Introduction
Colorectal cancer (CRC) is the third most common
cancer worldwide (1), with patients
frequently experiencing poor prognoses primarily due to acquired
drug resistance and mechanisms underlying immune escape (2,3). The
programmed cell death protein 1 (PD-1)/programmed cell death
ligand-1 (PD-L1) signaling axis is a vital pathway that facilitates
tumor immune escape (4), where
tumor cells expressing PD-L1 interact with PD-1 on T cells. The
identification of small molecule drugs that improve the efficacy of
PD-1/PD-L1 immunotherapy is critical for enhancing CRC treatment
outcomes.
There has been growing interest in investigating the
impact of intestinal microbiota and its metabolites on host
immunity (5,6). Among these metabolites, short-chain
fatty acids (SCFAs), such as acetate, propionate and butyrate, are
produced through the microbial fermentation of dietary
carbohydrates in the gut (7). SCFAs
are central to maintaining normal gastrointestinal functions, such
as immune modulation and host metabolism (8,9). SCFAs
exhibit protective effects by modulating the inflammatory cascade,
including the inhibition of the nuclear factor-κB and histone
deacetylase pathways (10,11). Acetate is the most abundant SCFA and
accounts for ~50% of all SCFAs in the colon (12). Researchers have elucidated its
potential role in cancer immunotherapy. For example, Tran et
al (13) reported that acetate
reduced the expression of poliovirus receptor/cluster of
differentiation (CD)155 in colorectal cancer cells, thus
upregulating CD8+ T cell activity against cancer. Peng
et al (14) demonstrated
that gut microbiota that produced SCFAs were positively associated
with an anti-PD-1/PD-L1 response in gastrointestinal malignancies.
These findings highlight the role of SCFAs in resisting tumor
immune escape. Certain reports have described the role and
molecular mechanisms underlying sodium propionate (SP) in cancer
progression. For instance, Park et al (15) reported that SP suppressed breast
cancer cell proliferation and induced cell apoptosis in
tumor-bearing mice in vivo via the Janus kinase 2/signal
transducer and activator of transcription 3/reactive oxygen
species/p38 mitogen-activated protein kinase pathway. Kim et
al (16) demonstrated that SP
promoted apoptosis and induced cell cycle arrest in lung cancer.
However, the role and mechanisms underlying SP in CRC remain
unclear.
Understanding the interplay between drug resistance
and immune escape mechanisms is vital, particularly since prolonged
or high-dose drug treatments may promote drug resistance through
pathways linked to immune evasion (17,18).
Tsai et al (19) reported
that cisplatin upregulated PD-L1 expression in bladder cancer by
targeting the extracellular signal-regulated protein kinase 1/2 and
activator protein-1 pathways, indicating that the immune escape
mechanism may induce chemoresistance. Additionally, cisplatin has
been shown to enhance PD-L1 expression in non-small cell lung
cancer, and its combined use with PD-1/PD-L1 pathway inhibitors
increased treatment efficacy in tumor-bearing models in vivo
(20).
The present study aimed to explore the effects of
high-dose SP on PD-L1 expression in CRC cells and to investigate
the underlying mechanisms to identify novel therapeutic strategies
for overcoming immune escape in CRC.
Materials and methods
Cell culture and transfection
The HCT116 and SW480 cell lines were acquired from
Procell Life Science & Technology Co., Ltd. and cultured in
Dulbecco's Modified Eagle Medium (Thermo Fisher Scientific, Inc.).
The medium was supplemented with 10% fetal bovine serum (Thermo
Fisher Scientific, Inc.) and 1% penicillin/streptomycin solution
(Beijing Solarbio Science & Technology Co., Ltd.). The cells
were maintained in a humidified incubator at 37°C with 5%
CO2. For the transfections, the short hairpin (sh)RNA
and negative control (NC) scrambled shRNA sequences were inserted
into the pLKO.1 plasmid (MilliporeSigma), resulting in the
formation of pLKO.1-shRNA and pLKO.1-sh-NC constructs. The
targeting portions of the sequence in sh-IGF2BP3#1, sh-IGF2BP3#2,
sh-IGF2BP3#3 and sh-NC were as follows: sh-IGF2BP3#1,
5′-GGTGCTGGATAGTTTACTA-3′; sh-IGF2BP3#2, 5′-GGTAAAGCAGCACCAACAA-3′;
sh-IGF2BP3#3, 5′-CGATGTCCACCGTAAAGAA-3′; and sh-NC:
5′-CAACAAGATGAAGAGCACCAA-3′. For the transfection, 2 µg of
pLKO.1-shRNA or pLKO.1-sh-NC plasmid DNA was diluted in 250 µl
Opti-MEM medium (Thermo Fisher Scientific, Inc.), mixed with 5 µl
Lipofectamine® 2000 reagent (pre-diluted in 250 µl
Opti-MEM) and incubated at room temperature for 20 min to form
transfection complexes. The complexes were added to cells and
incubated at 37°C with 5% CO2 for 6 h, after which the
medium was replaced with fresh complete medium. Cells were
harvested 48 h post-transfection for subsequent functional or
molecular analyses.
Cell Counting Kit-8 (CCK-8) assay
A commercial CCK-8 (Beijing Solarbio Science &
Technology Co., Ltd.) was used for this assay. First, the
half-maximal inhibitory concentration (IC50) of SP was
determined using the CCK-8 assay. For this, HCT116 cells were
seeded into 96-well plates and treated with SP at gradient
concentrations of 0.16, 0.8, 4, 20, 100 and 500 mM for 48 h at
37°C. After treatment, 10 µl CCK-8 reagent was added to each well
and incubated with the HCT116 cells for 2 h. Finally, the optical
density (OD) at 450 nm was recorded. Second, to analyze cell
viability, HCT116 and SW480 cells were seeded into 96-well plates
and treated with 10 mM SP for 48 h. Consequently, 10 µl CCK-8
reagent was added to the cells and incubated for 2 h. The OD at 450
nm was again detected.
Immunofluorescence (IF) assay
The abundance of proliferating cell nuclear antigen
(PCNA) in CRC cells was analyzed by an IF assay. After SP
treatment, the cells were fixed with 4% paraformaldehyde at room
temperature (25°C) for 20 min, permeabilized with 0.1% Triton X-100
in Tris-buffered saline and blocked with 3% donkey serum
(MilliporeSigma) at 25°C for 1 h. Subsequently, the cells were
incubated with an anti-PCNA antibody (cat no. GB11010; 1:500; Wuhan
Servicebio Technology Co., Ltd.) diluted in blocking buffer at 4°C
overnight, followed by incubation with Alexa Fluor 488-conjugated
goat anti-mouse IgG secondary antibody (cat no. GB25301; 1:800;
Wuhan Servicebio Technology Co., Ltd.) at 25°C for 1 h in the dark.
The nuclei were then stained using 4′,6-diamidino-2-phenylindole
(DAPI; MilliporeSigma; 1 µg/ml) at 25°C for 10 min. Fluorescence
images were captured under a Zeiss LSM 900 confocal microscope
(Carl Zeiss AG), with a scale bar of 100 µm. Fluorescence intensity
quantification was performed with ImageJ software (version 1.53;
National Institutes of Health).
Flow cytometry
Cell apoptosis was analyzed using flow cytometry.
First, CRC cells were seeded into 6-well plates and treated with 10
mM SP for 48 h. Second, the cells were harvested, washed three
times with ice-cold PBS and resuspended in ice-cold
phosphate-buffered saline. Third, the cells were stained with 50
µg/ml annexin V-fluorescein isothiocyanate (BD Biosciences) and 10
µg/ml propidium iodide (BD Biosciences) in the dark for 15 min.
Finally, the cells were analyzed using a FACSCalibur flow cytometer
(BD Biosciences), and the apoptosis rate was analyzed using FlowJo
software (version 10.8.1; FlowJo LLC; BD Biosciences).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA samples were extracted from the CRC cells
using the EastepTM Universal RNA Extraction Kit (Promega
Corporation), and cDNA was synthesized using the GoScriptTM Reverse
Transcription System (Promega Corporation). The kit was used
according to the manufacturer's instructions. qPCRs were conducted
using the GoTaq® qPCR Master Mix (Promega Corporation)
on the ABI 7500 Fast Real-Time PCR System (Applied Biosystems;
Thermo Fisher Scientific, Inc.) under the following thermocycling
conditions: Initial denaturation at 95°C for 2 min; 40 cycles of
denaturation at 95°C for 15 sec and annealing/extension at 60°C for
30 sec. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH), the
housekeeping gene, served as the internal control. Relative gene
expression was analyzed by the 2−ΔΔCq method (21). The primer sequences for PD-L1,
IGF2BP3 and GAPDH were as follows: PD-L1 forward primer (F),
5′-GGTAGAGTATGGTAGCAATATG-3′; PD-L1 reverse primer (R),
5′-CCTTCAGGTCTTCCTCTCCA-3′; IGF2BP3 F, 5′-TTCAAGGACGCCAAGATCCC-3′;
IGF2BP3 R, 5′-TCCCACTGTAAATGAGGCGG-3′; GAPDH F,
5′-GTCAAGGCTGAGAACGGGAA-3′; and GAPDH R,
5′-AAATGAGCCCCAGCCTTCTC-3′.
Western blot assay
The CRC cells were harvested, washed with PBS and
lysed in ice-cold radioimmunoprecipitation assay buffer (Beyotime
Institute of Biotechnology) for 30 min on ice. Then, protein
concentrations were measured using the bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.). The protein
samples (35 µg/lane) were then separated by 12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to
polyvinylidene difluoride membranes. The membranes were blocked
with 5% skimmed milk at room temperature for 1 h and then incubated
overnight with the following primary antibodies at 4°C overnight:
anti-PD-L1 (cat. no. GB11339A; 1:5,000; Wuhan Servicebio Technology
Co., Ltd.), anti-IGF2BP3 (cat. no. 14642-1-AP; 1:5,000; Proteintech
Group, Inc.) and anti-GAPDH (cat. no. GB15002; 1:20,000; Wuhan
Servicebio Technology Co., Ltd.). The next day, the membranes were
incubated with a horseradish peroxidase-conjugated secondary
antibody (cat. no. GB23303/GB23301; 1:20,000; Wuhan Servicebio
Technology Co., Ltd.) at 25°C for 1 h. Finally, the immunoreactive
protein bands were visualized using an enhanced chemiluminescence
kit (Beyotime Institute of Biotechnology). Densitometry was
conducted using ImageJ software (version 1.53; National Institutes
of Health), with GAPDH as the internal reference.
Bioinformatics analysis
The m6A2Target database (http://m6a2target.canceromics.org) was used to predict
N6-methyladenosine (m6A) modification-associated proteins upstream
of PD-L1. Genes co-expressed with PD-L1 were analyzed using the R
package ggplot2 (version 3.3.6; http://cran.r-project.org/package=ggplot2). IGF2BP3
expression data in colon adenocarcinoma (COAD) and rectum
adenocarcinoma (READ) tissues, along with their corresponding
healthy tissues, were downloaded from The Cancer Genome Atlas
(TCGA, http://portal.gdc.cancer.gov/) and
compared using the Wilcoxon signed-rank test using R statistical
software (version 4.2.1; http://www.r-project.org/). PD-L1 expression data were
downloaded from COAD and READ datasets in TCGA. Kaplan-Meier
survival curves was first constructed to visualize the association
between survival outcomes and pathological grades. The log-rank
test was then performed using R statistical software (version
4.2.1; http://www.r-project.org/).
Actinomycin D (ActD) assay
The stability of PD-L1 mRNA was analyzed using the
ActD assay. After transfection with sh-NC or sh-IGF2BP3#2 for 48 h,
2 µg/ml of the transcription inhibitor, ActD (MilliporeSigma), was
added to the wells. After ActD exposure for 0, 1, 2 or 4 h at 37°C,
the cells were harvested and PD-L1 expression was measured by
RT-qPCR.
Methylated RNA immunoprecipitation
(meRIP)-qPCR assay
The meRIP-qPCR assay was conducted as described
previously (22). To inhibit
m6A methylation, HCT116 cells were treated with 100 µM
S-adenosylhomocysteine (SAH) dissolved in serum-free DMEM for 48 h
at 37°C. Control cells were incubated with an equal volume of
solvent (0.1% DMSO in serum-free DMEM) under identical conditions.
Total RNA was extracted from HCT116 cells using TRIzol®
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and treated
with DNase I (Thermo Fisher Scientific Inc.) based on the
manufacturer's instructions. RNA (10 µg) was fragmented in 10 mM
ZnCl2 at 94°C for 5 min, followed by incubation with
mouse anti-m6A monoclonal antibodies diluted at a 1:300 ratio,
labeled Protein G Dynabeads (cat. no. 10003D; Thermo Fisher
Scientific, Inc.) at a ratio of 5 µg antibody per 1 mg Dynabeads at
4°C for 2 h. A 50 µl volume of conjugated Dynabeads slurry was
added to each RNA sample. After incubation, beads were washed three
times with high-salt buffer (50 mM Tris-HCl pH 7.4, 1 M NaCl, 1%
NP-40) and once with low-salt buffer (50 mM Tris-HCl pH 7.4, 0.1 M
NaCl). Beads were separated using a magnetic stand and bound RNA
was eluted in Elution Buffer (10 mM Tris-HCl pH 7.0, 1 mM EDTA,
0.5% SDS) at 65°C for 10 min. qPCR was performed using SYBR Green
Master Mix (Takara Bio, Inc.) on the ABI 7500 Fast Real-Time PCR
System (Applied Biosystems; Thermo Fisher Scientific, Inc.),
following the aforementioned protocol. The normalization was
performed using the ‘Input’ fraction. The PD-L1 primers used in the
RIP-qPCR assay were as follows: PD-L1 F,
5′-CCCATACAACAAAATCAACCAAAG-3′; and PD-L1 R:
5′-CTTGGAATTGGTGGTGGTGGTC-3′.
RIP-qPCR assay
The interaction between IGF2BP3 protein and PD-L1
mRNA was confirmed using a RIP assay as described previously
(23). Cells were lysed in RIPA
buffer (Thermo Fisher Scientific, Inc.). For each IP reaction, 500
µg of lysate was incubated with 5 µg of anti-IGF2BP3 antibodies,
diluted at a 1:200 ratio (cat. no. 14642-1-AP; Proteintech Group,
Inc.) and 50 µl Protein G magnetic beads (Thermo Fisher Scientific,
Inc.) at 4°C overnight. Beads were washed three times with
high-salt wash buffer (500 mM NaCl, 0.1% NP-40, 50 mM Tris-HCl, pH
7.4) and once with low-salt buffer (150 mM NaCl, 0.1% NP-40, 50 mM
Tris-HCl, pH 7.4), with each wash step involving centrifugation at
3,000 × g for 5 min at 4°C. RNA was isolated from the
immunoprecipitated complexes using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and quantified by qPCR
as aforementioned.
Statistical analysis
All experiments were conducted in triplicate. Data
were analyzed using GraphPad Prism 8.0 software (Dotmatics), with
the results expressed as the mean ± SD. The unpaired student's
t-test was conducted to compare the differences between two groups.
One-way analysis of variance followed by Tukey's test was used to
compare among multiple groups. P<0.05 was considered to indicate
a statistically significant difference.
Results
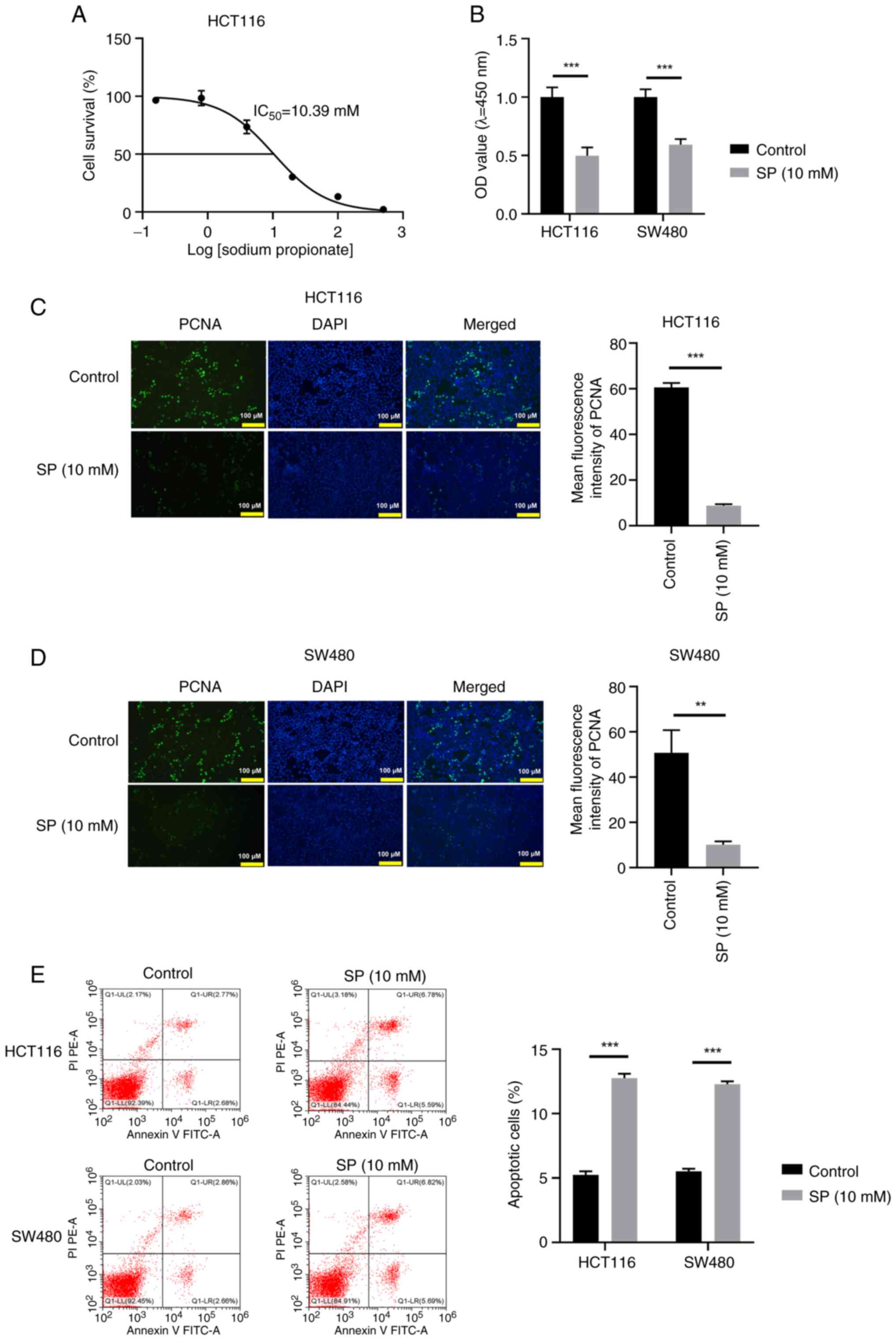
Low-dose SP restrains CRC cell
proliferation and triggers cell apoptosis in vitro
The effects of SP on the biological behavior of CRC
cells was assessed through a series of functional experiments. The
IC50 value of SP in the HCT116 cell line was 10.39 mM
(Fig. 1A). Subsequently, 10 mM SP
was selected for further assays. SP treatment markedly reduced CRC
cell viability (Fig. 1B). PCNA, a
marker of cell proliferation, was identified using the IF assay,
which indicated a notable reduction in PCNA abundance after SP
treatment (Fig. 1C and D).
Additionally, SP treatment triggered apoptosis in CRC cells
(Fig. 1E). Taken together, 10 mM SP
exerted an antitumor effect on CRC cells.
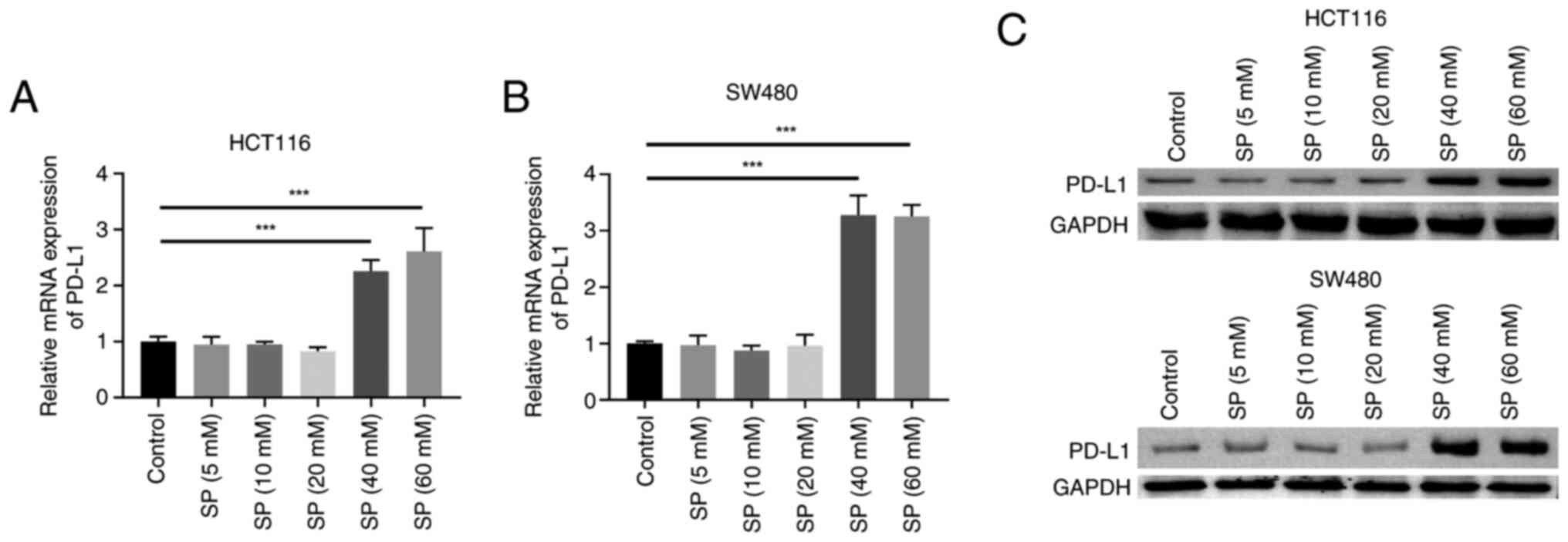
High-dose SP induces PD-L1 expression
in CRC cells
Acquired drug resistance in tumor cells has been
strongly associated with immune escape (24). PD-L1 is a pivotal mediator of T cell
activity, and the PD-L1/PD-1 pathway mediates tumor immune escape
(25). In the present study, the
effects of different doses of SP on PD-L1 expression in CRC cells
were investigated. SP (5, 10 and 20 mM) exerted marginal effects on
the PD-L1 mRNA and protein expression levels in the HCT116 and
SW480 cell lines. By contrast, SP (40 and 60 mM) significantly
induced PD-L1 mRNA and protein expression compared with the control
group (Fig. 2). Additionally, 40
and 60 mM SP exhibited comparable effects on promoting PD-L1 mRNA
and protein expression. Although 5, 10 and 20 mM SP could inhibit
cell viability, only 40 and 60 mM SP could promote PD-L1
expression. Therefore, relative to the 5, 10 and 20 mM doses, the
SP dose that could enhance PD-L1 expression (40 mM) was defined as
the ‘high’ dose. Considering that for most ‘cold’ tumors, tumor
cell expression of PD-L1 is a prerequisite for PD-1/PD-L1
immunotherapy, the present study focused more on the mechanisms by
which high dose SP (40 mM) regulated PD-L1 expression, to explore
new ways to enhance the sensitivity of PD-1/PD-L1
immunotherapy.
Screening for potential upstream
molecules that mediate PD-L1 expression
The present study explored whether m6A modification
mediated the SP-induced aberrant upregulation of PD-L1. Predictions
using the m6A2Target database identified 17 candidate
m6A-associated proteins upstream of PD-L1 (Tables I and II). Additionally, the R package ggplot2
(v3.3.6) analysis suggested that 12 proteins were positively
correlated with PD-L1 (Fig. 3A). Of
these, IGF2BP3 was aberrantly upregulated in tumor tissues compared
with healthy tissues (Fig. 3B).
Additionally, high IGF2BP3 expression was significantly associated
with poor prognosis in patients with high-grade tumors (T3/T4, N2
and M1; Fig. 3C-G). Therefore,
IGF2BP3 was selected for subsequent analyses.
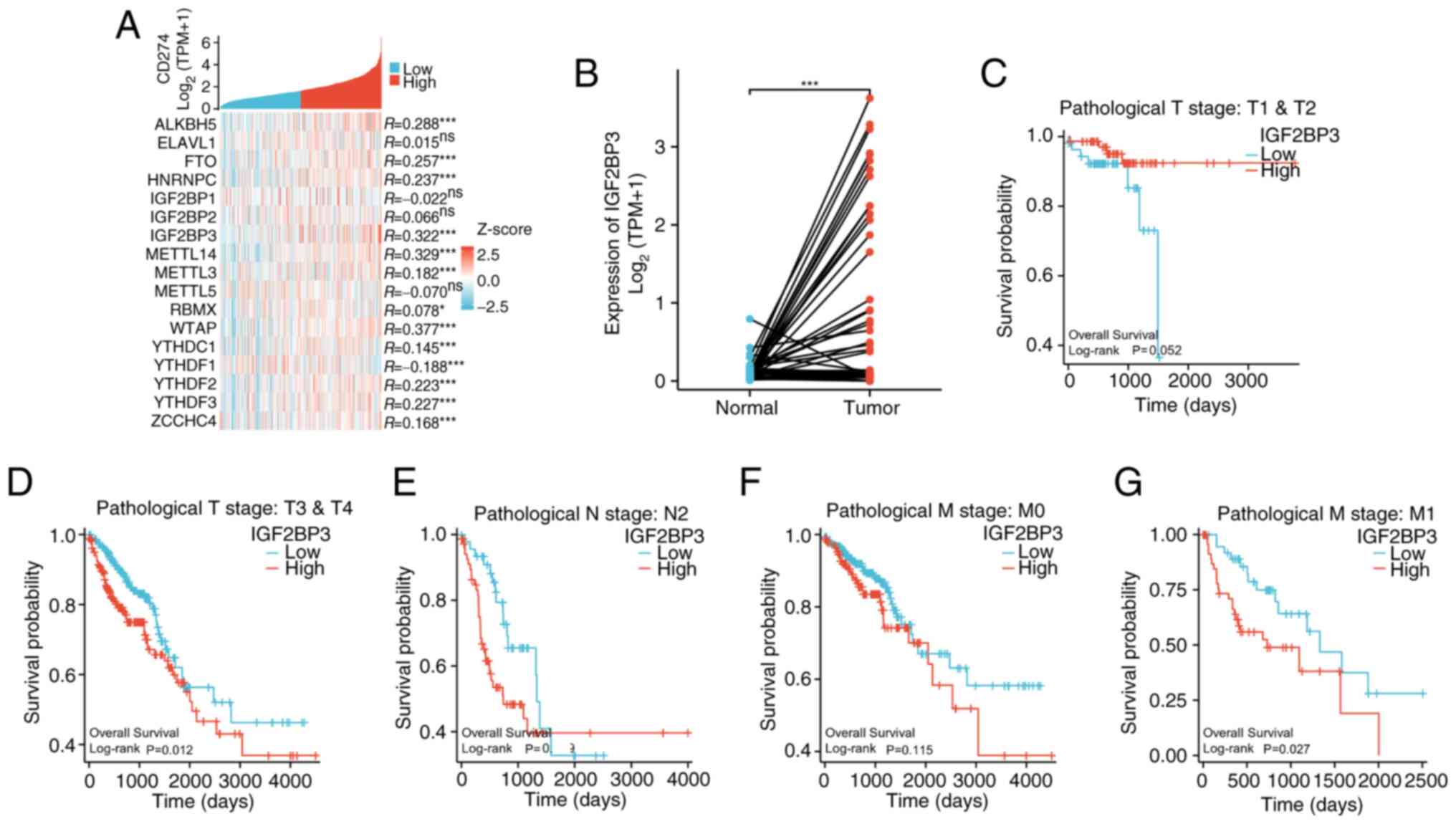 | Figure 3.Screening for potential upstream
molecules mediating PD-L1 expression regulation. (A) Genes with
similar expression pattern to PD-L1 were analyzed by R package
ggplot2 (v3.3.6). (B) The expression of IGF2BP3 in COAD and READ
tissues along with their paired normal tissues in TCGA database was
analyzed by Wilcoxon signed-rank test. (C-G) The PD-L1 expression
data was downloaded from the COAD and READ datasets in TCGA
database, and the associations between survival and different
pathological grades were analyzed by log-rank test. *P<0.05,
***P<0.001. ns, not statistically significant; PD-L1, programmed
cell death ligand-1; COAD, colon adenocarcinoma; READ, rectum
adenocarcinoma; TCGA, The Cancer Genome Atlas; TPM, transcripts per
million; T, tumor; N, node; M, metastasis. |
 | Table I.N6-methyladenosine-associated
proteins upstream of programmed cell death ligand-1 in Homo
sapiens, as predicted by the m6A2Target database. |
Table I.
N6-methyladenosine-associated
proteins upstream of programmed cell death ligand-1 in Homo
sapiens, as predicted by the m6A2Target database.
| RM2Target ID | WERs name | WERs type | Target gene |
|---|
|
RM2Target_629483 | ALKBH5 | Eraser | CD274 |
|
RM2Target_629484 | ALKBH5 | Eraser | CD274 |
|
RM2Target_629485 | ALKBH5 | Eraser | CD274 |
|
RM2Target_629486 | ALKBH5 | Eraser | CD274 |
|
RM2Target_795954 | ELAVL1 | Reader | CD274 |
|
RM2Target_795955 | ELAVL1 | Reader | CD274 |
|
RM2Target_690420 | FTO | Eraser | CD274 |
|
RM2Target_690418 | FTO | Eraser | CD274 |
|
RM2Target_690416 | FTO | Eraser | CD274 |
|
RM2Target_690417 | FTO | Eraser | CD274 |
|
RM2Target_690419 | FTO | Eraser | CD274 |
|
RM2Target_875022 | HNRNPC | Reader | CD274 |
|
RM2Target_905481 | IGF2BP1 | Reader | CD274 |
|
RM2Target_905482 | IGF2BP1 | Reader | CD274 |
|
RM2Target_905483 | IGF2BP1 | Reader | CD274 |
|
RM2Target_905484 | IGF2BP1 | Reader | CD274 |
|
RM2Target_969166 | IGF2BP2 | Reader | CD274 |
|
RM2Target_983717 | IGF2BP3 | Reader | CD274 |
|
RM2Target_174114 | METTL14 | Writer | CD274 |
|
RM2Target_174111 | METTL14 | Writer | CD274 |
|
RM2Target_174112 | METTL14 | Writer | CD274 |
|
RM2Target_174113 | METTL14 | Writer | CD274 |
|
RM2Target_268126 | METTL3 | Writer | CD274 |
|
RM2Target_268130 | METTL3 | Writer | CD274 |
|
RM2Target_268131 | METTL3 | Writer | CD274 |
|
RM2Target_268127 | METTL3 | Writer | CD274 |
|
RM2Target_268123 | METTL3 | Writer | CD274 |
|
RM2Target_268124 | METTL3 | Writer | CD274 |
|
RM2Target_268125 | METTL3 | Writer | CD274 |
|
RM2Target_268128 | METTL3 | Writer | CD274 |
|
RM2Target_268129 | METTL3 | Writer | CD274 |
|
RM2Target_268132 | METTL3 | Writer | CD274 |
|
RM2Target_390715 | METTL5 | Writer | CD274 |
|
RM2Target_1026942 | RBMX | Reader | CD274 |
|
RM2Target_562730 | WTAP | Writer | CD274 |
|
RM2Target_562729 | WTAP | Writer | CD274 |
|
RM2Target_562728 | WTAP | Writer | CD274 |
|
RM2Target_1102913 | YTHDC1 | Reader | CD274 |
|
RM2Target_1102912 | YTHDC1 | Reader | CD274 |
|
RM2Target_1141739 | YTHDF1 | Reader | CD274 |
|
RM2Target_1141737 | YTHDF1 | Reader | CD274 |
|
RM2Target_1141736 | YTHDF1 | Reader | CD274 |
|
RM2Target_1141738 | YTHDF1 | Reader | CD274 |
|
RM2Target_1181191 | YTHDF2 | Reader | CD274 |
|
RM2Target_1181189 | YTHDF2 | Reader | CD274 |
|
RM2Target_1181190 | YTHDF2 | Reader | CD274 |
|
RM2Target_1181188 | YTHDF2 | Reader | CD274 |
|
RM2Target_1233355 | YTHDF3 | Reader | CD274 |
|
RM2Target_611035 | ZCCHC4 | Writer | CD274 |
| Table II.Validation and contextual information
of predicted m6A-associated proteins interacting with PD-L1 in
various cell lines, as identified by the m6A2Target database. |
Table II.
Validation and contextual information
of predicted m6A-associated proteins interacting with PD-L1 in
various cell lines, as identified by the m6A2Target database.
| RM2Target ID | Cell line | Validated | Binding | Perturbation | Confidence | Motif |
|---|
|
RM2Target_629483 | THP1 | - | - | 44595 | 2 | 44638 |
|
RM2Target_629484 | HCCC9810 | 1 | - | - | 1 | 44638 |
|
RM2Target_629485 | RBE | 1 | - | - | 1 | 44638 |
|
RM2Target_629486 | MOLM13 | - | - | 44562 | 1 | 44638 |
|
RM2Target_795954 | THP1 | - | 44562 | 44562 | 2 | 44758 |
|
RM2Target_795955 | T24 | - | 44562 | - | 1 | 44758 |
|
RM2Target_690420 | HCT116 | 1 | - | - | 1 | 44731 |
|
RM2Target_690418 | HeLa | - | 44562 | - | 1 | 44731 |
|
RM2Target_690416 | 253J | - | - | 44562 | 1 | 44731 |
|
RM2Target_690417 | NB4 | - | - | 44562 | 1 | 44731 |
|
RM2Target_690419 | Huh7 | - | - | 44562 | 1 | 44731 |
|
RM2Target_875022 | MCF7 | - | - | 44562 | 1 | 44852 |
|
RM2Target_905481 | ES2 | - | - | 44594 | 2 | 44728 |
|
RM2Target_905482 | MV3 | - | - | 44594 | 2 | 44728 |
|
RM2Target_905483 | K562 | - | 44562 | - | 1 | 44728 |
|
RM2Target_905484 | SK-N-AS | - | - | 44562 | 1 | 44728 |
|
RM2Target_969166 | J82 | - | - | 44562 | 1 | 44669 |
|
RM2Target_983717 | PL45 | - | 44562 | - | 1 | 44729 |
|
RM2Target_174114 | HEK293T | - | 44562 | 44595 | 3 | 44578 |
|
RM2Target_174111 | A549 | - | - | 44562 | 1 | 44578 |
|
RM2Target_174112 | HSC3 | - | - | 44562 | 1 | 44578 |
|
RM2Target_174113 | IMR-90 | - | - | 44562 | 1 | 44578 |
|
RM2Target_268126 | HeLa | - | 44564 | 44598 | 3 | 44669 |
|
RM2Target_268130 | HEK293T | - | 44563 | 44596 | 3 | 44669 |
|
RM2Target_268131 | LNCaP | - | - | 44595 | 2 | 44669 |
|
RM2Target_268127 | SCC9 | 1 | - | - | 1 | 44669 |
|
RM2Target_268123 | HUVEC | - | - | 44562 | 1 | 44669 |
|
RM2Target_268124 | DKO-1 | - | - | 44562 | 1 | 44669 |
|
RM2Target_268125 | EndoC-βH1 | - | - | 44562 | 1 | 44669 |
|
RM2Target_268128 | U2OS | - | - | 44562 | 1 | 44669 |
|
RM2Target_268129 | HEC-1A | - | - | 44563 | 1 | 44669 |
|
RM2Target_268132 | HEL | - | - | 44562 | 1 | 44669 |
|
RM2Target_390715 | HeLa | - | - | 44563 | 1 | 44700 |
|
RM2Target_1026942 | HEK293T | - | 44563 | - | 1 | 44729 |
|
RM2Target_562730 | HEL | - | - | 44594 | 2 | 44574 |
|
RM2Target_562729 | HeLa | - | 44564 | - | 1 | 44574 |
|
RM2Target_562728 | MDA-LM2 | - | - | 44562 | 1 | 44574 |
|
RM2Target_1102913 | HeLa | - | 44562 | - | 1 | 44632 |
|
RM2Target_1102912 | MOLM13 | - | - | 44563 | 1 | 44632 |
|
RM2Target_1141739 | PC9 | 1 | - | - | 1 | 44633 |
|
RM2Target_1141737 | A549 | - | 44562 | - | 1 | 44633 |
|
RM2Target_1141736 | A172 | - | - | 44562 | 1 | 44633 |
|
RM2Target_1141738 | AGS | - | - | 44562 | 1 | 44633 |
|
RM2Target_1181191 | PC9 | 1 | - | - | 1 | 44758 |
|
RM2Target_1181189 | H1299 | - | 44562 | - | 1 | 44758 |
|
RM2Target_1181190 | GSC11 | - | 44562 | - | 1 | 44758 |
|
RM2Target_1181188 | HeLa | - | - | 44564 | 1 | 44758 |
|
RM2Target_1233355 | MDA-MB-231 | - | 44562 | - | 1 | 44664 |
|
RM2Target_611035 | HepG2 |
|
|
|
|
|
m6A ‘reader’ IGF2BP3 enhances PD-L1
mRNA stability in an m6A-dependent manner
First, the effects of SP on IGF2BP3 expression were
assessed. Treatment with 40 mM SP significantly upregulated IGF2BP3
mRNA and protein expression in HCT116 and SW480 cells (Fig. 4A and B). IGF2BP3 is an m6A reader
and functions by enhancing mRNA stability (26). Subsequently, it was investigated
whether IGF2BP3 could stabilize PD-L1 mRNA. ShRNA#2-induced IGF2BP3
knockdown exhibited the highest interference efficiency compared
with the sh-NC group (Fig. 4C and
D) and was selected for subsequent analyses. IGF2BP3 depletion
downregulated PD-L1 mRNA and protein expression in CRC cells
(Fig. 4E and F). Furthermore, the
addition of ActD suggested that IGF2BP3 knockdown reduced the PD-L1
mRNA stability in HCT116 and SW480 cells (Fig. 4G and H). SAH, a transmethylase
inhibitor, reduced m6A modifications of PD-L1 (Fig. 4I). Furthermore, the RIP assay
suggested that SAH weakened the interaction between PD-L1 and
IGF2BP3 (Fig. 4J). Taken together,
the results indicate that SP induces IGF2BP3 expression,
stabilizing PD-L1 mRNA in a m6A-dependent manner.
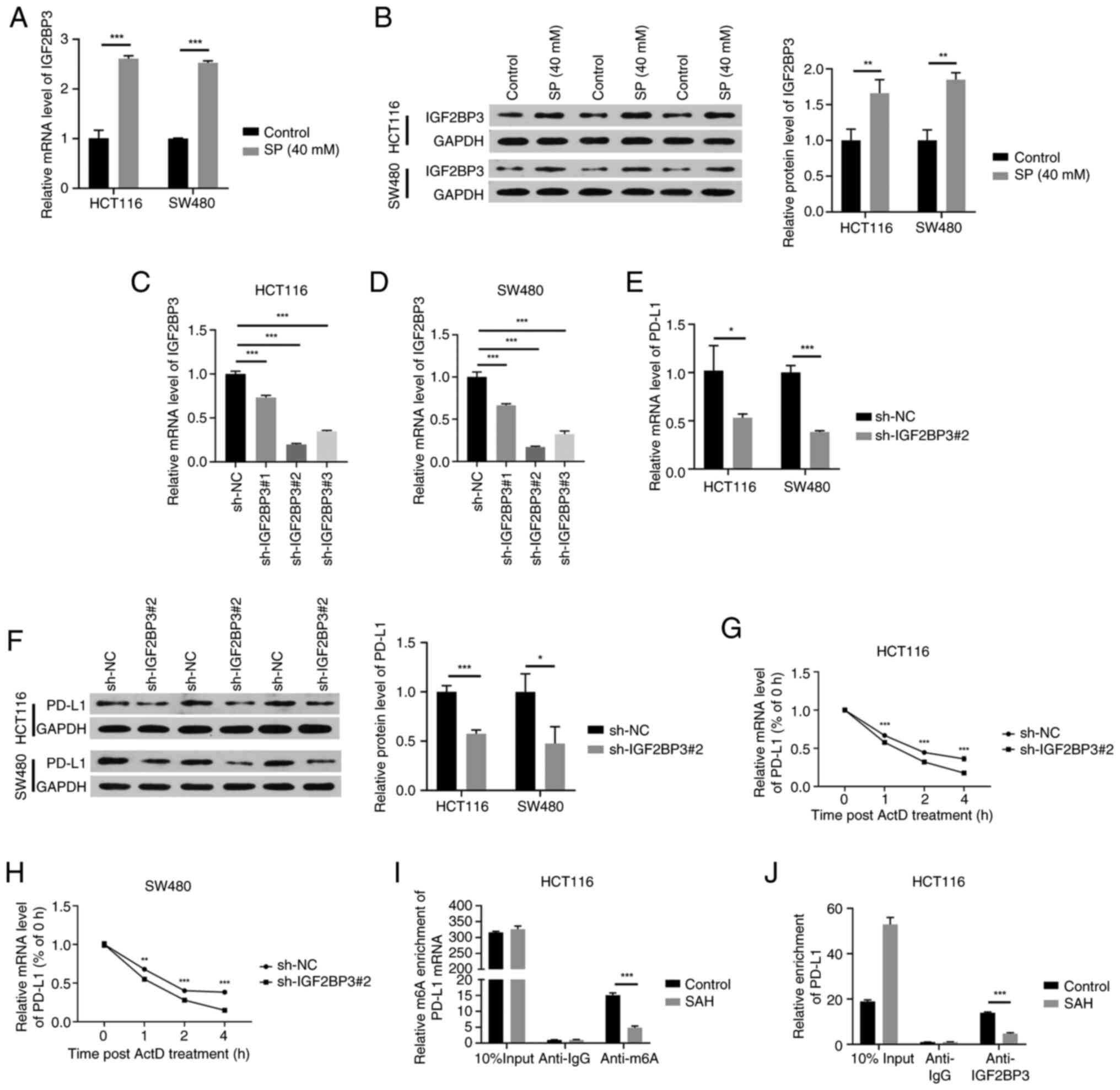 | Figure 4.m6A ‘reader’ IGF2BP3 enhances PD-L1
mRNA stability in an m6A-dependent manner. (A) RT-qPCR and (B)
western blotting were performed to detect the mRNA and protein
expression, respectively, of IGF2BP3 in CRC cells upon SP exposure
(40 mM for 48 h). The interference efficiencies of three IGF2BP3
shRNAs (sh-IGF2BP3#1, sh-IGF2BP3#2 and sh-IGF2BP3#3) in (C) HCT116
and (D) SW480 cells were evaluated by RT-qPCR. CRC cells were
transfected with sh-NC or sh-IGF2BP3#2 and the (E) mRNA and (F)
protein levels of PD-L1 were determined by RT-qPCR and western
blotting, respectively. The stability of PD-L1 mRNA upon IGF2BP3
knockdown in (G) HCT116 and (H) SW480 cells was analyzed by adding
ActD. (I) Methylated RIP-qPCR was performed to measure m6A
modification level of PD-L1 mRNA upon the addition of SAH. (J)
RIP-qPCR was implemented to test the interaction between IGF2BP3
and PD-L1 upon the addition of SAH. *P<0.05, **P<0.01,
***P<0.001. CRC, colorectal cancer; IGF2BP3, insulin-like growth
factor 2 mRNA binding protein 3; PD-L1, programmed cell death
ligand-1; SP, sodium propionate; shRNA, short hairpin RNA; NC,
negative control; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; ActD, actinomycin D; SAH,
S-adenosylhomocysteine; RIP, RNA immunoprecipitation; m6A,
N6-methyladenosine; GAPDH, glyceraldehyde-3-phosphate
dehydrogenase. |
IGF2BP3 mediates SP-induced
stabilization of PD-L1 mRNA in CRC cells
Rescue experiments were conducted to further confirm
the effects of SP on PD-L1 expression via IGF2BP3. IGF2BP3
knockdown decreased PD-L1 mRNA and protein expression; however,
these effects were partly reversed by SP (Fig. 5A and B). Additionally, IGF2BP3
depletion downregulated the stability of PD-L1 mRNA, whereas SP
treatment partly recovered its stability (Fig. 5C and D). Thus, the results indicate
that the stabilizing effect of SP on PD-L1 mRNA is dependent on
IGF2BP3 activity in CRC cells.
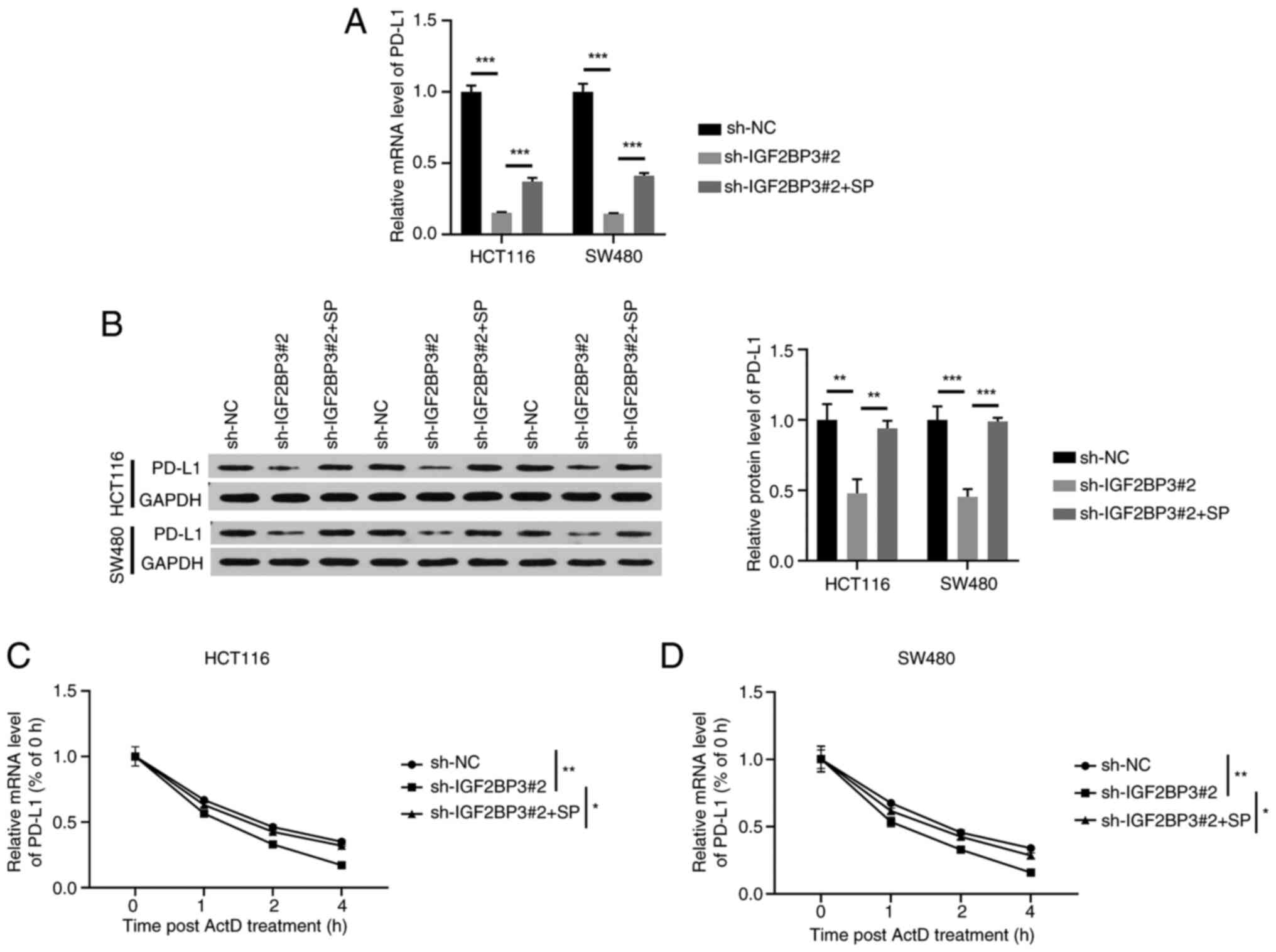 | Figure 5.IGF2BP3 mediates SP-induced
stabilization of PD-L1 mRNA in CRC cells. HCT116 and SW480 cells
were treated with sh-NC, sh-IGF2BP3#2 or sh-IGF2BP3#2 + SP and (A)
reverse transcription-quantitative polymerase chain reaction and
(B) western blotting were implemented to determine the mRNA and
protein expression of PD-L1 in CRC cells. The stability of PD-L1
mRNA in treated (C) HCT116 and (D) SW480 cells was analyzed by
adding the transcriptional inhibitor, ActD. The significance
indicated in the figures pertains to the time point at 4 h.
*P<0.05, **P<0.01, ***P<0.001. IGF2BP3, insulin-like
growth factor 2 mRNA binding protein 3; PD-L1, programmed cell
death ligand-1; SP, sodium propionate; shRNA, short hairpin RNA;
NC, negative control; ActD, actinomycin D; GAPDH,
glyceraldehyde-3-phosphate dehydrogenase. |
Discussion
SP exerts antitumor effects in multiple
malignancies, such as breast cancer (15), lung cancer (16) and CRC (27). Ryu et al (27) reported that SP treatment promoted
apoptosis in CRC by reducing the expression of protein arginine
methyltransferase 1. Similarly, in the present study, 10 mM SP
significantly restrained CRC cell proliferation and induced
apoptosis.
Cancer cells produce immunosuppressive proteins,
leading to the dysfunction and apoptosis of immune cells, thereby
promoting immune escape (28).
PD-L1, an immunosuppressive protein, interacts with PD-1 expressed
on T cells, B cells, dendritic cells and natural killer T cells,
and suppresses anticancer immune responses (29). In tumor immunology, tumors are
categorized as ‘cold’ or ‘hot’ based on their immune
microenvironment (30). ‘Hot’
tumors are characterized by high PD-L1 expression and significant
infiltration of effector T lymphocytes along with pro-inflammatory
signals, such as interferons and interleukin-2. This immunogenic
environment renders ‘hot’ tumors more responsive to PD-1/PD-L1
checkpoint inhibitors due to their active immune response (31,32).
By contrast, ‘cold’ tumors are characterized by low or no PD-L1
expression along with reduced or absent T-cell infiltration. Poor
immune activation leads to weak antitumor immune responses,
rendering these tumors less responsive to PD-1/PD-L1 blockade
therapies (32). CRC is a common
example of a ‘cold’ tumor and it demonstrates poor sensitivity to
immunotherapy. Researchers are focusing on transforming ‘cold’
tumors into ‘hot’ tumors to increase the efficacy and response
rates of cancer immunotherapy (33). A promising strategy involves
upregulating PD-L1 expression and promoting T-cell infiltration
within the tumor microenvironment (33). Ubiquitin-specific protease 8
inhibition can enhance the K63-linked ubiquitination of PD-L1
mediated by tumor necrosis factor receptor-associated factor 6
while counteracting its K48-linked ubiquitination; this results in
elevated PD-L1 protein expression (34). This regulatory pathway holds promise
in reshaping the tumor immune microenvironment to improve the
effectiveness of PD-1/PD-L1 blockade therapies. Therefore, the role
of high PD-L1 expression in CRC lies in its ability to alter the
tumor immune microenvironment from ‘cold’ to ‘hot’, improving T
cell activity and responsiveness to PD-1/PD-L1 inhibitors.
High-dose drugs can induce immune escape-related acquired drug
resistance in tumor cells (35).
Therefore, the role of high-dose SP on PD-L1 expression in CRC
cells was assessed in the present study. High-dose SP (40 mM)
significantly upregulated the PD-L1 mRNA and protein expression
levels, indicating the implication of SP in PD-L1-mediated immune
escape in CRC cells.
m6A methylation is crucial in regulating RNA
metabolism, including mRNA splicing, intracellular localization and
stability (36). m6A methylation is
dynamic and reversible, facilitated by RNA methyltransferases
(writers), RNA demethylases (erasers) and m6A reader proteins
(36). m6A modification affects
tumor onset and progression by regulating the expression of
oncogenes and tumor suppressor genes (37). Emerging evidence has suggested that
m6A modification plays a significant role in the immune response
and tumor microenvironment, influencing cancer immunotherapy
effectiveness (38). In the present
study, we hypothesized that m6A modification-related proteins
mediated the high-dose SP-induced upregulation of PD-L1. In the
bioinformatics analysis, the expression pattern of IGF2BP3 was
correlated with PD-L1. SP treatment notably upregulated both
IGF2BP3 mRNA and protein expression in CRC cells. Additionally,
IGF2BP3 positively regulated the PD-L1 mRNA and protein expression
levels in CRC cells. Furthermore, the meRIP and RIP assays
confirmed IGF2BP3 as an m6A reader for PD-L1. Data from rescue
experiments demonstrated that the stabilizing effect of SP on PD-L1
mRNA was dependent on IGF2BP3 activity in CRC cells.
The present study is unique in identifying the
specific role of high-dose SP in upregulating the m6A reader,
IGF2BP3. This, in turn, stabilizes PD-L1 mRNA and may facilitate
immune evasion in CRC. Previous studies have outlined the oncogenic
role of IGF2BP3 in CRC. For instance, Chen et al (39) illustrated the ability of IGF2BP3 to
stabilize EGFR mRNA in an m6A-dependent manner, contributing to
cetuximab resistance and CRC progression. In addition, Xu et
al (40) showed the involvement
of IGF2BP3 in promoting aggressive cancer phenotypes in CRC,
including enhanced proliferation and migration, which was
correlated with poor patient prognosis. Together, these studies
established IGF2BP3 as a key factor in CRC oncogenesis. Wan et
al (41) has elucidated the
role of the methyltransferase 3, N6-adenosine-methyltransferase
complex catalytic subunit/IGF2BP3 axis in PD-L1 expression in
breast cancer, and the present study extends these findings to CRC.
By introducing SP as a novel regulatory element in this pathway,
the present study adds a new dimension to the understanding of
IGF2BP3′s role in CRC.
While the present study revealed that high-dose SP
enhanced PD-L1 expression by upregulating IGF2BP3 in CRC cells and
highlighted the potential of SP combined with PD-1/PD-L1 blockade
for CRC treatment, there are several limitations to consider.
Primarily, the present study relied heavily on in vitro
experiments and lacked in vivo supporting data. In future
studies, we intend to validate the effects of SP and PD-L1
combination therapy in animal models to assess its actual efficacy
and safety in a complex biological environment. Additionally,
further exploration of the precise molecular interactions between
SP, IGF2BP3 and PD-L1 are needed to improve the understanding of
their roles in immune regulation and potential influencing factors.
These investigations will provide important scientific foundations
for the translation of SP and PD-L1 antibody combination therapy
strategies into clinical applications.
In conclusion, in the present study, high-dose SP
treatment enhanced PD-L1 expression by upregulating IGF2BP3
expression in CRC cells. This finding supports the potential for
combination therapies using SP and PD-L1 antibodies for CRC
treatment.
Acknowledgements
Not applicable.
Funding
This research was supported by the Natural Science Foundation
project of Hubei Province (grant no. 2017CFB571) and the Health
Commission of Hubei Province (grant no. WJ2023F051).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
XW designed and performed the experiments, analysed
and interpretated data and wrote the manuscript. YH contributed to
the conception and design of the study. XW and YH confirm the
authenticity of all the raw data. Both authors have read and
approved the final version of the manuscript.
Ethical approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Giaquinto AN and Jemal A:
Cancer statistics, 2024. CA Cancer J Clin. 74:12–49. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Biller LH and Schrag D: Diagnosis and
treatment of metastatic colorectal cancer: A review. JAMA.
325:669–685. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ganesh K, Stadler ZK, Cercek A, Mendelsohn
RB, Shia J, Segal NH and Diaz LA Jr: Immunotherapy in colorectal
cancer: Rationale, challenges and potential. Nat Rev Gastroenterol
Hepatol. 16:361–375. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yi M, Zheng X, Niu M, Zhu S, Ge H and Wu
K: Combination strategies with PD-1/PD-L1 blockade: Current
advances and future directions. Mol Cancer. 21:282022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arifuzzaman M, Collins N, Guo CJ and Artis
D: Nutritional regulation of microbiota-derived metabolites:
Implications for immunity and inflammation. Immunity. 57:14–27.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu H, Mu C, Xu L, Yu K, Shen L and Zhu W:
Host-microbiota interaction in intestinal stem cell homeostasis.
Gut Microbes. 16:23533992024. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hurst NR, Kendig DM, Murthy KS and Grider
JR: The short chain fatty acids, butyrate and propionate, have
differential effects on the motility of the guinea pig colon.
Neurogastroenterol Motil. 26:1586–1596. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meyer JH, Cervenka S, Kim MJ, Kreisl WC,
Henter ID and Innis RB: Neuroinflammation in psychiatric disorders:
PET imaging and promising new targets. Lancet Psychiatry.
7:1064–1074. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Morrison DJ and Preston T: Formation of
short chain fatty acids by the gut microbiota and their impact on
human metabolism. Gut Microbes. 7:189–200. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He J, Zhang P, Shen L, Niu L, Tan Y, Chen
L, Zhao Y, Bai L, Hao X, Li X, et al: Short-chain fatty acids and
their association with signalling pathways in inflammation, glucose
and lipid metabolism. Int J Mol Sci. 21:63562020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Filippone A, Casili G, Scuderi SA, Mannino
D, Lanza M, Campolo M, Paterniti I, Capra AP, Colarossi C, Bonasera
A, et al: Sodium propionate contributes to tumor cell growth
inhibition through PPAR-γ signaling. Cancers (Basel). 15:2172022.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wong JM, de Souza R, Kendall CW, Emam A
and Jenkins DJ: Colonic health: Fermentation and short chain fatty
acids. J Clin Gastroenterol. 40:235–243. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tran NL, Lee IK, Choi J, Kim SH and Oh SJ:
Acetate decreases PVR/CD155 expression via PI3K/AKT pathway in
cancer cells. BMB Rep. 54:431–436. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peng Z, Cheng S, Kou Y, Wang Z, Jin R, Hu
H, Zhang X, Gong JF, Li J, Lu M, et al: The gut microbiome is
associated with clinical response to anti-PD-1/PD-L1 immunotherapy
in gastrointestinal cancer. Cancer Immunol Res. 8:1251–1261. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park HS, Han JH, Park JW, Lee DH, Jang KW,
Lee M, Heo KS and Myung CS: Sodium propionate exerts anticancer
effect in mice bearing breast cancer cell xenograft by regulating
JAK2/STAT3/ROS/p38 MAPK signaling. Acta Pharmacol Sin.
42:1311–1323. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim K, Kwon O, Ryu TY, Jung CR, Kim J, Min
JK, Kim DS, Son MY and Cho HS: Propionate of a microbiota
metabolite induces cell apoptosis and cell cycle arrest in lung
cancer. Mol Med Rep. 20:1569–1574. 2019.PubMed/NCBI
|
|
17
|
Khalaf K, Hana D, Chou JT, Singh C,
Mackiewicz A and Kaczmarek M: Aspects of the tumor microenvironment
involved in immune resistance and drug resistance. Front Immunol.
12:6563642021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Peng S, Wang R, Zhang X, Ma Y, Zhong L, Li
K, Nishiyama A, Arai S, Yano S and Wang W: EGFR-TKI resistance
promotes immune escape in lung cancer via increased PD-L1
expression. Mol Cancer. 18:1652019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tsai TF, Lin JF, Lin YC, Chou KY, Chen HE,
Ho CY, Chen PC and Hwang TI: Cisplatin contributes to programmed
death-ligand 1 expression in bladder cancer through ERK1/2-AP-1
signaling pathway. Biosci Rep. 39:BSR201903622019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fournel L, Wu Z, Stadler N, Damotte D,
Lococo F, Boulle G, Ségal-Bendirdjian E, Bobbio A, Icard P,
Trédaniel J, et al: Cisplatin increases PD-L1 expression and
optimizes immune check-point blockade in non-small cell lung
cancer. Cancer Lett. 464:5–14. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang X, Wu R, Liu Y, Zhao Y, Bi Z, Yao Y,
Liu Q, Shi H, Wang F and Wang Y: m(6)A mRNA methylation controls
autophagy and adipogenesis by targeting Atg5 and Atg7. Autophagy.
16:1221–1235. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peritz T, Zeng F, Kannanayakal TJ, Kilk K,
Eiríksdóttir E, Langel U and Eberwine J: Immunoprecipitation of
mRNA-protein complexes. Nat Protoc. 1:577–580. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
MIto M, Mimura K, Nakajima S, Saito K, Min
AKT, Okayama H, Saito M, Momma T, Saze Z, Ohtsuka M, et al: Immune
escape mechanism behind resistance to anti-PD-1 therapy in
gastrointestinal tract metastasis in malignant melanoma patients
with multiple metastases. Cancer Immunol Immunother. 71:2293–2300.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu J, Chen Z, Li Y, Zhao W, Wu J and
Zhang Z: PD-1/PD-L1 checkpoint inhibitors in tumor immunotherapy.
Front Pharmacol. 12:7317982021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiang T, He X, Zhao Z, Zhang X, Wang T and
Jia L: RNA m6A reader IGF2BP3 promotes metastasis of
triple-negative breast cancer via SLIT2 repression. FASEB J.
36:e226182022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ryu TY, Kim K, Son MY, Min JK, Kim J, Han
TS, Kim DS and Cho HS: Downregulation of PRMT1, a histone arginine
methyltransferase, by sodium propionate induces cell apoptosis in
colon cancer. Oncol Rep. 41:1691–1699. 2019.PubMed/NCBI
|
|
28
|
Lei X, Lei Y, Li JK, Du WX, Li RG, Yang J,
Li J, Li F and Tan HB: Immune cells within the tumor
microenvironment: Biological functions and roles in cancer
immunotherapy. Cancer Lett. 470:126–133. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu Q, Jiang L, Li SC, He QJ, Yang B and
Cao J: Small molecule inhibitors targeting the PD-1/PD-L1 signaling
pathway. Acta Pharmacol Sin. 42:1–9. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang J, Huang D, Saw PE and Song E:
Turning cold tumors hot: From molecular mechanisms to clinical
applications. Trends Immunol. 43:523–545. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rameshbabu S, Labadie BW, Argulian A and
Patnaik A: Targeting innate immunity in cancer therapy. Vaccines
(Basel). 9:1382021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Karin N: Chemokines in the landscape of
cancer immunotherapy: How they and their receptors can be used to
turn cold tumors into hot ones? Cancers (Basel). 13:63172021.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin KX, Istl AC, Quan D, Skaro A, Tang E
and Zheng X: PD-1 and PD-L1 inhibitors in cold colorectal cancer:
Challenges and strategies. Cancer Immunol Immunother. 72:3875–3893.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiong W, Gao X, Zhang T, Jiang B, Hu MM,
Bu X, Gao Y, Zhang LZ, Xiao BL, He C, et al: USP8 inhibition
reshapes an inflamed tumor microenvironment that potentiates the
immunotherapy. Nat Commun. 13:17002022. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cao J and Yan Q: Cancer epigenetics, tumor
immunity, and immunotherapy. Trends Cancer. 6:580–592. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
An Y and Duan H: The role of m6A RNA
methylation in cancer metabolism. Mol Cancer. 21:142022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sun T, Wu R and Ming L: The role of m6A
RNA methylation in cancer. Biomed Pharmacother. 112:1086132019.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pan J, Huang T, Deng Z and Zou C: Roles
and therapeutic implications of m6A modification in cancer
immunotherapy. Front Immunol. 14:11326012023. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen LJ, Liu HY, Xiao ZY, Qiu T, Zhang D,
Zhang LJ, Han FY, Chen GJ, Xu XM and Zhu JH: IGF2BP3 promotes the
progression of colorectal cancer and mediates cetuximab resistance
by stabilizing EGFR mRNA in an m(6)A-dependent manner. Cell Death
Dis. 14:5812023. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu W, Sheng Y, Guo Y, Huang Z, Huang Y,
Wen D, Liu CY, Cui L, Yang Y and Du P: Increased IGF2BP3 expression
promotes the aggressive phenotypes of colorectal cancer cells in
vitro and vivo. J Cell Physiol. 234:18466–18479. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wan W, Ao X, Chen Q, Yu Y, Ao L, Xing W,
Guo W, Wu X, Pu C, Hu X, et al: METTL3/IGF2BP3 axis inhibits tumor
immune surveillance by upregulating N(6)-methyladenosine
modification of PD-L1 mRNA in breast cancer. Mol Cancer. 21:602022.
View Article : Google Scholar : PubMed/NCBI
|