Introduction
Tongue squamous cell carcinoma (TSCC) is a prevalent
and aggressive malignancy of the head and neck region, associated
with high morbidity and mortality rates, with an estimated global
incidence of 151,000 new cases and 48,000 deaths annually as of
2023, yielding an age-standardized incidence rate of 1.7 per
100,000 and mortality of 0.54 per 100,000 (1). The pathogenesis of TSCC is
multifactorial, driven by a complex interplay of genetic mutations,
cumulative genetic alterations, signaling aberrations, epigenetic
modifications, environmental exposures (such as tobacco use,
alcohol consumption and human papillomavirus infection) and
lifestyle-related risk factors (2).
In recent years, the concept of cancer stem cells
(CSCs) has gained considerable attention as a key driver of tumor
initiation, progression and therapeutic resistance. CSCs are a
subpopulation of tumor cells with the capacity for self-renewal and
differentiation into diverse cell types, thereby sustaining tumor
growth and heterogeneity (3). TSCC
CSCs exploit mitochondrial dynamics to reprogram lipid metabolism,
promoting chemoresistance and survival under metabolic stress,
while HPV+ TSCC subtypes exhibit distinct CSC properties that may
be targeted for relapse-free therapy (4). These cells are sustained by specific
molecular signaling pathways that are highly relevant in TSCC
(5).
The present review aimed to elucidate the
characteristics of CSCs in TSCC and the molecular signaling
mechanisms that govern their function, with the ultimate aim of
providing novel insights for future therapeutic strategies. A
deeper understanding of the unique biology of CSCs in TSCC may
facilitate the identification of new therapeutic targets and
support the development of more effective treatments for this
challenging disease.
Biological characteristics and significance
of CSCs
CSCs are a subpopulation of tumor cells that possess
the capacities for self-renewal and differentiation, which are
essential for tumor initiation, maintenance and recurrence. They
are often characterized by their ability to initiate tumor
formation when transplanted into immunocompromised mice, a
technique known as the xenograft assay (6,7).
Additionally, CSCs can be identified by the expression of specific
surface markers, such as CD44, CD24 and aldehyde dehydrogenase 1
(ALDH1), across various types of cancer, including TSCC (8). CD133 is also a recognized marker of
CSCs across multiple malignancies and is often co-expressed with
other stem cell markers (9); in
ER-positive/HER2-negative breast cancer, high expression of CD133
is associated with a better chemotherapy response and survival rate
(10).
In TSCC, CD133+ cells exhibit stem
cell-like properties and enhanced tumorigenic potential (11). Furthermore, a high density of α
smooth muscle actin (SMA)+ cancer-associated fibroblasts
has been related to disease recurrence and poor survival, whereas
the CD133+αSMA+ phenotype appears to be
mainly associated with vascular structures (12,13).
Furthermore, the CD44+CD133+ cell
subpopulation is regarded as a potential metastatic precursor,
demonstrating increased proliferation, clonogenicity, invasion and
migratory capacity (14).
Other approaches for identifying CSCs include
functional assays, such as the sphere-forming assay, which
evaluates the ability of cells to generate three-dimensional
aggregates under non-adherent conditions, a feature indicative of
stem-like properties (15). DNA
methylation profiling has also been used to distinguish CSCs from
non-CSCs. For example, A study has shown that, compared with
non-CSCs, the promoter regions of specific tumor suppressor genes
in CSCs are often hypermethylated, enabling clear differentiation
between these cell populations. Furthermore, genetic and epigenetic
profiling can be employed to distinguish CSCs from non-CSCs,
providing deeper insight into the molecular mechanisms underlying
their distinct characteristics (16).
Characteristics of TSCC-CSCs
In TSCC, CSCs exhibit several distinct biological
characteristics that contribute to tumor progression and
therapeutic resistance (6). A key
feature of CSCs is their high capacity for self-renewal, which
enables them to sustain the tumor cell population and drive
continued growth (17).
Additionally, TSCC-CSCs often display enhanced resistance to
chemotherapy and radiotherapy, attributed to their ability to
activate DNA repair pathways and evade apoptosis (18). These cells also exhibit a high
degree of plasticity, with the ability to undergo
epithelial-mesenchymal transition (EMT) and mesenchymal-epithelial
transition, thereby facilitating migration and invasion into
surrounding tissues (19,20).
CSCs and EMT are considered to form a bidirectional
reinforcing loop that promotes tumor progression (21). In one respect, the EMT process,
activated by pathways such as TGF-β and Wnt, induces epithelial
cells to lose polarity and acquire migratory and invasive
capabilities through the action of key transcription factors
(22). Concurrently, EMT confers
stem-like properties by reprogramming differentiated tumor cells
into CSCs, upregulating stem cell markers such as OCT4 and SOX2,
and enhancing self-renewal capacity. Notably, CSCs promote EMT by
secreting cytokines, including TGF-β and IL-6, thereby establishing
a self-perpetuating cycle within the tumor microenvironment that
drives surrounding cells toward a mesenchymal phenotype (23). Furthermore, TSCC-CSCs secrete
various cytokines and growth factors that contribute to a
supportive tumor microenvironment, promoting angiogenesis and
immune evasion (24).
Microenvironment and epigenetic
regulatory network
Hypoxia stabilizes HIF-1α, which activates
stemness-associated genes and promotes metabolic reprogramming in
CSCs, thereby enhancing self-renewal and chemoresistance. In TSCC,
hypoxic niches also upregulate ATP-binding cassette (ABC)
transporters, contributing to increased drug efflux (25). Extracellular matrix (ECM) components
such as collagen, laminin and fibronectin interact with integrin
receptors on CSCs to activate pro-survival signaling pathways
(26). These interactions trigger
focal adhesion kinase (FAK)/Src, PI3K/Akt and Ras/MAPK cascades,
enhancing CSC self-renewal, inhibiting apoptosis and promoting EMT,
thereby facilitating invasion and metastasis (27).
In TSCC, a stiffened ECM driven by the activation
and transdifferentiation of cancer-associated fibroblasts (CAFs),
which induces mechanotransduction via YAP/TAZ signaling, reinforces
CSC plasticity and EMT (28).
ECM-bound growth factors (TGF-β, EGF, HGF) maintain CSC quiescence
and niche retention in TSCC via three mechanisms: Sequestration by
ECM components, integrin-receptor crosstalk via FAK/Src and
PI3K/Akt, and MMP-mediated activation (29).
In addition, epigenetic dysregulation, including DNA
hypermethylation of tumor suppressor genes and histone
modifications, suppresses differentiation-related genes while
activating pathways such as Wnt/β-catenin and Notch (30).
Hepatocyte growth factor and its receptor c-MET
serve important roles in tongue development and the carcinogenesis
of head and neck squamous cell carcinoma (HNSCC) (31). Knockdown of c-MET reduces the
sphere-forming ability and stem cell marker expression of HNSCC
stem-like cells, while increasing sensitivity to cisplatin by
decreasing the side population fraction and downregulating the
ABCG2 transporter gene (31).
Non-coding RNAs also contribute to the derepression of
CSC-associated oncogenes. These ncRNAs form a regulatory triad that
destabilizes tumor-suppressive checkpoints, enabling CSCs to
maintain quiescence, evade therapy and initiate recurrence
(32). The H19/miR-let-7/HMGA2 and
miR-21/PTEN/STAT3 axes are particularly robust in TSCC, supported
by clinical tissue analyses and TCGA-derived co-expression networks
(33). In addition, hypoxia and ECM
stiffness synergistically remodel the epigenetic landscape,
maintaining CSCs in a stem-like state, with key therapeutic targets
including HIF-1α inhibitors (e.g., PX-478 in Phase II trials) and
YAP/TAZ pathway modulators (e.g., Verteporfin, reducing CSCs by 60%
in preclinical models) (34,35).
Collectively, these mechanisms support CSC resilience and present
potential targets for combination therapies in TSCC (36). The clinical significance of CSCs in
TSCC is substantial, as these cells are implicated in several key
aspects of cancer biology that directly affect patient outcomes
(37). CSCs serve a key role in
tumor recurrence and metastasis (38); owing to their resistance to
conventional therapies, CSCs can survive treatment and subsequently
drive tumor regrowth, contributing to poor prognosis (37). Additionally, the presence of CSCs in
TSCC is associated with an increased likelihood of metastasis, as
these cells possess the capacity to migrate and colonize distant
sites (39). Therefore, the
identification and targeting of CSCs in TSCC represents a promising
strategy for improving therapeutic efficacy and patient survival
(40). Furthermore, a deeper
understanding of the molecular and cellular mechanisms underlying
CSCs in TSCC may facilitate the development of novel targeted
therapies capable of overcoming current treatment limitations
(41,42).
Roles of the Wnt/β-catenin signaling pathway
in TSCC
The Wnt/β-catenin signaling pathway is a key
regulator of cell proliferation, differentiation and apoptosis
(43). In the absence of Wnt
ligands, β-catenin is phosphorylated by a destruction complex
composed of proteins such as axis inhibition protein, adenomatous
polyposis coli (APC) and glycogen synthase kinase-3b, which targets
it for proteasomal degradation (44). However, when Wnt ligands bind to
Frizzled receptors and low-density lipoprotein-related proteins 5
and 6 co-receptors, this phosphorylation is inhibited, allowing
β-catenin to accumulate in the cytoplasm and subsequently
translocate into the nucleus (45).
In the nucleus, β-catenin interacts with T cell factor
(TCF)/lymphoid enhancer factor family (LEF) transcription factors
to activate the expression of target genes involved in cell
proliferation and survival (Fig. 1)
(46,47). In healthy cells, this destruction
complex tightly regulates cytoplasmic β-catenin levels. Aberrant
activation of the pathway can occur through mutations in CTNNB1
(encoding β-catenin) or APC, epigenetic silencing of pathway
inhibitors such as secreted frizzled-related proteins and Dickkopf
proteins, or autocrine/paracrine upregulation of Wnt ligands
(48). This hyperactivation leads
to β-catenin accumulation and its translocation into the nucleus.
Within the nucleus, β-catenin forms complexes with TCF/LEF
transcription factors, driving the expression of oncogenic target
genes (49). These genes regulate
key cancer hallmarks in TSCC, including uncontrolled proliferation
(via c-Myc and Cyclin D1), evasion of apoptosis, EMT (which in turn
facilitates invasion and metastasis), maintenance of CSCs and
angiogenesis (50). The
Wnt/β-catenin signaling pathway molecular mechanism is highly
conserved across species and serves a critical role in both normal
developmental processes and disease pathogenesis, including TSCC
(51).
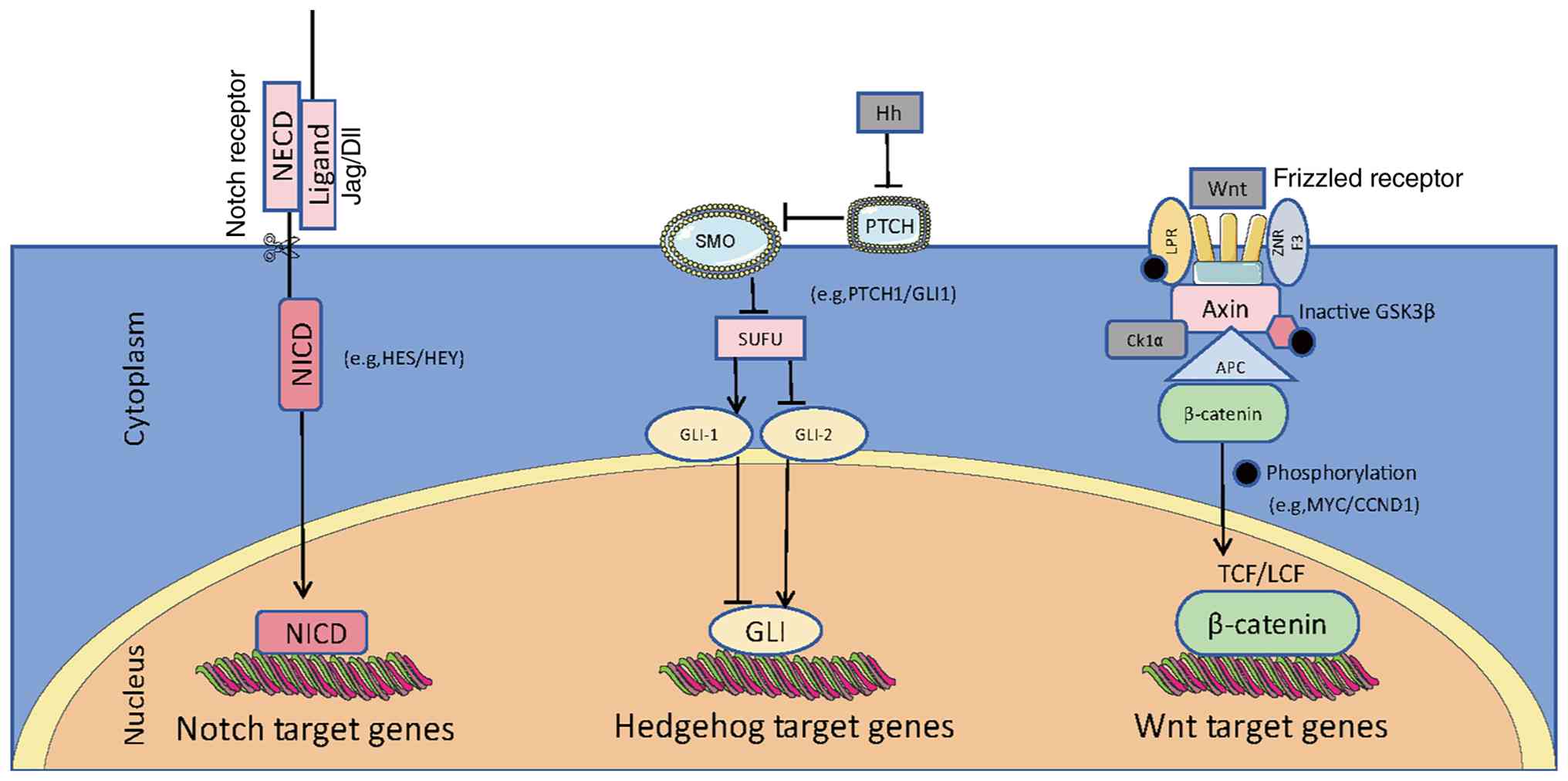 | Figure 1.Simplified view of canonical Notch,
Hh and Wnt signaling pathways in cancer. The Notch pathway involves
ligand binding to receptors, releasing NICD into the nucleus to
activate target genes like HES/HEY. The Hedgehog pathway activates
GLI transcription factors via SMO. The Wnt pathway stabilizes
β-catenin for nuclear translocation to regulate genes. Wnt ligands
bind to Frizzled receptors, preventing β-catenin degradation and
thus controlling cell proliferation and differentiation. NICD,
Notch intracellular domain; NECD, Notch extracellular domain;
Jag/DII, jagged/delta-like ligands; SMO, smoothened; PTCH, patched;
Hh, Hedgehog; GLI, glioma-associated oncogene homolog; LPR,
low-density lipoprotein receptor-related protein; Axin, axis
inhibition protein; SUFU, suppressor of fused homolog; ZNRF3, zinc
and ring finger 3; APC, adenomatous polyposis coli; TCF/LEF, T-cell
factor/lymphoid enhancer factor; GSK3b, glycogen synthase
kinase-3b; Ck1α, casein kinase 1α; HES, hairy and
enhancer-of-split; HEY, HES related with YRPW motif; MYC,
myelocytomatosis oncogene; CCND1, cyclin D1. |
The Wnt/β-catenin signaling pathway has been
implicated in the maintenance and expansion of CSCs in TSCC
(52). Accumulating evidence
suggests that activation of this pathway promotes the expression of
stemness-associated genes, such as SOX2 and OCT4, which are
essential for maintaining CSC properties (53,54).
Therefore, targeting the Wnt/β-catenin signaling pathway in TSCC
may represent a promising strategy for eliminating CSCs and
improving patient outcomes (55).
Roles of the Hedgehog (Hh) signaling pathway
in TSCC
The Hh signaling pathway is a key regulator of cell
growth and differentiation during embryonic development and in
adult tissues (56). The pathway is
initiated when Hh ligands bind to the Patched (Ptch) receptor,
relieving its inhibition of the Smoothened (Smo) protein (57). Activation of Smo promotes the
translocation of glioma-associated oncogene homolog (GLI)
transcription factors from the cytoplasm to the nucleus, where they
drive the transcription of Hh target genes (58,59).
The pathway is tightly regulated by several proteins, including
Suppressor of Fused, which negatively regulates Gli activity, and
Costal-2 (Cos2), which helps stabilize GLI proteins in the
cytoplasm (Fig. 1) (60,61).
Dysregulation of Hh signaling has been implicated in various types
of cancer, including TSCC, where it contributes to tumor
progression and maintenance (62).
In TSCC, the Hh signaling pathway is frequently
aberrantly activated, contributing to disease pathogenesis and
progression (63). This activation
may occur through multiple mechanisms, including upregulation of Hh
ligands, mutations in the Ptch receptor or alterations in other
regulatory components of the pathway (64). For example, mutations in the Ptch
gene can result in constitutive pathway activation even in the
absence of ligand binding (65,66).
In addition, the expression levels of Hh target genes, such as GLI1
and GLI2, are often upregulated in TSCC, indicating sustained
pathway activity (67,68). This persistent activation promotes
cell proliferation, inhibits apoptosis and enhances cancer cell
survival, thereby contributing to the aggressive behavior of TSCC
(69).
The Hh signaling pathway has been shown to serve a
critical role in the maintenance and expansion of CSCs in TSCC
(70), by upregulating stem cell
markers and genes involved in cell cycle regulation (71). For example, the transcription factor
GLI1, a key mediator of Hh signaling, enhances the expression of
Nanog and OCT4, which are essential for maintaining CSC stemness
(72,73). Furthermore, Hh signaling also
increases the expression of ABC transporters, which contributes to
CSC drug resistance (74). Thus,
targeting the Hh pathway may represent a promising therapeutic
strategy for eliminating CSCs and improving the prognosis of
patients with TSCC (75).
Roles of the Notch signaling pathway in
TSCC
Notch signaling mediates short-range cell-to-cell
communication through interactions between ligands and receptors on
adjacent cells. When a Notch ligand binds to its receptor, it
triggers cleavage at the S2 site by ADAM10 and ADAM17, resulting in
shedding of the extracellular domain (76). This is followed by
γ-secretase-mediated cleavage at the S3 site within the
transmembrane region. After S3 cleavage, the Notch intracellular
domain (NICD) is released from the plasma membrane and translocates
to the nucleus (77). In the
nucleus, NICD interacts with recombination signal binding protein
for immunoglobulin κ J region, also known as CBF1/Suppressor of
Hairless/Lag-1 (CSL), converting the transcriptional repressor
complex into a transcriptional activator complex and thereby
promoting the expression of Notch target genes (78).
The Notch signaling pathway is a highly conserved
intercellular communication system that plays a critical role in
cell fate determination, differentiation, and proliferation
(79). It involves a family of
transmembrane receptors (Notch1-4) that are activated upon binding
to ligands such as Jagged and Delta-like proteins (80). Following ligand engagement, the
Notch receptor undergoes two successive proteolytic cleavages,
resulting in the release of the NICD, which translocates to the
nucleus (81). In the nucleus, NICD
interacts with the DNA-binding protein CSL, displacing
co-repressors and recruiting co-activators to initiate
transcription of target genes (82). This tightly regulated mechanism
ensures precise control of multiple cellular processes, making
Notch signaling essential for tissue homeostasis and developmental
regulation (83).
In TSCC, Notch signaling is frequently upregulated
and serves a significant role in tumor progression and maintenance
(84). Activation of the pathway is
often driven by upregulation of ligands such as Jagged1 and
delta-like ligand 1, which promote tumor cell proliferation and
survival (Fig. 1). In addition,
mutations or amplifications in Notch receptors can lead to
constitutive pathway activation, further contributing to oncogenic
transformation (85). In HNSCC
(including TSCC), inactivation mutations of NOTCH1 are more common,
but amplification (such as an increase in copy number) of NOTCH3
can lead to excessive activation of the pathway, promoting tumor
invasion (86).
The regulatory mechanisms of Notch signaling in TSCC
are complex and involve both positive and negative feedback loops,
as well as crosstalk with other pathways. The Wnt signal
upregulates the expression of the Jagged1 ligand, further
activating Notch, and Akt directly phosphorylates Notch1, enhancing
its transcriptional activity (87).
Notch can activate NF-κB, maintaining the inflammatory
microenvironment by upregulating IL-6 and IL-8, enhancing the
invasiveness of tumors; furthermore, the intracellular segment of
Notch1 (NICD) can directly bind to YAP/TAZ, inhibiting the tumor
suppressive effect of the Hippo pathway and promoting the
characteristics of tumor stem cells (88,89).
These interactions create a dynamic regulatory network that can
either enhance or suppress Notch activity, depending on the tumor
microenvironment and specific genetic context (84).
Notch signaling has been implicated in the
maintenance and expansion of CSCs in TSCC (90). It promotes CSC properties by
activating transcriptional programs that enhance stemness,
including the expression of OCT4, SOX2 and NANOG (91). In addition, Notch signaling can
suppress differentiation pathways, thereby preserving the
undifferentiated state of CSCs (17). The role of Notch in CSCs extends
beyond transcriptional regulation; it also influences cell cycle
progression, apoptosis and EMT, all of which are critical for CSC
survival and metastatic potential (92,93).
Therefore, targeting Notch signaling represents a potential
therapeutic strategy to deplete CSCs and potentially improve
outcomes in TSCC (54).
Interactions among multiple signaling
pathways and their significance in TSCC
CSCs dynamically interact with key signaling
pathways to drive tumorigenesis and therapeutic resistance. In
TSCC, activation of the Wnt/β-catenin pathway stabilizes nuclear
β-catenin in CD44+ CSCs, leading to the upregulation of
c-MYC and OCT4 and sustaining self-renewal and chemoresistance
(94). Notch signaling, through
NICD cleavage, induces EMT by promoting HES1 expression, thereby
facilitating metastasis (95). Hh
pathway activation enhances CSC proliferation and contributes to
post-therapy recurrence (96).
Concurrently, PI3K/Akt/mTOR pathway alterations reprogram CSC
metabolism under hypoxic conditions, while NF-κB activation driven
by chronic inflammation suppresses apoptotic signaling. These
pathways engage in extensive crosstalk; for example, tumor
microenvironment-derived TGF-β amplifies Wnt and Notch signaling,
thereby locking CSCs into a drug-tolerant state (97). Targeting these signaling hubs can
disrupt CSC-driven progression, as demonstrated in TSCC xenograft
models where Wnt inhibition reduced tumor-initiating capacity by
~70% (98). Collectively, these
findings underscore the therapeutic potential of pathway-specific
targeting for CSC depletion in TSCC.
The interaction between multiple signaling pathways
is a complex yet critical aspect of cellular regulation,
particularly in TSCC (99). These
pathways often converge and crosstalk to regulate key cellular
processes such as proliferation, survival and differentiation
(100). For example, the PI3K/Akt
pathway, which is frequently activated in cancer, can interact with
the MAPK/ERK pathway to enhance cell survival and proliferation
(100,101). This interaction may occur through
shared downstream effectors or crosstalk at the level of upstream
receptors (102). In addition, the
NF-κB pathway, which is well known for its role in inflammation and
stress responses, integrates signals from multiple pathways,
including those activated by growth factors and cytokines, to
regulate gene expression and cellular fate (103). Understanding these mechanisms is
essential for the development of targeted therapies aimed at
disrupting these interactions and inhibiting cancer
progression.
The synergistic effects of multiple signaling
pathways in TSCC are profound and multifaceted. When concurrently
activated, these pathways can exert a stronger influence on cancer
cell behavior than when activated individually (101). For example, co-activation of the
PI3K/Akt and MAPK/ERK pathways enhances cell proliferation and
resistance to apoptosis, contributing to the aggressive nature of
TSCC (104,105); interactions between these pathways
can also promote EMT, a process closely associated with metastasis
(106). Additionally, EMT is
driven by the coordinated activity of pathways such as TGF-β and
Wnt/β-catenin, which may be further amplified through crosstalk
with additional signaling networks (Fig. 2) (107,108). In TSCC and HNSCC, the
co-activation of the PI3K/AKT and MAPK/ERK pathways significantly
enhances tumor proliferation, anti-apoptosis and the EMT process
(109). This synergistic
activation not only drives malignant transformation but also
contributes to therapeutic resistance, making TSCC particularly
challenging to treat (110).
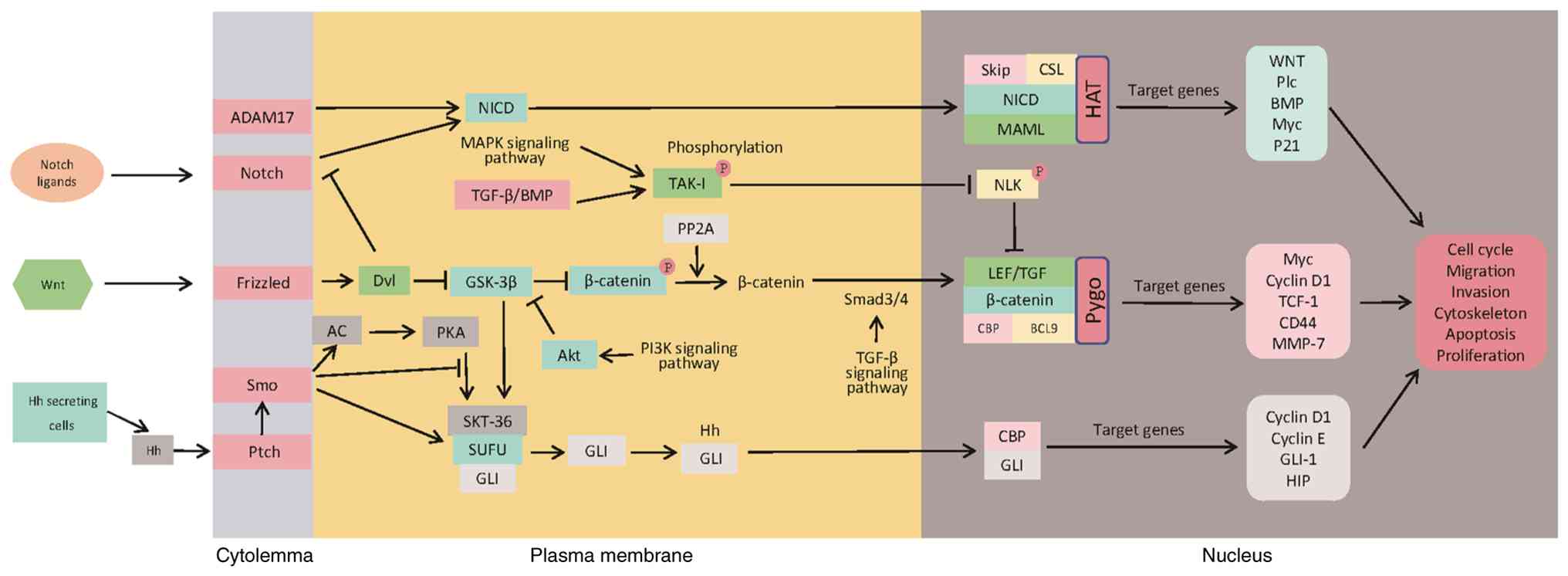 | Figure 2.Crosstalk between Notch, Wnt and Hh
signaling pathways in cancer. They coordinate cellular functions
through receptor-ligand binding, intracellular signal cascades,
nuclear translocation of effector proteins to regulate gene
expression, and feedback loops. ADAM17, a disintegrin and
metalloproteinase 17; SMO, smoothened; Ptch, patched; NICD, Notch
intracellular domain; BMP, bone morphogenetic protein; Dvl,
dishevelled; AC, adenylate cyclase; PKA, protein kinase A; SUFU,
suppressor of fused homolog; GLI, glioma-associated oncogene
homolog; Hh, hedgehog; TAK1, transforming growth factor-β-activated
kinase 1; PP2A, protein phosphatase 2A; SKIP, SKI-interacting
protein; CSL, CBF1/Su(H)/Lag-1; CBP, CREB-binding protein; BCL9,
B-cell lymphoma 9; LEF, lymphoid enhancer factor; HAT, histone
acetyltransferase; TCF, T-cell factor; HIP, Hh-interacting protein;
MAPK, mitogen-activated protein kinase; GSK-3, glycogen synthase
kinase-3; PI3K, phosphoinositide 3-kinase; Akt, protein kinase B;
Smad, mothers against decapentaplegic homolog; TGF, transforming
growth factor; MAML, mastermind-like; Pygo, pygopus; NLK, nemo-like
kinase. |
The interactions between multiple signaling pathways
also have a significant impact on CSCs, which are considered key
drivers of tumor initiation, progression and recurrence in TSCC
(3). CSCs exhibit a distinct
signaling landscape compared with non-stem cancer cells (17). For example, the Notch and
Wnt/β-catenin pathways are frequently upregulated in CSCs and serve
critical roles in maintaining stemness and self-renewal capacity
(93). These pathways interact with
additional signaling networks, such as PI3K/Akt and MAPK/ERK, to
establish a robust signaling environment that supports CSC survival
and proliferation (111).
Disrupting these interactions could potentially reduce the CSC
population and thereby inhibit tumor growth and metastasis
(112). Because CSCs drive tumor
initiation, growth, therapy resistance, recurrence and metastasis,
reducing their abundance may suppress primary tumor progression and
the formation of secondary metastatic lesions (113,114), thus targeting a fundamental source
of malignancy and relapse. Therefore, utilizing the crosstalk
between these signaling pathways to develop therapeutic targets
against TSCC may represent a promising strategy in the future
(115).
Therapeutic challenges of CSCs in TSCC
CSCs in TSCC exhibit marked resistance to
conventional chemotherapy and radiotherapy, thus posing significant
challenges to effective treatment, while also offering
opportunities for the development of novel treatment strategies
(116). A study has shown that
TSCC-CSCs express specific biomarkers, including ALDH, CD44, NANOG,
OCT4 and BMI1. These markers not only facilitate the identification
of CSCs, but may also serve as potential therapeutic targets
(117).
Complexity of molecular
heterogeneity
CSCs in TSCC exhibit significant molecular
heterogeneity. CSCs derived from different patients, and even
within the same tumor, may display distinct molecular profiles and
patterns of signaling pathway activation (99). This heterogeneity complicates the
development of a unified targeted therapeutic strategy against
CSCs. For example, upregulated of the ZFX gene has been associated
with tumor progression in certain TSCC cases; however, its
expression levels and functional roles vary considerably among
patients (118).
Redundancy and compensatory mechanisms
of signaling pathways
CSCs typically rely on multiple signaling pathways
to maintain their stemness and viability, including the
Wnt/β-catenin, Notch, Hh and HIF-1α/MCT4 pathways (119). When one pathway is inhibited,
others may become activated to compensate for the loss of function,
thereby contributing to treatment resistance. For example, in colon
cancer, after inhibiting the Wnt pathway, the Hh pathway will in a
compensatory manner activate, maintaining the CSC characteristics
by upregulating GLI1 (120). In
glioblastoma, blocking the Notch signal will lead to an increase in
HIF-1α expression, and through glycolytic reprogramming, it will
support the survival of CSCs (121). However, this multi-target
mechanism may encounter complex regulatory challenges in clinical
applications (122). For instance,
in pancreatic cancer, when both the Wnt and Hh pathways are
simultaneously inhibited, NF-κB will undergo compensatory
activation, which instead accelerates metastasis (123).
Rapid evolution of treatment
resistance
CSCs exhibit a high degree of genomic instability
and adaptability, enabling them to rapidly develop resistance
mechanisms under therapeutic pressure (124). For example, during chemotherapy,
CSCs may evade treatment by upregulating drug efflux pumps such as
ABC transporters, enhancing DNA repair capacity or reprogramming
cellular metabolic pathways (125). CSCs evade therapy through three
interconnected mechanisms: Overexpression of ABC transporters
(e.g., ABCB1/P-gp and ABCG2) actively expels chemotherapeutics like
paclitaxel and doxorubicin, reducing intracellular drug
concentrations; upregulation of DNA repair proteins (RAD51, ERCC1,
PARP1) enables efficient repair of radiation- and alkylator-induced
DNA double-strand breaks, suppressing apoptosis; and dynamic
metabolic reprogramming-shifting between glycolysis and oxidative
phosphorylation while upregulating MCT4 and GLUT1-maintains low ROS
levels and adapts to hypoxic, acidic microenvironments (126).
Barriers to clinical translation
Translating laboratory findings on CSCs into
clinical practice presents several challenges. The proportion of
CSCs within tumors is typically very low (for example,
ALDH+ cells account for only 1.3% of the Tca8113 cell
line), requiring highly specific therapeutic approaches to
effectively target these cells (127). CSCs impede clinical translation
because of their hypoxic microenvironment, drug resistance and
heterogeneity; low abundance and missing biomarkers hinder trials,
and current models fail to mimic human biology; they avoid immunity
via PD-L1 and Tregs, sustain stemness via methionine metabolism and
lack validated biomarkers without consensus on endpoints (128,129). Additionally, there is a lack of
reliable in vivo models that accurately recapitulate the
biological behavior and treatment responses of CSCs in TSCC.
Conclusion
The present review of CSC characteristics in TSCC
and key signaling pathways such as Wnt/β-catenin, Notch and Hh
aimed to advance the understanding of TSCC pathogenesis (130). These findings highlight the
critical role of CSCs in TSCC initiation and progression, providing
a theoretical basis for the development of targeted therapies
against CSCs and their regulatory networks (8). A balanced interpretation of existing
research is essential; while a study emphasized the role of
Wnt/β-catenin in CSC maintenance, others highlight the synergistic
effects of Notch and Hh signaling (108). CSCs resist therapy by leveraging
Wnt/β-catenin for self-renewal, while Notch and Hh signaling
synergistically enhance survival and immune evasion (131). In pancreatic cancer models where
dual inhibition of Notch and Hh reduced CSC frequency by 70%
compared to single-pathway targeting (132).
It could be considered that simultaneous targeting
of multiple pathways may be more effective in eliminating CSCs and
preventing recurrence (133).
Integrating molecular insights with clinical data is crucial for
translating these findings into practical therapies (134). Future research should further
elucidate pathway mechanisms and interactions to support the
development of personalized medicine approaches (135), potentially leading to more
effective, less toxic treatments for TSCC and improved patient
outcomes (75). Overall, the
exploration of CSCs in TSCC holds promise for transforming current
cancer treatment strategies (136). Bridging laboratory findings with
clinical application will be essential for the development of
targeted therapies that improve prognosis and quality of life for
patients (137).
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Authors' contributions
XZ and WC designed the present study. XG, WR and QZ
prepared the first draft. WR prepared the figures. CM and XG
reviewed and edited the manuscript. Data authentication is not
applicable. All authors were involved in revising the paper and had
full access to the data. All authors read and approved the final
version of the manuscript.
Availability of data and materials
Not applicable.
Ethical approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Hh
|
Hedgehog
|
|
GLI
|
glioma-associated oncogene homolog
|
|
Ptch
|
patched
|
|
SMO
|
smoothened
|
|
APC
|
adenomatous polyposis coli
|
|
ADAMs
|
A disintegrin and metalloproteases
|
|
Notch
|
neurogenic locus notch homolog
protein
|
|
NICD
|
Notch intracellular domain
|
|
Wnt
|
wingless-type MMTV integration
site
|
|
Myc
|
MYC proto-oncogene
|
|
TCF
|
T cell factor
|
|
CD44
|
cluster of differentiation 44
|
References
|
1
|
Burus T, Damgacioglu H, Huang B, Christian
WJ, Hull PC, Ellis AR, Arnold SM, Deshmukh AA and Lang Kuhs KA:
Trends in oral tongue cancer incidence in the US. JAMA Otolaryngol
Head Neck Surg. 150:436–443. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mohideen K, Krithika C, Jeddy N, Bharathi
R, Thayumanavan B and Sankari SL: Meta-analysis on risk factors of
squamous cell carcinoma of the tongue in young adults. J Oral
Maxillofac Pathol. 23:450–457. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Walcher L, Kistenmacher AK, Suo H, Kitte
R, Dluczek S, Strauß A, Blaudszun AR, Yevsa T, Fricke S and
Kossatz-Boehlert U: Cancer stem cells-origins and biomarkers:
Perspectives for targeted personalized therapies. Front Immunol.
11:12802020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu F, Chen S, Ren S, Wang R, Tan Y, Chen
R, Li B, Cao H and Li J: Regulating lipid metabolism via
mitochondrial dynamics in tongue squamous cell carcinoma cancer
stem cells. Recent Pat Anticancer Drug Discov. 20:445–459. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hoseinian SN, Saeedi M, Saravani ME,
Zenoozi S, Mehranfar F and Pouyan A: Navigating the molecular
signaling: Deciphering cancer stem cell self-renewal pathways. Int
J Mol Cell Med. 14:735–776. 2025.PubMed/NCBI
|
|
6
|
Najafi M, Mortezaee K and Majidpoor J:
Cancer stem cell (CSC) resistance drivers. Life Sci.
234:1167812019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hoque S, Dhar R, Kar R, Mukherjee S,
Mukherjee D, Mukerjee N, Nag S, Tomar N and Mallik S: Cancer stem
cells (CSCs): Key player of radiotherapy resistance and its
clinical significance. Biomarkers. 28:139–151. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gholamzad A, Khakpour N, Khosroshahi EM,
Asadi S, Koohpar ZK, Matinahmadi A, Jebali A, Rashidi M, Hashemi M,
Sadi FH and Gholamzad M: Cancer stem cells: The important role of
CD markers, signaling pathways, and MicroRNAs. Pathol Res Pract.
256:1552272024. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang K, Zhou XK, Wu M, Kang FW, Wang ZL
and Zhu Y: Role of CD133+ cells in tongue squamous
carcinomas: Characteristics of ‘stemness’ in vivo and in vitro.
Oncol Lett. 12:863–870. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sato T, Oshi M, Huang JL, Chida K, Roy AM,
Endo I and Takabe K: CD133 expression is associated with less DNA
repair, better response to chemotherapy and survival in
ER-positive/HER2-negative breast cancer. Res Sq [Preprint].
rs.3.rs-4148608. 2024.PubMed/NCBI
|
|
11
|
Kang FW, Wang K, Wu M, Wang ZL, Zhu Y and
Min R: Biological characteristics of CD133+ subpopulation in tongue
squamous cell carcinoma Tca8113 cell line. Hua Xi Kou Qiang Yi Xue
Za Zhi. 28:560–564. 2010.(In Chinese). PubMed/NCBI
|
|
12
|
Mascolo M, Ilardi G, Romano MF, Celetti A,
Siano M, Romano S, Luise C, Merolla F, Rocco A, Vecchione ML, et
al: Overexpression of chromatin assembly factor-1 p60,
poly(ADP-ribose) polymerase 1 and nestin predicts metastasizing
behaviour of oral cancer. Histopathology. 61:1089–1105. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vered M, Shnaiderman-Shapiro A,
Zlotogorski-Hurvitz A, Salo T and Yahalom R: Cancer-associated
fibroblasts in the tumor microenvironment of tongue carcinoma is a
heterogeneous cell population. Acta Histochem. 121:1514462019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun Y, Han J, Lu Y, Yang X and Fan M:
Biological characteristics of a cell subpopulation in tongue
squamous cell carcinoma. Oral Dis. 18:169–177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Biserova K, Jakovlevs A, Uljanovs R and
Strumfa I: Cancer stem cells: Significance in origin, pathogenesis
and treatment of glioblastoma. Cells. 10:6202021. View Article : Google Scholar
|
|
16
|
Babaei G, Aziz SG and Jaghi NZZ: EMT,
cancer stem cells and autophagy; the three main axes of metastasis.
Biomed Pharmacother. 133:1109092021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chu X, Tian W, Ning J, Xiao G, Zhou Y,
Wang Z, Zhai Z, Tanzhu G, Yang J and Zhou R: Cancer stem cells:
Advances in knowledge and implications for cancer therapy. Signal
Transduct Target Ther. 9:1702024. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Patil T, Rohiwal SS and Tiwari AP: Stem
cells: Therapeutic implications in chemotherapy and radiotherapy
resistance in cancer therapy. Curr Stem Cell Res Ther. 18:750–765.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Owusu-Akyaw A, Krishnamoorthy K, Goldsmith
LT and Morelli SS: The role of mesenchymal-epithelial transition in
endometrial function. Hum Reprod Update. 25:114–133. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bakir B, Chiarella AM, Pitarresi JR and
Rustgi AK: EMT, MET, plasticity, and tumor metastasis. Trends Cell
Biol. 30:764–776. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bayik D and Lathia JD: Cancer stem
cell-immune cell crosstalk in tumour progression. Nat Rev Cancer.
21:526–536. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen S, Du Y, Guan XY and Yan Q: The
current status of tumor microenvironment and cancer stem cells in
sorafenib resistance of hepatocellular carcinoma. Front Oncol.
13:12045132023. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lei ZN, Teng QX, Koya J, Liu Y, Chen Z,
Zeng L, Chen ZS, Fang S, Wang J, Liu Y and Pan Y: The correlation
between cancer stem cells and epithelial-mesenchymal transition:
Molecular mechanisms and significance in cancer theragnosis. Front
Immunol. 15:14172012024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Saw PE, Liu Q, Wong PP and Song E: Cancer
stem cell mimicry for immune evasion and therapeutic resistance.
Cell Stem Cell. 31:1101–1112. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Muñoz-Galván S, Verdugo-Sivianes EM,
Santos-Pereira JM, Estevez-García P and Carnero A: Essential role
of PLD2 in hypoxia-induced stemness and therapy resistance in
ovarian tumors. J Exp Clin Cancer Res. 43:572024. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li X, González-Maroto C and Tavassoli M:
Crosstalk between CAFs and tumour cells in head and neck cancer.
Cell Death Discov. 10:3032024. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li J, Wang J, Xie D, Pei Q, Wan X, Xing HR
and Ye T: Characteristics of the PI3K/AKT and MAPK/ERK pathways
involved in the maintenance of self-renewal in lung cancer
stem-like cells. Int J Biol Sci. 17:1191–1202. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pang X, He X, Qiu Z, Zhang H, Xie R, Liu
Z, Gu Y, Zhao N, Xiang Q and Cui Y: Targeting integrin pathways:
Mechanisms and advances in therapy. Signal Transduct Target Ther.
8:12023. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bierbaumer L, Katschnig AM, Radic-Sarikas
B, Kauer MO, Petro JA, Högler S, Gurnhofer E, Pedot G, Schäfer BW,
Schwentner R, et al: YAP/TAZ inhibition reduces metastatic
potential of Ewing sarcoma cells. Oncogenesis. 10:22021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lall SP, Alsafwani ZW, Batra SK and
Seshacharyulu P: ASPORIN: A root of the matter in tumors and their
host environment. Biochim Biophys Acta Rev Cancer. 1879:1890292024.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lim YC, Kang HJ and Moon JH: C-Met pathway
promotes self-renewal and tumorigenecity of head and neck squamous
cell carcinoma stem-like cell. Oral Oncol. 50:633–639. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rezakhani L, Salmani S, Eliyasi Dashtaki M
and Ghasemi S: Resveratrol: Targeting cancer stem cells and ncRNAs
to overcome cancer drug resistance. Curr Mol Med. 24:951–961. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kou N, Liu S, Li X, Li W, Zhong W, Gui L,
Chai S, Ren X, Na R, Zeng T and Liu H: H19 facilitates tongue
squamous cell carcinoma migration and invasion via sponging
miR-let-7. Oncol Res. 27:173–182. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wei J, Yang Y, Li Y, Mo X, Guo X, Zhang X,
Xu X, Jiang Z and You Q: Synthesis and evaluation of
N-(benzofuran-5-yl)aromaticsulfonamide derivatives as novel HIF-1
inhibitors that possess anti-angiogenic potential. Bioorg Med Chem.
25:1737–1746. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Condurat AL, Aminzadeh-Gohari S, Malnar M,
Schider N, Opitz L, Thomas R, Menon V, Kofler B and Pruszak J:
Verteporfin-induced proteotoxicity impairs cell homeostasis and
survival in neuroblastoma subtypes independent of YAP/TAZ
expression. Sci Rep. 13:37602023. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jokela TA and LaBarge MA: Integration of
mechanical and ECM microenvironment signals in the determination of
cancer stem cell states. Curr Stem Cell Rep. 7:39–47. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ayob AZ and Ramasamy TS: Cancer stem cells
as key drivers of tumour progression. J Biomed Sci. 25:202018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gurel C, Inetas G, Hortu I, Tunc E, Kuscu
GC, Dindaroglu FC, Sahin O, Buhur A and Oktem G: Cancer and cancer
stem cells: New molecular perspectives. Crit Rev Oncog. 24:99–104.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Atashzar MR, Baharlou R, Karami J,
Abdollahi H, Rezaei R, Pourramezan F and Zoljalali Moghaddam SH:
Cancer stem cells: A review from origin to therapeutic
implications. J Cell Physiol. 235:790–803. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dwivedi AR, Thakur A and Kumar V,
Skvortsova I and Kumar V: Targeting cancer stem cells pathways for
the effective treatment of cancer. Curr Drug Targets. 21:258–278.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee MY, Giraddi RR and Tam WL: Cancer stem
cells: Concepts, challenges, and opportunities for cancer therapy.
Methods Mol Biol. 2005:43–66. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Galassi C, Esteller M, Vitale I and
Galluzzi L: Epigenetic control of immunoevasion in cancer stem
cells. Trends Cancer. 10:1052–1071. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu J, Xiao Q, Xiao J, Niu C, Li Y, Zhang
X, Zhou Z, Shu G and Yin G: Wnt/β-catenin signalling: function,
biological mechanisms, and therapeutic opportunities. Signal
Transduct Target Ther. 7:32022. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yu F, Yu C, Li F, Zuo Y, Wang Y, Yao L, Wu
C, Wang C and Ye L: Wnt/β-catenin signaling in cancers and targeted
therapies. Signal Transduct Target Ther. 6:3072021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang X, Dong N and Hu X: Wnt/β-catenin
Signaling Inhibitors. Curr Top Med Chem. 23:880–896. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cheng X, Xu X, Chen D, Zhao F and Wang W:
Therapeutic potential of targeting the Wnt/β-catenin signaling
pathway in colorectal cancer. Biomed Pharmacother. 110:473–481.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hayat R, Manzoor M and Hussain A: Wnt
signaling pathway: A comprehensive review. Cell Biol Int.
46:863–877. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Agaimy A and Haller F: CTNNB1
(β-catenin)-altered neoplasia: A review focusing on soft tissue
neoplasms and parenchymal lesions of uncertain histogenesis. Adv
Anat Pathol. 23:1–12. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mccrea PD and Gottardi CJ: Beyond
β-catenin: Prospects for a larger catenin network in the nucleus.
Nat Rev Mol Cell Biol. 17:55–64. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Muhammad N, Bhattacharya S, Steele R,
Phillips N and Ray RB: Involvement of c-Fos in the promotion of
cancer stem-like cell properties in head and neck squamous cell
carcinoma. Clin Cancer Res. 23:3120–3128. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Reyes M, Flores T, Betancur D,
Peña-Oyarzún D and Torres VA: Wnt/β-catenin signaling in oral
carcinogenesis. Int J Mol Sci. 21:46822020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhang Y and Wang X: Targeting the
Wnt/β-catenin signaling pathway in cancer. J Hematol Oncol.
13:1652020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Safa AR: Resistance to cell death and its
modulation in cancer stem cells. Crit Rev Oncog. 21:203–219. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yan Q, Fang X, Li C, Lan P and Guan X:
Oncofetal proteins and cancer stem cells. Essays Biochem.
66:423–433. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wen X, Wu Y, Awadasseid A, Tanaka Y and
Zhang W: New advances in canonical Wnt/β-catenin signaling in
cancer. Cancer Manag Res. 12:6987–6998. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ingham PW: Hedgehog signaling. Curr Top
Dev Biol. 149:1–58. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Smith AE, Sigurbjörnsdóttir ES,
Steingrímsson E and Sigurbjörnsdóttir S: Hedgehog signalling in
bone and osteoarthritis: The role of Smoothened and cholesterol.
FEBS J. 290:3059–3075. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Jiang J: Hedgehog signaling mechanism and
role in cancer. Semin Cancer Biol. 85:107–122. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Jia J and Jiang J: Regulation of
smoothened trafficking and abundance in hedgehog signaling. Front
Cell Dev Biol. 10:8478442022. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Wu LF, Gao L, Hou XM, Zhang QH, Li S, Yang
YF and Lin XH: Drosophila miR-5 suppresses Hedgehog signaling by
directly targeting smoothened. FEBS Lett. 586:4052–4060. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Maier D: Membrane-anchored hairless
protein restrains notch signaling activity. Genes (Basel).
11:13152020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Salaritabar A, Berindan-Neagoe I, Darvish
B, Hadjiakhoondi F, Manayi A, Devi KP, Barreca D, Orhan IE, Süntar
I, Farooqi AA, et al: Targeting Hedgehog signaling pathway: Paving
the road for cancer therapy. Pharmacol Res. 141:466–480. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Shen D, Xia Y, Fu Y, Cao Q, Chen W, Zhu Y,
Guo K and Sun L: Hedgehog pathway and cancer: A new area (review).
Oncol Rep. 52:1162024. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhang Q and Jiang J: Regulation of
Hedgehog signal transduction by ubiquitination and
deubiquitination. Int J Mol Sci. 22:133382021. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Liu A: Proteostasis in the Hedgehog
signaling pathway. Semin Cell Dev Biol. 93:153–163. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Mohan M, Mannan A and Singh TG:
Therapeutic implication of Sonic Hedgehog as a potential modulator
in ischemic injury. Pharmacol Rep. 75:838–860. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Doheny D, Manore SG, Wong GL and Lo HW:
Hedgehog signaling and truncated GLI1 in cancer. Cells. 9:21142020.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Suchors C and Kim J: Canonical Hedgehog
pathway and noncanonical GLI transcription factor activation in
cancer. Cells. 11:25232022. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhang Y, Xie J, Wu H, Huang J, Zheng D,
Wang S, Jia X, He Z, Gong Y, Ju L and Sun Q: NK cell based
immunotherapy against oral squamous cell carcinoma. Front Immunol.
15:14407642024. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Raleigh DR and Reiter JF: Misactivation of
Hedgehog signaling causes inherited and sporadic cancers. J Clin
Invest. 129:465–475. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang J, Fan J, Zeng X, Nie M, Luan J,
Wang Y, Ju D and Yin K: Hedgehog signaling in gastrointestinal
carcinogenesis and the gastrointestinal tumor microenvironment.
Acta Pharm Sin B. 11:609–620. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Sigafoos AN, Paradise BD and
Fernandez-Zapico ME: Hedgehog/GLI signaling pathway: Transduction,
regulation, and implications for disease. Cancers (Basel).
13:34102021. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Koukourakis IM, Platoni K, Kouloulias V,
Arelaki S and Zygogianni A: Prostate cancer stem cells: Biology and
treatment implications. Int J Mol Sci. 24:148902023. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Izadpanah A, Mohammadkhani N, Masoudnia M,
Ghasemzad M, Saeedian A, Mehdizadeh H, Poorebrahim M and Ebrahimi
M: Update on immune-based therapy strategies targeting cancer stem
cells. Cancer Med. 12:18960–18980. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Köseer AS, Di Gaetano S, Arndt C, Bachmann
M and Dubrovska A: Immunotargeting of cancer stem cells. Cancers
(Basel). 15:16082023. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hori K, Sen A and Artavanis-Tsakonas S:
Notch signaling at a glance. J Cell Sci. 126:2135–2140.
2013.PubMed/NCBI
|
|
77
|
Kopan R: Notch signaling. Cold Spring Harb
Perspect Biol. 4:a0112132012. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wang H, Zang C, Liu XS and Aster JC: The
role of Notch receptors in transcriptional regulation. J Cell
Physiol. 230:982–988. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Shi Q, Xue C, Zeng Y, Yuan X, Chu Q, Jiang
S, Wang J, Zhang Y, Zhu D and Li L: Notch signaling pathway in
cancer: From mechanistic insights to targeted therapies. Signal
Transduct Target Ther. 9:1282024. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zhou B, Lin W, Long Y, Yang Y, Zhang H, Wu
K and Chu Q: Notch signaling pathway: Architecture, disease, and
therapeutics. Signal Transduct Target Ther. 7:952022. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zanotti S and Canalis E: Notch signaling
and the skeleton. Endocr Rev. 37:223–253. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Vázquez-Ulloa E, Lin KL, Lizano M and
Sahlgren C: Reversible and bidirectional signaling of notch
ligands. Crit Rev Biochem Mol Biol. 57:377–398. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Paniri A, Hosseini MM, Amjadi-Moheb F,
Tabaripour R, Soleimani E, Langroudi MP, Zafari P and Akhavan-Niaki
H: The epigenetics orchestra of Notch signaling: A symphony for
cancer therapy. Epigenomics. 15:1337–1358. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Meurette O and Mehlen P: Notch signaling
in the tumor microenvironment. Cancer Cell. 34:536–548. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Bellavia D, Checquolo S, Palermo R and
Screpanti I: The Notch3 receptor and its intracellular
signaling-dependent oncogenic mechanisms. Adv Exp Med Biol.
1066:205–222. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Sun W, Gaykalova DA, Ochs MF, Mambo E,
Arnaoutakis D, Liu Y, Loyo M, Agrawal N, Howard J, Li R, et al:
Activation of the NOTCH pathway in head and neck cancer. Cancer
Res. 74:1091–1104. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Suarez RF, Sanlidag S and Sahlgren C:
Mechanical regulation of the Notch signaling pathway. Curr Opin
Cell Biol. 85:1022442023. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Ranj T: Upcoming market catalysts in Q4
2019. Nat Rev Drug Discov. 18:7382019. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Zheng Y, Wei K, Jiang P, Zhao J, Shan Y,
Shi Y, Zhao F, Chang C, Li Y, Zhou M, et al: Macrophage
polarization in rheumatoid arthritis: Signaling pathways, metabolic
reprogramming, and crosstalk with synovial fibroblasts. Front
Immunol. 15:13941082024. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Falo-Sanjuan J and Bray SJ: Decoding the
Notch signal. Dev Growth Differ. 62:4–14. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yang L, Shi P, Zhao G, Xu J, Peng W, Zhang
J, Zhang G, Wang X, Dong Z, Chen F and Cui H: Targeting cancer stem
cell pathways for cancer therapy. Signal Transduct Target Ther.
5:82020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Espinoza I and Miele L: Deadly crosstalk:
Notch signaling at the intersection of EMT and cancer stem cells.
Cancer Lett. 341:41–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Zhu P and Fan Z: Cancer stem cells and
tumorigenesis. Biophys Rep. 4:178–188. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Xie J, Qi X, Wang Y, Yin X, Xu W, Han S,
Cai Y and Han W: Cancer-associated fibroblasts secrete
hypoxia-induced serglycin to promote head and neck squamous cell
carcinoma tumor cell growth in vitro and in vivo by activating the
Wnt/β-catenin pathway. Cell Oncol (Dordr). 44:661–671. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Djeungoue-Petga MA, Lurette O, Jean S,
Hamel-Côté G, Martín-Jiménez R, Bou M, Cannich A, Roy P and
Hebert-Chatelain E: Intramitochondrial Src kinase links
mitochondrial dysfunctions and aggressiveness of breast cancer
cells. Cell Death Dis. 10:9402019. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Garcia N, Al-Hendy A, Baracat EC, Carvalho
KC and Yang Q: Targeting hedgehog pathway and DNA
methyltransferases in uterine leiomyosarcoma cells. Cells.
10:532020. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Miyasaka A, Oda K, Ikeda Y, Sone K, Fukuda
T, Inaba K, Makii C, Enomoto A, Hosoya N, Tanikawa M, et al:
PI3K/mTOR pathway inhibition overcomes radioresistance via
suppression of the HIF1-α/VEGF pathway in endometrial cancer.
Gynecol Oncol. 138:174–180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Xin Y, Jiang Q, Liu C and Qiu J: Plumbagin
has an inhibitory effect on the growth of TSCC PDX model and it
enhances the anticancer efficacy of cisplatin. Aging (Albany NY).
15:12225–12250. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Joshi P and Waghmare S: Molecular
signaling in cancer stem cells of tongue squamous cell carcinoma:
Therapeutic implications and challenges. World J Stem Cells.
15:438–452. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Antra Parashar P, Hungyo H, Jain A, Ahmad
S and Tandon V: Unraveling molecular mechanisms of head and neck
cancer. Crit Rev Oncol Hematol. 178:1037782022. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Soltaninezhad P, Mohtasham N, Arab F,
Sadeghi M, EbrahimZadeh N, Azghadi SF and Mohajertehran F:
Therapeutic potential of siRNAs in tongue squamous cell carcinoma
by modulating the PI3K/AKT and ERK signaling pathways: A systematic
review. Cell J. 26:337–350. 2024.PubMed/NCBI
|
|
102
|
Asl ER, Amini M, Najafi S, Mansoori B,
Mokhtarzadeh A, Mohammadi A, Lotfinejad P, Bagheri M, Shirjang S,
Lotfi Z, et al: Interplay between MAPK/ERK signaling pathway and
MicroRNAs: A crucial mechanism regulating cancer cell metabolism
and tumor progression. Life Sci. 278:1194992021. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Didonato JA, Mercurio F and Karin M: NF-κB
and the link between inflammation and cancer. Immunol Rev.
246:379–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Peng Y, Wang Y, Zhou C, Mei W and Zeng C:
PI3K/Akt/mTOR pathway and its role in cancer therapeutics: Are we
making headway? Front Oncol. 12:8191282022. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Rocamora-Blanch G, Climent F and Solanich
X: Histiocytosis. Med Clin (Barc). 161:166–175. 2023.(In English,
Spanish). View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhang Y and Weinberg RA:
Epithelial-to-mesenchymal transition in cancer: Complexity and
opportunities. Front Med. 12:361–373. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Huang N, Sun X, Li P, Liu X, Zhang X, Chen
Q and Xin H: TRIM family contribute to tumorigenesis, cancer
development, and drug resistance. Exp Hematol Oncol. 11:752022.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Ang HL, Mohan CD, Shanmugam MK, Leong HC,
Makvandi P, Rangappa KS, Bishayee A, Kumar AP and Sethi G:
Mechanism of epithelial-mesenchymal transition in cancer and its
regulation by natural compounds. Med Res Rev. 43:1141–1200. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Saikia PJ, Pathak L, Mitra S and Das B:
The emerging role of oral microbiota in oral cancer initiation,
progression and stemness. Front Immunol. 14:11982692023. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Jin Y, Wang Z, He D, Zhu Y, Chen X and Cao
K: Identification of novel subtypes based on ssGSEA in
immune-related prognostic signature for tongue squamous cell
carcinoma. Cancer Med. 10:8693–8707. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Naponelli V, Rocchetti MT and Mangieri D:
Apigenin: Molecular mechanisms and therapeutic potential against
cancer spreading. Int J Mol Sci. 25:55692024. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Yadav AK and Desai NS: Cancer stem cells:
Acquisition, characteristics, therapeutic implications, targeting
strategies and future prospects. Stem Cell Rev Rep. 15:331–355.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Flaherty KT, Hodi FS and Fisher DE: From
genes to drugs: Targeted strategies for melanoma. Nat Rev Cancer.
12:349–361. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Freedman LP: On rigor and replication.
Science. 356:342017. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Kuşoğlu A and Biray Avcı Ç: Cancer stem
cells: A brief review of the current status. Gene. 681:80–85. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zou B, Sun S, Qi X and Ji P: Aldehyde
dehydrogenase activity is a cancer stem cell marker of tongue
squamous cell carcinoma. Mol Med Rep. 5:1116–1120. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Chaudhury S, Panda S, Mohanty N, Panda S,
Mohapatra D, Nagaraja R, Sahoo A, Gopinath D, Lewkowicz N and
Lapinska B: Can immunoexpression of cancer stem cell markers
prognosticate tongue squamous cell carcinoma? A systematic review
and meta-analysis. J Clin Med. 12:27532023. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Yin J, Jiang Y, Wu H, Wang J, Zhang S and
Liu H: Overexpression of ZFX and its involvement in squamous cell
carcinoma of the tongue. Oncol Rep. 33:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Liu Z, He Q, Ding X, Zhao T, Zhao L and
Wang A: SOD2 is a C-myc target gene that promotes the migration and
invasion of tongue squamous cell carcinoma involving cancer
stem-like cells. Int J Biochem Cell Biol. 60:139–146. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Varnat F, Siegl-Cachedenier I, Malerba M,
Gervaz P and Ruiz i Altaba A: Loss of WNT-TCF addiction and
enhancement of HH-GLI1 signalling define the metastatic transition
of human colon carcinomas. EMBO Mol Med. 2:440–457. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Lee J, Kim E, Chong K, Ryu SW, Kim C, Choi
K, Kim JH and Choi C: Atypical induction of HIF-1α expression by
pericellular Notch1 signaling suffices for the malignancy of
glioblastoma multiforme cells. Cell Mol Life Sci. 79:5372022.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Wu Y, Wan XW, Jiang L, Wang W, Zhu JJ and
Shao YS: Rhaponitin reverses cisplatin resistance and impairs
cancer stemness through HIF-1α/MCT4/Wnt pathway in tongue squamous
cell carcinoma. Kaohsiung J Med Sci. 41:e700692025. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Anglès F, Gupta V, Wang C and Balch WE:
COPII cage assembly factor Sec13 integrates information flow
regulating endomembrane function in response to human variation.
Sci Rep. 14:101602024. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Wang J, Li L, Gao L, Guan C, Su K, Li L,
Luo W, Chen H and Ji P: Identification of differentially expressed
genes in oral squamous cell carcinoma TCA8113 cells. Oncol Lett.
14:7055–7068. 2017.PubMed/NCBI
|
|
125
|
Li Y, Wang Z, Ajani JA and Song S: Drug
resistance and cancer stem cells. Cell Commun Signal. 19:192021.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Chinn LK, Ovchinnikova I, Sukmanova AA,
Davydova AO and Grigorenko EL: Early institutionalized care
disrupts the development of emotion processing in prosody. Dev
Psychopathol. 33:421–430. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Yang J, Ji P, Zou B, Sun S and Qi X:
Biological characteristics of cells expressing high level of
aldehyde dehydrogenase subpopulation in tongue squamous cell
carcinoma Tca8113 cell line. Hua Xi Kou Qiang Yi Xue Za Zhi.
30:439–443. 2012.(In Chinese). PubMed/NCBI
|
|
128
|
Sun X, Lv X, Yan Y, Zhao Y, Ma R, He M and
Wei M: Hypoxia-mediated cancer stem cell resistance and targeted
therapy. Biomed Pharmacother. 130:1106232020. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Najafi M, Farhood B, Mortezaee K,
Kharazinejad E, Majidpoor J and Ahadi R: Hypoxia in solid tumors: A
key promoter of cancer stem cell (CSC) resistance. J Cancer Res
Clin Oncol. 146:19–31. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zeng Z, Fu M, Hu Y, Wei Y, Wei X and Luo
M: Regulation and signaling pathways in cancer stem cells:
Implications for targeted therapy for cancer. Mol Cancer.
22:1722023. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Zhou M, Niu H, Cui D, Xu M, Li J, Huang G,
Zhou M, Xiong C, Liu Y, Xu X, et al: Cancer stem cell-driven drug
resistance in colorectal carcinoma: Molecular aspects and
therapeutic potentials. Mol Cancer. 25:542026. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
No authors listed. Deutsche
pharmakologische gesellschaft. Abstracts. 27th spring meeting,
March 11–14, 1986, Mainz. Naunyn Schmiedebergs Arch Pharmacol. 332
(Suppl):R1–R104. 1986.
|
|
133
|
Najafi M, Farhood B and Mortezaee K:
Cancer stem cells (CSCs) in cancer progression and therapy. J Cell
Physiol. 234:8381–8395. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Lathia J, Liu H and Matei D: The clinical
impact of cancer stem cells. Oncologist. 25:123–131. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Zhou H, Tan L, Liu B and Guan XY: Cancer
stem cells: Recent insights and therapies. Biochem Pharmacol.
209:1154412023. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Huang B, Yan X and Li Y: Cancer stem cell
for tumor therapy. Cancers (Basel). 13:48142021. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Akbar Samadani A, Keymoradzdeh A, Shams S,
Soleymanpour A, Elham Norollahi S, Vahidi S, Rashidy-Pour A, Ashraf
A, Mirzajani E, Khanaki K, et al: Mechanisms of cancer stem cell
therapy. Clin Chim Acta. 510:581–592. 2020. View Article : Google Scholar : PubMed/NCBI
|














