=35) | P-value |
|---|
| Methylated
BRCA1 | 4 (36.4) | 17 (48.6) |
|
| No recurrence | 3 (27.3) | 25 (71.4) | 0.0138a |
| Progression | 8 (72.7) | 10 (28.6) |
|
| Death | 6 (54.5) | 3 (8.6) | 0.0029a |
When the 21 patients with BRCA1-methylated OC
were analyzed separately, similar results were found, with 4
patients (19%) exhibiting higher miR-126 expression (HOC group), up
to 6-fold, compared with the control, and 17 patients (81%) with
unchanged miR-126 expression (UOC group) (Fig. 2B;). However, the association
between miR-126 expression and the risk of OC disease progression
and death was not statistically significant according to Fisher's
exact test (P=0.0797) (Table
VIII).
| Table VIII.MicroRNA-126 expression in the white
blood cells of patients with breast cancer type 1 gene methylated
OC (n=21). |
Table VIII.
MicroRNA-126 expression in the white
blood cells of patients with breast cancer type 1 gene methylated
OC (n=21).
| Cancer outcome | HOC (n=4) | UOC (n=17) | P-value |
|---|
| No recurrence | 1 (25.0) | 11 (64.7) |
|
| Progression | 3 (75.0) | 6 (35.3) |
|
| Death | 2 (50.0) | 1 (5.9) | 0.0797 |
mRNA in WBCs of patients with OC
When BRCA1 mRNA was measured in the WBCs of
the patients with BRCA1-methylated OC, expression was
significantly higher by up to 2.5-fold in the HOC group compared
with that in the control group (P=0.0056). However, the UOC group
did not significantly differ from the control group (P=0.1120)
(Fig. 2C).
Circulating miR-126 in OC
patients
When miR-126 expression was measured in the plasma
of patients with OC, there was less circulating miR-126 in the HOC
(P=0.0110) and UOC (P=0.0170) groups compared with that in
age-matched controls (Fig.
2D).
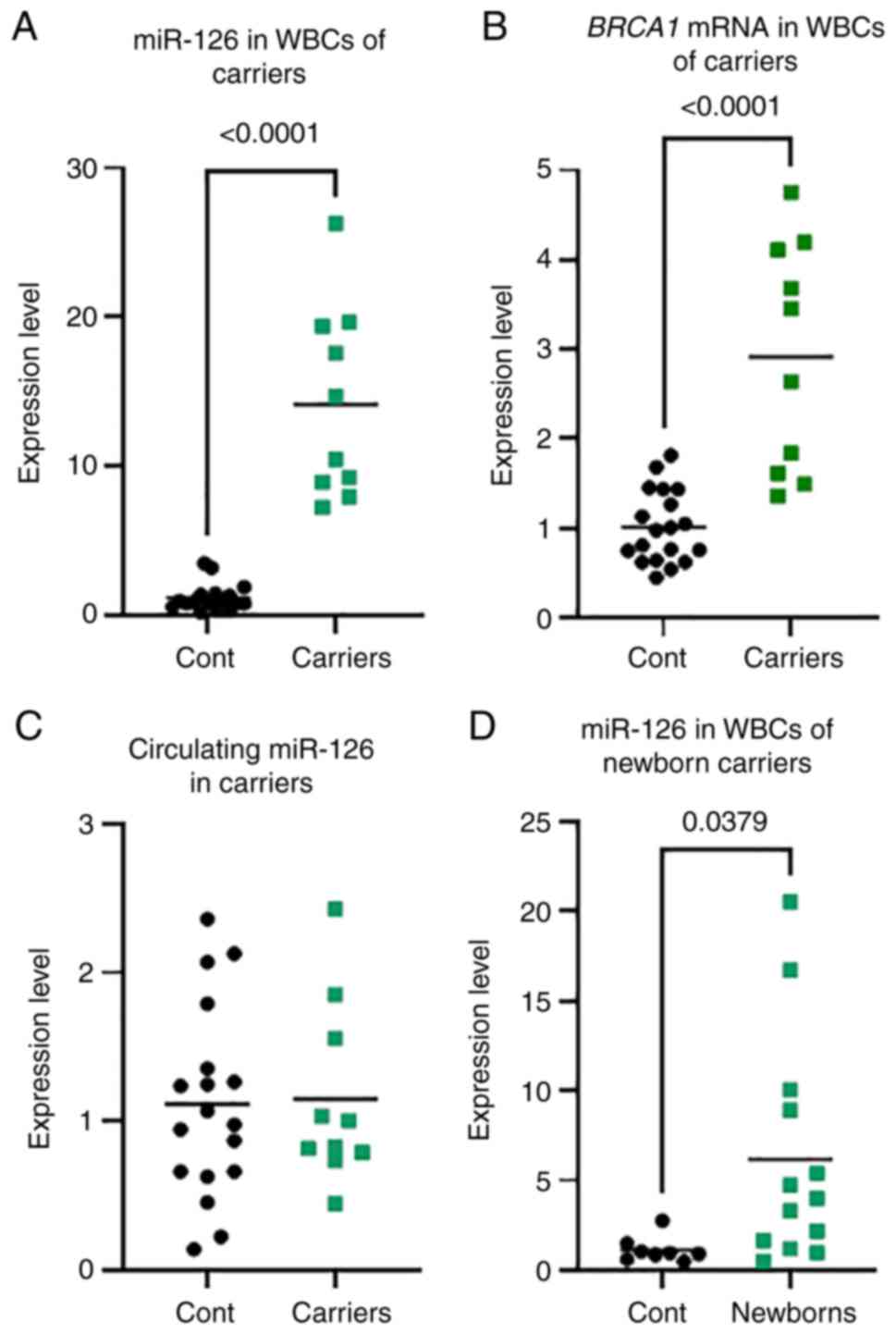
miR-126 is elevated in WBCs but
unchanged in the plasma of CF BRCA1-methylated female carriers
To further appreciate the use of miR-126 as a
biomarker for the early prediction of BC and OC, miR-126 levels
were measured in the WBC and plasma from 10 BRCA1 methylated
CF female carriers (age range, 18–27 years; median age, 20 years).
A greatly increased level of miR-126 expression, up to 27-fold
higher, was noted in the WBCs of all the carriers compared with
that in the age-matched control group. This result is similar to
the miR-126 expression in the WBCs from the patients with BC and OC
(Fig. 3A). In contrast to that in
the patients with BC and OC, there was no change in the level of
circulating miR-126 in the carrier group compared with the control
group (Fig. 3C). This result
revealed that, in the CF BRCA1-methylated carriers, miR-126
is altered in WBCs but not in the plasma.
mRNA in WBCs of CF carriers
Similar to the results in patients with BC and OC,
when BRCA1 mRNA was measured in the WBCs of the CF
BRCA1 methylation carriers, expression was significantly
higher, up to 20-fold, in the carrier group compared with that in
the age-matched control group (P<0.0001) (Fig. 3B).
miR-126 is elevated in the WBCs of CF
BRCA1-methylated newborn female carriers. As
BRCA1 promoter methylation is detectable from early on in
life in carriers (22,24), miR-126 was measured in the WBCs of
13 BRCA1-methylated newborn female carriers. A significantly
higher miR-126 expression level, up to 20-fold higher, was observed
in the newborn carrier group compared with that in the newborn
non-carrier control group (P=0.0391) (Fig. 3D). This result indicates an
alteration in miR-126 from early on in the life of the
carriers.
Discussion
There is a well-established association between
BRCA1 promoter methylation in peripheral blood cells and the
risk of BC and OC (20,22,23,36).
However, less is known about its role in the cancer risk of CF
BRCA1-methylated females. In the present study, BRCA1
promoter methylation was detected in peripheral WBCs in 9.4% of 425
CF female controls. This result agrees with our previous findings
(16,21). The detection of methylated
BRCA1 promoter in CF females raises the question of whether
those individuals are at increased risk of developing BC or OC
later in life. In the present study, miR-126 was investigated as a
potential molecular biomarker for predicting BC and OC risk in CF
BRCA1-methylated females.
In total, 502 patients with BC were screened for
BRCA1-methylation in peripheral WBCs. BRCA1 promoter
methylation was significantly associated with early onset BC
(P=0.0003), which agrees with the results of previous studies
(16,17,21,36).
Typically, TNG BC accounts for 10–20% of all BC cases (37). In the BRCA1 methylated BC
cases in the present study, TNG BC accounted for 34% compared with
17.4% of the unmethylated BC cases, which indicates enrichment of
TNG BC in BRCA1 methylation-positive cases. This finding
agrees with a recent study (18),
which investigated the association between the BRCA1
promoter methylation in peripheral blood and the risk of TNG BC.
The study reported that 30.2% of the TNG BC cases exhibited a
methylated BRCA1 gene status in the peripheral blood cells,
indicating a significant association between constitutional
BRCA1 promoter methylation and TNG BC (18).
Furthermore, in the present study, 187 patients with
OC were screened for methylated BRCA1. A significant
association was found between WBC methylated BRCA1 and OC
incidence (P=0.0266). The vast majority of the methylated OC cases
were HGSOC. These results agree with previously reported data,
where it was shown that the frequency of BRCA1 methylation
in WBCs was higher in patients with OC than in controls and that it
was associated with risk for HGSOC (22).
miR-126 is downregulated in tumor tissues compared
with that in normal adjacent tissues in several cancer types,
including BC, OC and colorectal cancer, and this decrease is
associated with higher malignancy tumor grade and metastasis
(31,32,38).
The restoration of miR-126 has been demonstrated to inhibit
metastasis properties in these cancer types (32,38).
By contrast, it has been reported that in BC,
miR-126 is increased significantly in DCIS tissue compared with
that in IDC and normal adjacent tissue. In addition, the
downregulation of miR-126 is associated with the later onset of IDC
(39). These findings suggest that
non-invasive tumor cells inside DCIS may counteract the progression
to an invasive lesion by increasing the level of miR-126 expression
(39). In the present study, the
results revealed a significant association between an increase in
miR-126 expression in WBCs and a lower risk of distant metastasis
in patients with BRCA1-methylated and unmethylated BC.
In patients with OC, the expression of miR-126 in
WBCs was significantly associated with a higher risk of disease
progression and death in the present study. These results suggest
that miR-126 is a dual-functional miRNA, functioning as a tumor
suppressor in BC and as an oncogene in OC, which may indicate
different targets and mechanisms of action in the two types of
cancer. Indeed, it was previously reported that the abundance of
miRNAs and their targets could contribute to their contradictory
roles in cancer (40). Notably, as
WBCs are considered normal cells with the body tissues, the present
findings might suggest an increase in miR-126 in the whole body.
However, further studies are needed to address these findings.
Similar to the findings in patients with BC and OC,
increased miR-126 expression was observed in the WBCs of CF
BRCA1-methylated carriers compared with that in age-matched
controls. Unlike that in patients with cancer, miR-126 expression
was not decreased in any of the carriers. Notably, the increase in
miR-126 appears to occur from early on in the life of the carriers,
as significant upregulation of miR-126 expression was observed in
newborn female carriers. As BRCA1 epimutation is present
from early on in the life of the carriers (24), we hypothesize that the increase in
miR-126 could be a protective mechanism activated by the whole
body, from the start of life, as a response to the epigenetically
altered cancer suppressor gene, BRCA1. This claim is
supported by the increase in BRCA1 mRNA expression in the
patients and carriers despite the methylation status of the
BRCA1 promoter. It has been reported that some
hypermethylated genes are overexpressed due to the interaction with
other factors (41). Based on the
present findings, it is tempting to speculate that the increase in
BRCA1 mRNA, which occurs from early on in the life of the
carriers (24), could be the
result of the interaction with other factors regulated by miR-126.
The inevitable activation of the carcinogenic mechanisms derived by
the constitutional methylation of the BRCA1 promoter, such
as genomic instability, gross chromosomal rearrangements and
generation of mutations, may result in the loss of miR-126 that
leads to carcinogenesis (42). We
therefore hypothesize that the upregulation of miR-126 could be
part of a mechanism linking constitutional BRCA1 promoter
methylation with the pathological consequences induced by this
epigenetic defect. Indeed, it has been reported that miR-126 is
reversibly increased in response to short-term exposure to asbestos
as a defensive process activating detoxifying mechanisms (42). However, long-lasting asbestos
exposure results in the irreversible downregulation of miR-126 due
to asbestos-induced DNA damage. The reduction in miR-126 activates
the IRS1/PI3K/AKT pathway leading to the development of malignant
mesothelioma (42). Thus, the
expression of miR-126 links asbestos exposure to malignant
mesothelioma.
Circulating miR-126 has been reported to
differentiate patients with cancer from controls (43). In the present study, decreased
circulating miR-126 was observed in the patients with BC and OC
compared with that in healthy controls, regardless of its level in
the WBCs, which agrees with previously reported data (27,29).
Notably, the fact that there was no derease in the level of
circulating miR-126 in the CF BRCA1 methylated carriers,
despite the increase in the WBCs, suggests the use of miR-126 as a
prognosticator for BC and OC risk for those carriers.
The present study has certain limitations. For
example, the physiological association between miR-126 and BRCA1 in
WBCs has not been explored. Further studies are needed to
investigate if there is any transcriptional regulation between
BRCA1 and miR-126. Additionally, future studies are needed to
search for the different targets of miR-126 in BC and OC cancer
that contribute to its contradictory actions in these two types of
cancer.
In conclusion, the present study revealed the likely
involvement of miR-126 in the constitutional methylation of
BRCA1 promoter-related malignancies. Significant
upregulation was observed in the level of miR-126 in WBCs, not only
in patients with BC and OC, but also in CF BRCA1 methylated
carriers. Overall, the increase in miR-126 could be a mechanism
activated by the body in response to the aberrantly methylated
cancer-suppressor gene BRCA1, which has different
pathological consequences according to cancer type.
Acknowledgements
The authors would like to thank Dr Ayodele Alaiya
(Department of Stem Cells, King Faisal Specialist Hospital and
Research Centre, Riyadh, Saudi Arabia) for providing help in
collecting the clinical data of the patients.
Funding
This project was funded by the King Faisal Specialist Hospital
and Research Centre.
Availability of data and materials
All data generated or analyzed during this study are
available from the corresponding author on reasonable request.
Authors' contributions
MAS, NAY and NAM performed the data analysis. MAS,
WA, SA and AA contributed to the sample and data collection. OA and
HA permitted sample collection and contributed to data acquisition.
NAM conceived and designed the study and drafted the manuscript.
All authors read and approved the final manuscript. OA, HA and AA
confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
Ethical approval (approval no. RAC #2170017) was
obtained by the Human Research Ethics Committee of King Faisal
Specialist Hospital and Research Centre. All participants provided
written informed consent. The guardian of the patient provided
written informed consent for participants <18 years old.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BC
|
breast cancer
|
|
BRCA1
|
breast cancer type 1 gene
|
|
CF
|
cancer-free
|
|
miR-126
|
microRNA-126
|
|
OC
|
ovarian cancer
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
WBC
|
white blood cell
|
References
|
1
|
McCormick B, Winter K, Hudis C, Kuerer HM,
Rakovitch E, Smith BL, Sneige N, Moughan J, Shah A, Germain I, et
al: RTOG 9804: A prospective randomized trial for good-risk ductal
carcinoma in situ comparing radiotherapy with observation. J Clin
Oncol. 33:709–715. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Corben AD: Pathology of invasive breast
disease. Surg Clin North Am. 93:363–392. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Duraker N, Hot S, Akan A and Nayir PO: A
comparison of the clinicopathological features, metastasis sites
and survival outcomes of invasive lobular, invasive ductal and
mixed invasive ductal and lobular breast carcinoma. Eur J Breast
Health. 16:22–31. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cho KR and Shih IM: Ovarian cancer. Annu
Rev Pathol. 4:287–313. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chan JK, Cheung MK, Husain A, Teng NN,
West D, Whittemore AS, Berek JS and Osann K: Patterns and progress
in ovarian cancer over 14 years. Obstet Gynecol. 108((3 Pt 1)):
521–528. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jacinto FV and Esteller M: Mutator
pathways unleashed by epigenetic silencing in human cancer.
Mutagenesis. 22:247–253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Futreal PA, Liu Q, Shattuck-Eidens D,
Cochran C, Harshman K, Tavtigian S, Bennett LM, Haugen-Strano A,
Swensen J and Miki Y: BRCA1 mutations in primary breast and ovarian
carcinomas. Science. 266:120–122. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Birgisdottir V, Stefansson OA,
Bodvarsdottir SK, Hilmarsdottir H, Jonasson JG and Eyfjord JE:
Epigenetic silencing and deletion of the BRCA1 gene in sporadic
breast cancer. Breast Cancer Res. 8:R382006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Esteller M, Silva JM, Dominguez G, Bonilla
F, Matias-Guiu X, Lerma E, Bussaglia E, Prat J, Harkes IC, Repasky
EA, et al: Promoter hypermethylation and BRCA1 inactivation in
sporadic breast and ovarian tumors. J Natl Cancer Inst. 92:564–569.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Butcher DT and Rodenhiser DI: Epigenetic
inactivation of BRCA1 is associated with aberrant expression of
CTCF and DNA methyltransferase (DNMT3B) in some sporadic breast
tumours. Eur J Cancer. 43:210–219. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sudbø J, Reith A and Lingjaerde OC:
Gene-expression profiles in hereditary breast cancer. N Engl J Med.
344:20292001.PubMed/NCBI
|
|
12
|
Veer LJ, Dai H, van de Vijver MJ, He YD,
Hart AA, Mao M, Peterse HL, van der Kooy K, Marton MJ, Witteveen
AT, et al: Gene expression profiling predicts clinical outcome of
breast cancer. Nature. 415:530–536. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sahnane N, Carnevali I, Formenti G,
Casarin J, Facchi S, Bombelli R, Di Lauro E, Memoli D, Salvati A,
Rizzo F, et al: BRCA methylation testing identifies a subset of
ovarian carcinomas without germline variants that can benefit from
PARP inhibitor. Int J Mol Sci. 21:97082020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang YQ, Yan Q, Zhang JR, Li SD, Yang YX
and Wan XP: Epigenetic inactivation of BRCA1 through promoter
hypermethylation in ovarian cancer progression. J Obstet Gynaecol
Res. 39:549–554. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Snell C, Krypuy M, Wong EM; kConFab
investigators, ; Loughrey MB and Dobrovic A: BRCA1 promoter
methylation in peripheral blood DNA of mutation negative familial
breast cancer patients with a BRCA1 tumour phenotype. Breast Cancer
Res. 10:R122008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Al-Moghrabi N, Al-Qasem AJ and Aboussekhra
A: Methylation-related mutations in the BRCA1 promoter in
peripheral blood cells from cancer-free women. Int J Oncol.
39:129–135. 2011.PubMed/NCBI
|
|
17
|
Wong EM, Southey MC, Fox SB, Brown MA,
Dowty JG, Jenkins MA, Giles GG, Hopper JL and Dobrovic A:
Constitutional methylation of the BRCA1 promoter is specifically
associated with BRCA1 mutation-associated pathology in early-onset
breast cancer. Cancer Prev Res (Phila). 4:23–33. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Prajzendanc K, Domagala P, Hybiak J, Rys
J, Huzarski T, Szwiec M, Tomiczek-Szwiec J, Redelbach W, Sejda A,
Gronwald J, et al: BRCA1 promoter methylation in peripheral blood
is associated with the risk of triple-negative breast cancer. Int J
Cancer. 146:1293–1298. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gupta S, Jaworska-Bieniek K, Narod SA,
Lubinski J, Wojdacz TK and Jakubowska A: Methylation of the BRCA1
promoter in peripheral blood DNA is associated with triple-negative
and medullary breast cancer. Breast Cancer Res Treat. 148:615–622.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Iwamoto T, Yamamoto N, Taguchi T, Tamaki Y
and Noguchi S: BRCA1 promoter methylation in peripheral blood cells
is associated with increased risk of breast cancer with BRCA1
promoter methylation. Breast Cancer Res Treat. 129:69–77. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Al-Moghrabi N, Nofel A, Al-Yousef N,
Madkhali S, Amer SM, Alaiya A, Shinwari Z, Al-Tweigeri T, Karakas
B, Tulbah A and Aboussekhra A: The molecular significance of
methylated BRCA1 promoter in white blood cells of cancer-free
females. BMC Cancer. 14:8302014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lønning PE, Berge EO, Bjørnslett M,
Minsaas L, Chrisanthar R, Høberg-Vetti H, Dulary C, Busato F,
Bjørneklett S, Eriksen C, et al: White blood cell BRCA1 promoter
methylation status and ovarian cancer risk. Ann Intern Med.
168:326–334. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jung Y, Hur S, Liu J, Lee S, Kang BS, Kim
M and Choi YJ: Peripheral blood BRCA1 methylation profiling to
predict familial ovarian cancer. J Gynecol Oncol. 32:e232021.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Al-Moghrabi N, Al-Showimi M, Al-Yousef N,
Al-Shahrani B, Karakas B, Alghofaili L, Almubarak H, Madkhali S and
Al Humaidan H: Methylation of BRCA1 and MGMT genes in white blood
cells are transmitted from mothers to daughters. Clin Epigenetics.
10:992018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bosviel R, Garcia S, Lavediaux G, Michard
E, Dravers M, Kwiatkowski F, Bignon YJ and Bernard-Gallon DJ: BRCA1
promoter methylation in peripheral blood DNA was identified in
sporadic breast cancer and controls. Cancer Epidemiol.
36:e177–e182. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tomasetti M, Gaetani S, Monaco F, Neuzil J
and Santarelli L: Epigenetic regulation of miRNA expression in
malignant mesothelioma: MiRNAs as biomarkers of early diagnosis and
therapy. Front Oncol. 9:12932019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tavazoie SF, Alarcon C, Oskarsson T, Padua
D, Wang Q, Bos PD, Gerald WL and Massague J: Endogenous human
microRNAs that suppress breast cancer metastasis. Nature.
451:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Meister J and Schmidt MHH: MiR-126 and
miR-126*: New players in cancer. ScientificWorldJournal.
10:2090–2100. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bockmeyer CL, Christgen M, Müller M,
Fischer S, Ahrens P, Länger F, Kreipe H and Lehmann U: MicroRNA
profiles of healthy basal and luminal mammary epithelial cells are
distinct and reflected in different breast cancer subtypes. Breast
Cancer Res Treat. 130:735–745. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang F, Zheng Z, Guo J and Ding X:
Correlation and quantitation of microRNA aberrant expression in
tissues and sera from patients with breast tumor. Gynecol Oncol.
119:586–593. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, Qin X, Jiang J and Zhao W:
MicroRNA-126 exerts antitumor functions in ovarian cancer by
targeting EGFL7 and affecting epithelial-to-mesenchymal transition
and ERK/MAPK signaling pathway. Oncol Lett. 20:1327–1335. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pan C, Stevic I, Muller V, Ni Q,
Oliveira-Ferrer L, Pantel K and Schwarzenbach H: Exosomal microRNAs
as tumor markers in epithelial ovarian cancer. Mol Oncol.
12:1935–1948. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wyman SK, Parkin RK, Mitchell PS, Fritz
BR, O'Briant K, Godwin AK, Urban N, Drescher CW, Knudsen BS and
Tewari M: Repertoire of microRNAs in epithelial ovarian cancer as
determined by next generation sequencing of small RNA cDNA
libraries. PLoS One. 4:e53112009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Azzollini J, Pesenti C, Pizzamiglio S,
Fontana L, Guarino C, Peissel B, Plebani M, Tabano S, Sirchia SM,
Colapietro P, et al: Constitutive BRCA1 promoter hypermethylation
can be a predisposing event in isolated early-onset breast cancer.
Cancers (Basel). 11:582019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nakai K, Hung MC and Yamaguchi H: A
perspective on anti-EGFR therapies targeting triple-negative breast
cancer. Am J Cancer Res. 6:1609–1623. 2016.PubMed/NCBI
|
|
38
|
Zhang Y, Wang X, Xu B, Wang B, Wang Z,
Liang Y, Zhou J, Hu J and Jiang B: Epigenetic silencing of miR-126
contributes to tumor invasion and angiogenesis in colorectal
cancer. Oncol Rep. 30:1976–1984. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Volinia S, Bertagnolo V, Grassilli S,
Brugnoli F, Manfrini M, Galasso M, Scatena C, Mazzanti CM, Lessi F,
Naccarato G, et al: Levels of miR-126 and miR-218 are elevated in
ductal carcinoma in situ (DCIS) and inhibit malignant potential of
DCIS derived cells. Oncotarget. 9:23543–23553. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xiang Y, Tian Q, Guan L and Niu SS: The
dual role of miR-186 in cancers: Oncomir battling with tumor
suppressor miRNA. Front Oncol. 10:2332020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Orgueira AM: Hidden among the crowd:
Differential DNA methylation-expression correlations in cancer
occur at important oncogenic pathways. Front Genet.
6:1632015.PubMed/NCBI
|
|
42
|
Andersen M, Grauslund M, Ravn J, Sorensen
JB, Andersen CB and Santoni-Rugiu E: Diagnostic potential of
miR-126, miR-143, miR-145, and miR-652 in malignant pleural
mesothelioma. J Mol Diagn. 16:418–430. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yan J, Ma S, Zhang Y, Yin C, Zhou X and
Zhang G: Potential role of microRNA-126 in the diagnosis of
cancers: A systematic review and meta-analysis. Medicine
(Baltimore). 95:e46442016. View Article : Google Scholar : PubMed/NCBI
|