Introduction
According to the World Health Organization, lung
cancer is one of the most rapidly growing malignancies in terms of
both morbidity and mortality worldwide (1). Histopathologically, lung cancer is
mainly classified into small-cell lung cancer and non-small cell
lung cancer (NSCLC). NSCLC accounts for 80–85% of all lung cancer
cases, and lung adenocarcinoma, squamous-cell carcinoma and
large-cell carcinoma are the three main subtypes of NSCLC (2,3).
Currently, surgical resection, radiotherapy and chemotherapy are
the most common treatment options for NSCLC. Although advances have
been made in the clinical diagnosis and treatment of lung cancer,
early-stage lung cancer is commonly asymptomatic and ~75% of
patients are diagnosed at advanced stages of the disease (2). Due to the lack of specific symptoms,
limited therapeutic efficacy and poor prognosis, the global overall
5-year survival rate of NSCLC remains at ~15% (4–6).
Therefore, the development of highly effective and low-toxicity
chemotherapeutic agents for NSCLC is of urgent importance.
Due to their diverse pharmacological properties and
low toxicity, natural products are considered important sources of
anticancer compounds. Jaceosidin, a flavonoid active ingredient
mainly derived from plants such as mugwort leaf and Eupatorium
lindleyanum, exhibits several biological and pharmacological
effects, including anti-inflammatory, antiviral, antioxidant,
antidiabetic, antiallergic, antibacterial and antitumor properties
(7,8). In vitro studies have suggested
that Jaceosidin exerts anticancer effects on several types of
cancer (9,10). A previous study showed that
Jaceosidin can induce apoptosis in Caki human kidney cancer cells
by downregulating myeloid cell leukemia 1 and cellular
FLICE-inhibitory protein at the transcriptional level (10). Jaceosidin can also induce apoptosis
in Ras-transformed human mammary epithelial cells (MCF10A-ras) by
upregulating Bax, downregulating Bcl-2 and enhancing reactive
oxygen species (ROS) production by ≤40% (11). Jeong et al (12) revealed that Jaceosidin could restore
TPA-induced cyclooxygenase 2 and matrix metallopeptidase 9
upregulation by inhibiting ERK1/2 phosphorylation in MCF10A cells,
eventually inhibiting cell invasion and migration. In addition,
Jaceosidin has been shown to induce G2/M cell cycle
arrest in U87 glioblastoma cells and Hec1A human endometrial cancer
cells through upregulating p53 and p21, and modulating the
ERK/ataxia-telangiectasia mutated kinase/checkpoint kinase 1/2
pathway, thereby attenuating cell proliferation (13,14).
These previous findings indicated that Jaceosidin may exert potent
anticancer effects; however, to the best of our knowledge, its
pro-apoptotic and anti-survival effects in NSCLC cells have not
been previously investigated. Therefore, the current study aimed to
explore the capacity of Jaceosidin in inhibiting cell viability and
inducing cell apoptosis in the human NSCLC cell line A549.
Materials and methods
Cell culture
The A549 NSCLC cell line and 293T cells were
purchased from Kunming Cell Bank of Type Culture Collection,
Chinese Academy of Science. A549 and 293T cells were cultured in
RPMI 1640 and DMEM (both from Gibco; Thermo Fisher Scientific,
Inc.), respectively, supplemented with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and
100 µg/ml streptomycin at 37°C in a constant temperature incubator
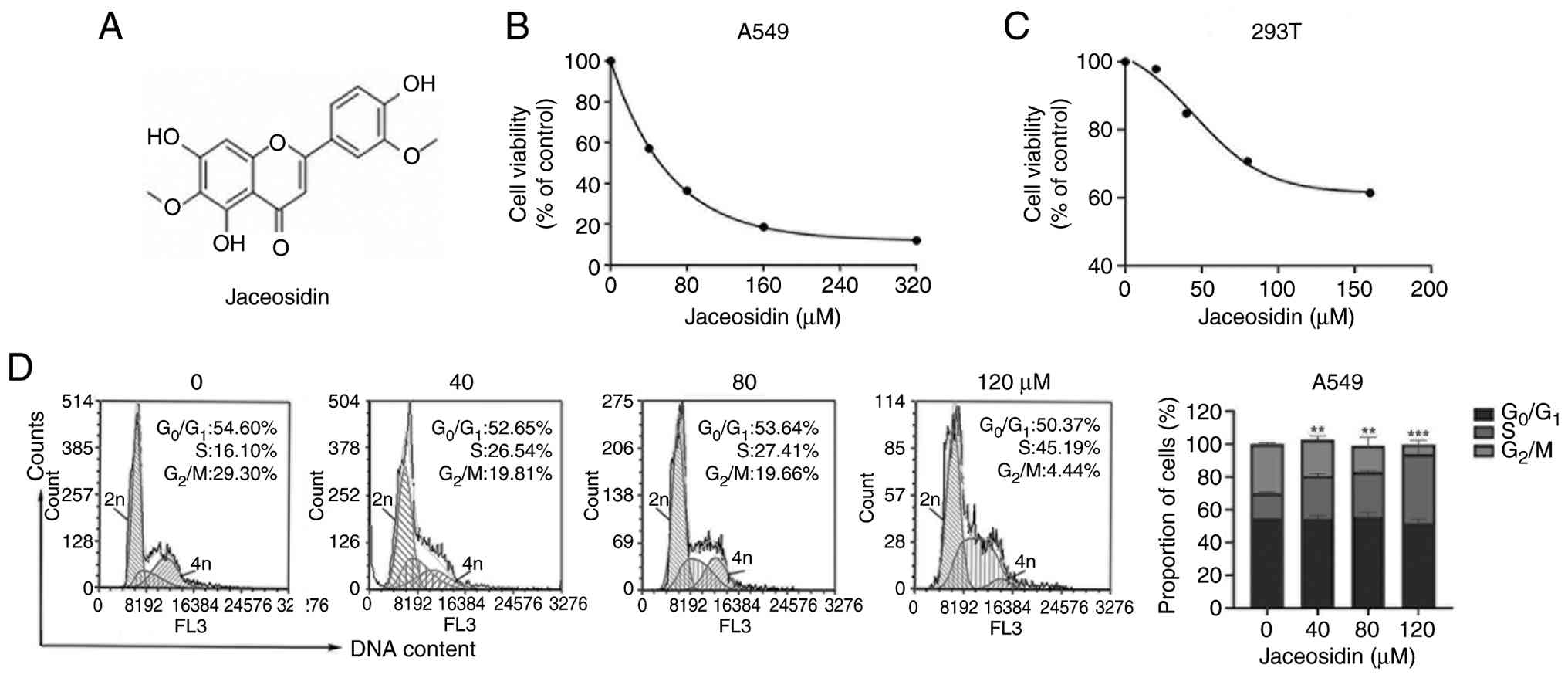
containing 5% CO2. Jaceosidin (Fig. 1A) was obtained from Wuhan ChemFaces
Biochemical Co., Ltd., and was prepared as a 100 mM storage
solution in DMSO.
Cell viability assay
Cell viability was assessed using the MTS method
(Promega Corporation). Briefly, A549 and 293T cells at a density of
1×104 cells/well were inoculated in 96-well culture
plates with three replicates per group, and the cells were cultured
under saturated humidity at 37°C and 5% CO2. Following
treatment of A549 cells with different concentrations of Jaceosidin
(0, 40, 80, 160 and 320 µM) for 68 h at 37°C, the cells were
co-treated with 20 µl MTS/PMS (20:1 v/v) for an additional 4 h. For
293T cells, the treatment procedure was identical, and the cells
were treated with difference concentrations of Jaceosidin (0, 20,
40, 80 and 160 µM). The optical density values in each well were
measured at 490 nm using a microplate reader (Nanjing Detai
Bioengineering Co., Ltd.). Finally, cell viability was calculated
using the following formula: Cell viability (%)=(absorbance value
of drug-treated group - absorbance value of blank
group)/(absorbance value of control group-absorbance value of blank
group) ×100. The IC50 values were calculated by
non-linear regression curve fitting using Microsoft Excel (version
2016; Microsoft Corporation).
Cell cycle distribution
Following treatment of A549 cells (2×105
cells/well in 6-well plates, 2 ml medium/well) with different
concentrations of Jaceosidin (0, 40, 80 and 120 µM) for 24 h at
37°C, cell precipitates were collected by centrifugation at 300 × g
for 5 min at 4°C. Subsequently, DNA content was measured using a
commercial cell cycle assay kit (Beijing Solarbio Science &
Technology Co., Ltd.). Briefly, cell pellets were fixed overnight
in 75% ice-cold ethanol at 4°C. Prior to staining, the cells were
washed with PBS, resuspended in 100 µl RNase A solution and
incubated for 30 min at 37°C in a water bath, followed by staining
with 400 µl propidium iodide (PI; MilliporeSigma) for 30 min at 4°C
in the dark. Cell cycle distribution was then analyzed by flow
cytometry using a BD FACSCanto flow cytometer (BD Biosciences) with
BD FACSDiva software (version 8.0; BD Biosciences).
Flow cytometric analysis of
apoptosis
Apoptosis was assessed using the Annexin V-FITC/PI
Apoptosis Detection Kit (Absin Bioscience Inc.). A549 cells
(2×105 cells/well in 6-well plates, 2 ml medium/well)
were treated with different concentrations of Jaceosidin (0, 40, 80
and 120 µM) for 24 h at 37°C, and subsequently harvested by
trypsinization. The cells were then stained with Annexin V-FITC and
PI for 30 min at room temperature in the dark. The proportions of
viable, necrotic, and early and late apoptotic cells were analyzed
by flow cytometry using a BD FACSCanto flow cytometer with BD
FACSDiva software (version 8.0).
Western blot analysis
Untreated control and Jaceosidin-treated A549 cells
(treated with 40, 80 and 120 µM Jaceosidin for 36 h at 37°C, or
with 120 µM Jaceosidin for 12, 24 and 36 h at 37°C) were collected
by centrifugation at 300 × g for 5 min at 4°C. The cells were then
washed twice with PBS and lysed with RIPA lysis buffer (99 ml 1X
PBS, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, supplemented
with 1X Roche protease inhibitor cocktail, 1X PMSF, 0.05 M NaF and
0.05 M glycerophosphate) on ice for 30 min. After sonication (20
kHz; 3 pulses, 10 sec each, with 30-sec intervals on ice), the cell
supernatants were collected by centrifugation at 12,000 × g for 10
min at 4°C. The protein concentration was quantified using a BCA
protein concentration assay kit (Beyotime Biotechnology), and equal
amounts of protein (40 µg/lane) were mixed with loading buffer,
denatured at 100°C for 5 min and separated by SDS-PAGE on a 12%
gel. Subsequently, the proteins were transferred onto a
polyvinylidene fluoride (PVDF) membranes (MilliporeSigma). After
blocking in 5% skim milk in PBS-0.05% Tween-20 (PBST) for 1 h at
room temperature, the membranes were incubated with primary
antibodies against poly-ADP ribose polymerase (PARP; cat. no.
9542), caspase-3 (cat. no. 9662), caspase-8 (cat. no. 4790),
caspase-9 (cat. no. 9502), cytochrome c (cyto-c; cat. no.
4280), p21 (cat. no. 2947), Ras (cat. no. 3965), Raf (cat. no.
9422), ERK1/2 (cat. no. 4695), phosphorylated (p)-ERK1/2 (cat. no.
4370), Akt (cat. no. 9272), p-Akt (cat. no. 4060), Bax (cat. no.
2772) and Bcl-2 (cat. no. 4223) (all from CST Biological Reagents
Co., Ltd.) at 4°C overnight. The primary antibodies were diluted in
PBST to 1:250 (for anti-Raf), 1:8,000 (for anti-β-actin; cat. no.
4970; CST Biological Reagents Co., Ltd.) and 1:1,000 for the
remaining antibodies. After washing three times with PBST, the PVDF
membranes were incubated with the corresponding horseradish
peroxidase-labeled anti-rabbit secondary antibodies (cat. no.
31460; Thermo Fisher Scientific, Inc.) diluted 1:5,000 in PBST at
room temperature for 1 h. Immunoreactive bands were visualized
using Immobilon Western Chemiluminescent HRP Substrate (cat. no.
WBKLS0500; MilliporeSigma). After incubation with ECL, the signals
were detected by exposure to photographic films, which were
subsequently developed, fixed and dried. Band intensities were
semi-quantified and normalized to β-actin using ImageJ software
(version 1.53t; National Institutes of Health).
Cyto-c release assay
Cyto-c is released from mitochondria into the
cytosol during apoptosis (10);
therefore, cytosolic cyto-c levels were analyzed in the present
study. Following treatment with jaceosidin (0, 40, 80 and 120 µM
for 36 h, or 120 µM for 12, 24 and 36 h at 37°C), A549 were
collected and centrifuged at 300 × g for 5 min at 4°C to remove the
culture supernatant and the pellet was washed twice with PBS.
Subsequently, the cells were lysed with self-prepared 1.5%
digitonin extraction buffer (containing 20 mM Tris-HCl, pH 7.4; 140
mM NaCl; 10 mM KCl; 1 mM MgCl2; 1.5% digitonin) and
incubated on ice for 10 min. The lysates were then centrifuged at
12,000 × g for 10 min at 4°C, and the supernatant was aspirated as
the cytosolic fraction. Finally, cyto-c levels were analyzed using
the aforementioned western blotting protocol.
Statistical analysis
Each experiment was performed at least three times
and all statistical analyses were performed using GraphPad 8.0
software (Dotmatics). Data are presented as the mean ± standard
deviation, and differences among groups were assessed by one-way
ANOVA followed by Tukey's post hoc test. P<0.05 was considered
to indicate a statistically significant difference.
Results
Jaceosidin inhibits the viability of
A549 cells and induces S-phase arrest
First, A549 cells were treated with increasing
concentrations of Jaceosidin for 72 h and cell viability was
determined by MTS assays. The results demonstrated that Jaceosidin
effectively inhibited A549 cell viability in a
concentration-dependent manner, with a half maximal inhibitory
concentration (IC50) value of 55 µM (Fig. 1B). However, Jaceosidin showed less
toxicity in 293T cells, with an extrapolated IC50 value
of 248.5 µM (Fig. 1C). Since cell
viability did not drop to 50% within the tested concentration range
(0–160 µM), the IC50 value was estimated by non-linear
regression curve fitting using Microsoft Excel (Microsoft
Corporation), and 248.5 µM represents an extrapolated value beyond
the experimental range. Cell cycle arrest serves a key role in
inhibiting cell proliferation; therefore, to reveal whether
Jaceosidin could inhibit A549 cell viability via dysregulation of
cell cycle distribution, PI staining followed by flow cytometry was
carried out. Following cell treatment with 40, 80 and 120 µM
Jaceosidin for 24 h, no significant changes were observed in the
proportion of G0/G1-phase cells, compared
with the control group (Fig. 1D).
However, the ratio of S-phase cells increased from 16.10% to 26.54,
27.41 and 45.19%, respectively, whereas G2/M-phase cells
decreased from 29.30% to 19.81, 19.66 and 4.44%, respectively,
compared with control cells. These results suggested that
Jaceosidin may inhibit A549 cell viability via inducing
dose-dependent S-phase arrest.
Jaceosidin induces apoptosis in A549
cells
To determine whether Jaceosidin exhibited
pro-apoptotic effects on A549 NSCLC cells, flow cytometric analysis
was conducted using Annexin V-FITC/PI double staining. As shown in
Fig. 2A and C, treatment with
Jaceosidin for 24 h dose-dependently increased the percentage of
apoptotic A549 cells compared with that in untreated cells, with
marked induction of both early and late apoptosis at 120 µM.
Subsequently, the expression levels of several apoptosis-related
proteins, including those of PARP and caspase-3, were detected by
western blot analysis. Notably, treatment of A549 cells with
Jaceosidin resulted in a significant decrease in PARP and caspase-3
expression levels in a dose- and time-dependent manner (Fig. 2B and D). Consistently, the protein
levels of both cleaved-PARP and cleaved-caspase-3 were markedly
enhanced in a dose- and time-dependent manner. Together, these data
verified that Jaceosidin could induce apoptosis in A549 cells.
Mechanism underlying
Jaceosidin-mediated inhibition of survival and induction of
apoptosis
To elucidate the mechanism underlying the effects of
Jaceosidin on promoting A549 cell apoptosis, the protein expression
levels of caspase-8, caspase-9, Bcl-2, Bax and cyto-c were
detected. Compared with in the control group, the protein
expression levels of cleaved-caspase-8 and Bax were significantly
increased, whereas those of Bcl-2 were notably decreased in a dose-
and time-dependent manner in Jaceosidin-treated cells (Fig. 3A and C). Mitochondria play a key
role in the regulation of apoptosis. To verify whether Jaceosidin
could induce cell apoptosis via the mitochondrial pathway, the
expression levels of cytoplasmic cyto-c and cleaved-caspase-9 were
detected in A549 cells. The results demonstrated that Jaceosidin
upregulated both cyto-c (Fig. 3B and
C) and cleaved-caspase-9 (Fig. 3A
and C) in A549 cells compared with those in the control group,
which indicated release of cyto-c from mitochondria and activation
of caspase-9. Furthermore, to investigate the mechanism involved in
Jaceosidin-mediated cell cycle arrest in NSCLC cells, the protein
expression levels of p21 were examined. Western blot analysis
demonstrated that the expression levels of p21 were enhanced in a
dose- and time-dependent manner in Jaceosidin-treated A549 cells
(Fig. 3B and C). Overall, the
aforementioned findings suggested that Jaceosidin could inhibit
NSCLC cell viability and induce apoptosis by activating
caspase-dependent mitochondrial signaling and upregulating p21.
 | Figure 3.Effects of Jaceosidin on
apoptosis-related signaling proteins and the cell cycle regulator
p21. (A) Western blot analysis was performed to detect the
expression levels of the apoptosis-related proteins caspase-8,
cleaved-caspase-8, caspase-9, cleaved-caspase-9, Bcl-2 and Bax in
A549 cells treated with the indicated concentrations of Jaceosidin
for 36 h, or with 120 µM Jaceosidin for the indicated time points.
(B) Western blot analysis was carried out to determine the
expression levels of cyto-c in cytosolic fractions of A549 cells
treated with the same concentrations of Jaceosidin. Western blot
analysis of p21 was also performed under identical treatment
conditions. (C) Densitometric analysis of caspase-8,
cleaved-caspase-8, caspase-9, cleaved-caspase-9, Bcl-2, Bax, cyto-c
and p21. Data are presented as the mean ± SD of three independent
experiments (n=3). Statistical differences were analyzed by one-way
ANOVA followed by Tukey's post hoc test. *P<0.05, **P<0.01,
***P<0.001 vs. control. Cyto-c, cytochrome c; ns, not
significant. |
Jaceosidin inhibits the MAPK
(Ras/Raf/MEK/ERK) and Akt signaling pathways in A549 cells
Given the potential of MAPK and Akt signaling in
treating NSCLC (4), as these
pathways are frequently dysregulated in NSCLC and drive tumor cell
proliferation and survival, representing key therapeutic targets,
the effect of Jaceosidin on the expression levels of Ras, Raf,
ERK1/2 and Akt in NSCLC cells were determined by western blot
analysis. The results demonstrated that Jaceosidin dose- and
time-dependently reduced the protein expression levels of Ras and
Raf (Fig. 4A and B), and those of
p-ERK1/2, ERK1/2, p-Akt and Akt in A549 cells (Fig. 4A and C), indicating that Jaceosidin
may inhibit both the expression and activation of these key
signaling proteins. These findings indicated that Jaceosidin could
suppress A549 cell viability by inhibiting the MAPK and Akt
signaling pathways.
Discussion
Currently, chemotherapy remains the primary
therapeutic strategy for NSCLC (15); however, conventional anticancer
drugs commonly cause strong toxic side effects. Natural products
have long been used in traditional medicine, and particular
bioactive compounds derived from plants have been investigated as
adjuvant agents in the treatment of cancer, capable of enhancing
antitumor efficacy and alleviating chemotherapy-related side
effects. Notable progress has thus been made in identifying novel
compounds with potent and relatively low toxicity against NSCLC
(16,17). Jaceosidin, a flavonoid extracted
from plants of the genus Artemisia, exhibits potential
antitumor activity against several types of cancer (8). The results of the present study
demonstrated that Jaceosidin treatment could attenuate the
viability of A549 cells compared with 293T cells, thus indicating
that this compound could display tumor-specific cytotoxicity,
further supporting its potential against NSCLC. 293T cells were
used as a non-malignant control to evaluate the tumor selectivity
of Jaceosidin.
Dysregulation of cell cycle progression is markedly
associated with aberrant tumor cell proliferation (18). It has been reported that several
anticancer drugs can trigger cell cycle arrest at
G0/G1, S and G2/M checkpoints,
thus enabling DNA injury repair prior to mitosis (19,20);
however, if the DNA damage cannot be repaired, the prolonged cell
cycle arrest ultimately leads to cell apoptosis, thereby exerting
an anticancer effect. The results of cell cycle analysis indicated
that Jaceosidin promoted S-phase arrest, whereas no notable changes
were reported in the G0/G1 phase. S-phase
arrest has been identified as one of the primary molecular
mechanisms underlying the action of several antitumor drugs. For
example, Kim et al (21)
demonstrated that dichloromethane fractions of Calystegia
soldanella induced S-phase arrest in human colorectal cancer
cells. Additionally, Li et al (22) reported that d-borneol could enhance
the sensitivity of NSCLC cells to cisplatin via p21/p27-mediated
S-phase arrest. In the current study, A549 cell treatment with
Jaceosidin induced S-phase arrest, thus inhibiting cell
viability.
p21, a cyclin-dependent kinase inhibitor downstream
of p53, serves a key role in linking DNA repair and cell cycle
regulation by suppressing cyclin-dependent kinase activity, thereby
inhibiting cell cycle progression (23). In the current study, Jaceosidin
upregulated p21 in a concentration- and time-dependent manner, thus
suggesting that p21 induction could be involved in the observed
Jaceosidin-mediated S-phase arrest in A549 cells. The induction of
S-phase arrest by Jaceosidin is of particular biological importance
for NSCLC therapy, as S phase is a critical stage for DNA
replication and cell proliferation; blocking this phase effectively
prevents the uncontrolled division of A549 cells (22). The upregulation of p21 not only
mediates cell cycle arrest, but also acts as a bridge between cell
cycle regulation and apoptosis (23), which may synergistically enhance the
anti-survival effect of Jaceosidin. Additionally, Jaceosidin
simultaneously activated the extrinsic (caspase-8-dependent) and
intrinsic (mitochondrial/caspase-9-dependent) apoptotic pathways,
which is a notable advantage for anticancer compounds, as it avoids
the drug resistance caused by the single pathway activation in
tumor cells (24). The crosstalk
between the two apoptotic pathways further amplifies the apoptotic
signal, leading to a more significant killing effect on A549 cells,
which is consistent with the characteristics of effective natural
anticancer products (8).
Apoptosis is closely associated with tumorigenesis.
Therefore, the majority of anticancer drugs exert their effects by
inducing apoptosis (25). Apoptosis
can be triggered by the mitochondria (intrinsic pathway) or cell
death receptors (extrinsic pathway), both promoting caspase
activation (25,26). PARP is a DNA repair enzyme that
serves as a cleavage substrate for caspases, key cystathionine
aspartase proteases involved in apoptosis (27). Once PARP is cleaved during
apoptosis, its DNA repair function is impaired. The caspase family
serves a central role in mediating apoptosis, with
cleaved-caspase-3 acting as the major effector downstream of both
the intrinsic and extrinsic pathways, thus activating PARP and
inducing apoptosis (28). Caspase-9
and caspase-8 are key proteins of the intrinsic and extrinsic
pathways, respectively (25,29).
In the present study, the experimental results showed that
Jaceosidin significantly increased the cleaved forms of PARP,
caspase-3, caspase-8 and caspase-9 during apoptosis. These findings
indicated that Jaceosidin could induce apoptosis in A549 cells by
activating both the intrinsic and extrinsic apoptotic pathways.
Consistent with the aforementioned results, a previous study
reported that Jaceosidin could induce caspase-dependent apoptosis
in the oral cancer cell lines HSC3 and Ca9.22 (30).
Mitochondria are notable regulators of cell death,
and members of the Bcl-2 family, including the pro-apoptotic
protein Bax and the anti-apoptotic protein Bcl-2, serve key roles
in maintaining mitochondrial stability, and regulating apoptosis
and tumorigenesis (31). Cyto-c is
an apoptotic regulator found in mitochondria. Following apoptotic
induction, cyto-c is released into the cytosol, where it binds
apoptotic protease activating factor 1 and initiates the caspase
cascade to induce apoptosis (32).
In the current study, Jaceosidin treatment downregulated Bcl-2 and
upregulated Bax expression in A549 cells in a dose- and
time-dependent manner, thereby decreasing the Bcl-2/Bax ratio, and
enhanced the release of cyto-c from mitochondria into the
cytoplasm, thus leading to caspase cleavage. Therefore, it was
hypothesized that Jaceosidin could induce apoptosis in A549 cells
via a mitochondria-dependent pathway. These results were consistent
with those reported by Kim et al (11), which demonstrated that Jaceosidin
could induce apoptosis in Ras-transformed human mammary epithelial
cells via the production of ROS, and those of Lv et al
(33), which revealed that
Jaceosidin could induce apoptosis in human ovarian cancer cells via
the mitochondrial pathway. Although cytosolic cyto-c was
significantly increased, the mitochondrial fraction was not
assessed in the present study. Further experiments including
mitochondrial separation and immunofluorescence staining for cyto-c
will be performed to verify cyto-c release.
The MAPK (Ras/Raf/MEK/ERK) signaling pathway has a
key regulatory role in the pathogenesis of NSCLC. The Ras gene
encodes a family of proteins involved in the regulation of cell
proliferation, differentiation and apoptosis (34). ERK is a downstream effector
activated by several growth factors, including epidermal growth
factor, nerve growth factor and platelet-derived growth factor.
Activated ERK1/2 mediates and amplifies tumor-related signals
during invasion and metastasis, and its phosphorylation triggers
the activation of downstream substrates, thus regulating cell
proliferation and apoptosis (35).
In parallel, the Akt pathway has been shown to be associated with
cell survival and proliferation, and its activation has been
involved in malignant growth and apoptosis escape in several types
of cancer (36,37). The findings of the present study
were consistent with those reported by previous studies, which
showed that Jaceosidin can decrease p-ERK and p-Akt to induce
apoptosis in Ras-transformed human mammary epithelial cells
(11), and that Akt signaling is
inhibited during Jaceosidin-mediated apoptosis in oral cancer cells
(30). Collectively, these results
suggested that Jaceosidin could inhibit A549 cell viability and
promote apoptosis via inhibition of the Ras/Raf/MEK/ERK and Akt
pathways. However, more extensive studies are needed to reveal the
detailed molecular mechanisms underlying Jaceosidin-mediated
pathway blockage. Additionally, Jaceosidin exhibited significant
tumor-selective cytotoxicity in the present study, with an
IC50 value of 55 µM in A549 cells and 248.5 µM in normal
293T cells (a 4.52-fold difference), which provides a solid basis
for its in vivo application and dose optimization (8). The 120 µM concentration used in the
current study is an in vitro concentration ~2.2-fold higher
than the IC50 value determined (55 µM), which was used
to clearly demonstrate the dose-dependent apoptotic effects and
signaling pathway changes, a common experimental design for initial
mechanistic research of natural products (17). Although the 120 µM concentration is
higher than the IC50, the significant tumor selectivity
indicated that Jaceosidin has a wide therapeutic window. For in
vivo application, the main challenge is the low bioavailability
of flavonoids; future strategies, such as structural modification,
nanocarrier delivery and combination therapy, may be used to
improve the in vivo bioavailability of Jaceosidin and to
reduce the effective therapeutic concentration (16). In addition, the inhibition of
Ras/Raf/MEK/ERK and Akt pathways by Jaceosidin is particularly
valuable for NSCLC, as these two pathways are frequently
dysregulated in NSCLC and are key targets for clinical anti-NSCLC
therapy (36).
It should be acknowledged that the present study has
several limitations that need to be addressed in future research.
First, the study was limited to the A549 lung adenocarcinoma cell
line, a classical and well-characterized NSCLC cell line for
mechanistic research; validation in other NSCLC cell lines (such as
H1299 and H460 cells) with different genetic backgrounds is
required to confirm the generalizability of the findings. Second,
the mechanistic link between the inhibition of Ras/Raf/MEK/ERK and
Akt pathways and apoptosis/cell cycle arrest is associative in the
present study; future studies using specific pharmacological
inhibitors (such as U0126 for MEK/ERK and LY294002 for PI3K/Akt)
and genetic knockdown approaches will further confirm the causal
relationship. Third, ROS, which has been implicated in
Jaceosidin-induced apoptosis in other cancer models (11), was not explored in the current
study; the role of ROS in Jaceosidin-mediated mitochondrial
apoptosis and pathway inhibition in A549 cells will be investigated
in subsequent work using DCFH-DA staining. Fourth, the present
study only included in vitro experiments; in vivo
xenograft models are needed to verify the anticancer effect of
Jaceosidin and its pharmacokinetic characteristics. Fifth, the
direct or indirect regulatory effect of Jaceosidin on the
Ras/Raf/MEK/ERK and Akt pathways remains unclear; future
biochemical experiments such as molecular docking will be performed
to explore the potential direct binding of Jaceosidin to key
proteins in these pathways (34).
In conclusion, given the high incidence and
mortality rates of NSCLC, identifying effective therapeutic drugs
is of great importance. Herein, the inhibitory effects of
Jaceosidin on A549 NSCLC cells and its underlying mechanism of
action were investigated. The results demonstrated that Jaceosidin
upregulated p21 and induced cell cycle arrest, thus attenuating
A549 cell viability. Furthermore, Jaceosidin promoted the cleavage
of caspases and PARP, triggering apoptosis through both extrinsic
death receptor and intrinsic mitochondrial pathways (Fig. 5). Notably, Jaceosidin may disrupt
mitochondrial stability by downregulating Bcl-2 and upregulating
Bax, thus facilitating cyto-c release from the mitochondria and the
subsequent activation of apoptosis through the intrinsic
mitochondrial pathway. Pathway analysis further indicated that the
anti-survival and pro-apoptotic effects of Jaceosidin were mediated
through inhibition of the Ras/Raf/MEK/ERK and Akt signaling
pathways.
Overall, although additional experiments are
required to fully uncover the regulatory network of Jaceosidin, the
results of the current study suggested that it could represent a
promising and innovative active agent for the treatment of
NSCLC.
Acknowledgements
Not applicable.
Funding
This research was funded by the Project of Yunnan Clinical
Research Center for Geriatric Diseases (grant no. 2023YJZX-LN12;
2022YJZX-LN19; 2023YJZX-LN14; 2023YJZX-LN21; 202102AA310069) and
the Basic Research Program of Yunnan Province (grant no.
202401AT070358).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
XC, CM, XH and LC conceived and designed the study.
Experiments were performed by XC, CM, XW, CW, LS, JW and RY. Data
processing, analysis and interpretation were performed by XC, CM
and XW. The manuscript was written by XC, CM and XW. Funding was
obtained by XH and LC. YL participated in study design, supervised
the study and revised the manuscript. XH and LC supervised the
study and revised the manuscript. XC and CM confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Giaquinto AN and Jemal A:
Cancer statistics, 2024. CA Cancer J Clin. 74:12–49.
2024.PubMed/NCBI
|
|
2
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
3
|
Bade BC and Dela Cruz CS: Lung cancer
2020: Epidemiology, etiology, and prevention. Clin Chest Med.
41:1–24. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hirsch FR, Scagliotti GV, Mulshine JL,
Kwon R, Curran WJ Jr, Wu YL and Paz-Ares L: Lung cancer: Current
therapies and new targeted treatments. Lancet. 389:299–311. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Duma N, Santana-Davila R and Molina JR:
Non-Small cell lung cancer: Epidemiology, screening, diagnosis, and
treatment. Mayo Clin Proc. 94:1623–1640. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Friedlaender A, Addeo A, Russo A, Gregorc
V, Cortinovis D and Rolfo CD: Targeted therapies in early stage
NSCLC: Hype or hope? Int J Mol Sci. 21:63292020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Park E, Hong K, Kwon BM, Kim Y and Kim JH:
Jaceosidin ameliorates insulin resistance and kidney dysfunction by
enhancing insulin receptor signaling and the antioxidant defense
system in type 2 diabetic mice. J Med Food. 23:1083–1092. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nageen B, Rasul A, Hussain G, Shah MA,
Anwar H, Hussain SM, Uddin MS, Sarfraz I, Riaz A and Selamoglu Z:
Jaceosidin: A natural flavone with versatile pharmacological and
biological activities. Curr Pharm Des. 27:456–466. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee HG, Yu KA, Oh WK, Baeg TW, Oh HC, Ahn
JS, Jang WC, Kim JW, Lim JS, Choe YK and Yoon DY: Inhibitory effect
of jaceosidin isolated from Artemisiaargyi on the function of E6
and E7 oncoproteins of HPV 16. J Ethnopharmacol. 98:339–343. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Woo SM and Kwon TK: Jaceosidin induces
apoptosis through Bax activation and down-regulation of Mcl-1 and
c-FLIP expression in human renal carcinoma Caki cells. Chem Biol
Interact. 260:168–175. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim MJ, Kim DH, Lee KW, Yoon DY and Surh
YJ: Jaceosidin induces apoptosis in ras-transformed human breast
epithelial cells through generation of reactive oxygen species. Ann
N Y Acad Sci. 1095:483–495. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jeong MA, Lee KW, Yoon DY and Lee HJ:
Jaceosidin, a pharmacologically active flavone derived from
Artemisia argyi, inhibits phorbol-ester-induced upregulation of
COX-2 and MMP-9 by blocking phosphorylation of ERK-1 and −2 in
cultured human mammary epithelial cells. Ann N Y Acad Sci.
1095:458–466. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Khan M, Yu B, Rasul A, Al Shawi A, Yi F,
Yang H and Ma T: Jaceosidin induces apoptosis in U87 glioblastoma
cells through G2/M phase arrest. Evid Based Complement Alternat
Med. 2012:7030342012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Khan M, Rasul A, Yi F, Zhong L and Ma T:
Jaceosidin induces p53-dependent G2/M phase arrest in U87
glioblastoma cells. Asian Pac J Cancer Prev. 12:3235–3238.
2011.PubMed/NCBI
|
|
15
|
Nagasaka M and Gadgeel SM: Role of
chemotherapy and targeted therapy in early-stage non-small cell
lung cancer. Expert Rev Anticancer Ther. 18:63–70. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang Y, Li N, Wang TM and Di L: Natural
products with activity against lung cancer: A review focusing on
the tumor microenvironment. Int J Mol Sci. 22:108272021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wen T, Song L and Hua S: Perspectives and
controversies regarding the use of natural products for the
treatment of lung cancer. Cancer Med. 10:2396–2422. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Z: Regulation of cell cycle
progression by growth Factor-induced cell signaling. Cells.
10:33272021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan W, Wu THY, Leung SSY and To KKW:
Flavonoids potentiated anticancer activity of cisplatin in
non-small cell lung cancer cells in vitro by inhibiting histone
deacetylases. Life Sci. 258:1182112020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pani S, Mohapatra S, Sahoo A, Baral B and
Debata PR: Shifting of cell cycle arrest from the S-phase to G2/M
phase and downregulation of EGFR expression by phytochemical
combinations in HeLa cervical cancer cells. J Biochem Mol Toxicol.
36:e229472022. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim IH, Eom T, Park JY, Kim HJ and Nam TJ:
Dichloromethane fractions of Calystegia soldanella induce
S-phase arrest and apoptosis in HT-29 human colorectal cancer
cells. Mol Med Rep. 25:602022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li J, Yuan J, Li Y, Wang J, Gong D, Xie Q,
Ma R, Wang J, Ren M, Lu D and Xu Z: d-Borneol enhances cisplatin
sensitivity via p21/p27-mediated S-phase arrest and cell apoptosis
in non-small cell lung cancer cells and a murine xenograft model.
Cell Mol Biol Lett. 27:612022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shamloo B and Usluer S: p21 in cancer
research. Cancers (Basel). 11:11782019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'Orazi G: Apoptosis as anticancer mechanism: Function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang X, Yang X, Chen M, Zheng S, Li J,
Lin S and Wang X: ST3Gal3 confers paclitaxel-mediated
chemoresistance in ovarian cancer cells by attenuating caspase-8/3
signaling. Mol Med Rep. 20:4499–4506. 2019.PubMed/NCBI
|
|
26
|
Ramadhani FJ, Kang SH, Kawala RA, Chung
BY, Bai HW and Kang BS: γ-irradiated prednisolone promotes
apoptosis of liver cancer cells via activation of intrinsic
apoptosis signaling pathway. Mol Med Rep. 23:4252021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jin N, Xia Y and Gao Q: Combined PARP
inhibitors and small molecular inhibitors in solid tumor treatment
(review). Int J Oncol. 62:282023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Santagostino SF, Assenmacher CA, Tarrant
JC, Adedeji AO and Radaelli E: Mechanisms of regulated cell death:
Current perspectives. Vet Pathol. 58:596–623. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim BI, Kim JH, Sim DY, Nam M, Jung JH,
Shim B, Lee J and Kim SH: Inhibition of JAK2/STAT3 and activation
of caspase-9/3 are involved in KYS05090S-induced apoptosis in
ovarian cancer cells. Int J Oncol. 55:203–210. 2019.PubMed/NCBI
|
|
30
|
Han HY, Kim HJ, Jeong SH, Kim J, Jeong SH,
Kim GC, Hwang DS, Kim UK and Ryu MH: The flavonoid jaceosidin from
artemisia princeps induces apoptotic cell death and inhibits the
Akt pathway in oral cancer cells. Evid Based Complement Alternat
Med. 2018:57650472018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Warren CFA, Wong-Brown MW and Bowden NA:
BCL-2 family isoforms in apoptosis and cancer. Cell Death Dis.
10:1772019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang Y, Xia C, Lun Z, Lv Y, Chen W and Li
T: Crosstalk between p38 MAPK and caspase-9 regulates
mitochondria-mediated apoptosis induced by
tetra-α-(4-carboxyphenoxy) phthalocyanine zinc photodynamic therapy
in LoVo cells. Oncol Rep. 39:61–70. 2018.PubMed/NCBI
|
|
33
|
Lv W, Sheng X, Chen T, Xu Q and Xie X:
Jaceosidin induces apoptosis in human ovary cancer cells through
mitochondrial pathway. J Biomed Biotechnol.
2008:3948022008.PubMed/NCBI
|
|
34
|
Korzeniecki C and Priefer R: Targeting
KRAS mutant cancers by preventing signaling transduction in the
MAPK pathway. Eur J Med Chem. 211:1130062021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Barbosa R, Acevedo LA and Marmorstein R:
The MEK/ERK network as a therapeutic target in human cancer. Mol
Cancer Res. 19:361–374. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tan AC: Targeting the PI3K/Akt/mTOR
pathway in non-small cell lung cancer (NSCLC). Thorac Cancer.
11:511–518. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Takeda T, Yamamoto Y, Tsubaki M, Matsuda
T, Kimura A, Shimo N and Nishida S: PI3K/Akt/YAP signaling promotes
migration and invasion of DLD-1 colorectal cancer cells. Oncol
Lett. 23:1062022. View Article : Google Scholar : PubMed/NCBI
|