Introduction
Src homology-domain-containing protein tyrosine
phosphatase (SHP) is a protein tyrosine phosphatase (PTP) family
member. SHP comprises SHP1, encoded by protein tyrosine phosphatase
non-receptor type (PTPN) 6, and SHP2, encoded by PTPN11 (1,2).
SHP2/PTPN11 is considered a signaling molecule that is involved in
the regulation of a number of cellular processes, such as cell
growth, differentiation, the mitotic cycle and oncogenic
transformation. SHP2 is widely expressed in a variety of human
tissues, such as breast and endometrial cancer (3,4).
SHP2 regulates cell proliferation, differentiation,
apoptosis and survival, affecting multiple signaling pathways, such
as the mitogen-activated protein kinase pathway. The abnormal
expression and mutation of SHP2 are related to human developmental
disorders, leukemia and solid tumors (such as lung, breast, ovarian
and endometrial cancer) (3,4). In different cancer types, activation
of SHP2 has been proposed as a disease cause. For example, Kim
et al (5) indicated that
SHP2 was not expressed in normal gastric mucosal cells, while it
was found to be expressed at high levels in 87% of gastric cancer
tissues, and was markedly related to the progression of gastric
cancer. Patients with gastric cancer and high SHP2 expression
demonstrate higher pathological grades, tumor stage (T stage) and
lymph node stage. Elevated expression of SHP2 indicates a poor
prognosis in patients with gastric cancer. SHP2 is widely
upregulated in invasive ductal carcinoma of the breast, and is
associated with HER2 expression, androgen receptor nuclear
localization, T stage and lymph node metastasis (6,7).
Inhibition of SHP2 expression can hinder the development of
receptor tyrosine kinase (RTK)-driven malignant growth cells, such
as those derived from KRAS-mutated breast carcinoma. In non-small
cell lung cancer (NSCLC) with a KRAS mutation, the blockage of SHP2
expression is enough to induce tumor senescence, which triggers the
clearance of cancer cells by the immune system (8). A limited number of studies have
demonstrated that SHP2 expression is decreased in certain cancer
types. For example, Jiang et al (9) observed that SHP2 expression was
reduced in 70.6% of patients with liver cancer, and decreased SHP2
expression was associated with a poor prognosis in hepatocellular
carcinoma. Therefore, given the close relationship between
SHP2/PTPN11 and human diseases, and its wide expression in tumor
tissues, as well as the dual function of SHP2/PTPN11 as a tumor
suppressor and tumor-promoting gene in tumors, the comprehensive
discussion of SHP2/PTPN11 has important clinical significance. The
present study assessed the expression levels of SHP2 in different
tumors, and its relationship with prognosis, immunity and the tumor
microenvironment.
Materials and methods
Data acquisition and
pre-processing
SHP2 expression levels of the 33 types of tumor with
associated data in The Cancer Genome Atlas (TCGA) database and the
corresponding tumor RNA-sequencing data were downloaded from the
Genomic Data Commons data portal website (https://portal.gdc.cancer.gov/). Clinical information
(SHP2 expression level, follow-up time and tumor type), tumor
mutational burden and microsatellite instability of the
corresponding patients were also downloaded. The 33 types of tumors
included the following: Adrenocortical carcinoma (ACC), bladder
urothelial carcinoma (BLCA), breast invasive carcinoma (BRCA),
cervical and endocervical cancer (CESC), cholangiocarcinoma (CHOL),
colon adenocarcinoma (COAD), lymphoid neoplasm diffuse large B-cell
lymphoma (DLBC), esophageal carcinoma (ESCA), glioblastoma
multiforme (GBM), head and neck squamous cell carcinoma (HNSC),
kidney chromophobe (KICH), kidney renal clear cell carcinoma
(KIRC), kidney renal papillary cell carcinoma (KIRP), acute myeloid
leukemia (LAML), brain lower grade glioma (LGG), liver
hepatocellular carcinoma (LIHC), lung adenocarcinoma (LUAD), lung
squamous cell carcinoma (LUSC), mesothelioma (MESO), ovarian serous
cystadenocarcinoma (OV), pancreatic adenocarcinoma (PAAD),
pheochromocytoma and paraganglioma (PCPG), prostate adenocarcinoma
(PRAD), rectal adenocarcinoma (READ), sarcoma (SARC), skin
cutaneous melanoma (SKCM), stomach adenocarcinoma (STAD),
testicular germ cell tumors (TGCT), thyroid carcinoma (THCA),
thymoma (THYM), uterine corpus endometrial carcinoma (UCEC),
uterine carcinosarcoma (UCS) and uveal melanoma (UVM) (10).
Expression levels, prognostic
evaluation and the tumor microenvironment
The R 4.2.2 software (R Core Team; www.r-project.org) was used for statistical analysis.
Univariate Cox regression analysis was used to evaluate the
association between SHP2 gene expression, overall survival (OS) and
disease free survival (DFS); the use of forest plots through the
‘forest plot’ R package (version 3.1.1) aimed to display the
P-value, hazard ratio (HR) and 95% confidence interval (CI) of
every variable (11). According to
the median value of the SHP2/PTPN11 gene expression, patients were
divided into SHP2/PTPN11 high and SHP2/PTPN11 low expression
groups. The Kaplan-Meier method was used to draw the survival curve
of every tumor type, and the log-rank test was used to analyze
survival status.
The matrix and immune scores of every patient were
calculated using the ‘ESTIMATE’ software package (version 1.0.13)
to evaluate the infiltration of immune cells in tumor tissue and
tumor purity. For reliable immune score evaluation, the ‘CIBERSORT’
algorithm was used to evaluate tumor immune cell infiltration
(12). Spearman's correlation
analysis was used to evaluate the correlation between SHP2/PTPN11
gene expression and the target, including matrix and immune scores,
immune cell infiltration, tumor mutation load and microsatellite
instability (13). Student's
unpaired t-test was used to evaluate the SHP2/PTPN11 gene
expression levels between tumor and corresponding non-tumor normal
tissues. P<0.05 was considered to indicate a statistically
significant difference.
Patients and ELISA
A total of 42 breast cancer and 42 normal human
serum specimens were collected. A total of 34 ovarian cancer and 36
normal human serum specimens were collected. A total of 29
endometrial cancer and 33 normal human serum specimens were
collected. A total of 46 cervical cancer and 40 normal human serum
specimens were collected. All samples were collected from the
Chongqing Health Center for Women and Children (Chongqing, China)
from February 2021 to December 2022. The patients' personal
information was not collected. The patients provided written
informed consent for the collection of blood samples. All specimens
were stored at −80°C and tested by ELISA in January 2023. The
present study was approved by the Ethics Association of Chongqing
Health Center for Women and Children (approval no.
cstc2020-jyk2.0).
The expression levels of SHP2/PTPN11 in serum were
detected by double antibody one-step sandwich ELISA. The PTPN11
ELISA kit (cat. no. P20220809; Shanghai Enzyme-linked Biotechnology
Co., Ltd.) was used at room temperature. A total of 50 µl standard
at different concentrations was used, and 50 µl sample was added
into the sample well. A total of 100 µl horseradish
peroxidase-labeled antibody was added to the standard and sample
wells, and incubated in a water bath at 37°C for 60 min. Following
washing, 50 µl substrates A and B was added to the plate wells and
incubated at 37°C for 15 min. A total of 50 µl stop solution was
added to each well, and the optical density (OD) value was measured
at a wavelength of 450 nm within 15 min. The test was repeated
three times.
Statistical analysis
SPSS version 25.0 (IBM Corp.) was used to analyze
the experimental data. The Shapiro-Wilk test was used to evaluate
whether experimental data conformed to a normal distribution.
Unpaired Student's t-test was used to evaluate the expression
levels of the SHP2/PTPN11 gene between tumor and normal tissues.
Spearman's correlation analysis was used to evaluate the
correlation between the SHP2/PTPN11 gene expression and the target
(matrix score, immune score, immune cell infiltration, tumor
mutation load and microsatellite instability) (13). The χ2 test was used to
analyze the difference in SHP2 expression level in the peripheral
blood of patients with breast, ovarian, endometrial and cervical
cancer. P<0.05 was considered to indicate a statistically
significant difference.
Results
Expression levels of the PTPN11 gene
in tumors
Among the 33 types of tumors that exhibit PTPN11 in
TCGA, 30 types of tumors could be compared with their corresponding
normal tissues as controls (LAML, MESO and UVM had no controls).
For a total of 26 out of the 30 types of tumors, statistically
significant differences were noted in the expression levels of the
PTPN11 gene compared with those of the normal tissues (P<0.05).
Among them, PTPN11 was expressed at higher levels in ACC, BRCA,
CHOL, COAD, DLBC, ESCA, GBM, HNSC, KICH, KIRC, KIRP, LGG, LIHC,
LUAD, LUSC, OV, PAAD, PRAD, READ, SKCM, STAD, TGCT and THCA, while
the expression levels of PTPN11 were decreased in THYM, UCEC and
UCS (Fig. 1).
 | Figure 1.Expression level of PTPN11 gene in 33
types of tumors and their corresponding normal tissues. SPSS
(version 25.0; IBM Corp.) was used for data normalization
processing (Z-score normalization). Unpaired Student's t-test was
used to analyze the data. *P<0.05, **P<0.01 and ***P<0.001
vs. normal tissues. ACC, adrenocortical carcinoma; BLCA, bladder
urothelial carcinoma; BRCA, breast invasive carcinoma; CESC,
cervical and endocervical cancers; CHOL, cholangiocarcinoma; COAD,
colon adenocarcinoma; DLBC, lymphoid neoplasm diffuse large B-cell
lymphoma; ESCA, esophageal carcinoma; GBM, glioblastoma multiforme;
HNSC, head and neck squamous cell carcinoma; KICH, kidney
chromophobe; KIRC, kidney renal clear cell carcinoma; KIRP, kidney
renal papillary cell carcinoma; LAML, acute myeloid leukemia; LGG,
brain lower grade glioma; LIHC, liver hepatocellular carcinoma;
LUAD, lung adenocarcinoma; LUSC, lung squamous cell carcinoma;
MESO, mesothelioma; OV, ovarian serous cystadenocarcinoma; PAAD,
pancreatic adenocarcinoma; PCPG, pheochromocytoma and
paraganglioma; PRAD, prostate adenocarcinoma; READ, rectal
adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous melanoma; STAD,
stomach adenocarcinoma; TGCT, testicular germ cell tumors; THCA,
thyroid carcinoma; THYM, thymoma; UCEC, uterine corpus endometrial
carcinoma; UCS, uterine carcinosarcoma; UVM, uveal melanoma;
PTPN11, protein tyrosine phosphatase non-receptor type 11. |
Expression levels of the PTPN11 gene
are associated with patient prognosis
According to the median value of the PTPN11 gene
expression, patients with 33 types of tumors were divided into
PTPN11 high and low expression groups. Survival analysis
demonstrated that the OS of patients in the PTPN11 high and low
expression groups was significantly different among the BRCA (HR,
1.70; 95% CI, 1.01–2.21; P=0.001), CESC (HR, 1.70; 95% CI,
1.23–1.98; P=0.023), MESO (HR, 1.80; 95% CI, 1.23–2.22; P=0.027),
BLCA (HR, 1.40; 95% CI, 1.12–1.68; P=0.031) and LIHC (HR, 1.40; 95%
CI, 1.23–1.87; P=0.049) tumor types. Patients in the PTPN11 high
expression group exhibited shorter OS times than those in the
control group (P<0.05). However, high expression levels of
PTPN11 improved the OS time of patients with the KIRC tumor type
(HR, 0.60; 95% CI, 0.54–0.89; P=0.001) (Fig. 2A). Considering that OS may be
affected by non-tumor-associated deaths, the relationship between
PTPN11 gene expression and DFS was evaluated further. High
expression of the PTPN11 gene reduced the DFS time of patients with
the BLCA (HR, 1.50; 95% CI; 1.22–1.76; P=0.021) and CESC (HR, 1.90;
95% CI, 1.65–2.22; P=0.043) tumor types. This phenomenon was not
observed in patients with other tumor types. However, high
expression levels of PTPN11 improved the DFS time of patients with
KIRC (HR, 0.65; 95% CI, 0.61–0.76; P=0.022) (Fig. 2B).
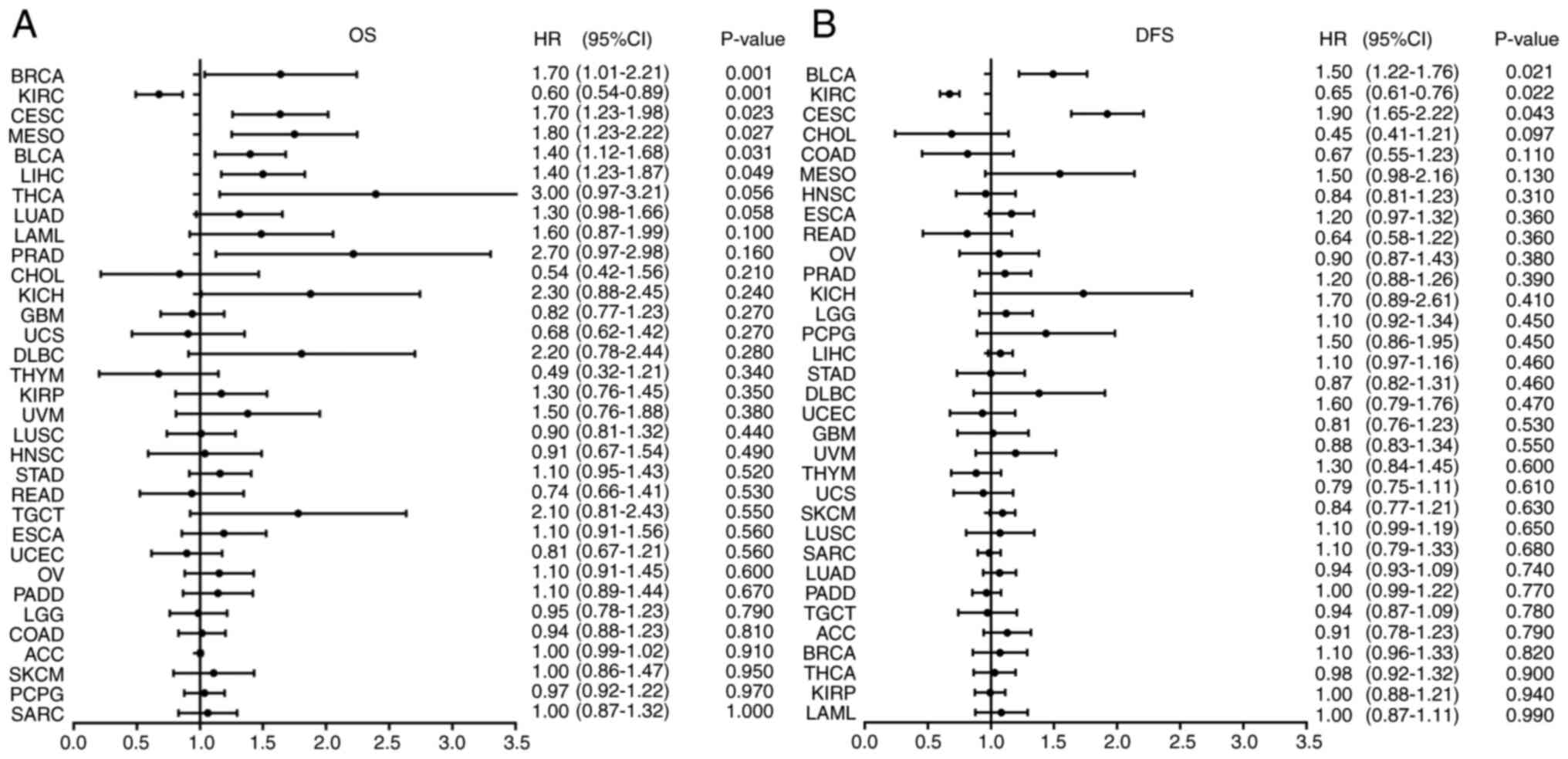 | Figure 2.Relationship between the expression
level of protein tyrosine phosphatase non-receptor type 11 gene and
prognosis in 33 types of tumors. (A) OS and (B) DFS. Univariate Cox
regression analysis was used to analyze the data; ‘forest’ plots
were used to display the P-value, HR and 95% CI of each variable.
HR, hazard ratio; CI, confidence interval; OS, overall survival;
DFS, disease-free survival; ACC, adrenocortical carcinoma; BLCA,
bladder urothelial carcinoma; BRCA, breast invasive carcinoma;
CESC, cervical and endocervical cancers; CHOL, cholangiocarcinoma;
COAD, colon adenocarcinoma; DLBC, lymphoid neoplasm diffuse large
B-cell lymphoma; ESCA, esophageal carcinoma; GBM, glioblastoma
multiforme; HNSC, head and neck squamous cell carcinoma; KICH,
kidney chromophobe; KIRC, kidney renal clear cell carcinoma; KIRP,
kidney renal papillary cell carcinoma; LAML, acute myeloid
leukemia; LGG, brain lower grade glioma; LIHC, liver hepatocellular
carcinoma; LUAD, lung adenocarcinoma; LUSC, lung squamous cell
carcinoma; MESO, mesothelioma; OV, ovarian serous
cystadenocarcinoma; PAAD, pancreatic adenocarcinoma; PCPG,
pheochromocytoma and paraganglioma; PRAD, prostate adenocarcinoma;
READ, rectal adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous
melanoma; STAD, stomach adenocarcinoma; TGCT, testicular germ cell
tumors; THCA, thyroid carcinoma; THYM, thymoma; UCEC, uterine
corpus endometrial carcinoma; UCS, uterine carcinosarcoma; UVM,
uveal melanoma. |
PTPN11 gene expression correlates with
immune checkpoints and immune scores
By using Spearman's correlation analysis, it was
found that the expression levels of the PTPN11 gene were
significantly correlated with cancer types in the presence of
immune cells. In BLCA, BRCA, CESC, CHOL, COAD, HNSC, KIRC, KIRP,
LGG, PRAD, READ, SKCM, STAD, THCA, THYM and UCEC, the expression
levels of the PTPN11 gene were significantly negatively correlated
with regulatory, follicular helper and cluster of differentiation
(CD)8+ T cells (P<0.05). In BRCA, CESC, COAD, ESCA,
HNSC, KICH, KIRC, LAML, LGG, LUAD, LUSC, OV, PCPG, PRAD, READ,
SARC, TGCT, THCA, THYM and UCEC, the expression levels of the
PTPN11 gene were significantly and positively correlated with the
number of CD4+ T cells (P<0.05). In BLCA, BRCA, HNSC,
LIHC, LUAD, PRAD and THCA, the expression levels of the PTPN11 gene
were significantly and positively correlated with the number of
neutrophils (P<0.05). In BRCA, KIRC, SKCM, STAD, TGCT, THCA,
THYM and UCEC, the expression levels of the PTPN11 gene were
significantly and positively correlated with the number of
macrophages M2 (P<0.05; Fig.
3).
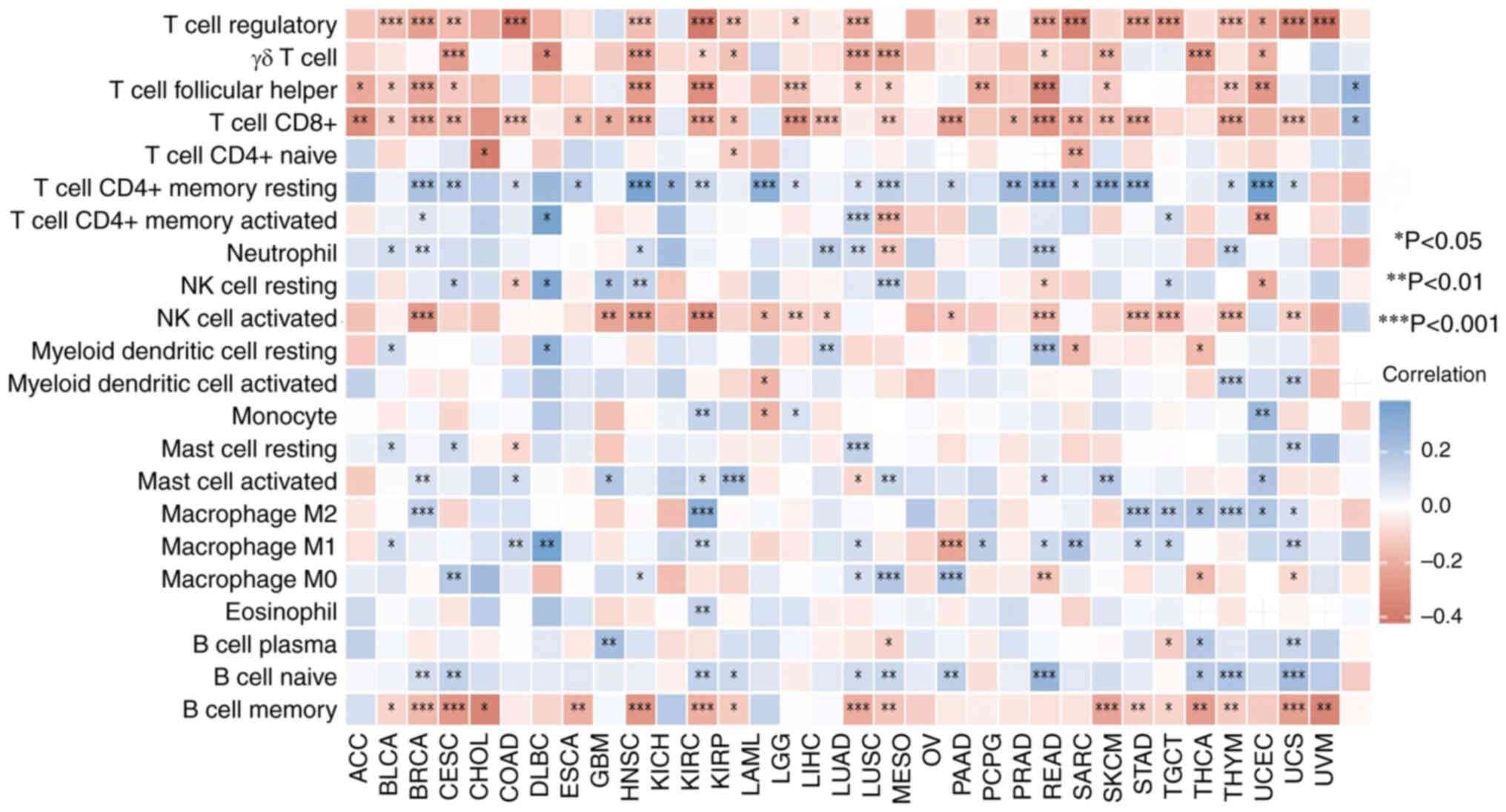 | Figure 3.Correlation between the expression
level of protein tyrosine phosphatase non-receptor type 11 gene and
immune cells in different cancer types. Spearman's correlation
analysis was used to assess the data. NK, natural killer; CD,
cluster of differentiation; ACC, adrenocortical carcinoma; BLCA,
bladder urothelial carcinoma; BRCA, breast invasive carcinoma;
CESC, cervical and endocervical cancers; CHOL, cholangiocarcinoma;
COAD, colon adenocarcinoma; DLBC, lymphoid neoplasm diffuse large
B-cell lymphoma; ESCA, esophageal carcinoma; GBM, glioblastoma
multiforme; HNSC, head and neck squamous cell carcinoma; KICH,
kidney chromophobe; KIRC, kidney renal clear cell carcinoma; KIRP,
kidney renal papillary cell carcinoma; LAML, acute myeloid
leukemia; LGG, brain lower grade glioma; LIHC, liver hepatocellular
carcinoma; LUAD, lung adenocarcinoma; LUSC, lung squamous cell
carcinoma; MESO, mesothelioma; OV, ovarian serous
cystadenocarcinoma; PAAD, pancreatic adenocarcinoma; PCPG,
pheochromocytoma and paraganglioma; PRAD, prostate adenocarcinoma;
READ, rectal adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous
melanoma; STAD, stomach adenocarcinoma; TGCT, testicular germ cell
tumors; THCA, thyroid carcinoma; THYM, thymoma; UCEC, uterine
corpus endometrial carcinoma; UCS, uterine carcinosarcoma; UVM,
uveal melanoma. |
Sialic acid binding Ig like lectin 15, indoleamine
2,3-dioxygenase 1, CD274, hepatitis A virus cellular receptor 2,
programmed cell death (PDCD) 1, cytotoxic T-lymphocyte associated
protein 4, lymphocyte activating 3 (LAG3) and PDCD 1 ligand 2 are
transcripts related to immune checkpoints. The expression levels of
these eight immune checkpoint-related genes were obtained and
assessed. Spearman's correlation analysis was used to analyze the
immune checkpoints genes related to PTPN11. Results were found in
the DLBC, PCPG, PAAD, PRAD, LIHC, UVM and READ tumor types, where
the expression levels of the PTPN11 gene were significantly and
positively correlated with CD274, which had a large number of
associations (P<0.05). However, in UCS, THCA and MESO
(correlation coefficient >±0.3), the expression levels of the
PTPN11 gene were significantly and negatively correlated with LAG3
(P<0.05; Fig. 4; Table I).
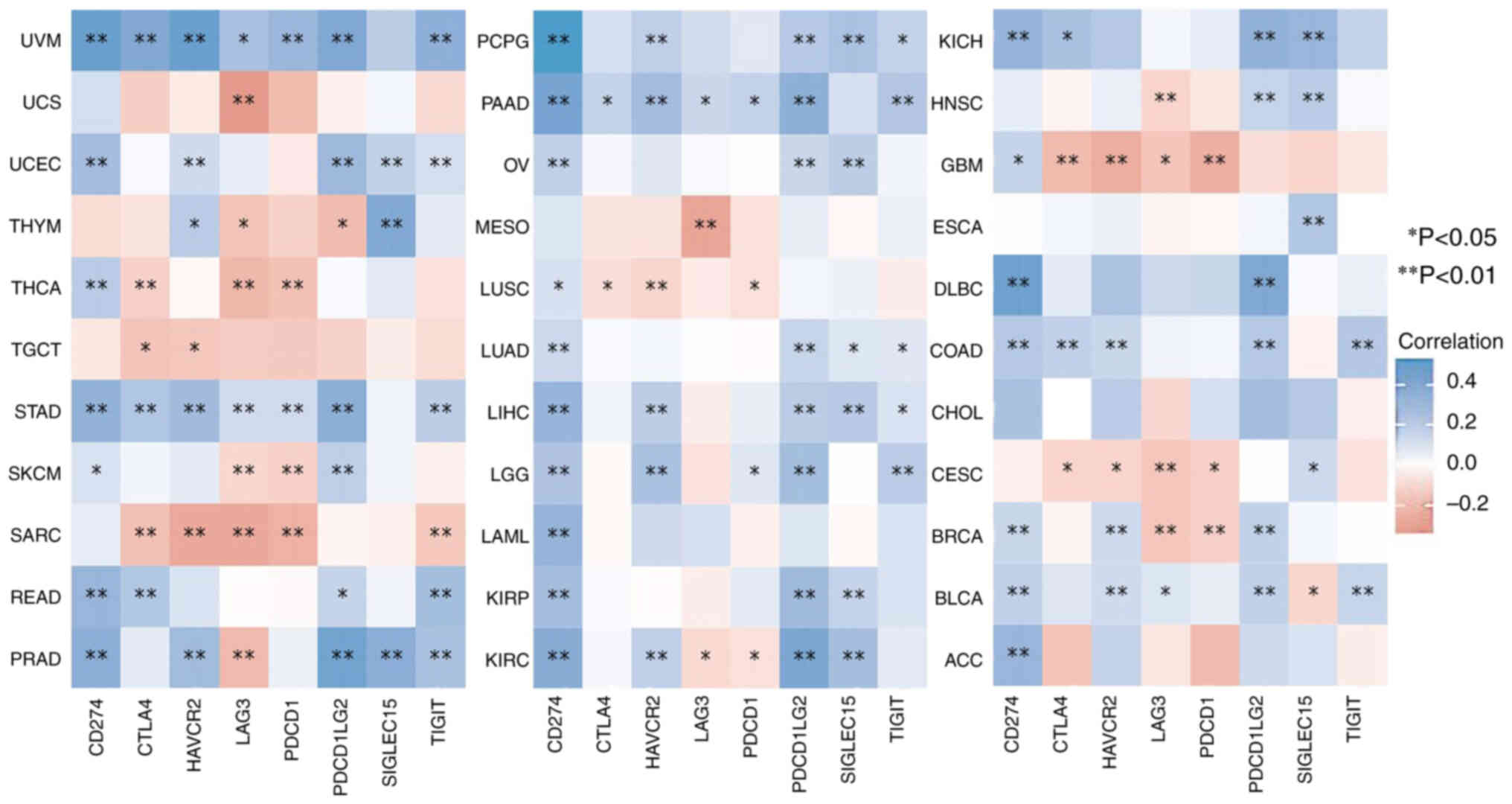 | Figure 4.Correlation between the expression
level of protein tyrosine phosphatase non-receptor type 11 gene and
immune checkpoints in different cancer types. Spearman's
correlation analysis was used to assess the data. ACC,
adrenocortical carcinoma; BLCA, bladder urothelial carcinoma; BRCA,
breast invasive carcinoma; CESC, cervical and endocervical cancers;
CHOL, cholangiocarcinoma; COAD, colon adenocarcinoma; DLBC,
lymphoid neoplasm diffuse large B-cell lymphoma; ESCA, esophageal
carcinoma; GBM, glioblastoma multiforme; HNSC, head and neck
squamous cell carcinoma; KICH, kidney chromophobe; KIRC, kidney
renal clear cell carcinoma; KIRP, kidney renal papillary cell
carcinoma; LAML, acute myeloid leukemia; LGG, brain lower grade
glioma; LIHC, liver hepatocellular carcinoma; LUAD, lung
adenocarcinoma; LUSC, lung squamous cell carcinoma; MESO,
mesothelioma; OV, ovarian serous cystadenocarcinoma; PAAD,
pancreatic adenocarcinoma; PCPG, pheochromocytoma and
paraganglioma; PRAD, prostate adenocarcinoma; READ, rectal
adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous melanoma; STAD,
stomach adenocarcinoma; TGCT, testicular germ cell tumors; THCA,
thyroid carcinoma; THYM, thymoma; UCEC, uterine corpus endometrial
carcinoma; UCS, uterine carcinosarcoma; UVM, uveal melanoma. |
 | Table I.Correlation between SHP2/PTPN11 and
immune checkpoint-related genes. |
Table I.
Correlation between SHP2/PTPN11 and
immune checkpoint-related genes.
| Cancer | Immune
checkpoint-related genes | ρ | P-value |
|---|
| THYM | SIGLEC15 | 0.394 | <0.01 |
| PRAD | SIGLEC15 | 0.325 | <0.01 |
| DLBC | PDCD1LG2 | 0.442 | <0.01 |
| PRAD | PDCD1LG2 | 0.439 | <0.01 |
| THCA | PDCD1 | −0.338 | <0.01 |
| UCS | LAG3 | −0.319 | <0.05 |
| THCA | LAG3 | −0.351 | <0.01 |
| MESO | LAG3 | −0.361 | <0.01 |
| UVM | HAVCR2 | 0.323 | <0.01 |
| SARC | HAVCR2 | −0.315 | <0.01 |
| UVM | CTLA4 | 0.342 | <0.01 |
| DLBC | CD274 | 0.473 | <0.01 |
| PCPG | CD274 | 0.402 | <0.01 |
| PAAD | CD274 | 0.367 | <0.01 |
| PRAD | CD274 | 0.342 | <0.01 |
| LIHC | CD274 | 0.331 | <0.01 |
| UVM | CD274 | 0.326 | <0.01 |
| READ | CD274 | 0.302 | <0.01 |
Expression levels of the PTPN11 gene
are related to microsatellite instability and tumor mutational
burden
The expression levels of the PTPN11 gene in LAML,
LUAD, SKCM and THYM were significantly and positively correlated
with microsatellite instability. However, in UVM and LGG, the
expression levels of the PTPN11 gene were significantly and
negatively correlated with microsatellite instability. Among them,
the correlation coefficient between the expression levels of the
PTPN11 gene and microsatellite instability was the highest in UVM
(ρ=−0.313), THYM (ρ=0.212) and LAML (ρ=0.203) (Fig. 5A).
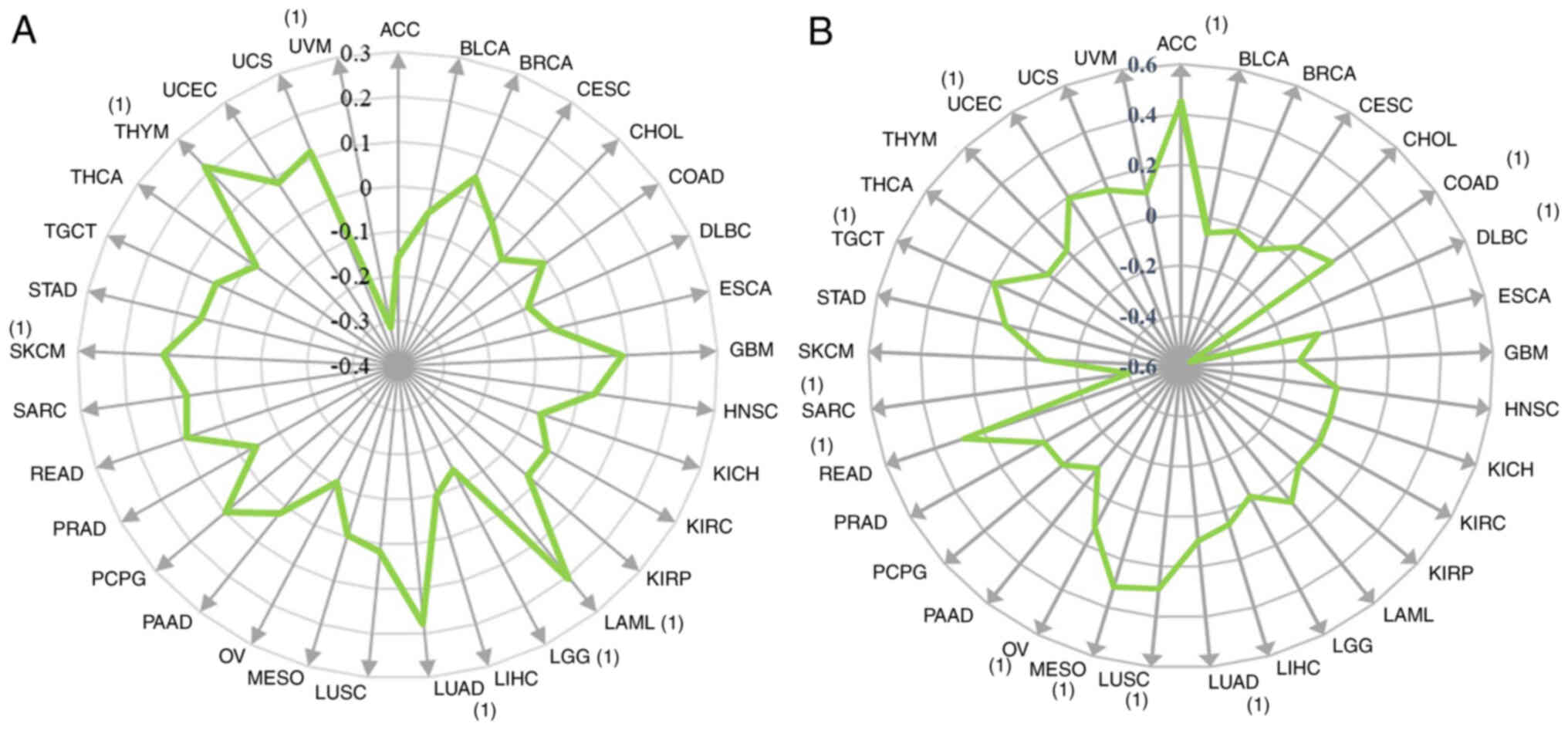 | Figure 5.Correlation between protein tyrosine
phosphatase non-receptor type 11 gene expression level and
microsatellite instability and tumor mutation burden. (A)
Microsatellite instability. (B) Tumor mutation burden. (1) indicates P<0.05. Spearman's
correlation analysis was used to assess the data. ACC,
adrenocortical carcinoma; BLCA, bladder urothelial carcinoma; BRCA,
breast invasive carcinoma; CESC, cervical and endocervical cancers;
CHOL, cholangiocarcinoma; COAD, colon adenocarcinoma; DLBC,
lymphoid neoplasm diffuse large B-cell lymphoma; ESCA, esophageal
carcinoma; GBM, glioblastoma multiforme; HNSC, head and neck
squamous cell carcinoma; KICH, kidney chromophobe; KIRC, kidney
renal clear cell carcinoma; KIRP, kidney renal papillary cell
carcinoma; LAML, acute myeloid leukemia; LGG, brain lower grade
glioma; LIHC, liver hepatocellular carcinoma; LUAD, lung
adenocarcinoma; LUSC, lung squamous cell carcinoma; MESO,
mesothelioma; OV, ovarian serous cystadenocarcinoma; PAAD,
pancreatic adenocarcinoma; PCPG, pheochromocytoma and
paraganglioma; PRAD, prostate adenocarcinoma; READ, rectal
adenocarcinoma; SARC, sarcoma; SKCM, skin cutaneous melanoma; STAD,
stomach adenocarcinoma; TGCT, testicular germ cell tumors; THCA,
thyroid carcinoma; THYM, thymoma; UCEC, uterine corpus endometrial
carcinoma; UCS, uterine carcinosarcoma; UVM, uveal melanoma. |
The expression levels of the PTPN11 gene were
significantly and positively correlated with the tumor mutation
burden in ACC, COAD, LUAD, LUSC, MESO, OV, READ, TGCT and UCEC.
However, the expression levels of the PTPN11 gene in DLBC and SARC
were significantly and negatively correlated with the tumor
mutational burden. Among them, the correlation coefficient between
the expression levels of the PTPN11 gene and the tumor mutational
burden was the highest in DLBC (ρ=−0.561), SARC (ρ=−0.400), ACC
(ρ=0.449), MESO (ρ=0.315), LUSC (ρ=0.289) and READ (ρ=0.281)
(Fig. 5B).
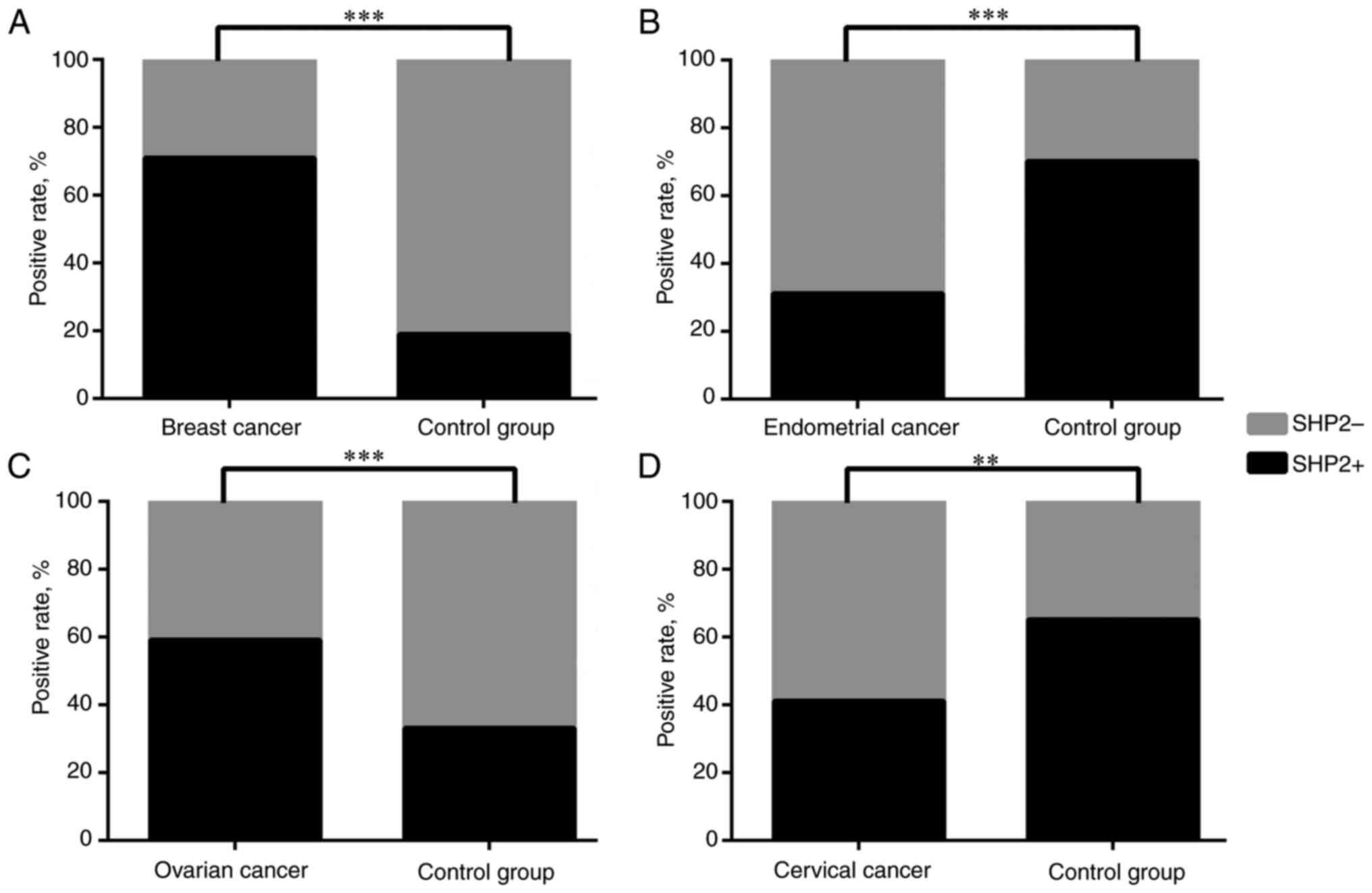
SHP2 is associated with the risk of
breast, ovarian, endometrial and cervical cancer
The peripheral blood samples from patients with 4
different tumor types were collected. ELISA was used to assess the
expression levels of SHP2. The standard OD value was 0.143. If the
OD value of the sample was higher than that of the standard OD
value, it was considered positive for SHP2, otherwise it was
considered negative. By using the χ2 test, it was found
that SHP2 was expressed at higher levels in breast cancer (71.43%;
P<0.001) and ovarian cancer (58.82%; P<0.001) than in the
control group; however, it was expressed at lower percentages in
endometrial cancer (31.03%; P<0.001) and cervical cancer
(41.30%; P=0.001) than in the control group (Table II; Fig.
6). This finding is consistent with the results of the
pan-cancer analysis in the present study.
| Table II.Expression of SHP2 in peripheral
blood. |
Table II.
Expression of SHP2 in peripheral
blood.
|
|
|
|
|
|
|
| Optical density
valuea |
|---|
|
| SHP2 |
|
|
|
|
|---|
|
|
|
|
|
|
|
|
|---|
| Cancer type | + | - | Total, n | Odds ratio | 95% confidence
interval | P-value | + | - |
|---|
| Breast |
|
|
|
|
|
|
|
|
| Cancer
group | 30 | 12 | 42 | 10.44 | 5.391–20.210 | <0.001 | 0.378 | 0.010 |
| Control
group | 8 | 34 | 42 |
|
|
| 0.303 | 0.017 |
| Ovarian |
|
|
|
|
|
|
|
|
| Cancer
group | 20 | 14 | 34 | 2.92 | 1.641–5.202 | <0.001 | 0.458 | 0.022 |
| Control
group | 12 | 24 | 36 |
|
|
| 0.404 | 0.020 |
| Endometrial |
|
|
|
|
|
|
|
|
| Cancer
group | 9 | 20 | 29 |
|
|
| 0.450 | 0.017 |
| Control
group | 23 | 10 | 33 | 0.19 | 0.105–0.352 | <0.001 | 0.488 | 0.011 |
| Cervical |
|
|
|
|
|
|
|
|
| Cancer
group | 19 | 27 | 46 |
|
|
| 0.523 | 0.029 |
| Control
group | 26 | 14 | 40 | 0.37 | 0.211–0.663 | 0.001 | 0.479 | 0.026 |
Discussion
SHP2 consists of the N-SH2 and C-SH2 domains, a
protein tyrosine phosphatase (PTP) domain and a C-terminal tail
with two tyrosine phosphorylation sites (14). The deviations in its biological
function can cause various disorders in the regulation of normal
body functions (such as normal development of the body,
cardiovascular production and immune response), and lead to the
formation of cancer, diabetes and autoimmune diseases, among others
(15). SHP2 serves a number of
roles in the formation and progression of tumors (4). SHP2 can regulate the proliferation and
metastasis of tumor cells by participating in numerous signaling
pathways such as the PI3K/AKT and RAS/ERK pathways (16–18).
Mutations or changes in the expression levels of PTPN11 can lead to
the formation of leukemia and various other tumors, such as liver,
cervical, ovarian and endometrial cancer (4,17).
Therefore, SHP2/PTPN11 may be an ideal target for cancer
intervention (4).
In the present study, the expression levels of
SHP2/PTPN11 were analyzed in tumors, and the data indicated that
SHP2/PTPN11 was expressed at high levels in KIRC. Concomitantly,
the prognostic analysis indicated that high expression of
SHP2/PTPN11 resulted in increased OS (HR, 0.60; P=0.001) and DFS
(HR, 0.65; P=0.022) times of patients with KIRC. Therefore,
SHP2/PTPN11 may be a tumor suppressor gene in KIRC, and its
elevated expression can improve patient prognosis. Increased
SHP2/PTPN11 expression decreased the OS (BLCA: HR, 1.40; P=0.031;
CESC: HR, 1.70; P=0.023) and DFS (BLCA: HR, 1.50; P=0.021; CESC:
HR, 1.90; P=0.043) times of patients with BLCA and CESC; moreover,
the high expression levels of SHP2/PTPN11 decreased the OS times of
patients with BRCA, MESO and LIHC, thereby worsening patient
prognosis, suggesting that SHP2/PTPN11 may act as an oncogene in
BLCA, CESC, BRCA, MESO and LIHC.
In a number of solid tumors, excessive SHP2
activation has been stated to serve an essential pathogenic role.
For example, Feng et al (19) indicated that SHP2 was expressed at
high levels in 94.1% of patients with NSCLC; its expression was
higher in the intratumoral area than in the stromal area. Moreover,
high expression of SHP2 is associated with an improved prognosis in
patients with NSCLC, and can increase OS and PFS time. Lei et
al (20) indicated that high
expression of SHP2 could promote the occurrence of breast cancer
and reduce lymph node metastasis. Hu et al (21) observed that the positive rate of
SHP2 in ovarian cancer tissues reached 81.67%, while the positive
rate of SHP2 in normal ovarian tissues was 0.00%. Moreover, SHP2
expression was associated with lymph node metastasis, clinical
stage and histological grade. The current study indicated that SHP2
was expressed in various tumor types. The expression levels of SHP2
were increased in the majority of tumors, with the exception of
THYM, UCEC and UCS. By using peripheral blood samples of patients
with cancer, it was found that SHP2 was not expressed at high
levels in all tumors; by contrast, it indicated tumor-promoting or
-suppressing functions in different tumors. The expression levels
of SHP2 were increased in breast and ovarian cancer, whereas they
were expressed at low levels in endometrial and cervical cancer.
Therefore, the role of SHP2 in the remaining tumors requires
additional investigation.
SHP2 is involved in the regulation of various
signaling pathways in organisms. SHP2 binding sites are present in
RTKs and backbone adaptors, such as growth factor receptor bound
(GRB)2-associated binding protein (GAB), insulin receptor
substrate, FAR1-related sequence and other proteins. Therefore,
this ‘molecular switch’ ensures that SHP2 is only activated in the
appropriate cellular regions (N-SH2 and PTP domain) (22). During the signal transduction
process of several growth factors and cytokines, SHP2 acts upstream
of RAS, and enables the full activation of the ERK/MAPK pathway
(6). The C-terminal tyrosine of
SHP2 is phosphorylated in response to the majority of the agonists,
and the tyrosine-phosphorylated SHP2 recruits the adaptor protein
GRB2 and the guanine nucleotide exchange factor son of sevenless,
which contribute to RAS activation (23). Certain studies (14,16,20)
have shown that SHP2, which is expressed at high levels in breast
cancer, can activate PI3K/AKT signaling to phosphorylate GSK3-β,
thereby promoting the proliferation of breast cancer cells.
However, SHP2 can also inhibit the PI3K/AKT signaling pathway. In
the EGFR signaling pathway, SHP2 can dephosphorylate the PI3K
binding site on GAB1 and decrease the GAB1-mediated activation of
the PI3K/AKT proteins. In addition, SHP2 can also bind to p85 to
form GAB2/SHP2/p85 complexes, thereby inhibiting the PI3K/AKT
pathway (24). SHP2 can inhibit the
gp130 pathway mediated by IL-6 and promote STAT3 dephosphorylation,
thereby negatively regulating the Janus kinase/STAT3 pathway and
eventually leading to juvenile bone marrow monocyte leukemia
(25,26). In addition, SHP2 is involved in
regulating T-cell activity by binding to the phosphotyrosine motif
of the immune checkpoint protein PDC1 through its N-SH2 domain
(27). Inhibitors that can block
the protein-protein interactions between PD-1 and SHP2 are expected
to be used as new tumor immunotherapy agents. Therefore, SHP2 has
also become a potential drug target in tumor immunotherapy
(28).
SHP2/PTPN11 may play an important role in promoting
tumor immune escape (29). As
important players in shaping the tumor microenvironment,
tumor-associated macrophages (TAMs) mediate tumor angiogenesis and
immune escape (30). TAMs are a
subset of macrophages with M2-like aggregate properties, which play
a major role in tumorigenesis, angiogenesis, grid renovation and
metastasis (31). For example, the
deletion of SHP2 in TAMs can significantly inhibit the growth of
melanoma. In response to IFN-γ or cytokine stimulation, the
deletion of SHP2 can notably enhance the ability of macrophages to
produce chemokine ligand (CXCL) 9, thereby recruiting additional T
cells, promoting the production of CXCL9 in the tumor
microenvironment, and forming a macrophage/CXCL9-T cell/IFN-γ
feedback loop to facilitate the antitumor immune function of T
cells (32,33). The aforementioned studies have
demonstrated that SHP2/PTPN11 is an expected target for managing
TAM function in immunotherapy. By inhibiting SHP2 expression, a
direct inhibition of tumor formation is facilitated by inhibiting
downstream pathways, such as that of RAS/ERK, PI3K/AKT and
JAK/STAT. By contrast, SHP2 inhibitors can also inhibit tumors by
activating T cells and promoting macrophage phagocytosis.
Therefore, SHP2 is a target for both immune and targeted
therapies.
In conclusion, the findings of the present study
showed that SHP2/PTPN11 was widely expressed in the majority of
tumors assessed, and that its expression was related to
tumorigenesis, tumor development and disease prognosis. SHP2/PTPN11
upregulation could improve OS and DFS time in patients with KIRC.
However, SHP2/PTPN11 upregulation could reduce OS time in patients
with BLCA, CESC, BRCA, MESO and LIHC. Moreover, the elevated
expression of SHP2 could also reduce patient DFS time in BLCA and
CESC. SHP2 serves an important role in maintaining the
immunosuppressive microenvironment via the inhibition of T-cell
activation and the promotion of M2 macrophage activation.
Inhibition of SHP2 may be a novel therapeutic approach with the
following dual applications: It can directly inhibit the growth of
cancer cells in specific tumors and it can change the tumor
microenvironment to promote antitumor immunity. Based on the
function of SHP2 in tumor cells, novel and effective antitumor
drugs targeting it can be developed.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Chongqing Science and
Technology Commission (grant nos. cstc2020jcyj-msxmX0588 and
cstc2019jcyj-msxmX0371).
Authors' contributions
SL conceptualized the study, visualized the data,
developed the methodology used, applied the software, performed the
statistical analysis, wrote the original draft, and reviewed and
edited the manuscript. JQ applied the software and developed the
methodology used. XW developed the methodology used, applied the
software and performed the statistical analysis. QZ developed the
methodology and performed the statistical analysis. CL
conceptualized the study, visualized the data, developed the
methodology, and wrote, reviewed and edited the manuscript. SL and
CL confirm the authenticity of all the raw data. All authors have
read and approved the final version of the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Association of Chongqing Health Center for Women and Children
(approval no. cstc2020-jyk2.0). The patients provided written
informed consent for the collection of blood samples and their use
in the present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ACC
|
adrenocortical carcinoma
|
|
BLCA
|
bladder urothelial carcinoma
|
|
BRCA
|
breast invasive carcinoma
|
|
CESC
|
cervical and endocervical cancer
|
|
CHOL
|
cholangiocarcinoma
|
|
COAD
|
colon adenocarcinoma
|
|
DFS
|
disease-free survival
|
|
DLBC
|
lymphoid neoplasm diffuse large B-cell
lymphoma
|
|
ESCA
|
esophageal carcinoma
|
|
GBM
|
glioblastoma multiforme
|
|
HNSC
|
head and neck squamous cell
carcinoma
|
|
KICH
|
kidney chromophobe
|
|
KIRC
|
kidney renal clear cell carcinoma
|
|
KIRP
|
kidney renal papillary cell
carcinoma
|
|
LAML
|
acute myeloid leukemia
|
|
LGG
|
brain lower grade glioma
|
|
LIHC
|
liver hepatocellular carcinoma
|
|
LUAD
|
lung adenocarcinoma
|
|
LUSC
|
lung squamous cell carcinoma
|
|
MESO
|
mesothelioma
|
|
NSCLC
|
non-small cell lung cancer
|
|
OS
|
overall survival
|
|
OV
|
ovarian serous cystadenocarcinoma
|
|
PAAD
|
pancreatic adenocarcinoma
|
|
PCPG
|
pheochromocytoma and paraganglioma
|
|
PRAD
|
prostate adenocarcinoma
|
|
READ
|
rectal adenocarcinoma
|
|
SARC
|
sarcoma
|
|
SKCM
|
skin cutaneous melanoma
|
|
STAD
|
stomach adenocarcinoma
|
|
TAMs
|
tumor-associated macrophages
|
|
TGCT
|
testicular germ cell tumors
|
|
THCA
|
thyroid carcinoma
|
|
THYM
|
thymoma
|
|
UCEC
|
uterine corpus endometrial
carcinoma
|
|
UCS
|
uterine carcinosarcoma
|
|
UVM
|
uveal melanoma
|
References
|
1
|
Yuan H, Zhao J, Yang Y, Wei R, Zhu L, Wang
J, Ding M, Wang M and Gu Y: SHP-2 Interacts with CD81 and regulates
the malignant evolution of colorectal cancer by inhibiting
epithelial-mesenchymal transition. Cancer Manag Res.
12:13273–13284. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhou L, Feng Y, Ma YC, Zhang Z, Wu JW, Du
S, Li WY, Lu XH, Ma Y and Wang RL: Exploring the mechanism of the
potent allosteric inhibitor compound2 on SHP2WT and
SHP2F285S by molecular dynamics study. J Mol Graph
Model. 103:1078072021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mitra R and Ayyannan SR: Small-Molecule
Inhibitors of Shp2 phosphatase as potential chemotherapeutic agents
for Glioblastoma: A Minireview. ChemMedChem. 16:777–787. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li S, Wang X, Li Q and Li C: Role of
SHP2/PTPN11 in the occurrence and prognosis of cancer: A systematic
review and meta-analysis. Oncol Lett. 25:192023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim JS, Shin OR, Kim HK, Cho YS, An CH,
Lim KW and Kim SS: Overexpression of protein phosphatase
non-receptor type 11 (PTPN11) in gastric carcinomas. Dig Dis Sci.
55:1565–1569. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kerr DL, Haderk F and Bivona TG:
Allosteric SHP2 inhibitors in cancer: Targeting the intersection of
RAS, resistance, and the immune microenvironment. Curr Opin Chem
Biol. 62:1–12. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sausgruber N, Coissieux MM, Britschgi A,
Wyckoff J, Aceto N, Leroy C, Stadler MB, Voshol H, Bonenfant D and
Bentires-Alj M: Tyrosine phosphatase SHP2 increases cell motility
in triple-negative breast cancer through the activation of
SRC-family kinases. Oncogene. 34:2272–2278. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mainardi S, Mulero-Sánchez A, Prahallad A,
Germano G, Bosma A, Krimpenfort P, Lieftink C, Steinberg JD, de Wit
N, Gonçalves-Ribeiro S, et al: SHP2 is required for growth of
KRAS-mutant non-small-cell lung cancer in vivo. Nat Med.
24:961–967. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jiang C, Hu F, Tai Y, Du J, Mao B, Yuan Z,
Wang Y and Wei L: The tumor suppressor role of Src homology
phosphotyrosine phosphatase 2 in hepatocellular carcinoma. J Cancer
Res Clin Oncol. 138:637–646. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xinhua S: Role of CAD gene in the
prognosis evaluation of pan-cancer. J Basic Clinical Oncol.
33:485–90. 2020.
|
|
11
|
Zhang Z, Lin E, Zhuang H, Xie L, Feng X,
Liu J and Yu Y: Construction of a novel gene-based model for
prognosis prediction of clear cell renal cell carcinoma. Cancer
Cell Int. 20:272020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sturm G, Finotello F, Petitprez F, Zhang
JD, Baumbach J, Fridman WH, List M and Aneichyk T: Comprehensive
evaluation of transcriptome-based cell-type quantification methods
for immuno-oncology. Bioinformatics. 35:i436–i45. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Thorsson V, Gibbs DL, Brown SD, Wolf D,
Bortone DS, Ou Yang TH, Porta-Pardo E, Gao GF, Plaisier CL, Eddy
JA, et al: The Immune landscape of cancer. Immunity. 51:411–412.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gu J, Han T, Ma RH, Zhu YL, Jia YN, Du JJ,
Chen Y, Jiang XJ, Xie XD and Guo X: SHP2 promotes laryngeal cancer
growth through the Ras/Raf/Mek/Erk pathway and serves as a
prognostic indicator for laryngeal cancer. Int J Oncol. 44:481–490.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chalfant CE, Kim M, Hartman Z, Geldenhuys
WJ and Agazie YM: Novel Small-molecule inhibitor for the oncogenic
tyrosine phosphatase SHP2 with anti-breast cancer cell effects. Mol
Cancer Res. 5:25113–25124. 2020.
|
|
16
|
Vazhappilly CG, Saleh E, Ramadan W, Menon
V, Al-Azawi AM, Tarazi H, Abdu-Allah H, El-Shorbagi AN and El-Awady
R: Inhibition of SHP2 by new compounds induces differential effects
on RAS/RAF/ERK and PI3K/AKT pathways in different cancer cell
types. Invest New Drugs. 37:252–261. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ahmed TA, Adamopoulos C, Karoulia Z, Wu X,
Sachidanandam R, Aaronson SA and Poulikakos PI: SHP2 Drives
adaptive resistance to ERK signaling inhibition in molecularly
defined subsets of ERK-Dependent tumors. Cell Rep. 26:65–78.e5.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fedele C, Ran H, Diskin B, Wei W, Jen J,
Geer MJ, Araki K, Ozerdem U, Simeone DM, Miller G, et al: SHP2
inhibition prevents adaptive resistance to MEK inhibitors in
multiple cancer models. Cancer Discov. 8:1237–1249. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feng HB, Chen Y, Xie Z, Jiang J, Zhong YM,
Guo WB, Yan WQ, Lv ZY, Lu DX, Liang HL, et al: High SHP2 expression
determines the efficacy of PD-1/PD-L1 inhibitors in advanced KRAS
mutant non-small cell lung cancer. Thorac Cancer. 12:2564–2573.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lei W, Qin X, Wang X and Zhu F: Expression
and clinical significance of Shp2 protein in breast cancer. Jilin
Med J. 32:7692–7694. 2011.
|
|
21
|
Hu Z, Li J, Gao Q, Wei S and Yang B: SHP2
overexpression enhances the invasion and metastasis of ovarian
cancer in vitro and in vivo. Onco Targets Ther. 10:3881–3891. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liotti F, Kumar N, Prevete N, Marotta M,
Sorriento D, Ieranò C, Ronchi A, Marino FZ, Moretti S, Colella R,
et al: PD-1 blockade delays tumor growth by inhibiting an intrinsic
SHP2/Ras/MAPK signalling in thyroid cancer cells. J Exp Clin Cancer
Res. 40:222021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao Y, Yu H, Ida CM, Halling KC, Kipp BR,
Geiersbach K, Rumilla KM, Gupta S, Lin MT and Zheng G: Assessment
of RAS dependency for BRAF alterations using cancer genomic
databases. JAMA Netw Open. 4:e20354792021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan X, Bu H and Zhou J: Recent advances
of SHP2 inhibitors in cancer therapy: Current development and
clinical application. J Med Chem. 63:11368–11396. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim M, Morales LD and Jang IS: Protein
tyrosine phosphatases as potential regulators of STAT3 signaling.
Int J Mol Sci. 19:27082018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang W, Chan RJ, Chen H, Yang Z, He Y,
Zhang X, Luo Y, Yin F, Moh A, Miller LC, et al: Negative regulation
of Stat3 by activating PTPN11 mutants contributes to the
pathogenesis of Noonan syndrome and juvenile myelomonocytic
leukemia. J Biol Chem. 284:22353–22363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang J, Pollard K, Allen AN, Tomar T,
Pijnenburg D, Yao Z and Rodriguez FJ: Combined inhibition of SHP2
and MEK is effective in models of NF1-Deficient malignant
peripheral nerve sheath tumors. Cancer Res. 80:5367–5379. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu G, Xie J, Kong W, Xie J, Li Y, Du L,
Zheng Q, Sun L, Guan M, Li H, et al: Phase separation of
disease-associated SHP2 mutants underlies MAPK hyperactivation.
Cell. 183:490–502.e18. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gong M, Liu P, He T, Zhang M and Li G:
Demonstrating the effect of SHP2 inhibitor on cervical squamous
cell carcinoma from the perspective of ZAP70. Anticancer Drugs.
32:477–483. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cassetta L and Pollard JW: Targeting
macrophages: Therapeutic approaches in cancer. Nat Rev Drug Discov.
17:887–904. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xiao P, Zhang H, Zhang Y, Zheng M, Liu R,
Zhao Y, Zhang X, Cheng H, Cao Q and Ke Y: Phosphatase Shp2
exacerbates intestinal inflammation by disrupting macrophage
responsiveness to interleukin-10. J Exp Med. 216:337–349. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hui E, Cheung J, Zhu J, Su X, Taylor MJ,
Wallweber HA, Sasmal DK, Huang J, Kim JM, Mellman I and Vale RD: T
cell costimulatory receptor CD28 is a primary target for
PD-1-mediated inhibition. Science. 355:1428–1433. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Torres-Ayuso P and Brognard J: Shipping
out MEK inhibitor resistance with SHP2 inhibitors. Cancer Discov.
8:1210–1212. 2018. View Article : Google Scholar : PubMed/NCBI
|