Introduction
Angiogenesis, the process of forming new capillaries
from existing blood vessels, is a physiological process essential
for embryonic development and tissue repair (1). It is also involved in the transition
of premalignant lesions to malignant tumors and is regarded as a
hallmark of metastasis (2). Highly
proliferative cancer cells in solid tumor masses require expansion
of vascular networks to transport oxygen and essential nutrients,
leading to the production and secretion of angiogenic factors
(3). Vascular endothelial growth
factor (VEGF), also known as VEGF-A, is considered the major
mediator of angiogenesis in certain types of cancer in humans,
including glioblastoma (GB), breast, colon and lung cancers
(4). In addition, VEGF also serves
a key role in the proliferation, migration, invasion and
capillary-like tube formation of endothelial cells (5). VEGF binds to and activates the
high-affinity transmembrane receptor tyrosine kinases, VEGF
receptor (VEGFR)1 (also known as Flt-1) and VEGFR2 (also known as
KDR/Flk-1) (6). VEGFR2 has a high
affinity for VEGF and is the major effector of VEGF-induced
proangiogenic signaling in endothelial cells (6). The binding of VEGF to VEGFR2 triggers
receptor dimerization and autophosphorylation of intracellular
domains, resulting in the activation of downstream signal
transduction mediators, including PI3K/AKT/mTOR and MAPK/ERK
(7). Subsequently, certain key
intracellular signaling molecules, such as vascular cell adhesion
molecule-1 (VCAM-1), intercellular adhesion molecule-1 (ICAM-1) and
matrix metalloproteinases (MMPs), that are responsible for
endothelial cell proliferation, migration and invasion, are
produced and activated (8,9). Therefore, blockade of
VEGF/VEGFR2-mediated signal transduction has been reported to be an
effective anti-angiogenic strategy against certain cancers
(4–6).
According to the Central Brain Tumor Registry of the
United States Statistical Report in the United States in 2012-2016,
gliomas are the among the most common intracranial neoplastic
diseases, accounting for >80% of malignant tumors of the central
nervous system (10). GB,
categorized as grade IV astrocytoma by the World Health
Organization, is the most common and malignant brain tumor and the
overall incidence rate in elderly patients (≥65 years) was 13.16
per 100,000 in the United States from 2000 to 2017 (11). Even if the patient receives standard
treatment, including surgical resection, chemotherapy and
radiotherapy, the median survival is 12–15 months, and the 5-year
survival rate is <5% (12). A
significant characteristic of GB is a high degree of angiogenesis
due to the release of proangiogenic factors that promote tumor
vasculature development, including VEGF, transforming growth factor
β, nitric oxide, proteolytic enzymes and proangiogenic chemokines
(13). Previous studies have
correlated VEGF expression with glioma grade and prognosis
(14,15). Therefore, angiogenesis induced by
VEGF is one of the main targets of GB therapy (13).
Essential oils extracted from Cedrus species
have traditionally been used in clinical applications of
aromatherapy for musculoskeletal, genitourinary and skin systems
(16). Previous studies have
demonstrated that extracts from Cedrus atlantica possess
anticancer, antibacterial and antihyperalgesic effects (17–20).
Cedrol, one of the active ingredients in oils extracted from C.
atlantica, is a natural crystalline sesquiterpene alcohol with
numerous pharmacological activities, including antioxidant,
anti-inflammatory, analgesic, antimicrobial, sedative and
anticancer effects (21–23) Although our previous studies
demonstrated that cedrol suppressed GB growth by inducing DNA
damage, cell cycle arrest and apoptosis in vitro and in
vivo (24,25), the anti-angiogenic role of cedrol
remains unclear. Thus, in the present study, the mechanisms
underlying the anti-angiogenic effects of cedrol on VEGF-induced
angiogenesis in human umbilical vein endothelial cells (HUVECs)
were analyzed.
Materials and methods
Agents and antibodies
Cedrol was purchased from Tokyo Chemical Industry
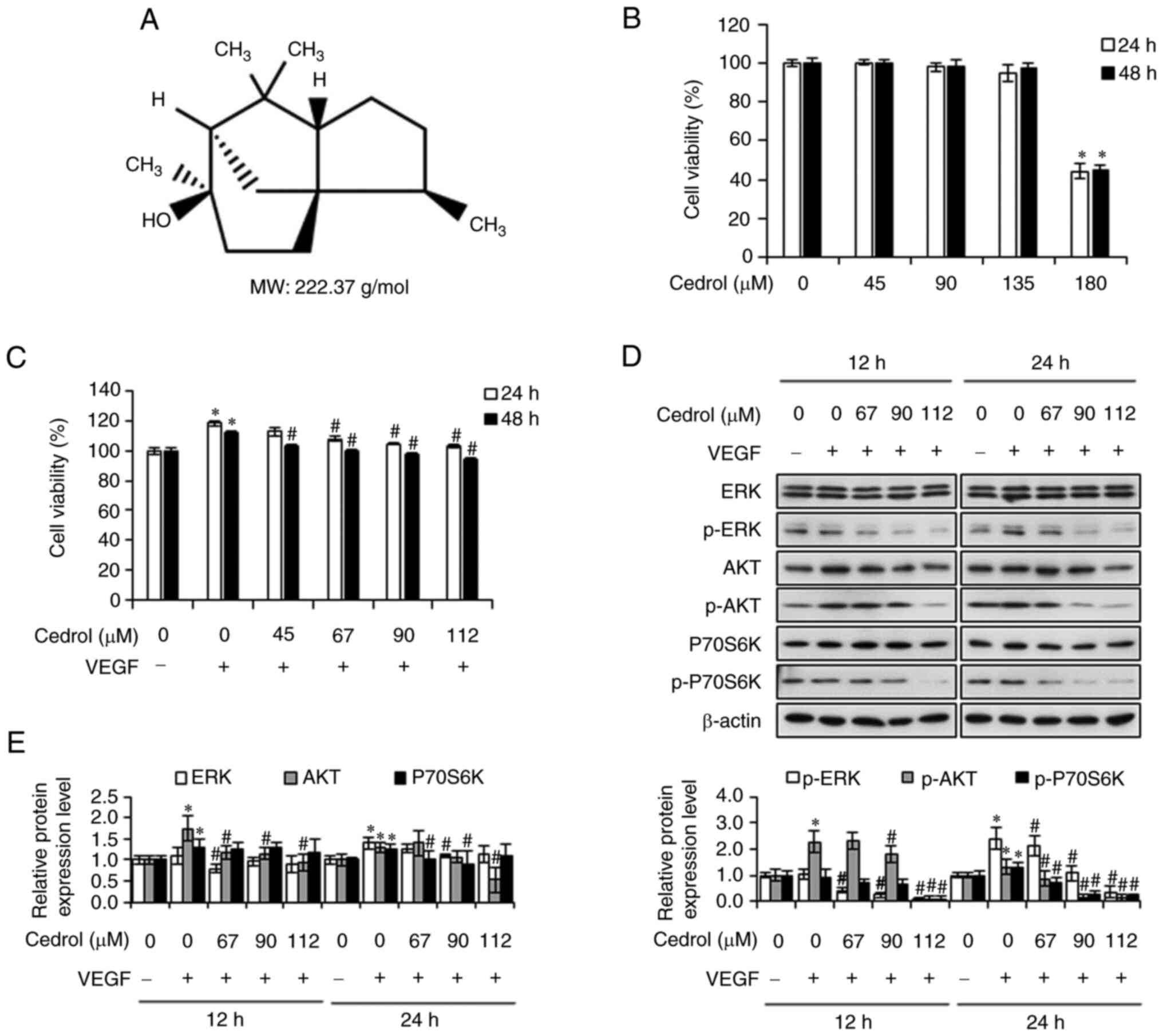
Co., Ltd., and its chemical structure, with a molecular weight of
222.37 g/mol, is presented in Fig.
1A. Recombinant human VEGF165 was purchased from
PeproTech, Inc. Primary antibodies, such as anti-P70S6K (cat. no.
SC-8418), anti-p-P70S6K (cat. no. SC-8416), anti-VEGF (cat. no.
SC-152), anti-VEGFR2 (cat. no. SC-6251) and MMP-9 (SC-6840) were
purchased from Santa Cruz Biotechnology (all 1:250 dilution; Santa
Cruz Biotechnology, Inc.). The antibody against p-VEGFR2 was
purchased from Biorbyt (1:1,000; cat. no. orb159521; Biorbyt Ltd).
Specific antibodies against ERK (cat. no. IR187-705), p-ERK (cat.
no. IR188-706), AKT (IR171-666), p-AKT (IR172-668), CD31 (IR218-1),
VCAM-1 (IR79-272), ICAM-1 (IR78-268) and β-actin (IR2-7) were
obtained from iReal Biotechnology, Inc. (all 1:1,000).
Cell culture
HUVECs (CRL-1730) and human GB cells (DBTRG-05MG)
were purchased from the American Type Culture Collection. HUVECs
were cultured in M199 medium supplemented with 10% heat-inactivated
fetal bovine serum, endothelial cell growth supplement (30 µg/ml;
MilliporeSigma), heparin (20 U/ml; MilliporeSigma), L-glutamine (2
mM), penicillin-streptomycin mixture (100 U/ml penicillin and 100
µg/ml streptomycin), sodium pyruvate (1 mM) and HEPES buffer
solution (10 mM). DBTRG-05MG cells were cultured in Roswell Park
Memorial Institute (RPMI)-1640 medium supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.),
penicillin-streptomycin mixture (100 U/ml penicillin and 100 µg/ml
streptomycin), sodium pyruvate (1 mM) and HEPES buffer solution (10
mM). Cells were incubated at 37°C in a humidified atmosphere of 5%
CO2. All other reagents were purchased from Gibco
(Thermo Fisher Scientific, Inc.), unless otherwise noted.
Cell viability assay
Cell viability was determined using an MTT assay.
The 0.2% gelatin solution (MilliporeSigma) in PBS was added in
96-well culture plates and incubated at 37°C overnight. After
removing gelatin solution and washing with PBS, 5×103
cells were seeded in gelatin-coated 96-well culture plates and
allowed to adhere overnight. When cells attained 50–60% confluence,
they were treated with cedrol dissolved in dimethyl sulfoxide
(DMSO; final concentration <0.5%) with or without VEGF (20
ng/ml) for 24 and 48 h. After the medium was replaced with the MTT
solution (400 µg/ml; Amresco, LLC), the cells were incubated for
4–6 h. The formed formazan crystals was dissolved in DMSO and
detected using a Spectra Max Plus 384 Microplate Reader (Molecular
Devices, LLC) at a wavelength of 550 nm. Cell viability in the
control group was considered to be 100%. Each assay was performed
in triplicate.
Western blotting
For whole protein extraction, cells were harvested,
washed twice with PBS, and lysed in RIPA buffer (Bio Basic, Inc.)
containing protease inhibitors (Amresco, LLC) and phosphatase
inhibitors (Bionovas Biotechnology Co., Ltd.) for 30 min on ice.
After centrifugation at 14,000 × g for 15 min at 4°C, the proteins
in the supernatant were quantified using the BCA Protein Assay Kit
(Pierce; Thermo Fisher Scientific, Inc.) and then stored at −20°C.
Cell lysates (20 µg/lane) were separated by 8-12.5% SDS-PAGE and
transferred to polyvinylidene fluoride (PVDF) membranes (Pall Life
Sciences). The blots were blocked with 5% skim milk at 25°C for 1 h
and immunolabeled with primary antibodies at 4°C overnight. After
three rounds of washing with 0.5% Tween-20 in TBS, membranes were
incubated with biotin-conjugated secondary antibody (Santa Cruz
Biotechnology, Inc.) at 25°C for 2 h, followed by
peroxidase-conjugated streptavidin (Jackson ImmunoResearch
Laboratories, Inc.) for 1 h at 25°C. The immunoreactive bands were
visualized using an enhanced chemiluminescence reagent (T-Pro
Biotechnology), and grayscale analysis was performed using ImageJ
software (version 1.8.0; National Institutes of Health). β-actin
was used as a loading control.
Scratch wound healing assays
HUVECs were seeded onto gelatin-coated six-well
plates, as aforementioned, at a density of 5×105
cells/well in complete medium (10% FBS) and cultured to 90%
confluence. After treatment with 5 µg/ml mitomycin C
(MedChemExpress) at 37°C for 2 h, the monolayer was wounded using a
200 µl pipette tip to create cross scratches and washed with PBS to
remove non-adherent cells. After replacing complete medium with
low-serum medium (2% FBS), the cells were treated with cedrol
(0–112 µM) and VEGF (20 ng/ml) and incubated at 37°C for up to 12
h. Images of the area of migrated cells in the scratch were
captured at time intervals of 0, 6 and 12 h under an inverted
microscope and quantified using the ImageJ software (version 1.8.0;
National Institutes of Health).
Boyden chamber assay
Cell migration and invasion assays were performed in
a 48-well Boyden chamber. Briefly, the upper and lower chambers
were separated using nitrocellulose filters (pore size, 8 µm; GVS
North America, Inc.) incubated with 0.2% gelatin (MilliporeSigma)
dissolved in PBS at 37°C overnight and washed with PBS. The upper
surface of filters was coated with or without 20 µl Matrigel (0.5
mg/ml; Corning Corp.) and incubated for 30 min at 37°C for gelling.
Next, 5×104 cells in serum-free medium were placed in
the upper chambers, and the bottom chamber was filled with M199
complete medium containing 10% FBS, cedrol (0–112 µM) and VEGF (20
ng/ml). After incubation at 37°C for 24 h, the cells on the upper
surface were removed by wiping with cotton swabs and the cells on
the lower surface of the membrane were fixed and stained with 0.1%
crystal violet at 25°C for 10 min. After the membrane was dried,
cells were counted in five independent areas per membrane using a
bright-field microscope to determine the migration and invasion
capacity of cedrol-treated cells.
Capillary-like tube formation
assay
To prepare the thin collagen layer, 10 µl of
Matrigel was poured into wells of µ-Slide 15 Well 3D (Ibidi GmbH)
and incubated at 37°C for 30 min to solidify gels. HUVECs were
plated on µ-slides at a density of 6×103 cells/well in
M199 serum-free medium containing cedrol (0–90 µM) and VEGF (20
ng/ml). After incubation at 37°C for 3 h, the formation of
tubular-like structures were observed and images captured using an
inverted microscope and quantified by counting the total number of
branch points in five randomly selected fields of view to evaluate
the anti-angiogenic capacity of cedrol.
Semi-quantitative reverse
transcription-polymerase chain reaction (RT-PCR)
Total RNA from cells subjected to treatment with
cedrol (67, 90 and 112 µM) and VEGF (20 ng/ml) at 37°C for 12 and
24 h was extracted using RareRNA reagent (Genepure), and cDNA was
converted using a HiSpec Reverse Transcriptase kit (Yeastern
Biotech Co., Ltd.), following the manufacturer's instructions. PCR
amplification was performed on a Thermo Cycler PX2 PCR instrument
(Thermo Fisher Scientific, Inc.) using single-stranded cDNA,
specific primers (listed in Table
I; Mission Biotech), 5X Taq PCR MasterMix (Biomate) and
distilled H2O. The total reaction system volume was 20
µl and the following thermocycling conditions were used for PCR:
Initial denaturation at 95°C for 10 min; 30 cycles of 95°C for 30
sec, 55–56°C for 30 sec, 72°C for 30 sec; and a final extension at
72°C for 10 min. PCR products were electrophoresed on 2% agarose
gels, stained with ethidium bromide, and imaged using a UV
transilluminator (Alpha Innotech). The quantification of each band
was performed by ImageJ software (version 1.8.0; National
Institutes of Health). GAPDH was used as an internal control.
 | Table I.Primers used for semi-quantitative
RT-PCR reactions. |
Table I.
Primers used for semi-quantitative
RT-PCR reactions.
| Gene | Primer sequence
(5′→3′) | Annealing
temperature |
|---|
| VEGF | F:
CGCTCGGTGCTGGAATTTGA | 56°C |
|
| R:
AGTGGGGAATGGCAAGCAAA |
|
| KDR | F:
GTGATCGGAAATGACACTGGAG | 56°C |
|
| R:
CATGTTGGTCACTAACAGAAGCA |
|
| PECAM1 | F:
AACAGTGTTGACATGAAGAGCC | 55°C |
|
| R:
TGTAAAACAGCACGTCATCCTT |
|
| VCAM-1 | F:
TCAGATTGGAGACTCAGTCATGT | 56°C |
|
| R:
ACTCCTCACCTTCCCGCTC |
|
| ICAM-1 | F:
GGCCGGCCAGCTTATACAC | 55°C |
|
| R:
TAGACACTTGAGCTCGGGCA |
|
| MMP-9 | F:
TATGACATCCTGCAGTGCCC | 55°C |
|
| R:
TTGTATCCGGCAAACTGGCT |
|
| GADPH | F:
GAGTCAACGGATTTGGTCGT | 56°C |
|
| R:
GACAAGCTTCCCGTTCTCAG |
|
Tumor cell-induced angiogenesis
assay
DBTRG-05MG cells were plated, cultured to 90%
confluence and treated with cedrol (0–90 µM) in serum-free medium
at 37°C for 24 h. Conditioned media (CM) from DBTRG-05MG cells in
each culture condition was used for the tube formation assays.
HUVECs were plated on µ-slides at a density of 6×103
cells/well in CM from DBTRG-05MG cells, incubated at 37°C for 3 h,
and observed the formation of tubular-like structures as
aforementioned. Following washing with PBS and centrifugation at
300 × g at 4°C for 10 min, pellets of DBTRG-05MG cells were
collected and mRNA and protein expression were analyzed by
semi-quantitative RT-PCR and western blotting, respectively,
according to the aforementioned methods.
Statistical analysis
All values are presented as the mean ± standard
deviation of at least three independent experiments and
statistically significant differences were determined by one-way
ANOVA using SPSS software (version 22.0, IBM Corp.) followed by
Tukey post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Cedrol inhibits VEGF-induced
proliferation of HUVECs
To determine the appropriate treatment dose of
cedrol with no cytotoxic effects for angiogenesis assays, a range
of concentrations of cedrol were applied to HUVECs for 24 and 48 h
and cell viability was measured by MTT assay. Cedrol reduced the
viability of HUVECs with an IC50 value of 178.50±3.76 µM
(24 h) and 179.66±8.71 µM (48 h) and demonstrated no significant
cytotoxic effect on HUVECs at concentrations up to 135 µM (Fig. 1B). Therefore, the dose of cedrol
used in the present study was <135 µM for the in vitro
angiogenesis assays. The proliferation of endothelial cells is key
in the multi-step process of angiogenesis. The effects of a range
of concentrations of cedrol on VEGF-induced proliferation of HUVECs
were examined. Treatment of HUVECs with VEGF for 24 and 48 h
significantly increased the cell viability to 118.44±1.87 and
112.04±1.12% compared with the control, however treatment of HUVECs
with cedrol significantly inhibited VEGF-induced cell proliferation
in a concentration-dependent manner (Fig. 1C). The MAPK/ERK and mTOR/PI3K/AKT
signaling pathways can be stimulated by VEGF-VEGFR2 attachment and
are known regulators of cell survival and proliferation (26). To understand the molecular mechanism
of the cedrol-mediated anti-proliferative properties, the protein
expression levels of ERK/p-ERK, AKT/p-ATK and P70S6K/p-P70S6K were
analyzed using western blotting. Treatment with cedrol
significantly downregulated VEGF-induced phosphorylation of ERK,
AKT and P70S6K proteins in a dose-dependent manner (Fig. 1D and E), suggesting that the
inhibitory effect of cedrol on HUVECs is mediated through the
MAPK/ERK and mTOR/PI3K/AKT signaling pathways. Taken together,
these results suggest that cedrol may act as a potent inhibitor of
VEGF-induced signaling pathways in endothelial cells.
Cedrol suppresses VEGF-induced
migration and invasion of HUVECs
Since cell migration and invasion are critical steps
in angiogenesis, the inhibitory activities of cedrol on
VEGF-induced migration and invasion of HUVECs were examined using a
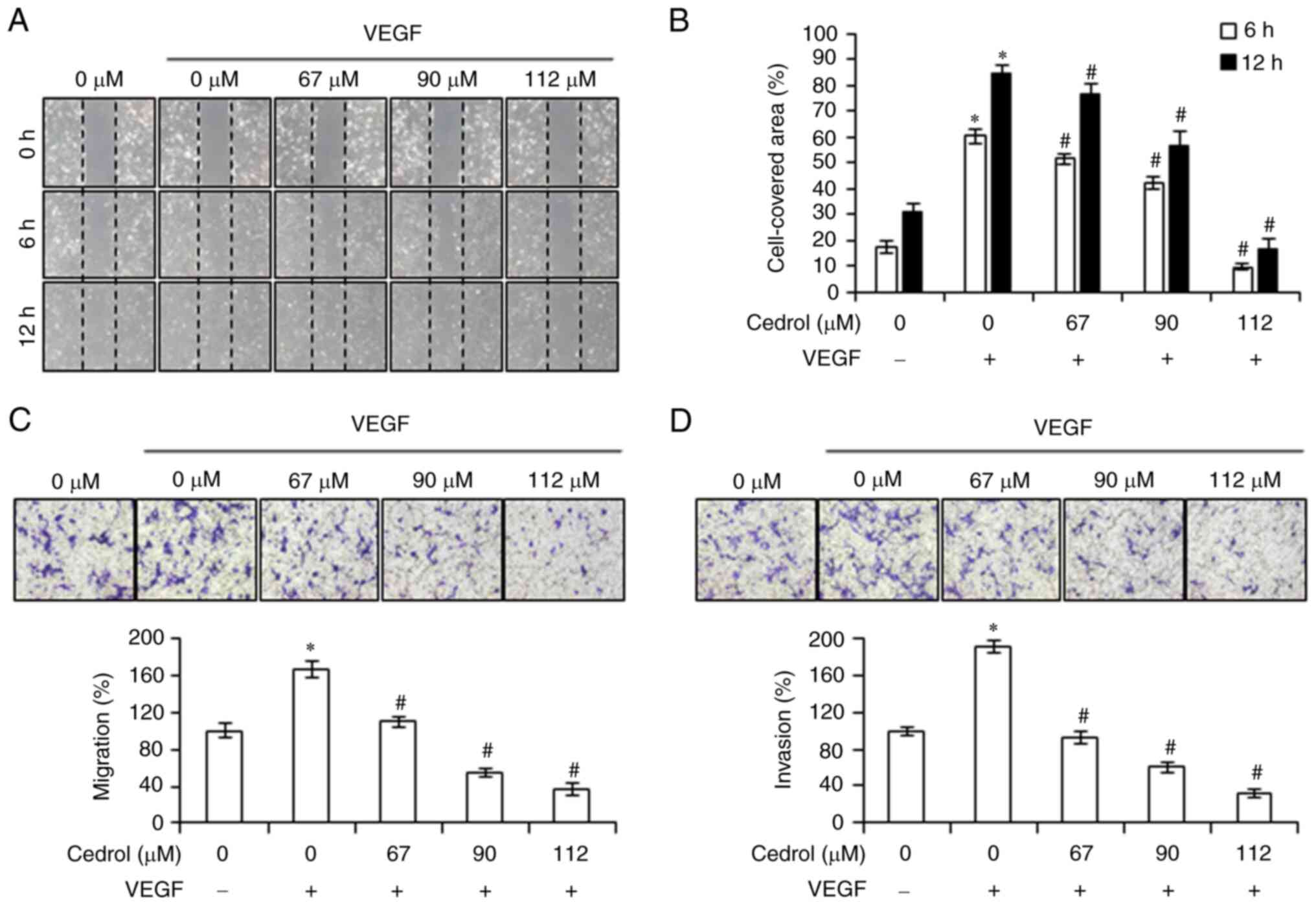
wound healing assay or Boyden chamber assay. As demonstrated by the
wound healing assay, the cell-covered area in the VEGF group was
84.73±2.69% and treatment with 67, 90 and 112 µM cedrol
significantly decreased the migration of HUVECs after 12 h to
76.43±3.77, 56.82±5.68 and 16.71±3.67%, respectively, compared with
the control (Fig. 2A and B).
Similarly, the results of Boyden chamber assay demonstrated that
the average percentage of migrated and invasive cells significantly
reduced from 167.03±9.21 and 191.60±6.62% for VEGF-stimulated
HUVECs to 109.75±5.20, 54.80±4.91 and 36.38±6.02% (Fig. 2C); 92.02±6.28, 60.40±5.84 and
31.91±4.57% (Fig. 2D) for 67, 90
and 112 µM cedrol-treated HUVECs, respectively. Thus, these results
demonstrated that VEGF-induced migration and invasion of HUVECs
were markedly inhibited by cedrol in a dose-dependent manner.
Cedrol prevents VEGF-induced
capillary-like tube formation of HUVECs
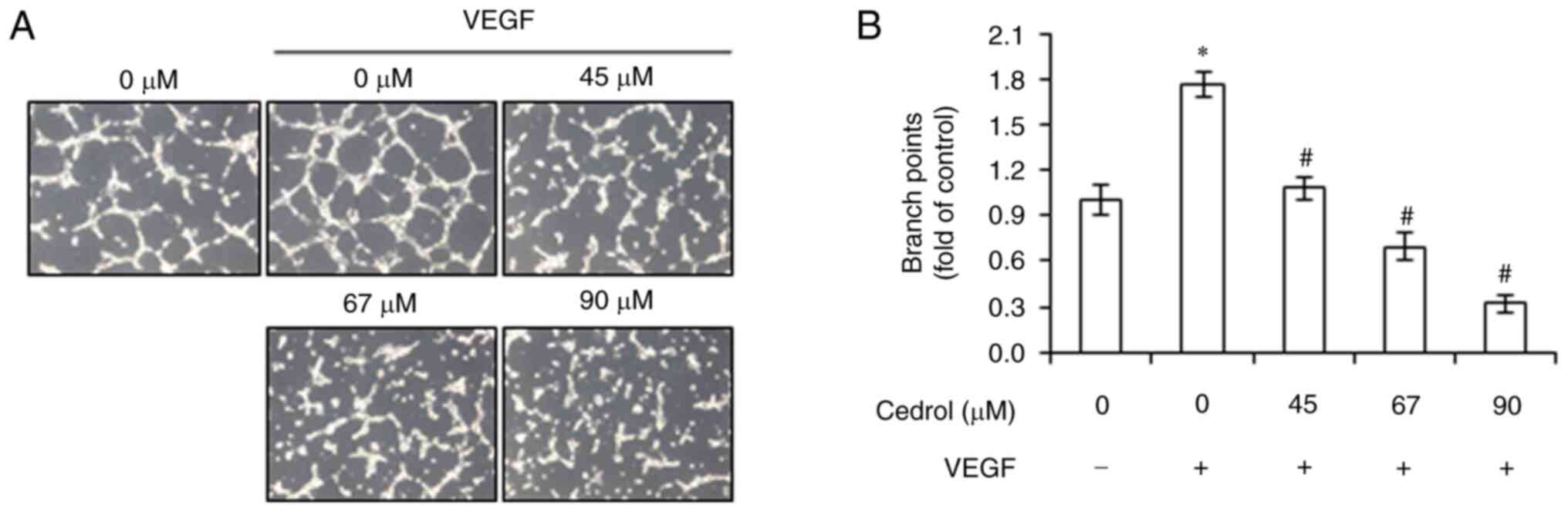
Serum-starved HUVECs are able to spontaneously
arrange themselves in a capillary-like tube formation stimulated by
VEGF when cultured on Matrigel matrix, which is another essential
step in angiogenesis (27).
Therefore, the effects of cedrol on VEGF-induced tube formation in
HUVECs were investigated. HUVECs were plated on the surface of the
Matrigel and treated with cedrol (0–90 µM) and VEGF (20 ng/ml) for
3 h. The results demonstrated that elongated and robust tubule-like
networks were formed in the VEGF group, whereas disorganized
structures were observed in the cedrol exposure groups (Fig. 3A). After statistical analysis, the
fold-change of branch points in the VEGF group reached 1.77±0.08
fold, compared with the control. The cedrol group displayed a
significant concentration-dependent reduction of branch points at
45, 67 and 90 µM with 1.08±0.08, 0.69±0.09 and 0.32±0.06 fold
number of branches, compared with the control, respectively
(Fig. 3B). These results
demonstrated that cedrol interfered with VEGF-induced
capillary-like tube formation in HUVECs.
Cedrol regulates expression of
angiogenesis and adhesion molecules
VEGF, one of the most important regulators of
angiogenesis, induces autophosphorylation of VEGFR2 in endothelial
cells, thereby activating numerous downstream signaling substrates,
involved in processes such as cell proliferation, migration and
tube formation (7). The effects of
cedrol on VEGFR2 signaling pathways in HUVECs were investigated
using semi-quantitative RT-PCR and western blotting. VEGF treatment
significantly increased the mRNA and protein expression levels of
KDR/VEGFR2 and the endothelial marker PECAM1/CD31, as well as
phosphorylation of VEGFR2 protein, which was downregulated upon
treatment with cedrol (Fig. 4). In
addition, as expected, mRNA and protein expression levels of
adhesion molecules, ICAM-1 and VCAM-1, were significantly increased
by VEGF treatment, which were both significantly reduced by cedrol
treatment in a concentration-dependent manner. During angiogenesis,
MMPs) are vital for the degradation of the extracellular matrix
(28). The present study also
assessed the mechanism of the invasive ability by evaluating the
mRNA and protein expression levels of MMP-9. Cedrol significantly
suppressed the expression of endothelial cell-derived MMP-9
stimulated by VEGF in a dose-dependent manner (Fig. 4). Collectively, these findings
demonstrated that cedrol prevented migration, invasion and
angiogenesis by blocking VEGFR2-mediated downstream signaling
cascades in HUVECs.
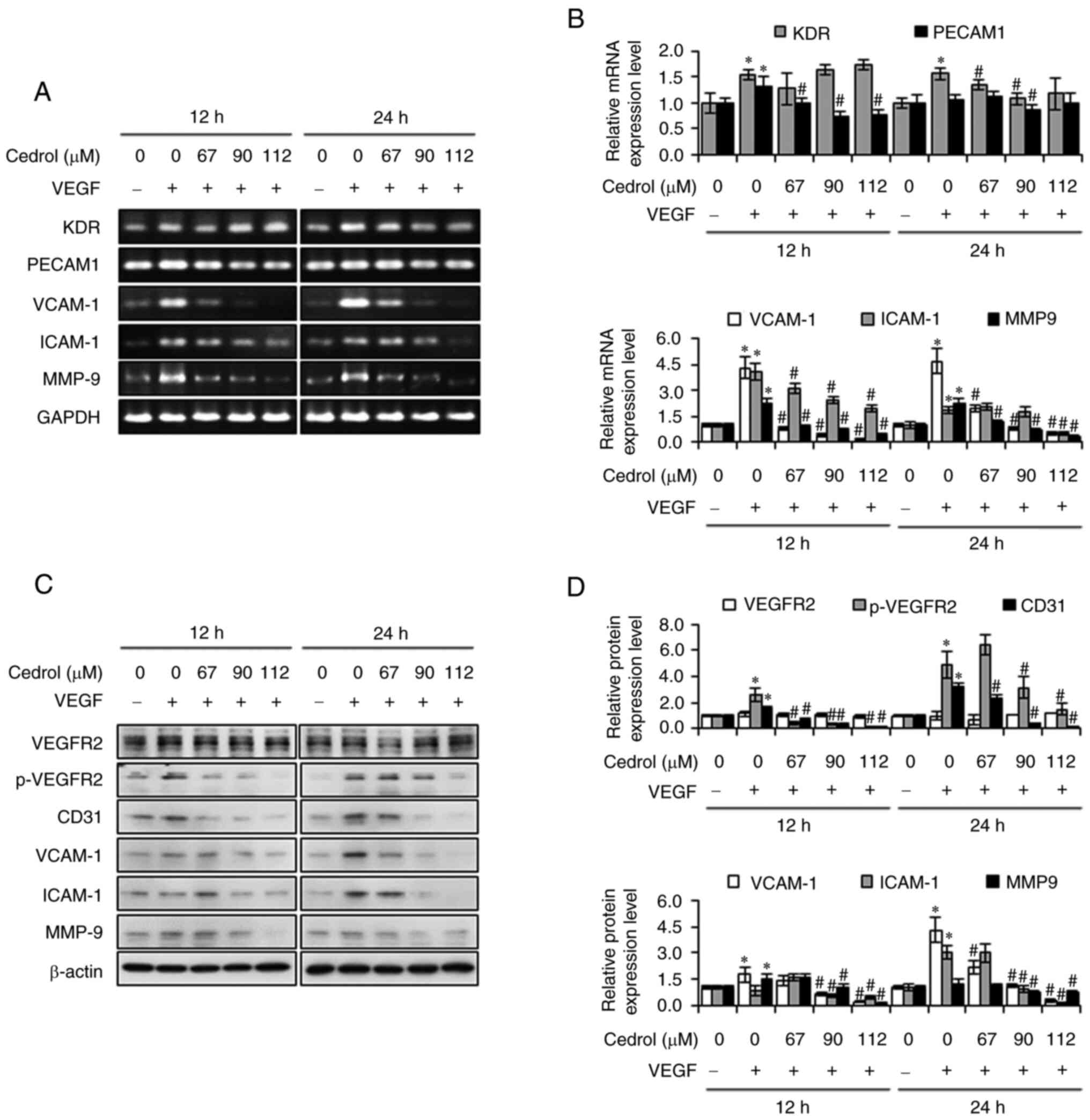 | Figure 4.Effects of cedrol on expression of
angiogenesis and adhesion molecules in HUVECs. HUVECs were treated
with cedrol (0–112 µM) and VEGF (20 ng/ml) for 12 and 24 h and
total cell lysates were subjected to (A and B) semi-quantitative
RT-PCR and (C and D) western blotting to evaluate expression levels
of mRNA and proteins, respectively. The levels of GAPDH or β-actin
were used as an internal control. Data are presented as the mean ±
standard deviation from three independent experiments. *P<0.05
vs. control group. #P<0.05 vs. VEGF group. VEGF,
vascular endothelial growth factor. VEGF, vascular endothelial
growth factor; KDR, kinase insert domain receptor; PECAM1, platelet
and endothelial cell adhesion molecule 1; VCAM-1, vascular cell
adhesion molecule 1; ICAM-1, intracellular adhesion molecule 1;
MMP-9, matrix metallopeptidase 9; GADPH, glyceraldehyde 3-phosphate
dehydrogenase; p, phosphorylated. |
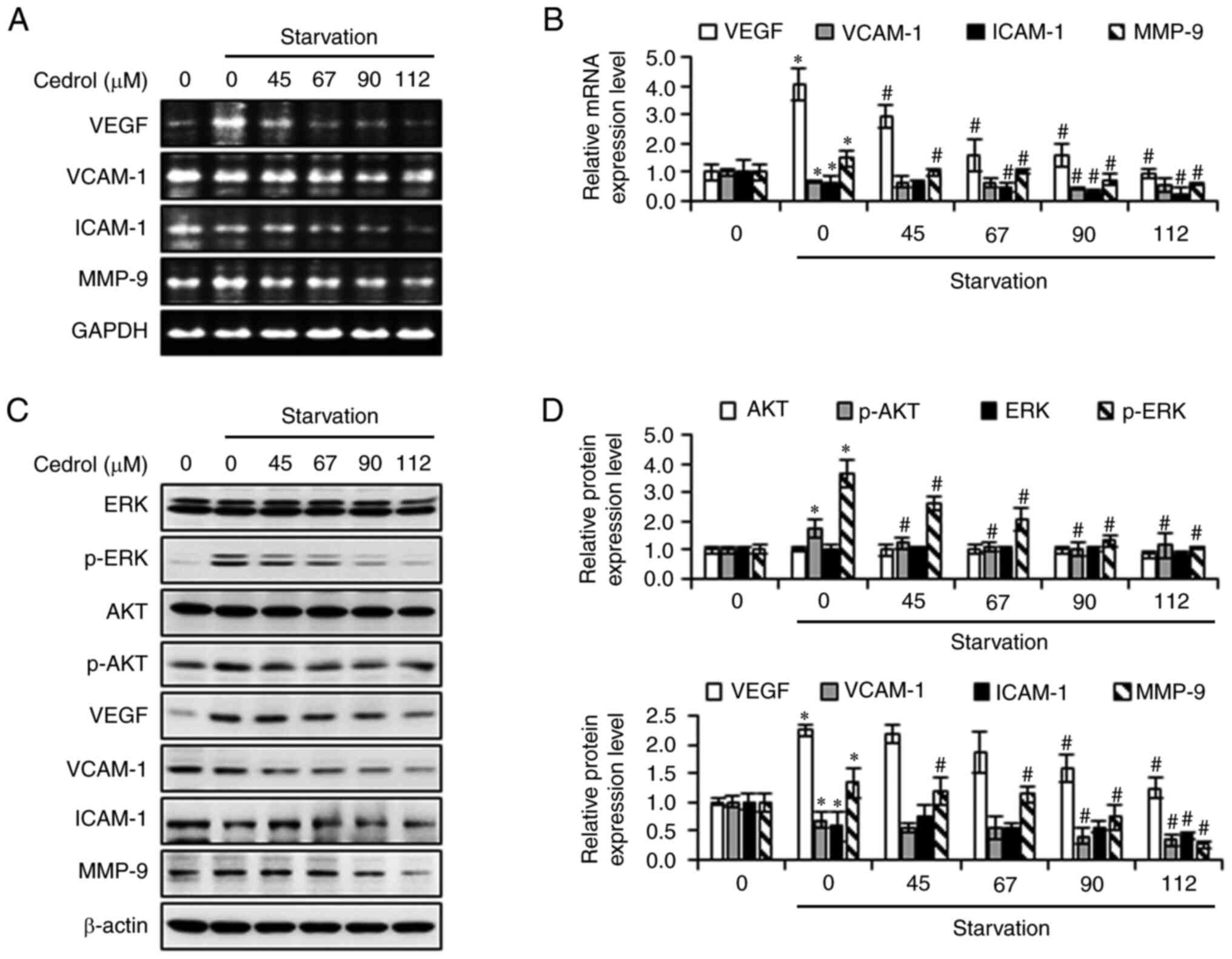
Cedrol suppresses tumor cell-induced
angiogenesis
Angiogenesis serves a crucial role in tumor growth
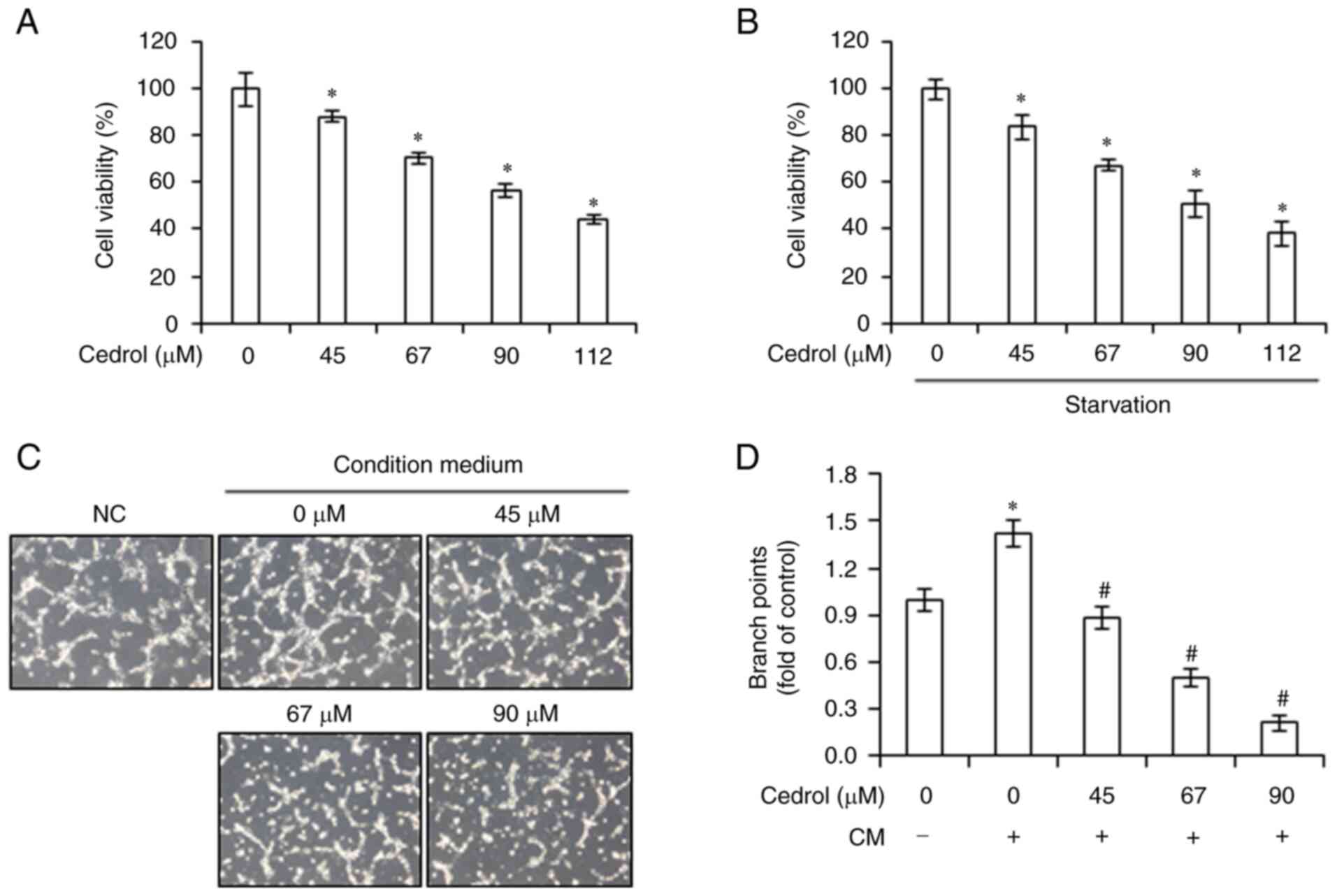
and metastasis. First, the effects of cedrol on the viability of
human GB DBTRG-05MG cells were determined by MTT assay. Cedrol, in
10% FBS medium or serum-free medium, significantly decreased the
viability of DBTRG-05MG cells and demonstrated IC50
values at 24 h of 101.55±3.05 and 93.72±4.78 µM, respectively
(Fig. 5A and B). To investigate the
effect of cedrol on the angiogenic potential of GB, the CM of
DBTRG-05MG cells cultured in serum-free medium with or without
cedrol was used in an in vitro angiogenesis assay. Compared
with the control (medium cultured without tumor cells), CM
significantly activated capillary-like tube formation in HUVECs,
whereas DBTRG-05MG cells treated with cedrol demonstrated
significantly attenuated tumor cell-stimulated development of
tubule-like networks in a dose-dependent manner (Fig. 5C). Diminishing relative branch
points from 1.42±0.08 (CM group) to 0.88±0.07, 0.50±0.06 and
0.21±0.05 were produced in 45, 67 and 90 µM cedrol-treated groups,
respectively (Fig. 5D). To explore
the anti-angiogenesis mechanism of cedrol in tumor cells, the
expression of VEGF mRNA and protein was assessed using
semi-quantitative RT-PCR and western blot analysis. Serum
starvation induced VEGF mRNA and protein expression, which was
significantly downregulated upon treatment with cedrol (Fig. 6). In addition, cedrol significantly
reduced the expression of p-ERK, p-AKT, VCAM-1, ICAM-1 and MMP-9 in
starved cells. These results suggested that cedrol inhibited VEGF
expression in tumor cells stimulated by nutrient deficiency, which
resulted in the attenuation of the angiogenic ability of
HUVECs.
Discussion
Angiogenesis has emerged as an attractive
therapeutic target because of its critical role in several
diseases, such as cancer, neovascular age-related macular
degeneration and diabetic retinopathy (29). Anti-angiogenic therapy using
angiogenesis inhibitors, including bevacizumab, sunitinib,
sorafenib and pazopanib, has previously been reported (30). However, previous clinical studies
have reported that these inhibitors have insufficient efficacy in
blocking the complex biological processes involved in angiogenesis
and tumor development and induce possible accompanying side effects
such as bleeding, hypertension, gastrointestinal perforation,
hypothyroidism, vomiting, diarrhea and skin toxicity (31). Therefore, there is still a need to
develop new anti-angiogenic drugs to improve treatment efficacy of
diseases, such as cancer, neovascular age-related macular
degeneration and diabetic retinopathy.
In the present study, the potent anti-angiogenic
activity of cedrol, a natural sesquiterpene alcohol isolated from
C. atlantica, was evaluated. It has previously been reported
that cedrol exhibits antioxidant, anti-inflammatory, antimicrobial,
analgesic, sedative and anticancer activities (21–25).
Cedrol attenuates rheumatoid arthritis symptoms by blocking the
phosphorylation of ERK/MAPK and p65/NF-κB signaling pathways in
LPS-mediated fibroblast-like synoviocytes and suppresses
pro-survival signaling in human cancer cells by inhibiting proteins
in the PI3/AKT/mTOR/ERK1/2 and NF-κB signaling pathways (32,33).
Moreover, our previous studies demonstrated that cedrol suppressed
the growth of GB via downregulation of mTOR/AKT/P70S6K (24,25).
Here, expression of VEGF, VEGFR2 and vessel marker CD31 in tumor
tissues was decreased after cedrol treatment. In VEGFR2-dependent
angiogenesis, PI3K/AKT and ERK signaling pathways serve important
roles in the proliferation and survival of endothelial cells
(26). These previous results
suggested that cedrol may have high anti-angiogenic potential. The
results of the present study demonstrated that cedrol significantly
inhibited the major angiogenic phenotypes, including proliferation,
migration, invasion and tube formation of VEGF-stimulated HUVECs at
low or non-toxic concentrations. Furthermore, these results
demonstrated that the VEGFR2 signaling pathway in endothelial cells
and VEGF levels in GB cells were downregulated by cedrol treatment,
which suggested that cedrol suppressed tumor-induced angiogenesis.
Taken together, these findings suggested that cedrol could serve as
a novel drug for the prevention or therapy of cancer and
angiogenesis-related diseases.
Phosphorylation of VEGFR-2 is critical for
VEGF-mediated proliferation, migration and microvascular
permeability of endothelial cells, and further stimulates several
signaling networks to induce angiogenesis, including MAPK/ERK and
AKT/mTOR (7,34). MAPK family members, such as ERK, are
important signaling components in VEGFR2-regulated cell migration
and differentiation of vascular progenitor cells, and in response
to external stimuli, may lead to changes in the cytoskeleton
(35). Concomitantly, AKT/mTOR
signaling activated by VEGFR2 improves survival and migration and
induces cytoskeletal rearrangement in HUVECs (7,36). In
the present study, it was demonstrated that VEGF-induced
phosphorylation of VEGFR2, and the activation of ERK, AKT and
P70SK6 were significantly inhibited by cedrol. This suggested that
cedrol inhibits the VEGFR2-ERK and AKT/P70S6K pathways and
subsequently alters the cytoskeleton, resulting in the reduction of
cell migration and tube formation, and exhibits anti-angiogenic
effects in HUVECs. In addition, adhesion molecules such as VCAM-1
and ICAM-1 were previously reported to be upregulated in diseased
states, including angiogenesis, inflammation and vascular injury,
and are required for the attachment of endothelial cells to the
extracellular matrix to form new capillaries (8). Activation of VEGFR2 by VEGF also
induces the expression of certain cytoplasmic proteins, including
FAK and MMP, in tumor-derived HUVECs (37). Degradation of the extracellular
matrix by MMPs, primarily MMP-2 and MMP-9, has been associated with
angiogenesis-dependent intravasation and metastasis (38). In the present study, it was
demonstrated that the mRNA and protein expression levels of VCAM-1,
ICAM-1 and MMP-9 were increased by VEGF in the experimental system
and were abrogated by cedrol. This suggested that decreased VCAM-1,
ICAM-1 and MMP-9 might also be responsible for interfering with the
expression of VEGF/VEGFR2 signaling, thus inhibiting the
neo-angiogenesis process.
Solid tumors require new blood vessels to obtain
oxygen and essential nutrients to support tumor cell survival,
invasion and metastasis when they grow beyond a 1–2 mm diameter
(2,3). There is increasing evidence that VEGF
can provide pro-survival and pro-angiogenesis signals to tumor
stimulated HUVECs, which are regulated by VEGF receptors and have
been regarded as a direct target against angiogenesis (39). VEGF is one of the most critical
mediators involved in angiogenesis in cancers (4) and is correlated with glioma grade and
prognosis (14,15). Hence, disruption or blockage of
tumor angiogenesis or VEGF/VEGFR2 signaling may be a therapeutic
option for the treatment of solid tumors. Serum starvation induces
upregulation of VEGF in cancer cells and triggers
neovascularization (40). In the
present study, a model of tumor cell-induced angiogenesis was
established using CM from serum-free starved DBTRG-05MG cells
treated with cedrol. The results demonstrated that cedrol not only
inhibited the proliferation of DBTRG-05MG cells, but also
suppressed CM-induced tube formation in HUVECs. mRNA and protein
expression levels of VEGF in DBTRG-05MG cells were reduced by
cedrol in a dose-dependent manner, which suggested that cedrol
inhibited tumor cell-induced angiogenesis by decreasing VEGF
expression. Moreover, cedrol inhibited the expression of growth
(p-AKT and p-ERK), adhesion (VCAM-1 and ICAM-1) and invasion
(MMP-9) markers. Therefore, these findings indicated that cedrol
could be used to treat angiogenesis, growth and metastasis in
future cancer treatments.
In conclusion, the present study demonstrated that
cedrol suppressed VEGF-induced cell proliferation, migration and
invasion in a dose-dependent manner and attenuated tube formation
triggered by VEGF or CM from GB cells. This evidence suggested that
cedrol may have the potential to be developed as a therapeutic
agent for GB treatment and angiogenesis-related diseases.
Acknowledgements
The absorbance values in MTT assay was detected
using the Spectra Max Plus 384 Microplate Reader, performed in the
Instrument Center of Chung Shan Medical University, which is
supported by the National Science Council, Ministry of Education
and Chung Shan Medical University.
Funding
The present study was supported by the Ministry of Science and
Technology (grant nos. MOST 109-2320-B-040-012, MOST
110-2320-B-040-006 and MOST 111-2320-B-040-022), Taiwan, R.O.C.
Availability of data and materials
The datasets used and/or analyzed in this study are
available from the corresponding authors on reasonable request.
Authors' contributions
CYH and NMT conceived the study, obtained funding
and edited the manuscript. KFC and CYL performed the experiments,
analyzed data and wrote the manuscript. YCH performed statistical
analysis. NMT and KFC confirm the authenticity of all the raw data.
All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tahergorabi Z and Khazaei M: A review on
angiogenesis and its assays. Iran J Basic Med Sci. 15:1110–1126.
2012.PubMed/NCBI
|
|
2
|
Welch DR and Hurst DR: Defining the
hallmarks of metastasis. Cancer Res. 79:3011–3027. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kroemer G and Pouyssegur J: Tumor cell
metabolism: Cancer's Achilles' heel. Cancer Cell. 13:472–482. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ghalehbandi S, Yuzugulen J, Pranjol MZI
and Pourgholami MH: The role of VEGF in cancer-induced angiogenesis
and research progress of drugs targeting VEGF. Eur J Pharmacol.
949:1755862023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Niu G and Chen X: Vascular endothelial
growth factor as an anti-angiogenic target for cancer therapy. Curr
Drug Targets. 11:1000–1017. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sharma PS, Sharma R and Tyagi T:
VEGF/VEGFR pathway inhibitors as anti-angiogenic agents: Present
and future. Curr Cancer Drug Targets. 11:624–653. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Holmes K, Roberts OL, Thomas AM and Cross
MJ: Vascular endothelial growth factor receptor-2: Structure,
function, intracellular signalling and therapeutic inhibition. Cell
Signal. 19:2003–2012. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim I, Moon SO, Kim SH, Kim HJ, Koh YS and
Koh GY: Vascular endothelial growth factor expression of
intercellular adhesion molecule 1 (ICAM-1), vascular cell adhesion
molecule 1 (VCAM-1), and E-selectin through nuclear factor-kappa B
activation in endothelial cells. J Biol Chem. 276:7614–7620. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Turunen MP and Yla-Herttuala S: Epigenetic
regulation of key vascular genes and growth factors. Cardiovasc
Res. 90:441–446. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ostrom QT, Cioffi G, Gittleman H, Patil N,
Waite K, Kruchko C and Barnholtz-Sloan JS: CBTRUS statistical
report: Primary brain and other central nervous system tumors
diagnosed in the United States in 2012-2016. Neurooncology. 21
(Suppl 5):v1–v100. 2019.
|
|
11
|
Chen B, Chen C, Zhang Y and Xu J: Recent
incidence trend of elderly patients with glioblastoma in the United
States, 2000-2017. BMC Cancer. 21:542021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Krex D, Klink B, Hartmann C, von Deimling
A, Pietsch T, Simon M, Sabel M, Steinbach JP, Heese O, Reifenberger
G, et al: Long-term survival with glioblastoma multiforme. Brain.
130:2596–2606. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Groblewska M and Mroczko B: Pro- and
antiangiogenic factors in gliomas: Implications for novel
therapeutic possibilities. Int J Mol Sci. 22:61262021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen W, He D, Li Z, Zhang X, Pan D and
Chen G: Overexpression of vascular endothelial growth factor
indicates poor outcomes of glioma: A systematic review and
meta-analysis. Int J Clin Exp Med. 8:8709–8719. 2015.PubMed/NCBI
|
|
15
|
Luo Y, Hou WT, Zeng L, Li ZP, Ge W, Yi C,
Kang JP, Li WM, Wang F, Wu DB, et al: Progress in the study of
markers related to glioma prognosis. Eur Rev Med Pharmacol Sci.
24:7690–7697. 2020.PubMed/NCBI
|
|
16
|
Mojay G: The aromatic and acupressure
treatment of common musculoskeletal disorders: An Oriental medicine
approach. Int J Aromatherapy. 14:81–88. 2004. View Article : Google Scholar
|
|
17
|
Huang CY, Chien JH, Chang KF, Hsiao CY,
Huang YC, Chen YT, Hsu MY, Hsieh MC and Tsai NM: Cedrus atlantica
extract exerts antiproliferative effect on colorectal cancer
through the induction of cell cycle arrest and apoptosis. Food Sci
Nutr. 10:1638–1648. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chang KF, Chang JT, Huang XF, Huang YC, Li
CY, Weng JC, Hsiao CY, Hsu HJ and Tsai NM: Cedrus atlantica extract
suppress glioblastoma growth through promotion of genotoxicity and
apoptosis: In vitro and in vivo studies. Int J Med Sci.
18:2417–2430. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dakir M, El Hanbali F, Mellouki F, Akssira
M, Benharref A, Quilez Del Moral JF and Barrero AF: Antibacterial
diterpenoids from Cedrus atlantica. Nat Prod Res. 19:719–722. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Emer AA, Donatello NN, Batisti AP,
Oliveira Belmonte LA, Santos ARS and Martins DF: The role of the
endocannabinoid system in the antihyperalgesic effect of Cedrus
atlantica essential oil inhalation in a mouse model of
postoperative pain. J Ethnopharmacol. 210:477–484. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sakhaee MH, Sayyadi SAH, Sakhaee N,
Sadeghnia HR, Hosseinzadeh H, Nourbakhsh F and Forouzanfar F:
Cedrol protects against chronic constriction injury-induced
neuropathic pain through inhibiting oxidative stress and
inflammation. Metab Brain Dis. 35:1119–1126. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang JW, Chen SS, Zhang YM, Guan J, Su GY,
Ding M, Li W and Zhao YQ: Anti-inflammatory and analgesic activity
based on polymorphism of cedrol in mice. Environ Toxicol Pharmacol.
68:13–18. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oh I, Yang WY, Park J, Lee S, Mar W, Oh KB
and Shin J: In vitro Na+/K+-ATPase inhibitory activity and
antimicrobial activity of sesquiterpenes isolated from Thujopsis
dolabrata. Arch Pharm Res. 34:2141–2147. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chang KF, Huang XF, Chang JT, Huang YC, Lo
WS, Hsiao CY and Tsai NM: Cedrol, a sesquiterpene alcohol, enhances
the anticancer efficacy of temozolomide in attenuating drug
resistance via regulation of the DNA damage response and MGMT
Expression. J Nat Prod. 83:3021–3029. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chang KF, Huang XF, Chang JT, Huang YC,
Weng JC and Tsai NM: Cedrol suppresses glioblastoma progression by
triggering DNA damage and blocking nuclear translocation of the
androgen receptor. Cancer Lett. 495:180–190. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Claesson-Welsh L and Welsh M: VEGFA and
tumour angiogenesis. J Intern Med. 273:114–127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo S, Lok J, Liu Y, Hayakawa K, Leung W,
Xing C, Ji X and Lo EH: Assays to examine endothelial cell
migration, tube formation, and gene expression profiles. Methods
Mol Biol. 1135:393–402. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Quintero-Fabian S, Arreola R,
Becerril-Villanueva E, Torres-Romero JC, Arana-Argáez V,
Lara-Riegos J, Ramírez-Camacho MA and Alvarez-Sánchez ME: Role of
matrix metalloproteinases in angiogenesis and cancer. Front Oncol.
9:13702019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fallah A, Sadeghinia A, Kahroba H, Samadi
A, Heidari HR, Bradaran B, Zeinali S and Molavi O: Therapeutic
targeting of angiogenesis molecular pathways in
angiogenesis-dependent diseases. Biomed Pharmacother. 110:775–785.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Verheul HM and Pinedo HM: Possible
molecular mechanisms involved in the toxicity of angiogenesis
inhibition. Nat Rev Cancer. 7:475–485. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen X, Shen J, Zhao JM, Guan J, Li W, Xie
QM and Zhao YQ: Cedrol attenuates collagen-induced arthritis in
mice and modulates the inflammatory response in LPS-mediated
fibroblast-like synoviocytes. Food Funct. 11:4752–4764. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mishra SK, Bae YS, Lee YM, Kim JS, Oh SH
and Kim HM: Sesquiterpene alcohol cedrol chemosensitizes human
cancer cells and suppresses cell proliferation by destabilizing
plasma membrane lipid rafts. Front Cell Dev Biol. 8:5716762020.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zachary I and Gliki G: Signaling
transduction mechanisms mediating biological actions of the
vascular endothelial growth factor family. Cardiovasc Res.
49:568–581. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu J, Liu X, Jiang Y, Chu L, Hao H, Liua
Z, Verfaillie C, Zweier J, Gupta K and Liu Z: MAPK/ERK signalling
mediates VEGF-induced bone marrow stem cell differentiation into
endothelial cell. J Cell Mol Med. 12:2395–2406. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao M, Bai H, Wang E, Forrester JV and
McCaig CD: Electrical stimulation directly induces pre-angiogenic
responses in vascular endothelial cells by signaling through VEGF
receptors. J Cell Sci. 117:397–405. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Weis SM and Cheresh DA: Tumor
angiogenesis: Molecular pathways and therapeutic targets. Nat Med.
17:1359–1370. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Deryugina EI and Quigley JP: Tumor
angiogenesis: MMP-mediated induction of intravasation- and
metastasis-sustaining neovasculature. Matrix Biol. 44–46. 94–112.
2015.
|
|
39
|
Saraswati S, Kumar S and Alhaider AA:
α-santalol inhibits the angiogenesis and growth of human prostate
tumor growth by targeting vascular endothelial growth factor
receptor 2-mediated AKT/mTOR/P70S6K signaling pathway. Mol Cancer.
12:1472013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu XQ, Kiefl R, Roskopf C, Tian F and
Huber RM: Interactions among lung cancer cells, fibroblasts, and
macrophages in 3D Co-cultures and the impact on MMP-1 and VEGF
expression. PLoS One. 11:e01562682016. View Article : Google Scholar : PubMed/NCBI
|