Introduction
Breast cancer is one of the most common cancers
among women globally and its incidence rate is increasing annually
(1). Breast cancer is the second
leading cause of cancer deaths among women (2). Breast cancer is a metastatic cancer
that initially spreads through the lymphatic system to various
levels of lymph nodes and can then metastasize to distant organs
through the bloodstream, such as the bones, liver, lungs and brain,
which contributes to the difficulties in effectively treating this
disease (3). Accurate metastasis
prediction is related to the formulation of treatment plans,
prognosis, survival rate and quality of life (4). However, breast cancer is a highly
heterogeneous malignant disease with various functional phenotypes
(5). Previous proteomic, genomic
and transcriptomic studies have explored different cellular
subtypes and the development of breast cancer lymphatic metastasis
biomarkers (6,7). Therefore, finding protein biomarkers
related to breast cancer is important for predicting the
progression of the disease, implementing early drug or intervention
treatment and thus reducing mortality caused by metastasis.
The process of lymphatic metastasis involves the
lymphatic circulation and tumor-related lymphatic vessels provide a
direct route to the lymph nodes, which enables primary tumors to
transmit cytokine signals, gradually reshaping and hijacking lymph
node function from afar (8). After
a tumor develops, various aspects of the internal tumor environment
will undergo complex biological changes (9). Lymph fluid, an essential component of
the lymph node environment, is subject to these changes, with
variations occurring in its component cytokines and antibodies; for
example, programmed death ligand 1 and tumor-infiltrating
lymphocytes (10). However, due to
the difficulties associated with obtaining lymph fluid [due to
individualized vessel routing and the technical difficulty of
extraction via contrast-enhanced ultrasound (CEUS)], this issue has
been under-researched to date. Percutaneous CEUS technology can
provide support in this area (11),
enabling observation of the lymphatic vessels entering and leaving
the sentinel lymph nodes and assisting in the accurate extraction
of lymph fluid. Therefore, in the present study, CEUS technology
was used to precisely extract lymph fluid from rabbits with breast
cancer metastasis and healthy rabbits.
Materials and methods
Establishment of the rabbit breast
cancer model
Purebred New Zealand white rabbits (male, n=1;
female, n=6; 6 months old), weighing 2.0–2.5 kg were purchased from
the Experimental Animal Center of Ningxia Medical University.
Maintenance conditions for the rabbits were as follows:
Temperature, 18–29°C; relative humidity, 40–70%; noise, ≤60 dB;
animal illumination, 100–001×; and food and water, sufficient food
and water sources to meet the physiological needs of the rabbits.
The VX2 rabbit mesenchymal squamous cell carcinoma tumor cell line
(cat. no. RP-0097) was purchased from Shanghai Yinxi Biotechnology
Co., Ltd. Cells were cultured in RPMI 1640 culture medium. The
ratio was basic culture medium:serum (FBS) at 9:1, and the double
antibody was added at 1% volume. Cells were cultivated in a 5%
CO2 incubator at 37°C. The aforementioned reagents were
purchased from Thermo Fisher Scientific, Inc. To establish the
tumor model, 1 ml of a 1×107 cells/ml suspension of VX2
cells was injected into the lateral muscles of the hind legs of the
male rabbit. After 2 weeks, a solid mass was formed. Animals were
euthanized by an intravenous injection of pentobarbital sodium (100
mg/kg) into tumor bearing rabbits. Under sterile conditions, the
solid tumor from the hind leg of the tumor bearing rabbit was
removed, washed with physiological saline and placed in a glass
dish containing 20 ml of RPMI 1640 solution. A fish-shaped piece of
tissue (~5×5×5 mm in size) with vigorous growth at the edge of the
tumor was cut into tissue blocks with a diameter of 0.5–1.0 mm
using ophthalmic scissors. The tumor blocks were mixed well and
placed with 10 ml RPMI1640 solution into a 20 ml syringe to make a
tissue block suspension. Tissues were placed on ice until used.
Three female rabbits were held in a supine position using a fixator
and administered inhalant isoflurane anesthesia (induction
concentration, 2%; maintenance concentration, 1.5%) for 2 min. Once
the corneal reflex weakened, muscles relaxed or breathing slowed
down, the injection was performed. A 10 ml volume of tissue block
suspension was injected under the breast pad of the second nipple
on the left side of the rabbit chest wall. After 2 weeks, the
rabbits were observed to have a 100% tumor formation rate and 1
tumor per animal. Humane endpoints were as follows: During the
experiment, if any uncontrollable pain or other conditions were
found in the animal, euthanasia would be carried out promptly.
During the experiment, the size of the tumor was observed and if
the tumor volume exceeded 10% of the experimental rabbit's body
weight, the experiment would be terminated in a timely manner. The
in vivo experiments performed in the present study were
approved by the Medical Research Ethics Review Committee of Ningxia
Medical University General Hospital (approval no.
KYLL-2022-0173).
Precision extraction of lymph
fluid
A total of three healthy rabbits and three rabbits
with lymph node metastasis of breast cancer were analyzed.
Ultrasound contrast agent (Sonazoid; GE Healthcare) was injected
subcutaneously at the edge of the tumor in the breast cancer model
rabbits. The first (group) lymph node traced from the injection
point along the lymphatic vessel was the sentinel lymph node (SLN).
CEUS can clearly display the input lymphatic vessels of the SLN,
which may reflect the state of the lymph fluid after tumor
microenvironment formation (11,12).
CEUS can used to dynamically observe the progression of lymph node
metastasis. Lymph fluid from the rabbit's SLN at 8 weeks after
tumor implantation was used to ensure that the metastatic lymph
node was malignant. Accurate lymph fluid extraction from normal
rabbits under CEUS guidance was performed as a control. This
process occurred once every 2 days and lasted for 2 weeks, with 10
µl of lymph extracted each time. During the extraction process,
animals were anaesthetized through the inhalation of isoflurane
(induction concentration, 2%; maintenance concentration, 1.5%) for
3 min. After the experiment was completed, all rabbits were
euthanized by intravenous injection of pentobarbital sodium (100
mg/kg). The criteria for determining death were: No breathing or
pulse in the rabbit, no heartbeat lasting for more than 5 min when
auscultating or touching the chest with a stethoscope, rabbit
corneal reflex disappeared, pupil dilation occurred and nerve
reflexes ceased. At the end of the experiment, the maximum tumor
diameter of both female and male rabbits did not exceed 4 cm.
Reagents and equipment
Ammonium bicarbonate buffer
(NH4HCO3; pH 8.0), dithiothreitol (DTT),
iodoacetamide (IAA) and sodium carbonate were purchased from
MilliporeSigma. Urea, SDS and the BCA protein assay kit were
purchased from Bio-Rad Laboratories, Inc. Trypsin was purchased
from Promega Corporation and the Q Exactive™ Plus mass spectrometer
and EASY-nLC™ 1200 were purchased from Thermo Fisher Scientific,
Inc.
Label-free quantification (LFQ) sample
preparation
After freeze-drying the lymph fluid, each sample was
mixed with 100–200 µl SDT lysis buffer [4% sodium dodecyl sulfate,
100 mM dithiothreitol, 100 mM Tris-HCl (pH 8.0); Bio-Rad
Laboratories, Inc.] based on the volume of lymph fluid obtained
(2:1 ratio). Lysate was homogenized and transferred to an
Eppendorf® tube. The lysate was then incubated in a
boiling water bath (100°C) for 3 min, sonicated for 2 min (50 W,
60°C for 2 sec), centrifuged at 16,000 × g and 4°C for 20 min and
the supernatant was collected. The BCA method was used for protein
quantification. A total of 100 µg protein was taken from each
sample for filter-aided sample preparation digestion. The following
steps were performed: DTT (1 M to a final concentration of 100 mM)
was added to each sample followed by incubation in a boiling water
bath (100°C) for 5 min, after which the samples were cooled to room
temperature. A 200 µl volume of urea buffer (8 M urea; 150 mM
Tris-HCl; pH 8.0) was added to each sample and mixed before
transferring to a 10 kDa ultrafiltration centrifuge tube and
centrifuged at 12,000 × g for 15 min at 4°C. UA buffer (200 µl) was
added to samples and centrifuged at 12,000 × g for 15 min at 4°C
before the filtrate was discarded. A 100 µl volume of IAA (50 mM
IAA in UA) was added and samples were shaken at 600 rpm for 1 min.
Next, samples were kept in the dark at room temperature for 30 min,
then centrifuged at 12,000 × g for 10 min at 4°C. UA buffer (100
µl) was added and samples were centrifuged at 12,000 × g for 10 min
at 4°C and repeated twice. A 100-µl volume of
NH4HCO3 buffer was added and samples were
centrifuged at 14,000 × g for 10 min at 4°C and repeated twice. A
40 µl volume of trypsin buffer (6 µg trypsin in 40 µl
NH4HCO3 buffer) was added to each sample
before shaking at 600 rpm for 1 min and incubating at 37°C for
16–18 h. Samples were moved to a new collection tube and
centrifuged at 12,000 × g for 10 min at 4°C. The filtrate was
collected and an appropriate volume (100 µl) of 0.1%
trifluoroacetic acid (TFA) solution was added. Then, the digested
peptides were desalted using the C18 Cartridge (Bio-Rad
Laboratories, Inc.) and freeze-dried under vacuum. After drying,
the digested peptides were reconstituted with 0.1% TFA. The peptide
concentration was measured and samples were prepared for liquid
chromatography with tandem mass spectrometry (LC-MS/MS)
analysis.
LC-MS/MS analysis
Chromatographic separation of peptide samples was
performed using a nanoflow Easy nLC 1200 chromatography system
(Thermo Fisher Scientific, Inc.). The buffer solutions were
prepared as follows: Buffer A was a 0.1% formic acid aqueous
solution and buffer B was a formic acid (0.1%), acetonitrile (80%)
and water solution. The chromatographic column was equilibrated
with 95% buffer A solution. The sample was injected into a trap
column (100 µm; 20 mm; 5 µm; C18; Dr. Maisch HPLC GmbH) and
separated by a gradient through the chromatographic analysis column
(75 µm; 150 mm; 3 µm; C18; Dr. Maisch HPLC GmbH) at a flow rate of
300 nl/min. The liquid phase separation gradient was as follows:
0–2 min, linear gradient of buffer B solution from 5–8%; 2–90 min,
linear gradient of buffer B solution from 8–23%; 90–100 min, linear
gradient of buffer B solution from 23–40%; 100–108 min, linear
gradient of buffer B solution from 40–100%; 108–120 min, buffer B
solution was maintained at 100%. After peptide separation, a
Q-Exactive HF-X mass spectrometer (Thermo Fisher Scientific, Inc.)
was used for data-dependent acquisition mass spectrometry analysis.
The analysis time was 120 min, the detection mode was positive
ionization, the parent ion scan range was 300–1,800 m/z, the
primary mass spectrometry resolution was 60,000 at m/z 200, the AGC
target was 3×106 and primary mass spectrometry maximum
ion time (IT) was 50 msec. Peptide secondary mass spectrometry was
collected according to the following methods: After each full scan,
20 secondary mass spectra (MS2 scans) of the highest intensity
parent ions were collected, with a secondary mass spectrometry
resolution of 15,000 at m/z 200, an AGC target of 1×105,
secondary mass spectrometry maximum IT of 50 msec, MS2 activation
type of HCD, an isolation window of 1.6 m/z and a normalized
collision energy of 28.
Database retrieval
The resulting LC-MS/MS raw files were imported into
the Proteome Discoverer software (version 2.4; Thermo Fisher
Scientific, Inc.) and the search engine Sequest HT was used for
database retrieval. The database used for searching was
uniprot-Oryctolagus cuniculus (Rabbit) [9986]-43526-20211222.fasta,
which was sourced from the Uniprot protein database (https://www.uniprot.org/taxonomy/9986),
with a protein entry of 43,526 and a download date of
22.12.2021.
Parallel reaction monitoring (PRM)
sample preparation
Urea (8 M) was added to 200-µl samples, which were
then sonicated in an ice bath and centrifuged at 4°C and 16,000 × g
for 20 min to collect the supernatant. BCA quantification was
performed on the supernatant and 15 µg of each sample was run on a
1% agarose gel. DTT (final concentration 10 mM) was added to 200 µg
of each sample and incubated at 37°C for 1 h. IAA was added to a
final concentration of 50 mM and samples were incubated in darkness
for 30 min at 25°C. Trypsin was added to each sample (1:50) and
incubated at 37°C overnight. After quantitative desalting, samples
were mixed in equal volumes for testing.
LC-PRM/MS analysis
A total of 2 µg of peptide from each sample was used
for LC-PRM/MS analysis. After sample loading, chromatographic
separation was performed using a nanoflow Easy nLC1200
chromatography system (Thermo Fisher Scientific, Inc.). The liquid
phase separation gradient was as follows: 0–5 min, linear gradient
of buffer B solution from 2–5%; 5–45 min, the linear gradient of
buffer B solution from 5–23%; 45–50 min, linear gradient of buffer
B solution from 23–40%; 50–52 min, linear gradient of buffer B
solution from 40–100%; and 52–60 min, buffer B solution was
maintained at 100%. The Q Exactive HF-X mass spectrometer (Thermo
Fisher Scientific, Inc.) was used for targeted PRM mass
spectrometry analysis. The original PRM raw data files of the mass
spectra obtained were analyzed using Skyline software (version 4.1)
(13).
Bioinformatics analysis
Proteins with fold-change (FC) >1.5 or <0.667
and P<0.05 were considered as significantly differentially
expressed proteins. Bioinformatics data were analyzed using Perseus
(14), Microsoft Excel (Microsoft
Corporation) and R (version 4.0.3; RStudio, Inc.) statistical
computing software. Sequence annotations were extracted from
UniProtKB/Swiss-Prot, Kyoto Encyclopedia of Genes and Genomes
(KEGG) and Gene Ontology (GO) databases. GO and KEGG enrichment
analyses were performed using Fisher's exact test and false
discovery rate adjustment for multiple testing. GO terms were
divided into three categories: Biological processes (BP), molecular
functions (MF) and cellular components (CC). Enriched GO and KEGG
pathways were classed as statistically significant if
P<0.05.
Statistical analysis
Data were analyzed using SPSS software (version
20.0; IBM Corp.). Measurement data were presented as the mean ±
standard deviation of three replicates for each group and the
unpaired t-test was used for inter-group comparisons. Count data
were presented as percentages and inter-group differences were
compared using the χ2 test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Observation of breast cancer lymph
node metastasis under CEUS guidance
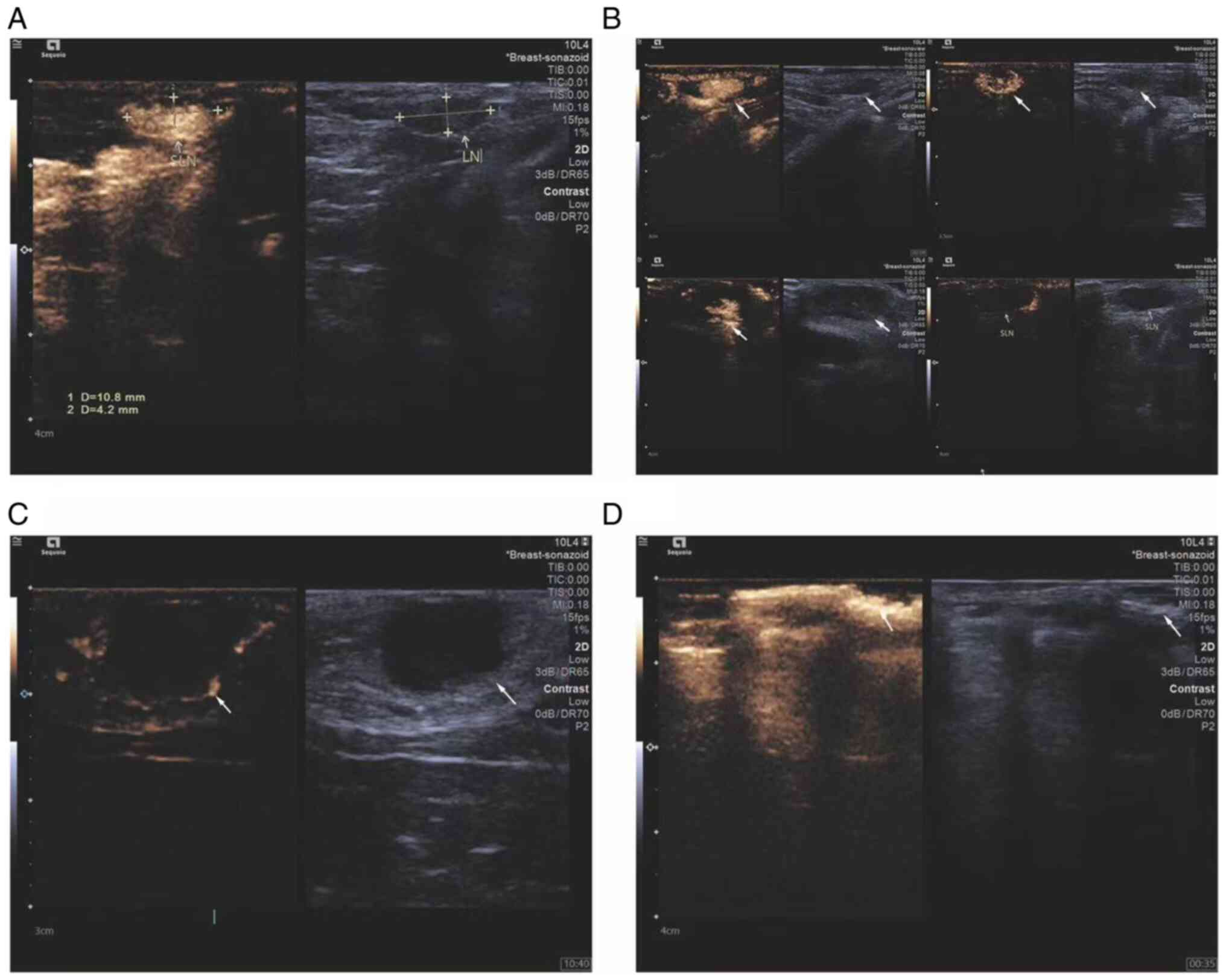
CEUS was used to observe the changes in sentinel
lymph nodes during the metastasis process of the breast cancer
lymphatic metastasis group of rabbits. Percutaneous superficial
ultrasound with Sonozoid demonstrated four contrast patterns in
sentinel lymph nodes during breast cancer progression (Fig. 1): i) Uniform enhancement; ii)
non-uniform enhancement; iii) peripheral ring enhancement with no
internal enhancement; and iv) complete absence of enhancement in
the lymph node with the presence of surrounding lymphatic vessels
bypassing. To ensure that the extracted lymph originated from
metastatic lymph nodes, the status of rabbit lymph nodes using CEUS
was observed and ultrasound-guided lymph extraction was performed
when there was no enhancement under contrast-enhanced ultrasound.
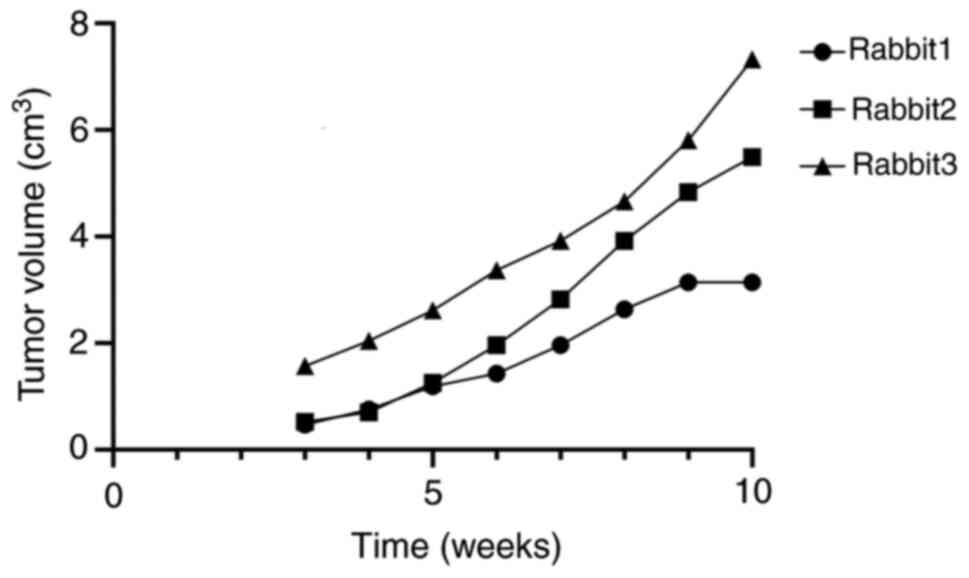
During the experiment, the volume, maximum diameter and minimum
diameter of the rabbit tumor were measured and recorded (Fig. 2) (Table
I).
 | Table I.Maximum diameter of tumors observed in
rabbits at the end of the experiment. |
Table I.
Maximum diameter of tumors observed in
rabbits at the end of the experiment.
| Animal | Long diameter of
tumor, cm | Short diameter of
tumor, cm |
|---|
| Male rabbit | 2.0 | 1.5 |
| Female rabbit
1 | 3.0 | 2.0 |
| Female rabbit
2 | 3.5 | 3.0 |
| Female rabbit
3 | 4.0 | 3.0 |
Analysis of lymphatic fluid
differentially expressed proteins
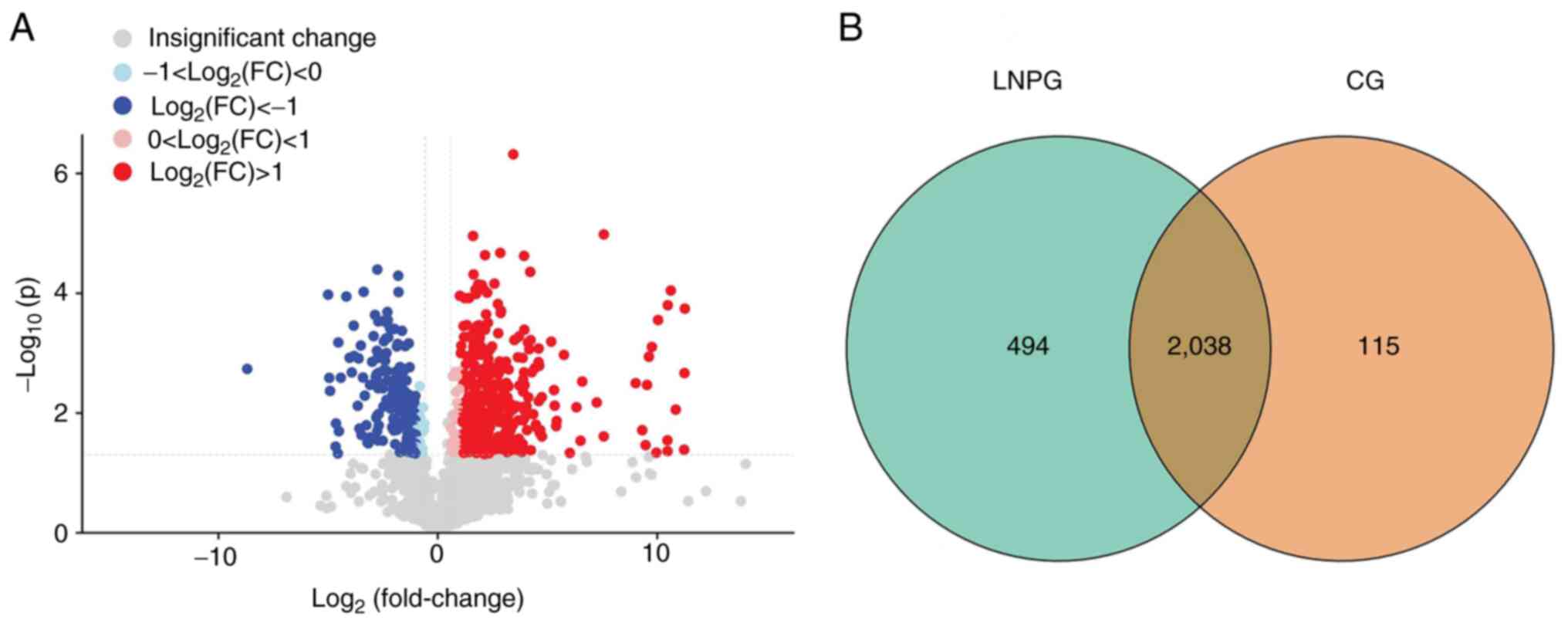
A total of 2,647 proteins were detected in six
samples using the LFQ method. Using a 1.2-fold increase or decrease
in protein expression as the criterion for significant changes, 547
differentially expressed proteins were identified. Compared with
the normal group (CG), the metastasis group (LNPG) demonstrated 371
upregulated proteins and 176 downregulated proteins (Fig. 3).
Differentially expressed protein GO
functional enrichment analysis
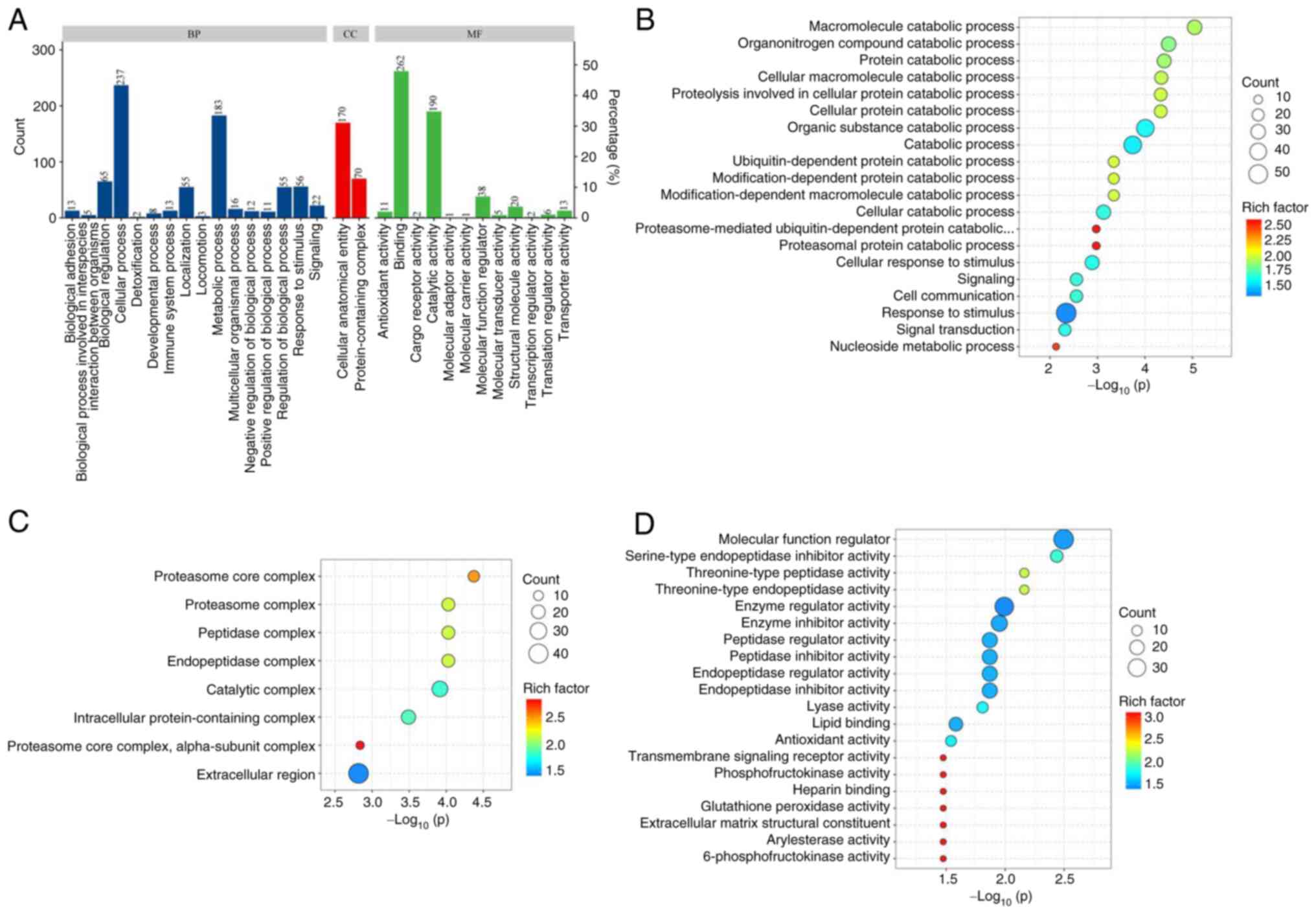
GO functional enrichment analysis was conducted on
the differentially expressed proteins in the lymphatic fluid of
breast cancer rabbits and normal rabbits (Fig. 4A). The main BPs included
‘macromolecule catabolic process’, ‘organonitrogen compound
catabolic process’ and ‘protein catabolic process’ (Fig. 4B). The main CCs included ‘proteasome
core complex’, ‘proteasome complex’, ‘peptidase complex’,
‘endopeptidase complex’ and ‘catalytic complex’ (Fig. 4C). The main MFs included ‘molecular
function modulator’, ‘serine-type endopeptidase inhibitor activity’
and ‘threonine-type peptidase activity’ (Fig. 4D).
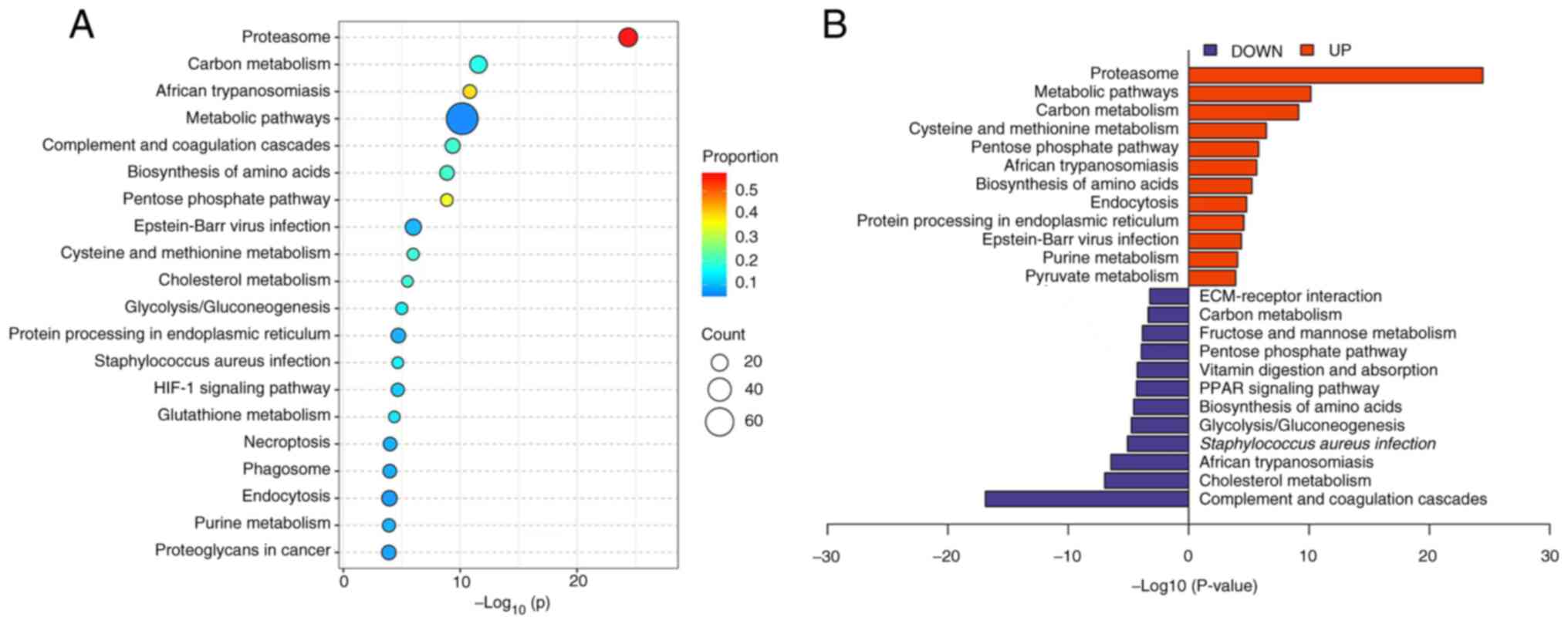
Differentially expressed protein KEGG
enrichment analysis
KEGG analysis was used to subject the differentially
expressed proteins to pathway enrichment analysis. It was
demonstrated that the differentially expressed proteins were
involved in 20 main pathways (Fig.
5A), which included the proteasome pathway, complement and
coagulation cascades pathway and pentose phosphate pathway. The
upregulated proteins were mainly involved in pathways such as the
proteasome pathway, carbon metabolism pathway and cysteine and
methionine metabolism pathway. The downregulated proteins were
mainly involved in pathways such as the complement and coagulation
cascade pathway, cholesterol metabolism pathway and
Staphylococcus aureus infection pathway (Fig. 5B).
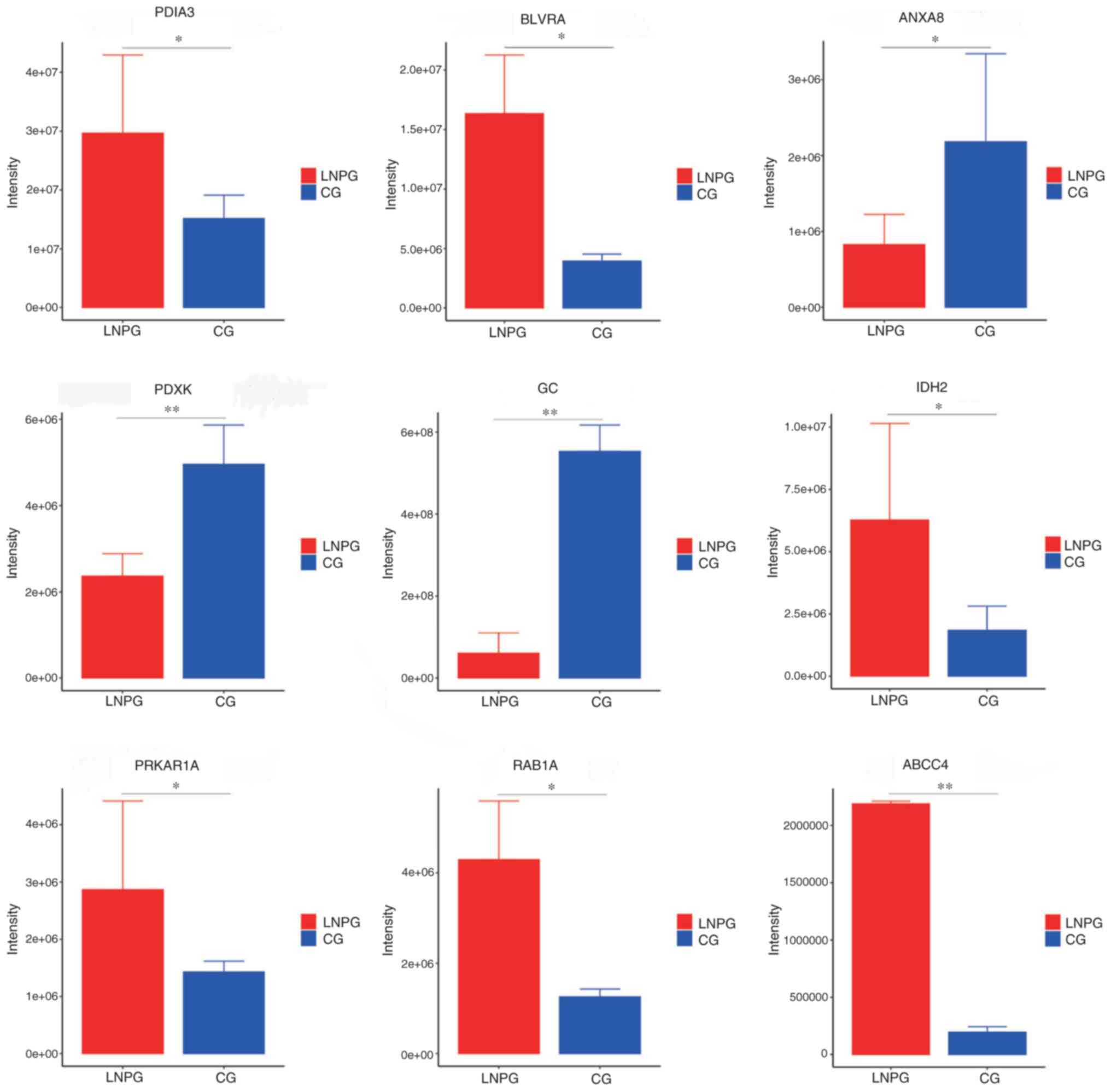
PRM validation
PRM was conducted on the differentially expressed
proteins in the lymphatic fluid of the metastasis group and healthy
group of rabbits. A total of 10 differentially expressed proteins
were demonstrated to have consistent trends with the results from
the LFQ proteomics technology assay (Table II). The upregulated proteins
included protein disulfide-isomerase 3 (PDIA3), biliverdin
reductase A, isocitrate dehydrogenase 2, protein kinase
cAMP-dependent type I regulatory subunit α (PRKAR1A), Ras-related
protein Rab-1A, ATP-binding cassette sub-family C member 4 (ABCC4)
and microtubule-associated protein 4, while the downregulated
proteins included annexin A8, pyridoxal kinase and envelope
glycoprotein C (Fig. 6).
| Table II.Label-free quantification proteomic
validation of differentially expressed proteins in the normal and
metastasis groups. |
Table II.
Label-free quantification proteomic
validation of differentially expressed proteins in the normal and
metastasis groups.
| Gene | Protein | NCBI accession
no. | Peptide
sequence | Score | Fold-change | P-value |
|---|
| PDIA3 | Protein
disulfide-isomerase | B7NZF1 | ELSDFISYLQR | 187.53 | 0.62 | <0.05 |
| BLVRA | Biliverdin
reductase A | G1SRZ6 | FGFPAFSGISR | 21.76 | 1.88 | <0.05 |
| IDH2 | Isocitrate
dehydrogenase (NADP) | G1SZF7 |
LNEHFLNTTDFLDTIK | 130.9 | 2.35 | <0.05 |
| PRKAR1A | Protein kinase
cAMP-dependent type I regulatory subunit alpha | G1TDN4 | NVLFSHLDDNER | 48.67 | 6.97 | <0.05 |
| RAB1A | RAB1A, member RAS
oncogene family | G1TCS8 | QWLQEIDR | 61.72 | 1.78 | <0.05 |
| ABCC4 | ATP binding
cassette subfamily C member 4 | A0A5F9CTH3 | SFAELIASLR | 59.42 | 0.97 | <0.05 |
| ANXA8 | Annexin | G1T6W4 | GAGTLDGTLIR | 141.5 | 7.97 | <0.01 |
| PDXK | Pyridoxal
kinase | G1UZB5 |
GQVLTSDELHELHELYEGLR | 21.46 | 4.49 | <0.05 |
| GC | Gc-globulin | G1SU8Z | HLSLLTTLSNR | 628.2 | 2.35 | <0.05 |
Discussion
Lymphatic metastasis is a major factor affecting the
prognosis of various cancers, including breast cancer (15). Although cancer cells spread in a
number of different ways, the structure of the lymphatic system
makes it the prime site of cancer metastasis (16). Despite the current understanding of
the pathophysiology of breast cancer, the molecular mechanisms,
especially those related to lymphatic metastasis, are still
unclear. This may be due to the difficulty in obtaining lymphatic
fluid. Identifying differentially expressed proteins and signaling
pathways involved in breast cancer metastasis is critical to
understanding its mechanism and exploring biomarkers related to
metastasis. In the present study, CEUS technology was used to
extract lymphatic fluid for comparative analysis, quantitative
evaluation and functional identification of LFQ proteomics and
validated differentially expressed proteins with PRM
technology.
In the present study, using LC-MS/MS analysis,
proteins in the lymph fluid were quantified and bioinformatics
analysis was performed on differentially expressed proteins to
identify protein biomarkers for breast cancer lymphatic metastasis
and explore the function of differentially expressed proteins and
the signaling pathways involved. The present study laid the
foundation for further exploration of predictive indicators and
therapeutic targets for breast cancer metastasis in the future. As
breast cancer develops, the tumor microenvironment inevitably
changes, as does the body's metabolic environment. For example,
amino acids are essential nutrients for all living cells and are
vital for the proliferation and maintenance of tumor cells. Since
tumor cells grow faster compared with normal cells, tumor cells
have a higher demand for amino acids (17). Previous studies have reported that
some amino acid metabolic pathways, such as glutamine, serine,
glycine and proline pathways, are altered in breast cancer, which
suggests that amino acid transport may be crucial for the
proliferation and progression of breast cancer (18–20).
At the same time, mechanisms of hypoxia-adaptive metabolic
responses (21), including
increased glycolysis and decreased tricarboxylic acid cycle, serve
to reduce the production of mitochondrial reactive oxygen species
(22).
The present study identified upregulated proteins
PDIA3, PRKAR1A and ABCC4 in the lymphatic fluid of metastatic
breast cancer rabbits using LC-MS/MS. PDIA3, also known as ERp57,
is a 58 kDa glucose-regulated protein that also acts as a
chaperone, modifying and folding proteins, and has redox functions
(23,24). PDIA3 serves a role in the quality
control of newly synthesized glycoproteins, participates in the
assembly of major histocompatibility complex class I molecules and
regulates immune responses and immunogenic cell death (25). Despite, to the best of our
knowledge, no reports of a direct link between PDIA3 and breast
cancer development to date, PDIA3 has previously been reported to
be upregulated in various cancers and is involved in cancer
initiation, progression and chemosensitivity, which suggests its
potential as a cancer biomarker and therapeutic target (26–28).
PRKAR1A is a gene that directs the synthesis of protein kinase A
(PKA) regulatory subunits and is one of the critical components of
the PKA tetramer (29). It is the
primary mediator of cAMP function in various mammalian cellular
processes, including cell differentiation, proliferation and
apoptosis (30). PRKAR1A
inactivating mutations can lead to Carney syndrome, which is
characterized by cardiac myxoma and multiple endocrine tumors
(31). Therefore, PRKAR1A mutations
may serve as a predictor of breast cancer metastasis. ABCC4 is the
fourth member of the ATP-binding cassette protein C subfamily, also
known as multidrug resistance-associated protein 4 (MRP4). ABCC4
was discovered due to its role in mediating drug resistance in
various tumor types (32). High
expression levels of ABCC4 have been reported in drug-resistant
tumors, including neuroblastoma, prostate cancer, pancreatic cancer
and acute myeloid leukemia, where MRP4 expression is associated
with poor prognosis (33–35). Thus, the increased drug resistance
in breast cancer may be linked to the high expression levels of
ABCC4.
However, the technical difficulty in accurately
extracting lymphatic fluid using CEUS limits its application to
animal research. Additionally, proteomics technology has high
economic costs, making it challenging to promote in clinical
practice.
In conclusion, the present study employed CEUS
technology for lymphatic fluid extraction, as well as proteomic and
mass spectrometry analysis techniques to investigate differentially
expressed proteins and related functions in metastatic breast
cancer rabbits. The up-regulated or down-regulated proteins and
multiple enrichment pathways identified in the present study were
related to the pathophysiological process of the occurrence and
development of breast cancer. The data provided by the present
study may provide new ideas for future follow-up research on new
metastasis targets of breast cancer.
Acknowledgments
The authors would like to thank Shanghai Bioprofile
(Shanghai, China) for providing technical support by performing the
proteomic testing and analysis.
Funding
The present study was supported by the National Science
Foundation of China (grant no. 82260351).
Availability of data and materials
The datasets used in the present study can be found
in online repositories. The LC-MS/MS data was uploaded to iProX
(accession no. IPX0007648000).
Authors' contributions
JX, XZ, GY, WS, WW and CM designed the research
concept. WW and CM coordinated the study. Lymph extraction was
performed by JX, XZ, GY and WS. XZ conducted the statistical
analysis. JX is responsible for the language editing of this
manuscript and writing the main manuscript. JX, XZ, GY, WS, WW and
CM participated in the planning and supervision of the work, as
well as in the interpretation of data. JX and XZ confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
All experimental animals in the present study were
provided by the Experimental Animal Center of Ningxia Medical
University and all experimental procedures met the review standards
of the Medical Research Ethics Review Committee of Ningxia Medical
University General Hospital (approval no. KYLL-2022-0173).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD, Wagle NS and Jemal
A: Cancer statistics, 2023. CA Cancer J Clin. 73:17–48. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun YS, Zhao Z, Yang ZN, Xu F, Lu HJ, Zhu
ZY, Shi W, Jiang J, Yao PP and Zhu HP: Risk factors and preventions
of breast cancer. Int J Biol Sci. 13:1387–1397. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Park M, Kim D, Ko S, Kim A, Mo K and Yoon
H: Breast cancer metastasis: Mechanisms and therapeutic
implications. Int J Mol Sci. 23:68062022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gouri A, Benarba B, Dekaken A, Aoures H
and Benharkat S: Prediction of late recurrence and distant
metastasis in early-stage breast cancer: Overview of current and
emerging biomarkers. Curr Drug Targets. 21:1008–1025. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yeo SK and Guan JL: Breast cancer:
Multiple subtypes within a tumor? Trends Cancer. 3:753–760. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mani DR, Krug K, Zhang B, Satpathy S,
Clauser KR, Ding L, Ellis M, Gillette MA and Carr SA: Cancer
proteogenomics: Current impact and future prospects. Nat Rev
Cancer. 22:298–313. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Y, Kong X, Wang Z and Xuan L: Recent
advances of transcriptomics and proteomics in triple-negative
breast cancer prognosis assessment. J Cell Mol Med. 26:1351–1362.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
du Bois H, Heim TA and Lund AW:
Tumor-draining lymph nodes: At the crossroads of metastasis and
immunity. Sci Immunol. 6:eabg35512021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Arneth B: Tumor microenvironment. Medicina
(Kaunas). 56:152019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rizzo A and Ricci AD: Biomarkers for
breast cancer immunotherapy: PD-L1, TILs, and beyond. Expert Opin
Investig Drugs. 31:549–555. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cui Q, Dai L, Li J and Xue J: Accuracy of
CEUS-guided sentinel lymph node biopsy in early-stage breast
cancer: A study review and meta-analysis. World J Surg Oncol.
18:1122020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Karaman S and Detmar M: Mechanisms of
lymphatic metastasis. J Clin Invest. 124:922–928. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
MacLean B, Tomazela DM, Shulman N,
Chambers M, Finney GL, Frewen B, Kern R, Tabb DL, Liebler DC and
MacCoss MJ: Skyline: An open source document editor for creating
and analyzing targeted proteomics experiments. Bioinformatics.
26:966–968. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tyanova S, Temu T, Sinitcyn P, Carlson A,
Hein MY, Geiger T, Mann M and Cox J: The Perseus computational
platform for comprehensive analysis of (prote)omics data. Nat
Methods. 13:731–740. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Das S and Skobe M: Lymphatic vessel
activation in cancer. Ann N Y Acad Sci. 113:235–241. 2008.
View Article : Google Scholar
|
|
16
|
Liu Z, Wang R, Zhou J, Zheng Y, Dong Y,
Luo T, Wang X and Zhan W: Ultrasound lymphatic imaging for the
diagnosis of metastatic central lymph nodes in papillary thyroid
cancer. Eur Radiol. 31:8458–8467. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cha YJ, Kim ES and Koo JS: Amino acid
transporters and glutamine metabolism in breast cancer. Int J Mol
Sci. 19:9072018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
El Ansari R, McIntyre A, Craze ML, Ellis
IO, Rakha EA and Green AR: Altered glutamine metabolism in breast
cancer; subtype dependencies and alternative adaptations.
Histopathology. 72:183–190. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim S, Kim DH, Jung WH and Koo JS:
Expression of glutamine metabolism-related proteins according to
molecular subtype of breast cancer. Endocr Relat Cancer.
20:339–348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Craze ML, Cheung H, Jewa N, Coimbra NDM,
Soria D, El-Ansari R, Aleskandarany MA, Wai Cheng K, Diez-Rodriguez
M, Nolan CC, et al: MYC regulation of glutamine-proline regulatory
axis is key in luminal B breast cancer. Br J Cancer. 118:258–265.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Semenza GL: Hypoxia-inducible factors:
Coupling glucose metabolism and redox regulation with induction of
the breast cancer stem cell phenotype. EMBO J. 36:252–259. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
de Heer EC, Jalving M and Harris AL: HIFs,
angiogenesis, and metabolism: Elusive enemies in breast cancer. J
Clin Invest. 130:5074–5087. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chichiarelli S, Altieri F, Paglia G,
Rubini E, Minacori M and Eufemi M: ERp57/PDIA3: New insight. Cell
Mol Biol Lett. 27:122022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hettinghouse A, Liu R and Liu CJ:
Multifunctional molecule ERp57: From cancer to neurodegenerative
diseases. Pharmacol Ther. 181:34–48. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Song D, Liu H, Wu J, Gao X, Hao J and Fan
D: Insights into the role of ERp57 in cancer. J Cancer.
12:2456–2464. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park S, Park JH, Jung HJ, Jang JH, Ahn S,
Kim Y, Suh PG, Chae S, Yoon JH, Ryu SH and Hwang D: A secretome
profile indicative of oleate-induced proliferation of HepG2
hepatocellular carcinoma cells. Exp Mol Med. 50:1–14. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen Y, Lin MC, Wang H, Chan CY, Jiang L,
Ngai SM, Yu J, He ML, Shaw PC, Yew DT, et al: Proteomic analysis of
EZH2 downstream target proteins in hepatocellular carcinoma.
Proteomics. 7:3097–3104. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li S, Zhao X, Chang S, Li Y, Guo M and
Guan Y: ERp57-small interfering RNA silencing can enhance the
sensitivity of drug-resistant human ovarian cancer cells to
paclitaxel. Int J Oncol. 54:249–260. 2019.PubMed/NCBI
|
|
29
|
Czarnecka AM, Synoradzki K, Firlej W,
Bartnik E, Sobczuk P, Fiedorowicz M, Grieb P and Rutkowski P:
Molecular Biology of Osteosarcoma. Cancers (Basel). 12:21302020.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pitsava G, Stratakis CA and Faucz FR:
PRKAR1 and thyroid tumors. Cancers (Basel). 13:38342021. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kirschner LS: PRKAR1A and the evolution of
pituitary tumors. Mol Cell Endocrinol. 326:3–7. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kochel TJ and Fulton AM: Multiple drug
resistance-associated protein 4 (MRP4), prostaglandin transporter
(PGT), and 15-hydroxyprostaglandin dehydrogenase (15-PGDH) as
determinants of PGE2 levels in cancer. Prostaglandins Other Lipid
Mediat. 116–117. 99–103. 2015.PubMed/NCBI
|
|
33
|
Huynh T, Norris MD, Haber M and Henderson
MJ: ABCC4/MRP4: A MYCN-regulated transporter and potential
therapeutic target in neuroblastoma. Front Oncol. 2:1782012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rasmuson A, Kock A, Fuskevåg OM, Kruspig
B, Simón-Santamaría J, Gogvadze V, Johnsen JI, Kogner P and
Sveinbjörnsson B: Autocrine prostaglandin E2 signaling promotes
tumor cell survival and proliferation in childhood neuroblastoma.
PLoS One. 7:e293312012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Montani M, Hermanns T, Müntener M, Wild P,
Sulser T and Kristiansen G: Multidrug resistance protein 4 (MRP4)
expression in prostate cancer is associated with androgen signaling
and decreases with tumor progression. Virchows Arch. 462:437–443.
2013. View Article : Google Scholar : PubMed/NCBI
|