Introduction
Liver cancer poses a substantial public health
challenge, as 8.3% of patients with cancer worldwide die from liver
cancer (1). Within the realm of
primary liver cancer, primary hepatocellular carcinoma (HCC) is the
predominant histopathological entity (2,3). There
have been advancements in therapeutic modalities for liver cancer,
encompassing interventions such as surgical resection,
chemotherapeutic interventions, radiotherapy and targeted
therapies. Nevertheless, due to the high recurrence rates and
metastatic dissemination of liver cancer, the efficacy of different
types of treatment remains suboptimal (4,5).
Therefore, exploring the underlying molecular therapeutic
mechanisms specific to liver cancer could result in the refinement
of therapeutic approaches against this malignancy.
The function of long non-coding (lnc)RNAs has
steadily emerged as a hot topic of study in tumor biology (6). Numerous previous studies have reported
that lncRNAs exert marked regulatory roles in tumor processes by
acting as ‘sponges’ of micro (mi)RNAs (miR) or competing endogenous
(ce)RNAs (7–10). lncRNAs are engaged in a range of
biological processes within liver cancer cells, including cell
proliferation, apoptosis, migration, metastasis, cell cycle
regulation, tumor stem cell maintenance and modulation of the tumor
microenvironment via multiple signaling pathways (11–14).
The newly discovered lncRNA, progression and
angiogenesis-associated RNA in HCC (PAARH), is associated both with
liver cancer development and angiogenesis, and with the mechanisms
underlying liver cancer progression. A previous study reported that
PAARH is overexpressed in liver cancer and exhibits a targeted
association with miR-6512-3p (15).
However, to the best of our knowledge, the precise mechanistic
interplay between PAARH and miR-6512-3p in liver cancer remains
unknown.
Therefore, the present study used bioinformatic
techniques to analyze the binding sites of lncRNA PAARH and
miR-6512-3p, as well as miR-6512-3p and the LIM and SH3 domain
protein 1 (LASP1) gene. The present study aimed to assess the
functional and regulatory associations between PAARH, miR-6512-3p
and LASP1 that are relevant to liver cancer. The purpose of the
approach in the present study was to elucidate the underlying
mechanism via which the lncRNA PAARH modulates the biological
behavior of liver cancer cells through miR-6512-3p-mediated
targeting of LASP1. The ultimate goal was to provide a theoretical
foundation for the identification of valuable therapeutic targets
in liver cancer.
Materials and methods
Cell culture
The HepG2, HEP3B2.1.7, HCCLM3, Huh-7 and MHCC97-H
liver cancer cell lines and the THLE-2 epithelial cell lines were
used in the present study. All cells were purchased from Aoruisai
Biotechnology (Shanghai) Co. Ltd., and underwent STR
identification. Cell incubations were performed using DMEM (Wuhan
Servicebio Technology Co., Ltd.) supplemented with 100
U.ml−1 penicillin, 10% FBS (Procell Life Science &
Technology Co., Ltd.) and 100 mg.l−1 streptomycin
(Beijing Solarbio Science & Technology Co., Ltd.). The cells
were incubated in a cell culture incubator maintained at 37°C in a
humidified atmosphere containing 5% CO2. Subsequent
experiments were performed after the cell confluence had reached
80–90%.
Bioinformatics analysis
The Encyclopedia of RNA Interactomes (ENCORI)
database (https://starbase.sysu.edu.cn/index.php) was used to
predict the binding sites of miR-6512-3p and LASP1. Additionally,
gene and clinical data from The Cancer Genome Atlas (TCGA;
http://portal.gdc.cancer.gov/) LIHC
dataset were downloaded to confirm the expression of LASP1 in liver
cancer, and to analyze its expression pattern across different
tumor (T), lymph node (N) and metastasis (M) stages of liver
cancer. Using information retrieved from the Gene Expression
Profiling Interactive Analysis (GEPIA; http://gepia.cancer-pku.cn/) and TCGA databases, the
association between LASP1 expression and the survival of patients
with liver cancer was also assessed.
Cell transfection
To evaluate the effects of PAARH interference on
liver cancer cell proliferation, lentiviral vectors for PAARH
[short-hairpin(sh)-PAARH: (5′-CTCCTCAGCAAACCACAGATG-3′) Shanghai
GenePharma Co., Ltd.], were constructed, with a non-targeting
sequence sh-negative control (NC; 5′-TTCTCCGAACGTGTCACGT-3′;
Shanghai GenePharma Co., Ltd.) serving as the negative control. The
packaging plasmids pGag/Pol, pRev and pVSV-G were utilized
(Shanghai GenePharma Co., Ltd.), and the shuttle plasmid was LV3
(Shanghai GenePharma Co., Ltd.). The construction and packaging of
the lentivirus were completed by Shanghai GenePharma Co., Ltd.
HepG2 cells were seeded at a density of 1×105 cells per
well in 6-well cell culture plates. Transfection was performed
using a lentivirus-containing medium (GenePharma, Shanghai, China)
after the cell confluence had reached 50–60%. A multiplicity of
infection (MOI) value of 50 was chosen for the transfection, which
was carried out at room temperature and continued for 24 h.
Subsequently, puromycin (2 µg/ml) was used for cell selection and
maintenance. Transfection efficiency was assessed 96 h
post-transfection using reverse transcription-quantitative
(RT-q)PCR.
Furthermore, to assess the impact of miR-6512-3p on
liver cancer cell proliferation, miR-6512-3p mimic [sense (S),
5′-UUCCAGCCCUUCUAAUGGUAGG-3′ and antisense (AS),
5′-CCUACCAUUAGAAGGGCUGGAA-3′] was used to overexpress miR-6512-3p,
with miR-NC (S, 5′-UCACAACCUCCUAGAAAGAGUAGA-3′ and AS,
5′-UCUACUCUUUCUAGGAGGUUGUGA-3′) serving as the control. An
miR-6512-3p inhibitor (5′-CCUACCAUUAGAAGGGCUGGAA-3′) was used to
suppress miR-6512-3p expression, with inhibitor-NC
(5′-UCUACUCUUUCUAGGAGGUUGUGA-3′) serving as the control. All
aforementioned plasmids were acquired from Hanbio Biotechnology,
Co., Ltd. For these experiments, cells were seeded at a density of
1×105 cells per well in 6-well cell culture plates.
Lipofectamine 3000™ (Invitrogen™; Thermo Fisher Scientific, Inc.)
was used as the cell transfection reagent. Transfection was
performed with a transfection complex composed of 10 µl
Lipofectamine 3000™ and 5 µl of the plasmid, after the cells had
reached a confluency of 40–50%. The transfection was conducted at
room temperature and lasted for 8 h. Transfection efficiency was
evaluated 48 h post-transfection using RT-qPCR.
RT-qPCR analysis
After transfecting cells with different target genes
until the specified time, total RNA from each group of cells was
extracted using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). An RT kit (Monad Biotech Co., Ltd. and
Sangon Biotech Co., Ltd.) was used to synthesize the cDNA according
to the manufacturer's instructions. qPCR was performed using SYBR
Green method (Shanghai Yeasen Biotechnology Co., Ltd.) on a Light
Cycler 96 Real-Time PCR System (Roche Diagnostics). GAPDH was used
as the reference gene for PAARH, U6 was used as the reference gene
for miR-6512-3p, and β-tubulin was used as the reference gene for
LASP1. Analysis of relative gene expression data was performed
using the 2−ΔΔCq method (16). Each experiment set was performed in
triplicate. Primer sequences used were as follows: miR-6512-3p S,
5′-CGCGTTCCAGCCCTTCTAAT-3′ and AS, 5′-AGTGCAGGGTCCGAGGTATT-3′;
PAARH S, 5′-CCTGAAAGTCTCCAAGGCCA-3′ and AS,
5′-TGTGTCTTCAGGCAGCAACT-3′; LASP1 S, 5′-CAAGGGCAAAGGTTTCAGCGTAG-3′
and AS, 5′-ATGCGGCTCTTCTCAAACTCCTC-3′; GAPDH S,
5′-GGACCTGACCTGCCGTCTAG-3′ and AS, 5′-GTAGCCCAGGATGCCCTTGA-3′; U6
S, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ and AS,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; and β-tubulin S,
5′-GGAGAACACGGATGAGACCTACTG-3′ and AS, 5′-CCAGCTTGAGGGTGCGGAAG-3′.
The thermocycling conditions for the RT-qPCR protocol were as
follows: Pre-denaturation at 95°C for 5 min, denaturation at 95°C
for 10 sec, annealing at 60°C for 20 sec, and extension at 72°C for
20 sec. The aforementioned steps constituted one cycle, and a total
of 40 cycles were performed.
Colony formation assay
After the appropriate cell transfections and
treatments had been performed, HepG2 cells in the exponential
growth phase from each group were seeded at a density of
3×103 cells per well in 6-well cell culture plates.
During 7 days of cell culture, the medium was replaced every 2
days. Subsequently, the cells were fixed with 4% paraformaldehyde
(Beijing Solarbio Science & Technology Co., Ltd.) for 30 min at
room temperature, then rinsed twice with PBS (Wuhan Servicebio
Technology Co., Ltd.), followed by staining with crystal violet for
10 min at room temperature. Finally, the cells were imaged (IX71 +
DP80 microscope; Olympus Corporation) and colonies were counted
using ImageJ version 1.54 g (National Institutes of Health), with
colonies defined as having >50 cells each.
Immunofluorescence studies
The 6-well cell culture plates were pre-coated with
cell crawling tablets (Shanghai Hongwo Biotechnology Co., Ltd.),
and HepG2 cells were seeded at a density of 1.5×105
cells per well. When the cell density reached 70–80%, the cells
were fixed with 4% paraformaldehyde at room temperature for 30 min.
Permeabilization was performed using 0.5% Triton-X100 (Beijing
Solarbio Science & Technology Co., Ltd.) for 20 min, with three
5-min PBS washes performed between each step. After the washing
steps were completed, PBS was discarded, and the cells were blocked
for 30 min at room temperature using 10% goat serum (Beijing
Solarbio Science & Technology Co., Ltd.), after which the
blocking solution was discarded. Subsequently, 500 µl of anti-LASP1
primary antibody solution (1:100 dilution; cat. no. ab156872;
Abcam) was added to each well and the plates were incubated
overnight at 4°C (at least 12 h). Fluorescent secondary antibodies
(1:500; Fluor594-conjugated; cat. no. S0006; Affinity Biosciences)
and DAPI (10 µg/ml) were added to the solution and the plates were
incubated for 1 h at room temperature in the dark. Following the
incubation, the coverslips were carefully lifted from the wells and
placed onto glass slides, with the addition of 10 µl of anti-fade
mounting medium. Finally, the samples were imaged using a laser
confocal microscope (Olympus FV3000; Olympus Corporation) and
stored in a refrigerator at 4°C.
Western blotting analysis
A 30-min incubation on ice was performed to lyse the
HepG2 cells using pre-chilled RIPA lysis buffer (Invent
Biotechnologies, Inc.). The lysates were then centrifuged for 20
min at 4°C and 14,000 × g. The protein levels in the supernatants
were quantified using a BCA protein quantification kit (Monad
Biotech Co., Ltd.). Protein concentrations were adjusted before
being mixed with SDS protein loading buffer, followed by
denaturation at 95°C for 5 min and western blot analysis.
After separating the proteins (30 µg protein per
lane) using 12% SDS-PAGE (using the WIX-easyPRO4 Easy Mini Vertical
Electrophoresis Cell System; Interscience Sdn Bhd), an equivalent
amount of denatured proteins was transferred to a PVDF membrane
(MilliporeSigma) using a transfer apparatus (WIX-easyPRO4). After
blocking for 1 h with 5% skimmed milk, the membrane was incubated
overnight at 4°C with primary antibodies, including anti-LASP1
(1:20,000; cat. no. ab156872; Abcam) and anti-β-tubulin (1:50,000;
cat. no. S0001; Affinity Biosciences), diluted in TBST containing
0.05% Tween-20 and 3% BSA. The following day, the samples were
washed three times with TBST for 5 min each. Following the washes,
the membrane was incubated for 1 h at room temperature with
secondary antibodies (1:50,000; 488 Fluorescent Dye; cat. no.
66240-1-Ig; Proteintech Group, Inc.). An enhanced chemiluminescence
kit (Monad Biotech Co., Ltd.) was used to visualize the protein
signals. ImageJ software (version 1.54d; National Institutes of
Health) was used to semi-quantify the band intensities. The loading
control was β-tubulin, and the relative expression of the target
protein was calculated as the ratio of the intensity of the target
protein band to the intensity of the β-tubulin protein band.
Luciferase reporter assay
Luciferases test-human-LASP1-3UTR-wild-type
(LT-h-LASP1-3UTR-WT) and Luciferases test-human-LASP1-3UTR-mutant
(LT-h-LASP1-3UTR-MUT) plasmids were constructed by Hanbio
Biotechnology, Co., Ltd. These plasmids were co-transfected into
293T cells with miR-6512-3p mimic and miR-NC using Lipofectamine
3000™ (Invitrogen; Thermo Fisher Scientific, Inc.) transfection
reagent. At 48 h post-transfection, the luciferase activity in each
cell group was assessed using the Dual-Luciferase Reporter Assay
Kit (Hanbio Biotechnology, Co., Ltd), with Renilla
luciferase activity used as the normalization reference. All the
aforementioned experiments were performed in triplicate.
Statistical analysis
For data analysis, the statistical software GraphPad
Prism version 9 (Dotmatics) and SPSS version 27 (IBM Corp.) were
used. The mean ± standard deviation was used to present
quantitative data. One-way analysis of variance was used to compare
multiple groups of data, followed by the Least Significant
Difference post hoc test for datasets containing three groups and
the Tukey post hoc test for datasets with >3 groups. The
Kruskal-Wallis test was used for non-normally distributed data, and
Dunn's post hoc test was applied to datasets containing more than
three groups. P<0.05 was used to indicate a statistically
significant difference.
Results
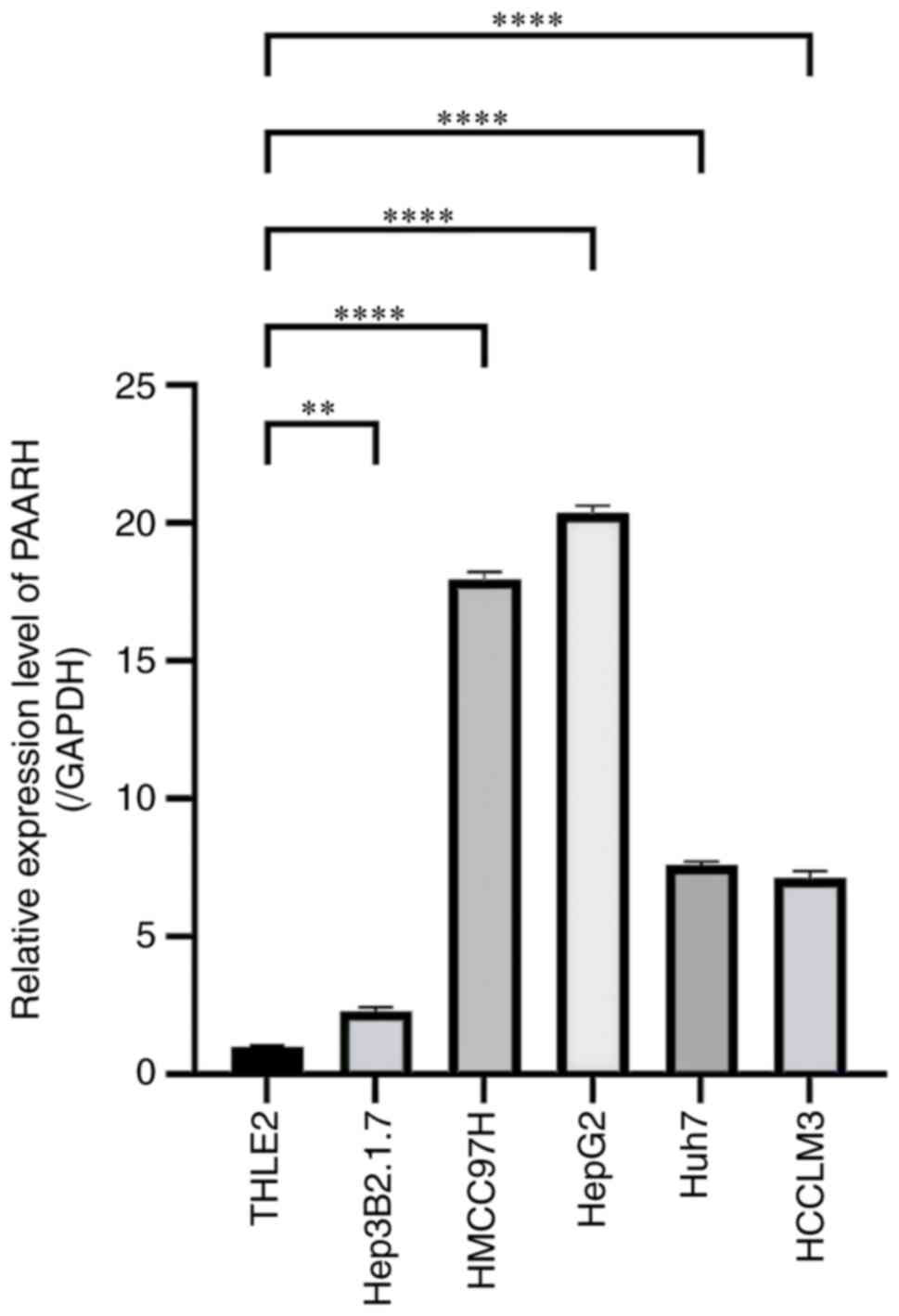
Liver cancer cells exhibit an elevated
level of PAARH expression
RT-qPCR was performed to evaluate the expression
level of PAARH in the HEP3B2.1.7, HepG2, HCCLM3, Huh-7, MHCC97-H
and THLE-2 cell lines to assess the differential expression of
PAARH between the liver cancer cells and the normal liver cell line
(THLE-2). The expression of PAARH was demonstrated to be
significantly upregulated in HepG2, HEP3B2.1.7, HCCLM3, Huh-7 and
MHCC97-H cells compared with the normal liver cell line, THLE-2
(Fig. 1). Notably, the HepG2 cell
line exhibited the highest level of PAARH expression, and therefore
HepG2 cells were chosen for use in future experiments.
PAARH modulates the proliferation of
liver cancer cells via miR-6512-3p
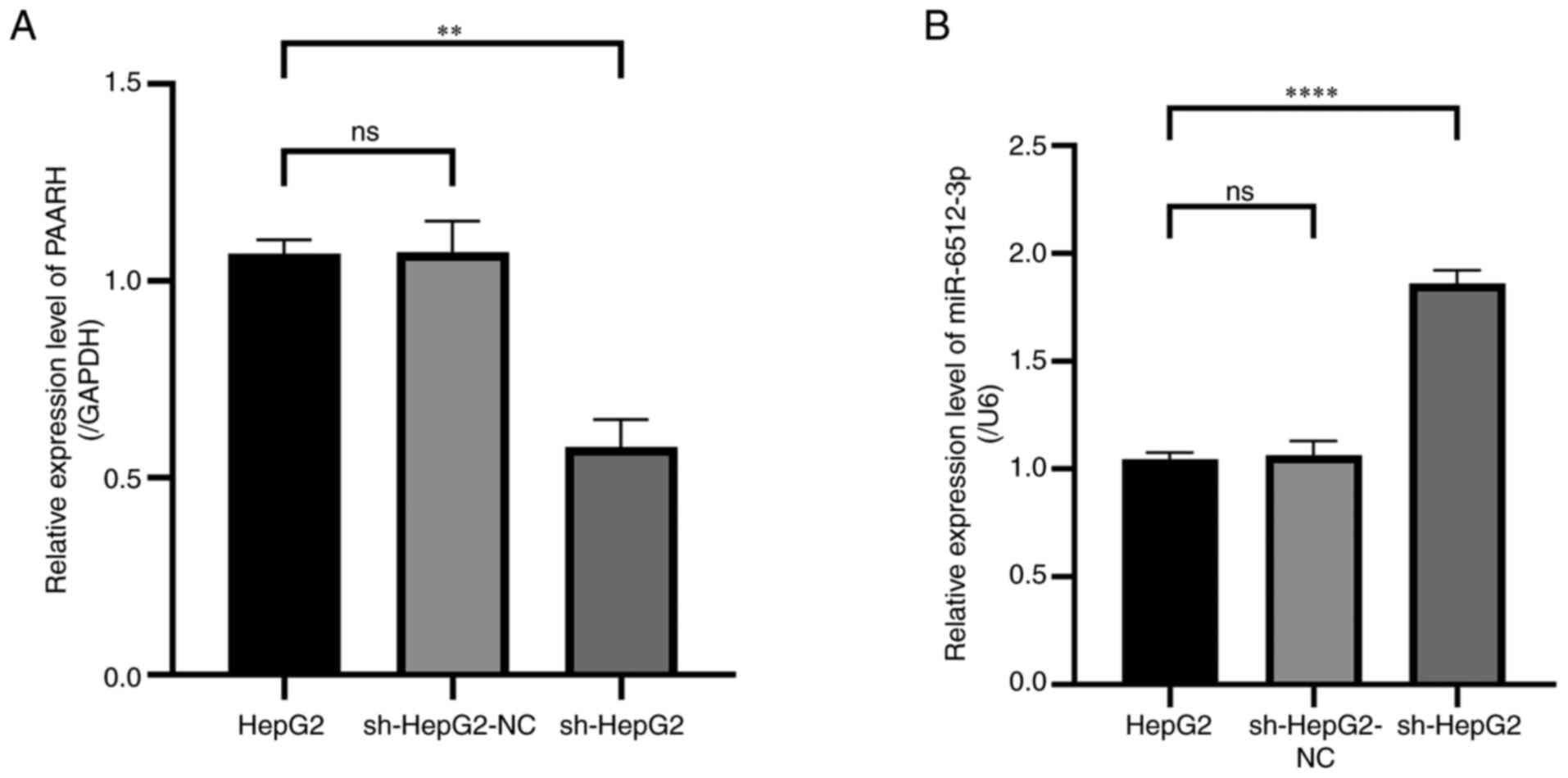
In the initial phases of the present study, an
interaction between PAARH and miR-6512-3p was observed (15). To elucidate the influence of PAARH
on the proliferation of liver cancer cells via miR-6512-3p, a
lentiviral vector was constructed (sh-PAARH) to knock-down PAARH
expression in liver cancer cells. RT-qPCR analysis revealed that,
after sh-PAARH transfection, there was a significant reduction in
PAARH expression in sh-HepG2 cells compared with that in HepG2
cells (Fig. 2A), with a
corresponding significant increase in the level of miR-6512-3p
expression (Fig. 2B), which
indicated that PAARH may regulate miR-6512-3p as a ceRNA.
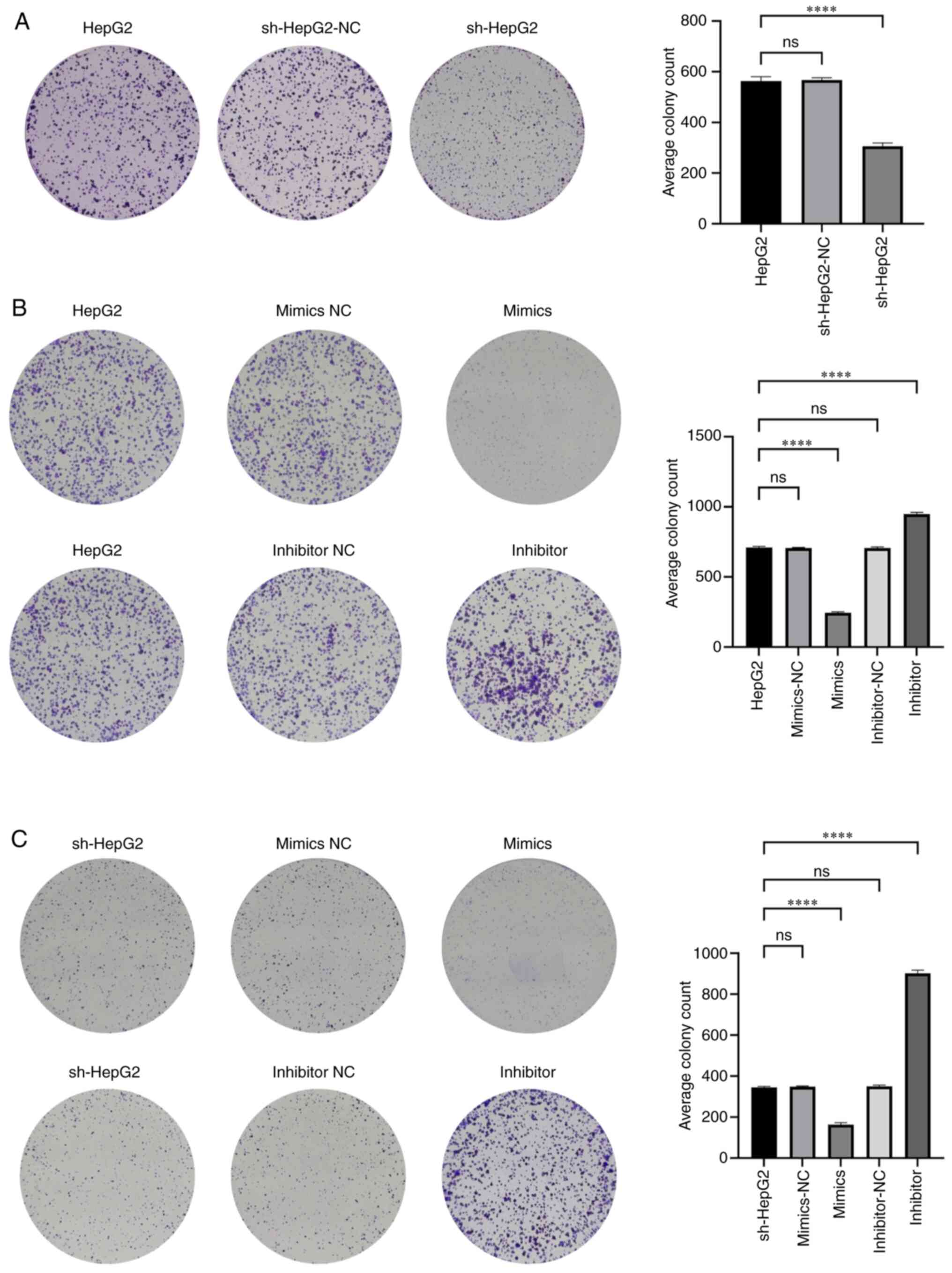
Subsequently, miR-6512-3p mimic and miR-6512-3p
inhibitor plasmids were transfected into HepG2 and sh-HepG2 cells,
respectively (Fig. S1). Colony
formation assays showed that, compared with that in HepG2 cells,
cell proliferation was significantly reduced in sh-HepG2 cells
following PAARH knockdown (Fig.
3A). miR-6512-3p overexpression led to a significant reduction
in cell proliferation in both HepG2 and sh-HepG2 cells, whereas
suppression of miR-6512-3p expression in these cells led to a
significant increase in cell proliferation (Fig. 3B and C). These results revealed that
silencing miR-6512-3p in sh-HepG2 cells partially compensated for
the decreased proliferation observed after PAARH silencing. Taken
together, these experiments demonstrated that PAARH may promote
liver cancer cell proliferation through competitively inhibiting
miR-6512-3p expression.
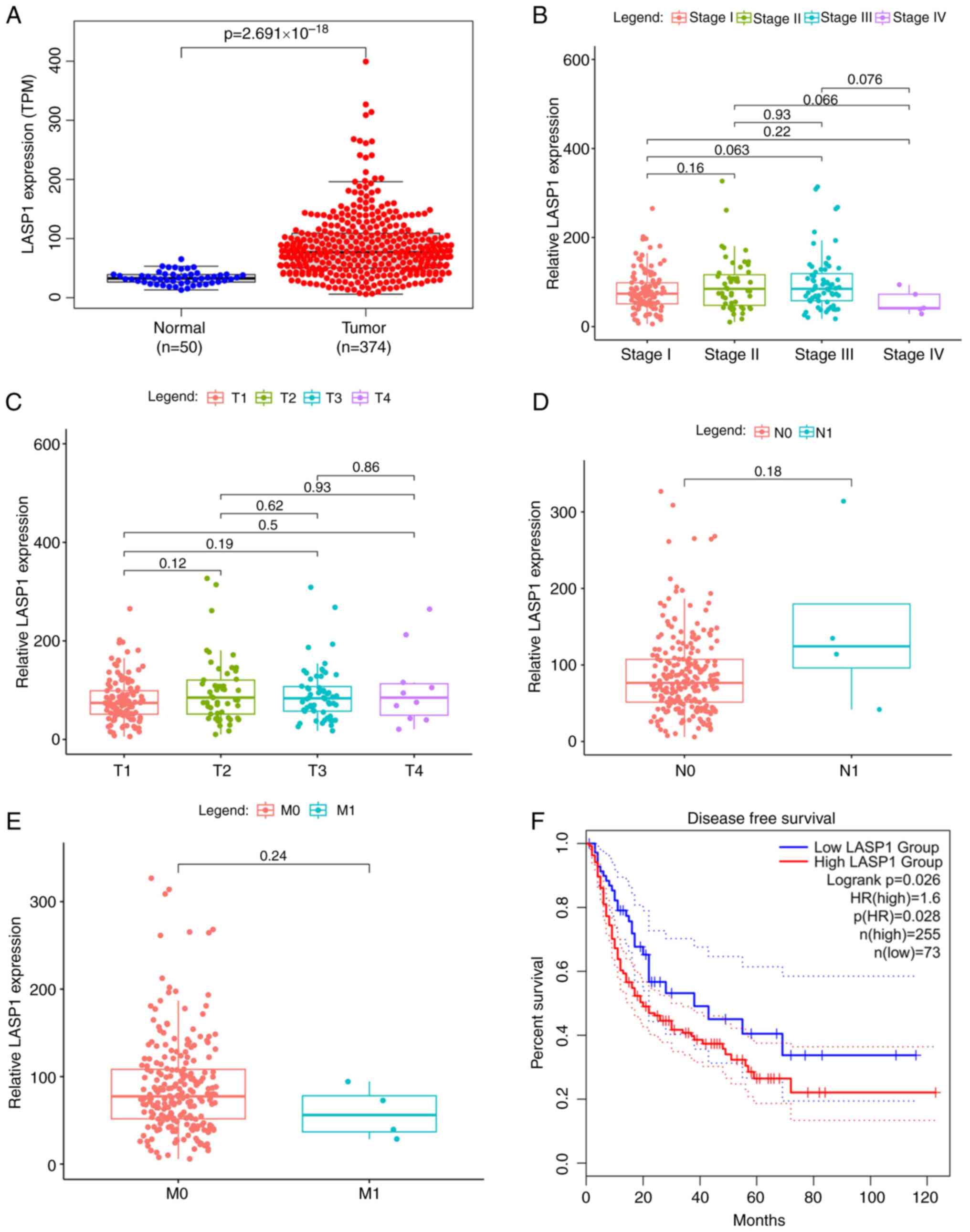
Increased levels of LASP1 expression
suggest a poor prognosis in patients with liver cancer
Using ENCORI database analysis, the binding region
between LASP1 and miR-6512-3p was predicted. TCGA and GEPIA
databases were used to access the gene and clinical data for 424
samples from patients with liver cancer, including 374 liver cancer
tissues and 50 adjacent non-cancerous tissues. The LASP1 expression
level was demonstrated to be substantially and significantly
increased in liver cancer tissues, compared with normal tissues
(Fig. 4A). Across several stages of
liver cancer, the expression level of LASP1 was not demonstrated to
be statistically different in terms of TNM staging, lymph node
involvement or distant metastasis (Fig.
4B-E). However, compared to patients with a low expression
level, those with a high expression level of LASP1 in cancer
exhibited significantly lower disease-free survival rates (Fig. 4F).
Regulation of LASP1 expression via
miR-6512-3p
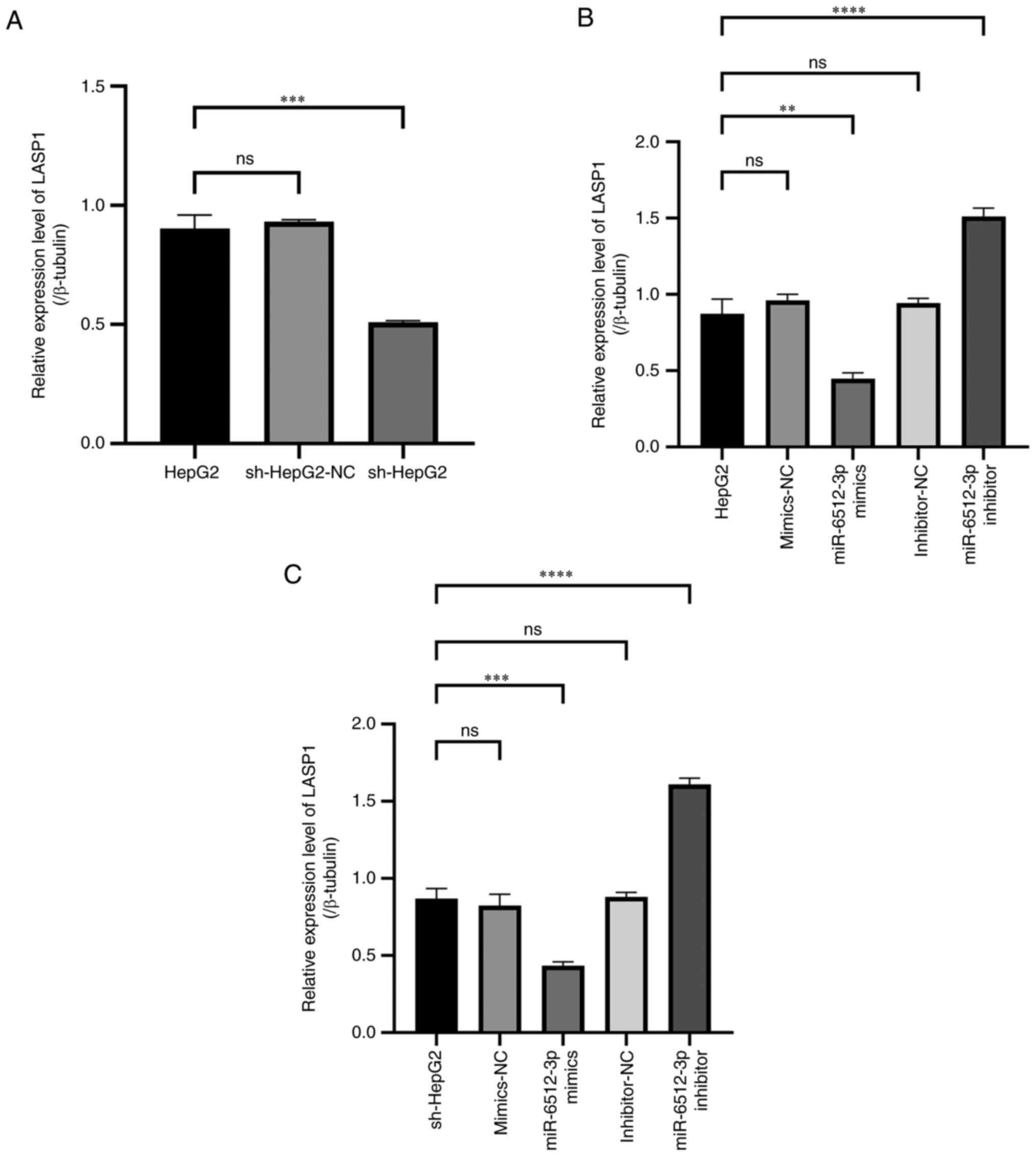
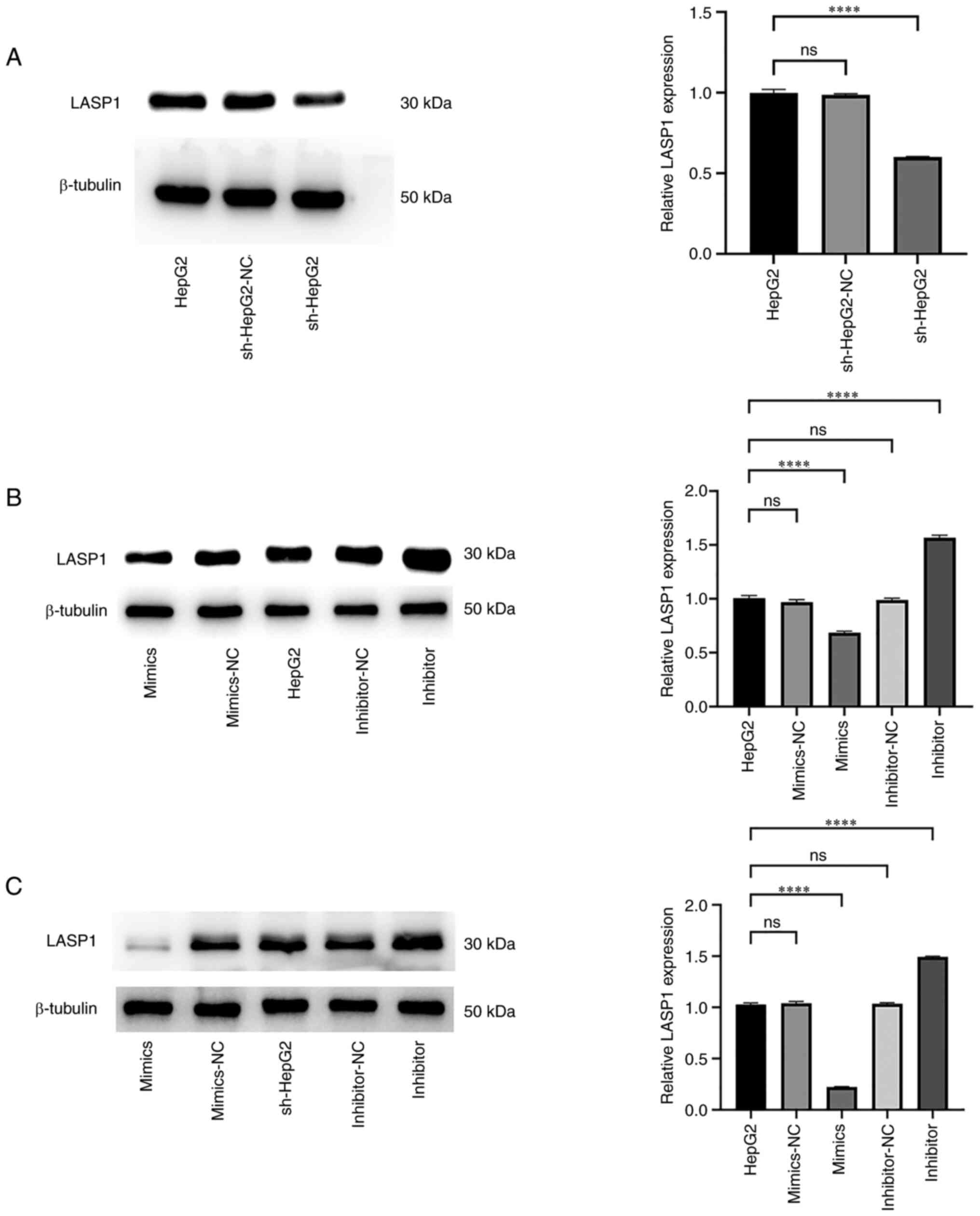
To assess changes in the LASP1 expression level
following miR-6512-3p regulation, LASP1 expression was initially
evaluated in HepG2 cells and HepG2 cells transfected with sh-PAARH.
RT-qPCR analysis revealed that, compared with that in HepG2 cells,
LASP1 expression was significantly reduced in HepG2 cells
transfected with sh-PAARH (Fig.
5A). In HepG2 cells, transfection with a miR-6512-3p mimic led
to a significant decrease in LASP1 expression, whereas transfection
with a miR-6512-3p inhibitor significantly increased LASP1
expression (Fig. 5B). Similarly, in
sh-HepG2 cells, transfection with a miR-6512-3p mimic led to a
significant decrease in LASP1 expression, whereas transfection with
a miR-6512-3p inhibitor significantly reversed the lowered
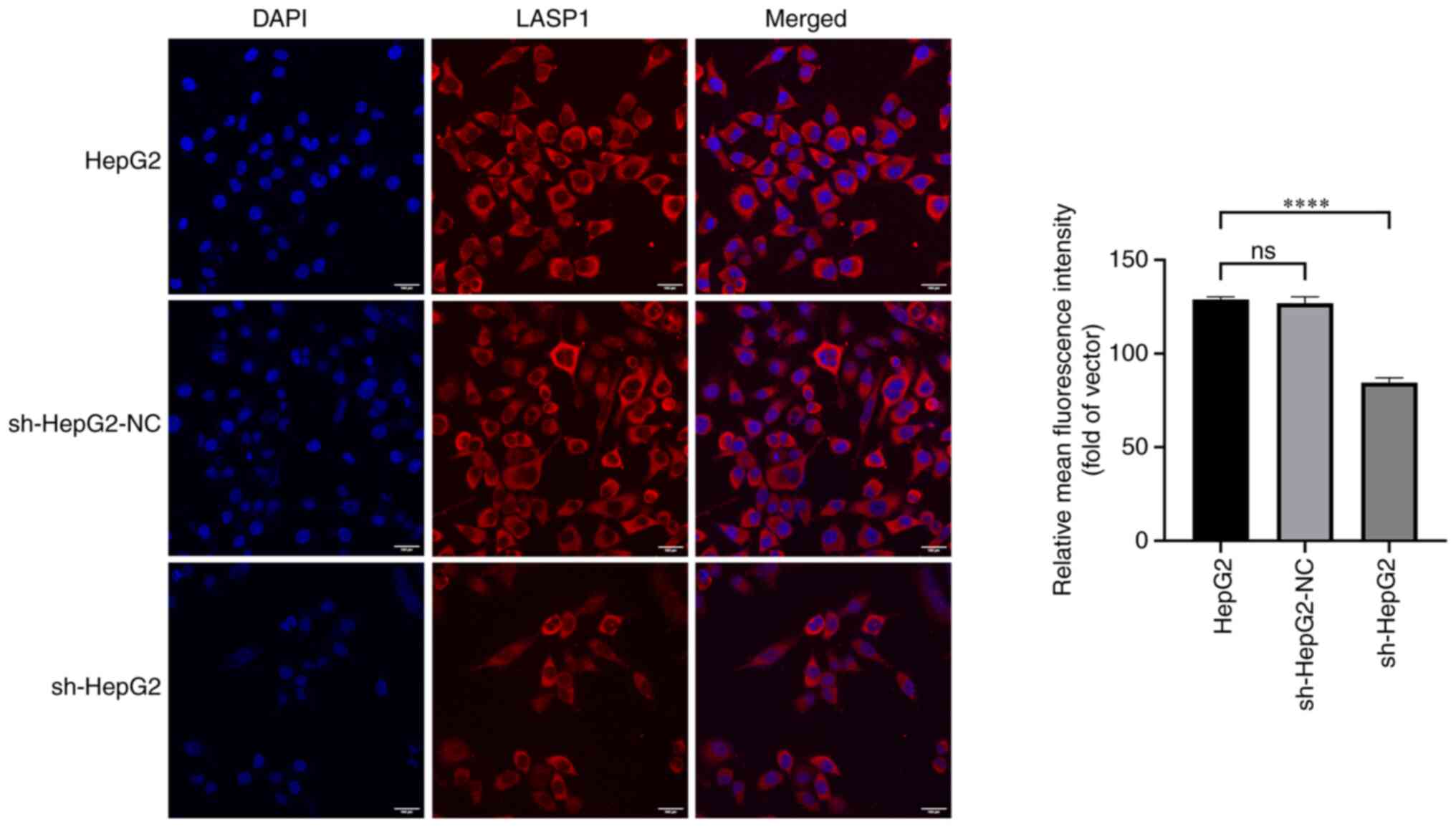
expression level of LASP1 caused by sh-PAARH interference (Fig. 5C). The immunofluorescence (Fig. 6) and western blot (Fig. 7) experiments that were subsequently
performed also confirmed these findings. Overall, the results
collectively indicated that miR-6512-3p could suppress LASP1
expression.
miR-6512-3p directly targets
LASP1
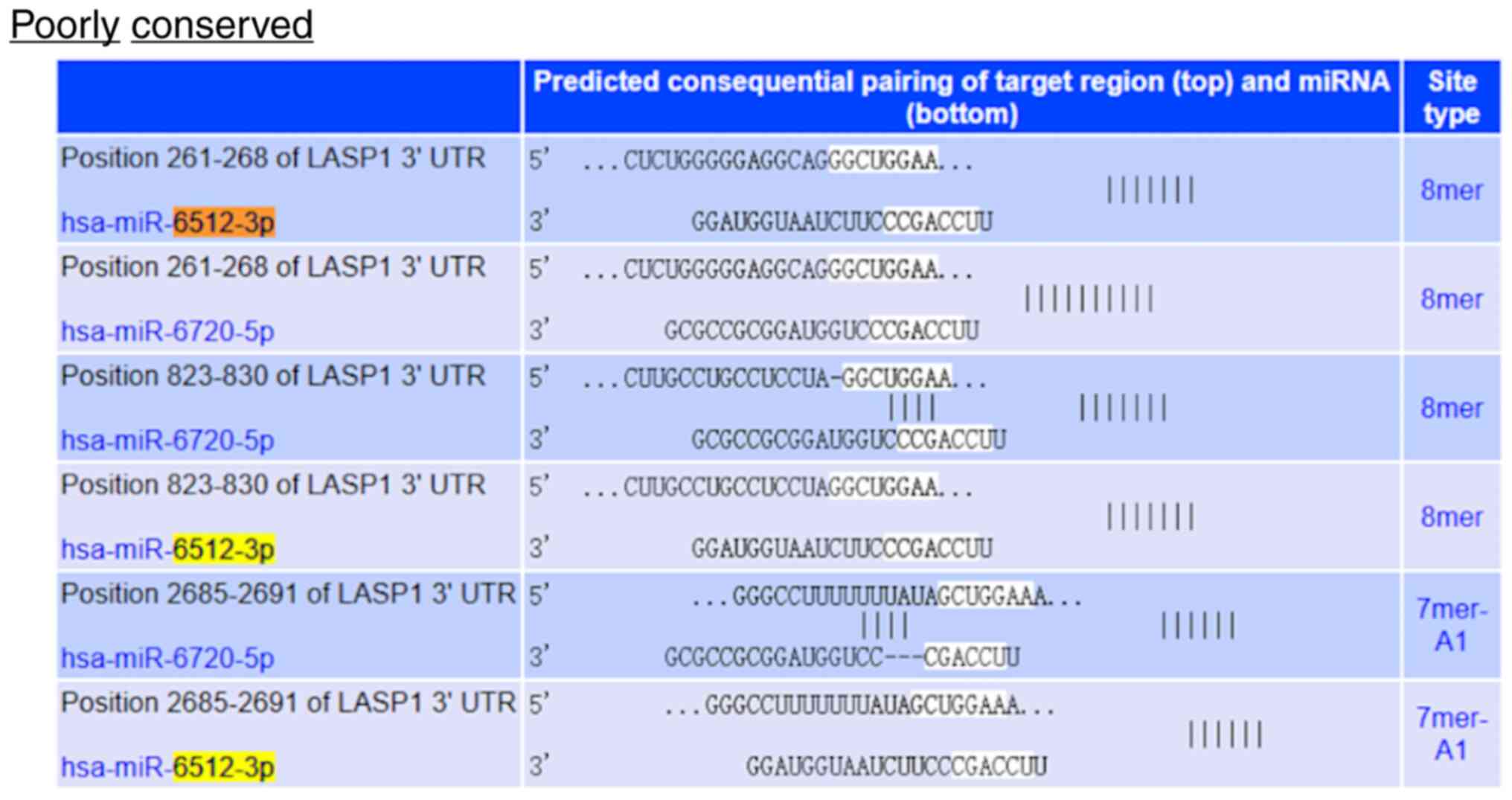
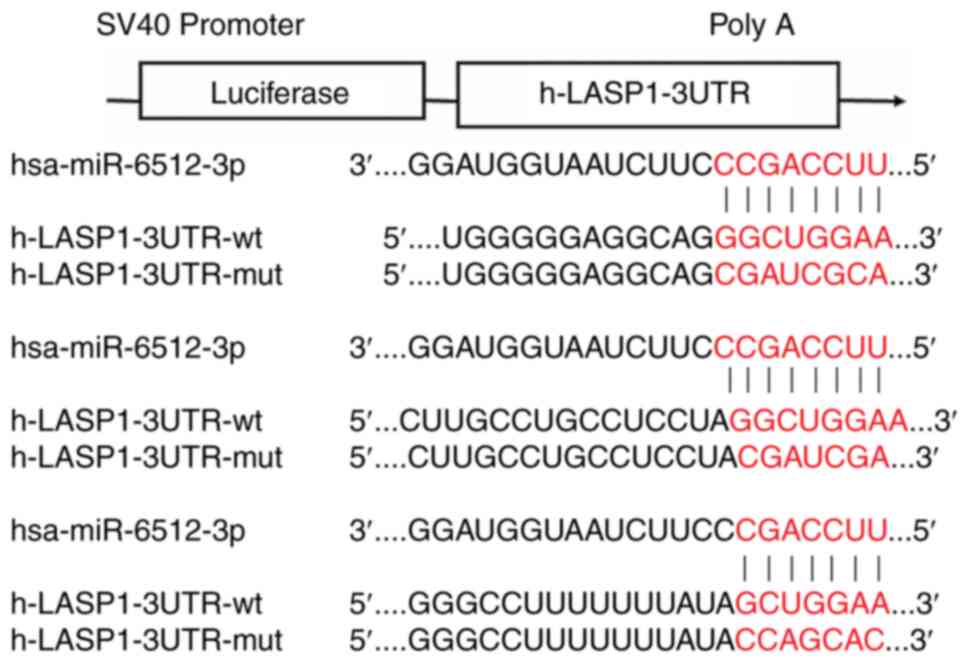
ENCORI database analysis revealed that miR-6512-3p
bound to the 3′-untranslated region (UTR) of LASP1 at nucleotide
positions 261–268, 823–830, 2,685-2,691 and 1604–1610 (Figs. 8 and 9). To assess the binding between
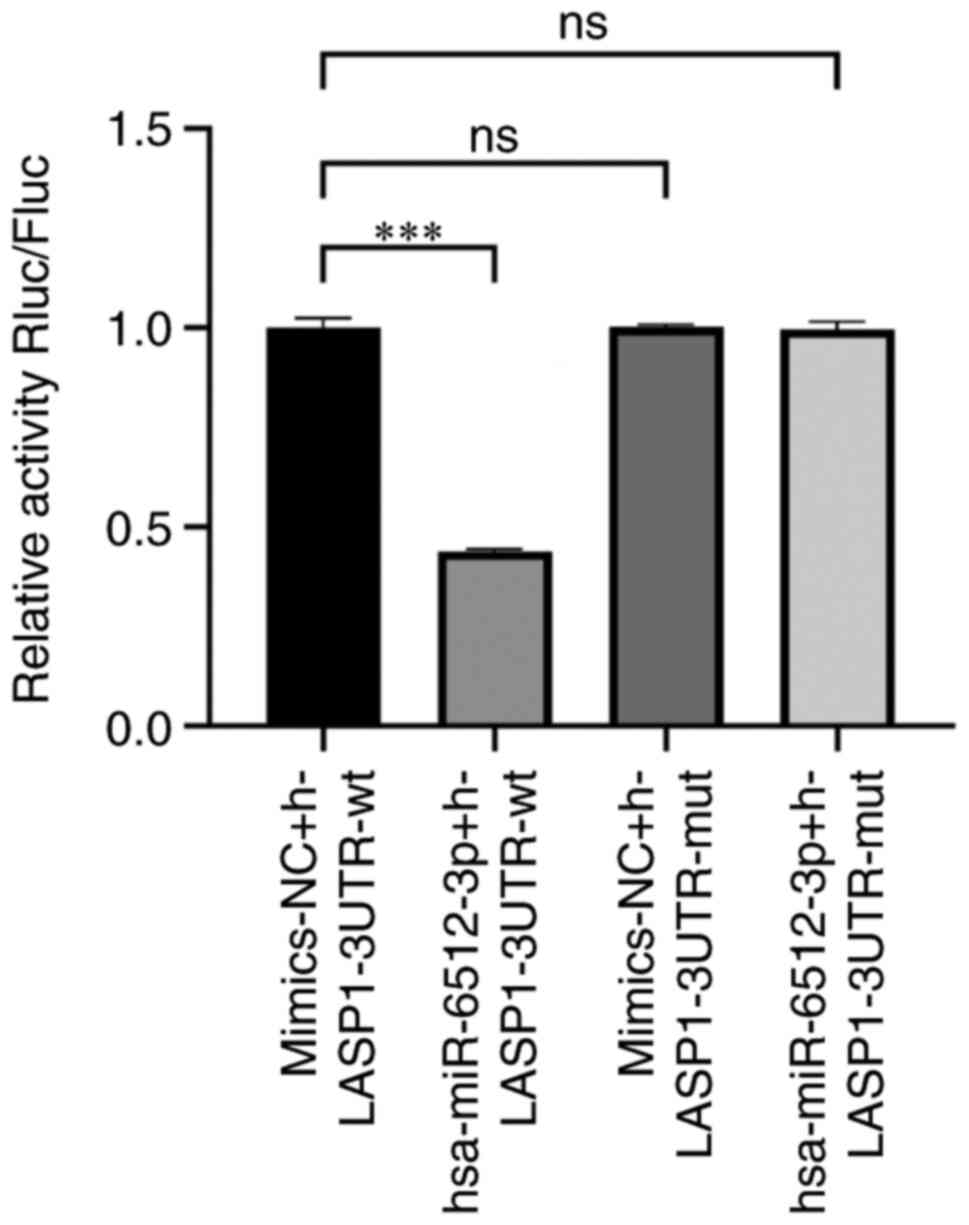
miR-6512-3p and LASP1, a luciferase reporter gene assay was
performed. In the h-LASP1-3UTR-WT-transfected cells,
hsa-miR-6512-3p led to a significant reduction in the level of
luciferase expression compared with the NC group, demonstrating
their interaction during binding. In contrast, after mutation,
hsa-miR-6512-3p was not demonstrated to significantly suppress
luciferase expression in h-LASP1-3UTR-MUT-transfected cells
compared with the NC group, demonstrating that the mutation was
successful (Figs. 10 and S2). Taken together, these experiments
demonstrated that there was a targeted association between
miR-6512-3p and LASP1.
Discussion
Cancer metastasis and recurrence are major
contributing factors in the low survival rates of patients with
liver cancer (17). The role of
lncRNAs in the proliferative processes of several types of human
cancer, including liver cancer, has been confirmed (18,19).
In both our previous and present studies, an increased expression
in the lncRNA PAARH was observed in liver cancer cells and this
increased expression has been reported to facilitate the
proliferation of HCC cells (15).
The mechanistic basis for this effect involves the previously
identified role of lncRNA PAARH in promoting the progression and
angiogenesis of liver cancer through HOXA transcript at the distal
tip upregulation and hypoxia-inducible factor-1α/VEGF signaling
pathway activation (15). In the
present study, it was identified that the mechanistic basis for the
proliferative effect of lncRNA PAARH in liver cancer cells may also
be achieved through competitively inhibition of the expression of
miR-6512-3p. These findings indicated that PAARH served a diverse,
complex and notable regulatory role in the occurrence and
development of liver cancer, representing a potential prognostic
biomarker and therapeutic target for liver cancer. However, several
crucial functions of PAARH have not yet been elucidated, including
its roles in epigenetic regulation, control at the transcriptional
level, modulation of signaling pathways and structural
contributions, among others; therefore, an in-depth exploration of
the role and significance of PAARH in tumor initiation and
progression is still required.
The mechanism of lncRNA action involving ceRNAs
refers to the process whereby lncRNAs share common miRNA response
elements with miRNAs, thereby inhibiting miRNA binding to other
target mRNAs. This subsequently leads to the regulation of the
expression of these mRNAs (20).
Within tumor cells, miRNAs are able to bind to the 3′-UTR of their
target genes, leading to mRNA degradation or translational
repression, thereby suppressing the expression of the target genes.
However, lncRNAs can also competitively bind to miRNA binding
sites, forming lncRNA-miRNA complexes that prevent miRNAs from
binding to other target genes. This alleviates miRNA-mediated
inhibition and upregulates these genes. This action can lead to
changes that may either promote or inhibit cancer (21). Numerous studies have reported that
lncRNAs exert an influence on the functions of HCC cells through
the aforementioned ceRNA mechanism. For example, according to Kong
et al (22), lncRNA
AC006329.1 acts as a ceRNA by inhibiting miR-127-5p and stimulating
the SHC3/ERK signaling pathway. This promotes the progression,
metastasis and epithelial-mesenchymal transition (EMT) of HCC
cells. According to Yao et al (23), phosphatidylinositol transfer protein
α-antisense RNA 1 functions as a ceRNA to sequester miR-363-5p,
thereby regulating the expression of platelet-derived growth
factor-D. This then promotes carcinogenesis. Similarly, Wang et
al (24) reported that lncRNA
02027 served as a ceRNA by competitively binding to miR-625-3p to
regulate the expression of PDZ and LIM domain protein 5, thereby
suppressing HCC cell invasion, proliferation, EMT and
migration.
lncRNA PAARH, also known as CMB9-22P13.1 with a gene
ID of 101927789, is a lncRNA located on the long arm of chromosome
11, q13.1, with a length of 1,116 nucleotides. Studies have
reported that this lncRNA not only enhances the malignant phenotype
of liver cancer cells but also promotes angiogenesis (15). In earlier investigations, it was
demonstrated that lncRNA PAARH functions as a sponge, recruiting
miR-6512-3p, miR-6760-5p and other miRNAs (15). In the present study, to elucidate
the mechanism of action between PAARH and miR-6512-3p in promoting
liver cancer cell proliferation, a lentiviral vector was
constructed for PAARH knockdown to reduce its expression in liver
cancer cells. The results demonstrated a significant reduction in
PAARH expression in HepG2 cells following transfection with
sh-PAARH (Fig. 2A), accompanied by
an elevation in miR-6512-3p expression (Fig. 2B). This suggested a negative
association between PAARH and miR-6512-3p, indicating that PAARH
may regulate miR-6512-3p through a ceRNA mechanism. Subsequently,
HepG2 and sh-HepG2 cells were separately transfected with
miR-6512-3p mimics and miR-6512-3p inhibitors, respectively,
following PAARH silencing. Proliferation was significantly
decreased following PAARH silencing (Fig. 3A). miR-6512-3p overexpression in
HepG2 and sh-HepG2 cells also led to a decrease in proliferation
(Fig. 3B), whilst miR-6512-3p
expression inhibition in these cells resulted in an increase in
proliferation (Fig. 3C). This
indicated that the inhibition of miR-6512-3p expression in sh-HepG2
cells counteracted the decrease in proliferation observed following
PAARH silencing. Therefore, PAARH may promote liver cancer cell
proliferation by competitively inhibiting miR-6512-3p
expression.
In addition, the present study further assessed the
ceRNA mechanism of lncRNAs. To evaluate the ceRNA network mechanism
of PAARH, bioinformatics tools were used to predict miR-6512-3p
target genes. An increased expression level of LASP1 was associated
with a poor prognosis in patients with liver cancer. LASP1 is a
protein extensively expressed within cells, capable of promoting
cellular cytoskeletal remodeling and migration by binding with
cytoskeletal proteins, such as actin. This interaction enhances the
invasive and metastatic capabilities of tumor cells, demonstrating
that LASP1 is crucial for cellular adhesion, proliferation,
migration and transformation (25).
LASP1 also interacts with extracellular matrix receptors such as
integrins, thereby regulating cell adhesion and infiltration
(26). In tumor tissues, the
overexpression of LASP1 may alter cell adhesion properties to the
extracellular matrix, which has the effect of facilitating the
invasion of tumor cells into surrounding tissues and promoting
tumor development (27). Previous
studies have also reported an aberrant expression of LASP1 in
several types of cancer, with its expression closely associated
with tumor prognosis. For example, a study by Ke et al
(28) reported that LASP1,
regulated by the miR-181c-5p/LASP1 axis, experienced changes in
expression levels that affected the proliferation, invasion and
migration of squamous cell carcinoma cells. Furthermore, a study by
Herrmann et al (29)
suggested that there was an association between LASP1 and
chemotherapy resistance in chronic myeloid leukemia. Bioinformatics
analyses in the present study were able to confirm the association
between the upregulation of LASP1 expression in liver cancer and
liver cancer prognosis. Further studies have reported that LASP1 is
involved in cellular migration, invasion, proliferation and
apoptosis through several mechanisms, such as participation in
signaling pathways, m6A methylation modification or influencing the
tumor microenvironment (30–34).
In addition, progress has been made in terms of ongoing therapeutic
research seeking to target LASP1 (35). Therefore, LASP1 is widely regarded
as a prognostic marker for determining tumor prognosis (36–38).
In our previous study, the binding sites between
lncRNA PAARH and miR-6512-3p were identified (16). In the present study, to elucidate
the regulatory role of PAARH via the action of miR-6512-3p on LASP1
expression, the expression levels of LASP1 in HepG2 cells and
PAARH-knockdown HepG2 cells were initially assessed. A significant
decrease in LASP1 expression was observed in the PAARH-knockdown
cells, with synchronized expression patterns between PAARH and
LASP1. Furthermore, miR-6512-3p was demonstrated to suppress LASP1
expression; miR-6512-3p overexpression in liver cancer cells
reduced LASP1 expression levels, whilst the inhibition of
miR-6512-3p enhanced LASP1 expression, indicating a negative
regulatory relationship between miR-6512-3p and LASP1. lncRNA PAARH
deletion increased miR-6512-3p expression in liver cancer cells,
concomitant with a decrease in LASP1 expression. In addition,
interference with miR-6512-3p expression suppressed the low
expression of LASP1 in PAARH-knockdown liver cancer cells,
suggesting that PAARH may regulate LASP1 expression via
miR-6512-3p. Subsequent dual-luciferase reporter gene assays
confirmed their binding relationship, where miR-6512-3p
overexpression led to a reduction in LASP1 expression. These
findings collectively confirmed the existence of the
PAARH-miR-6512-3p-LASP1 ceRNA network.
The present study exhibits some limitations,
including an insufficient exploration of the cellular biological
functions and the absence of animal trials, which constitute
considerable deficiencies. In conclusion, the present study
demonstrated that lncRNA PAARH may regulate the expression of LASP1
by competitively sequestering miR-6512-3p, thereby promoting the
proliferation of liver cancer cells. These findings further suggest
that PAARH may serve as a potential prognostic biomarker and
therapeutic target in liver cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present project received support from the National Natural
Science Foundation of China (grant no. 8226160150), the Guangxi
Science and Technology Project (grant no. GK-AA21196004), and the
Research Foundation Enhancement Project for Middle-aged and Young
Teachers in Higher Education Institutions (grant no.
2020KY13023).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
GL and ZiH conducted the analysis and interpretation
of the experimental data for reverse transcription-quantitative
PCR. JN analyzed and interpreted the experimental data for
immunofluorescence and western blotting, and was responsible for
creating and modifying all figures. YH, ZhH and LH performed
bioinformatics analysis and interpretation. QW led the design and
implementation of the experiments and was the primary contributor
to manuscript writing. JP was the principal investigator, conceived
and designed the experimental content, supervised the entire
experimental process, made substantial revisions to the discussion
section of the manuscript to ensure the accuracy or completeness of
the research work, and undertook the task of drafting responses to
reviewer comments. QW and GL confirm the authenticity of all the
raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cai Y, Lyu T, Li H, Liu C, Xie K, Xu L, Li
W, Liu H, Zhu J, Lyu Y, et al: LncRNA CEBPA-DT promotes liver
cancer metastasis through DDR2/β-catenin activation via interacting
with hnRNPC. J Exp Clin Cancer Res. 41:3352022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ganesan P and Kulik LM: Hepatocellular
carcinoma: New developments. Clin Liver Dis. 27:85–102. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vogel A, Meyer T, Sapisochin G, Salem R
and Saborowski A: Hepatocellular carcinoma. Lancet. 400:1345–1362.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gilles HT, Garbutt T and Landrum J:
Hepatocellular carcinoma. Crit Care Nurs Clin North Am. 34:289–301.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou H, Hao X, Zhang P and He S: Noncoding
RNA mutations in cancer. Wiley Interdiscip Rev RNA. 14:e18122023.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang J, Zhang X, You Z, Meng Y, Fan X,
Qiao G and Pang D: RNA atlas and competing endogenous RNA
regulation in tissue-derived exosomes from luminal B and
triple-negative breast cancer patients. Front Oncol.
13:11131152023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Leng D, Yang Z, Sun H, Song C, Huang C, Ip
KU, Chen G, Deng CX, Zhang XD and Zhao Q: Comprehensive analysis of
tumor microenvironment reveals prognostic ceRNA network related to
immune infiltration in sarcoma. Clin Cancer Res. 29:3986–4001.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fu Z, Yang G, Wang T, Gao D, Lin D and Liu
X: Comprehensive analysis of the Sorafenib-associated druggable
targets on differential gene expression and ceRNA network in
hepatocellular carcinoma. J Environ Pathol Toxicol Oncol. 42:47–60.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Z, Qiu X, He Q, Fu X, Ji F and Tian X:
CCND1-associated ceRNA network reveal the critical pathway of
TPRG1-AS1-hsa-miR-363-3p-MYO1B as a prognostic marker for head and
neck squamous cell carcinoma. Sci Rep. 13:118312023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Y, Hu J, Guo D, Ma W, Zhang X, Zhang Z,
Lu G and He S: LncRNA SNHG5 promotes the proliferation and cancer
stem cell-like properties of HCC by regulating UPF1 and
Wnt-signaling pathway. Cancer Gene Ther. 29:1373–1383. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu Q, Wang H, Lei X, Ma Q, Zhao J, Sun W,
Guo C, Huang D and Xu Q: LncRNA ALKBH3-AS1 enhances ALKBH3 mRNA
stability to promote hepatocellular carcinoma cell proliferation
and invasion. J Cell Mol Med. 26:5292–5302. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang C, Yu H, Zheng Y, Feng W and Shen J:
lncRNA TINCR regulates proliferation and invasion of hepatocellular
carcinoma cells by regulating the miR-375/ATG7 axis. J Oncol.
2022:81324032022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang J, Zhou Y, Gu C, Ming F and Zhang Y:
LncRNA SAMD12-AS1 suppresses proliferation and migration of
hepatocellular carcinoma via p53 signaling pathway. J Oncol.
2022:90963652022.PubMed/NCBI
|
|
15
|
Wei H, Xu Z, Chen L, Wei Q, Huang Z, Liu
G, Li W, Wang J, Tang Q and Pu J: Long non-coding RNA PAARH
promotes hepatocellular carcinoma progression and angiogenesis via
upregulating HOTTIP and activating HIF-1α/VEGF signaling. Cell
Death Dis. 13:1022022. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Brown ZJ, Tsilimigras DI, Ruff SM, Mohseni
A, Kamel IR, Cloyd JM and Pawlik TM: Management of hepatocellular
carcinoma: A review. JAMA Surg. 158:410–420. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Farzaneh M, Ghasemian M, Ghaedrahmati F,
Poodineh J, Najafi S, Masoodi T, Kurniawan D, Uddin S and
Azizidoost S: Functional roles of lncRNA-TUG1 in hepatocellular
carcinoma. Life Sci. 308:1209742022. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pu J, Wang J, Wei H, Lu T, Wu X, Wu Y,
Shao Z, Luo C and Lu Y: lncRNA MAGI2-AS3 prevents the development
of HCC via recruiting KDM1A and promoting H3K4me2 Demethylation of
the RACGAP1 promoter. Mol Ther Nucleic Acids. 18:351–362. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tay Y, Kats L, Salmena L, Weiss D, Tan SM,
Ala U, Karreth F, Poliseno L, Provero P, Di Cunto F, et al:
Coding-independent regulation of the tumor suppressor PTEN by
competing endogenous mRNAs. Cell. 147:344–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kong XX, Yang X, Jiang WJ, Zhu M and Kong
LB: The long non-coding RNA AC006329.1 facilitates hepatocellular
carcinoma progression and metastasis by regulating
miR-127-5p/SHC3/ERK axis. J Hepatocell Carcinoma. 10:1085–1103.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yao J and Liu X: lncRNA PITPNA-AS1
promotes cell proliferation and metastasis in hepatocellular
carcinoma by upregulating PDGFD. Aging (Albany NY). 15:4071–4083.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang J, Zhu Y, Ai X, Wan H, Jia W, Chu J,
Xu B, Kong X and Kong L: Long noncoding RNA 02027 inhibits
proliferation, migration and invasion of hepatocellular carcinoma
via miR-625-3p/PDLIM5 pathway. J Gene Med. 25:e34852023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Butt E, Howard CM and Raman D: LASP1 in
cellular signaling and gene expression: More than just a
cytoskeletal regulator. Cells. 11:38172022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Duvall-Noelle N, Karwandyar A, Richmond A
and Raman D: LASP-1: A nuclear hub for the UHRF1-DNMT1-G9a-snail1
complex. Oncogene. 35:1122–1133. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Niimi A, Limsirichaikul S, Kano K,
Mizutani Y, Takeuchi T, Sawangsri P, Tran DQ, Kawamoto Y and Suzuki
M: LASP1, CERS6, and actin form a ternary complex that promotes
cancer cell migration. Cancers (Basel). 15:27812023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ke S, Fang M, Li R, Wang J and Lu J:
Downregulation of long noncoding RNA breast cancer anti-estrogen
resistance 4 inhibits cell proliferation, invasion, and migration
in esophageal squamous cell carcinoma by regulating the
microRNA-181c-5p/LIM and SH3 protein 1 axis. Bioengineered.
13:12998–13010. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Herrmann AB, Müller ML, Orth MF, Müller
JP, Zernecke A, Hochhaus A, Ernst T, Butt E and Frietsch JJ:
Knockout of LASP1 in CXCR4 expressing CML cells promotes cell
persistence, proliferation and TKI resistance. J Cell Mol Med.
24:2942–2955. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yan P, Liu J, Zhou R, Lin C, Wu K, Yang S,
Yang S, Zhou J, Xu L, Wang H and Zhao L: LASP1 interacts with
N-WASP to activate the Arp2/3 complex and facilitate colorectal
cancer metastasis by increasing tumour budding and worsening the
pattern of invasion. Oncogene. 39:5743–5755. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen N, Han X, Bai X, Yin B and Wang Y:
LASP1 induces colorectal cancer proliferation and invasiveness
through Hippo signaling and Nanog mediated EMT. Am J Transl Res.
12:6490–6500. 2020.PubMed/NCBI
|
|
32
|
Li ZX, Zheng ZQ, Yang PY, Lin L, Zhou GQ,
Lv JW, Zhang LL, Chen F, Li YQ, Wu CF, et al: WTAP-mediated m(6)A
modification of lncRNA DIAPH1-AS1 enhances its stability to
facilitate nasopharyngeal carcinoma growth and metastasis. Cell
Death Differ. 29:1137–1151. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou L, Zhang L, Guan X, Dong YI and Liu
T: Long noncoding RNA PPP1R14B-AS1 imitates microRNA-134-3p to
facilitate breast cancer progression by upregulating LIM and SH3
protein 1. Oncol Res. 29:251–262. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen X, Zhao S, Li Q, Xu C, Yu Y and Ge H:
LncRNA NEAT1 knockdown inhibits retinoblastoma progression by
miR-3619-5p/LASP1 axis. Front Genet. 11:5741452020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dejima T, Imada K, Takeuchi A, Shiota M,
Leong J, Tombe T, Tam K, Fazli L, Naito S, Gleave ME and Ong CJ:
Suppression of LIM and SH3 domain protein 1 (LASP1) negatively
regulated by androgen receptor delays castration resistant prostate
cancer progression. Prostate. 77:309–320. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou R, Shao Z, Liu J, Zhan W, Gao Q, Pan
Z, Wu L, Xu L, Ding Y and Zhao L: COPS5 and LASP1 synergistically
interact to downregulate 14-3-3σ expression and promote colorectal
cancer progression via activating PI3K/AKT pathway. Int J Cancer.
142:1853–1864. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Butt E, Ebbing J, Bubendorf L and Ardelt
P: Influence of hematuria and infection on diagnostic accuracy of
urinary LASP1: A new biomarker for bladder carcinoma. Biomark Med.
11:347–357. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen Q, Wu K, Qin X, Yu Y, Wang X and Wei
K: LASP1 promotes proliferation, metastasis, invasion in head and
neck squamous cell carcinoma and through direct interaction with
HSPA1A. J Cell Mol Med. 24:1626–1639. 2020. View Article : Google Scholar : PubMed/NCBI
|