Introduction
Gliomas, a series of malignant tumors originating
from the neuroglial tissues, primarily occur in the brain and
spinal cord. As the most common primary malignant tumor of the
central nervous system (CNS), gliomas exhibit a relatively high
incidence rate of 6.6 to 7.4 per 100,000 individuals and are
particularly prevalent among adults, accounting for ~75% of all
brain malignancies (1,2). Gliomas are typically classified into
several subtypes, among which glioblastoma (GBM), a highly
aggressive variant characterized by rapid growth and invasive
behavior, is the most prevalent (3). Due to its complex anatomical location
within the CNS, surgical resection of GBM poses a notable challenge
(4).
Temozolomide (TMZ), an alkylating chemotherapeutic
agent, has shown marked clinical efficacy in the treatment of
gliomas (5). TMZ induces tumor cell
apoptosis and inhibits DNA synthesis and repair in tumor cells.
Moreover, it has been widely incorporated into glioma chemotherapy
regimens, with notable survival benefits in clinical trials,
extending median overall survival in newly diagnosed patients with
glioblastoma to ~15 months (6,7).
However, the responses of patients with GBM to TMZ vary, with
certain individuals developing drug resistance, which greatly
limits therapeutic effectiveness (8). Therefore, novel therapeutic strategies
are being explored to enhance TMZ efficacy and overcome drug
resistance.
As TMZ chemotherapy failure in gliomas is primarily
attributed to acquired drug resistance, research suggests that
immunosuppressive cytokines serve notable roles in the progression
of gliomas and TMZ resistance acquisition in gliomas (9). However, the precise mechanisms by
which these cytokines mediate TMZ resistance remain poorly
understood.
IL-19, an immunosuppressive cytokine associated with
poor GBM prognosis, has been reported to promote invasiveness via
the IL-19/WNT1-inducible signaling pathway protein 1 pathway,
whilst its blockade suppressed tumor growth by enhancing antitumor
immunity and reducing tumor-associated macrophage (TAM)-mediated
suppression (9). Moreover, elevated
IL-18 expression in GBM cells has been demonstrated to induce TMZ
resistance and form an immunosuppressive microenvironment by
activating the PI3K/Akt pathway and upregulating CD274 expression
(10). These findings emphasize
that targeting cytokines may be a critical strategy for overcoming
TMZ resistance in GBM, providing the foundation for novel
therapeutic approaches.
IL-37, another immunosuppressive cytokine, has
emerged as a potential prognostic biomarker in gliomas, where
reduced IL-37 expression is associated with the poor survival of
patients (11). Among the five
transcriptional variants of IL-37, IL-37b is the most extensively
studied (12). Concurrently,
TMZ-resistant gliomas secrete immunosuppressive factors such as the
cytokine IL-10 to recruit regulatory T cells (Tregs) and
subsequently suppress the functions of cytotoxic T lymphocytes and
natural killer cells, which fosters an immunosuppressive
microenvironment (13,14). This suggests that IL-37 may
similarly contribute to the immune evasion and chemoresistance of
GBM through similar mechanisms.
Dysregulated mitogen-activated protein kinase (MAPK)
signaling drives oncogenic proliferation, invasion and metastasis
by activating downstream effectors such as VEGF (15). The MAPK signaling pathway includes
three major categories of kinases: MAPK kinase kinase (MAPKKK),
MAPK kinase (MAPKK) and MAPK (16).
MAPKKK family proteins are the upstream kinases of MAPK cascades
and are responsible for activating the downstream MAPKK, including
rapidly accelerated fibrosarcoma (RAF)-like kinases, MAPK/ERK
kinase kinases and other members such as apoptosis
signal-regulating kinase 1 (17,18).
MAPKs are the terminal kinases of MAPK cascades and are mainly
divided into extracellular signal-regulated kinase-1/2 (ERK-1/2),
c-Jun N-terminal kinase-1/2/3, p38 MAPK and ERK5, which are
activated by extracellular cytokines, neurotransmitters, hormones
and stress signals (19,20). Mutated RAF-like kinases in tumor
cells often lead to constitutive activation of the MAPK signaling
pathway and drive the uncontrolled proliferation of tumors
(21), whereas mutations in
melanomas result in persistent activation of the RAS/MAPK pathway,
aberrantly hyperactivated CDK12 and enhanced DNA repair capability,
eventually promoting the establishment of drug resistance (22). Activation of ERK signaling also
protects leukemic monocytes from chronic myelomonocytic leukemia by
upregulating the anti-apoptotic protein myeloid cell leukemia-1
(23). For TMZ resistance in GBM,
inhibiting expression of MAPK8 and subsequent activation of the
MAPK signaling pathway were reported to markedly induce apoptotic
responses in GBM cells (24).
To address this, the present study aimed to
investigate the functional role of IL-37 in the malignant
progression and TMZ resistance of GBM. Using a combination of in
vitro gain- and loss-of-function approaches in GBM cell lines,
including rIL-37 treatment, gene silencing and overexpression
alongside transcriptomic profiling via RNA sequencing, the present
study aimed to elucidate the underlying mechanisms by which IL-37
modulates cellular survival and apoptotic responses under
chemotherapeutic stress.
Materials and methods
Cell culture and treatments
The human GBM cell lines U-251 MG (U251; cat. no.
SCSP-559), U-87 MG (U87; cat. no. TCHu138; glioblastoma of unknown
origin) and A172 (TCHu171) were obtained from The Cell Bank of Type
Culture Collection of The Chinese Academy of Sciences, and
maintained in high-glucose DMEM/MEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin under standard
conditions (37°C and 5% CO2). All cell lines have been
STR validated. To establish TMZ-resistant (TR) sublines, parental
cells were chronically exposed to progressively increasing
concentrations of TMZ (starting from 5 µM, with a 5 µM increment
every 2 weeks) over a period of 6 months. The resistant phenotype
was confirmed by Cell Counting Kit-8 (CCK-8) assay (cat. no.
HY-K0301; MedChemExpress) and remained stable for at least 10
passages in the absence of TMZ. For TMZ treatment, varying
concentrations of TMZ (cat. no. HY-17364; MedChemExpress) dissolved
in DMSO, corresponding to log concentrations of 0, 1.5, 2, 2.5, 3
and 3.5 µM, were added to the culture medium of U251 cells for 48
h. To evaluate the effects of IL-37 on cellular response to TMZ,
U251 cells received recombinant human IL-37b (rIL-37; 10 ng/ml;
cat. no. 7585-IL; R&D Systems, Inc.) dissolved in sterile PBS
in combination with TMZ and finally harvested for the subsequent
analysis. To evaluate the necessity of MAPK signaling, cells were
pretreated with the MAPK inhibitor Adezmapimod (10 µM; cat. no.
HY-10256; MedChemExpress) for 1 h prior to TMZ incubation or
co-incubation with TMZ and rIL-37 at 37°C for 48 h.
Transfection of small interfering
(si)RNAs and plasmids
siRNAs targeting IL-37/single immunoglobulin
interleukin-1 related receptor (SIGIRR) and human IL-37
overexpression plasmids were manufactured by GenScript Biotech
Corporation using the pcDNA3.1 backbone. U251 or U251-TR cells were
transfected with siRNAs (50 nM) or plasmids (1 µg/µl) using
Lipofectamine 2000™ (cat. no. 11668-019; Invitrogen; Thermo Fisher
Scientific, Inc.), and subsequent experiments were performed 48 h
post-transfection following standardized protocols provided by the
manufacturer. The siRNA targeting IL-37 (si-IL-37) consisted of a
sense strand 5′-CAAUGUGUUUCCUGUUCUC-3′ and an antisense strand
5′-GAGAACAGGAAACACAUUG-3′. The siRNA targeting SIGIRR (si-SIGIRR)
consisted of a sense strand 5′-AUGAAGUUCACGAACUUGC-3′ and an
antisense strand 5′-GCAAGUUCGUGAACUUCAU-3′. The negative control
siRNA (si-NC) consisted of a sense strand 5′-ACGUGACACGUUCGGAGAA-3′
and an antisense strand 5′-UUCUCCGAACGUGUCACGU-3′.
Flow cytometry
Apoptosis in TMZ-treated U251 cells with or without
rIL-37 was analyzed using flow cytometry. After resuspension, the
harvested cells were incubated with an annexin V-FITC and propidium
iodide kit (PI; cat. no. 40302ES60; Shanghai Yeasen Biotechnology
Co., Ltd.) for 20 min in the dark at room temperature. The samples
were analyzed using a CytoFLEX flow cytometer (Beckman Coulter,
Inc.) with excitation at 488 nm to detect early apoptotic (annexin
V+/PI−) and late apoptotic (annexin
V+/PI+) populations, which were quantified as
the total apoptosis rate. Data analysis was performed using FlowJo
software (version 10.8.1; BD Biosciences).
Senescence-associated
(SA)-β-galactosidase staining
Cellular senescence was detected using a
SA-β-galactosidase staining kit (cat. no. HY-K1089;
MedChemExpress). After finishing corresponding treatments,
1.0×106 U251 cells were fixed with paraformaldehyde for
15 min at room temperature and then incubated with β-galactosidase
staining solution overnight at 37°C. The following day, U251 cells
were captured using a bright-field microscope (Nikon Corporation).
Senescent cells exhibited blue cytoplasmic precipitates, and the
ratio of cellular senescence was calculated by comparing
blue-stained cells to total cells using Fiji software (version
2.9.0; National Institutes of Health).
Immunofluorescence staining
SA DNA damage in U251 cells was determined using a
DNA Damage Assay Kit by γ-H2A histone family member X (H2AX)
immunofluorescence (cat. no. C2035S; Beyotime Biotechnology).
Briefly, after TMZ and rIL37 treatments, U251 cells were fixed with
paraformaldehyde (cat. no. P0099; Beyotime Biotechnology) for 15
min at room temperature and permeabilized with 0.1% Triton X-100
(cat. no. BL934B; Biosharp Life Sciences) for another 15 min at
room temperature. After blocking with 5% bovine serum albumin (BSA)
for 1 h at room temperature, the samples were incubated with
anti-γ-H2AX antibodies (cat. no. C2035S-4; Beyotime Biotechnology)
at 4°C overnight, followed by AF488-labeled goat anti-rabbit IgG
(cat. no. C2035S-5; Beyotime Biotechnology) for 1 h at room
temperature in the dark. The samples were counterstained with DAPI
(cat. no. C2035S-6; Beyotime Biotechnology) for 5 min and then
subjected to fluorescence microscopy (Zeiss AG) to capture the
images. The mean immunofluorescence intensities of γ-H2AX-positive
nuclei were quantified using Fiji software (version 2.9.0; National
Institutes of Health).
Colony formation assay
The proliferative capabilities of U251 cells were
assessed using a colony formation assay. Briefly, U251 cells seeded
in 6-well plates (1×103 cells/well) underwent TMZ or
rIL-37 treatment and were cultured at 37°C for 2 weeks. The
aforementioned culture medium (Gibco; Thermo Fisher Scientific,
Inc.) was refreshed every 3 days to maintain the nutrient supply.
Colonies were fixed with paraformaldehyde (cat. no. P0099; Beyotime
Biotechnology) for 15 min and stained with 0.1% crystal violet
solution (cat. no. C0121; Beyotime Biotechnology) for 20 min at
room temperature. Visible colonies (>50 cells/colony) were
quantified using ImageJ software (National Institutes of Health)
under bright-field microscopy.
CCK-8 assay
The survival of TMZ-treated U251 cells, with or
without rIL-37, was determined using the CCK-8 assay. U251 cells
were seeded in 96-well plates at a density of 5×103
cells/well. After TMZ treatments, 10 µl CCK-8 reagent (cat. no.
BS350A; Biosharp Life Sciences) was added to each well and
incubated for another 4 h in the dark at 37°C. Optical density at
450 nm was determined using a microplate reader (Thermo Fisher
Scientific, Inc.) every 1 h during a 4 h-incubation period.
Reverse transcription-quantitative PCR
(RT-qPCR) assay
The efficiency of IL-37 knockdown and overexpression
was assessed using RT-qPCR. U251 cells were lysed with
TRIzol® reagent (cat. no. RC202-01; Vazyme Biotech Co.,
Ltd.) to extract total RNA. The total RNA concentrations were
quantified using NanoDrop™ 2000 (Thermo Fisher Scientific, Inc.)
and the quality was determined by the A260/A280 ratio. Reverse
transcription was performed using PrimeScript™ RT Master Mix (cat.
no. RR036B; Takara Bio, Inc.) to synthesize cDNA templates, and
qPCR was performed using SYBR® Premix Ex Taq II (cat.
no. RR390A; Takara Bio, Inc.) following the manufacturer's
instructions on a QuantStudio™ 6 Flex Real-Time PCR System (Thermo
Fisher Scientific, Inc.). Relative expression of target genes was
calculated using the 2−ΔΔCq method (25) and normalized to GAPDH. The RNA
primers used were as follows: IL-37-forward (F),
5′-TTCTTTGCATTAGCCTCATCCTT-3′ and reverse (R),
5′-CGTGCTGATTCCTTTTGGGC-3′; IL-1B-F, 5′-AATCTGTACCTGTCCTGCGTGTT-3′
and R, 5′-TGGGTAATTTTTGGGATCTACACTCT-3′; IL-6-F,
5′-TTCTCCACAAGCGCCTTCGGTC-3′ and R, 5′-TCTGTGTGGGGCGGCTACATCT-3′;
and SIGIRR-F, 5′-CTCCCCGTCTGAAGACCAG-3′ and R,
5′-CCCCAATTCCCAATGGAAGC-3′ GAPDH-F, 5′-AGATCCCTCCAAAATCAAGTGG-3′
and R, 5′-GGCAGAGATGATGACCCTTTT-3′.
RNA-sequencing (RNA-seq) analysis
RNA-seq analyses were performed by GENE DENOVO
(https://www.genedenovo.com/). After TMZ
exposure with or without rIL-37 treatment with three biological
replicates set for each treatment group, the total RNA of U251
cells was extracted using a Trizol reagent kit (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
RNA quality was assessed on an Agilent 2100 Bioanalyzer (Agilent
Technologies, Inc.) and checked using RNase free agarose gel
electrophoresis. After the total RNA was extracted, eukaryotic mRNA
was enriched using Oligo(dT) beads. Then the enriched mRNA was
fragmented into short fragments using fragmentation buffer and
reverse transcribed into cDNA by using NEBNext Ultra RNA Library
Prep Kit for Illumina (cat. no. #7530; New England BioLabs, Inc.).
The purified double-stranded cDNA fragments were end repaired, A
base added and ligated to Illumina sequencing adapters. The
ligation reaction was purified with the AMPure XP Beads (1.0X) and
PCR amplified. The resulting cDNA library was sequenced using
Illumina Novaseq6000 by GENE DENOVO. Differentially expressed genes
(DEGs) were identified with a threshold set at a false discovery
rate (FDR) of <0.05 and fold-change (FC) of >2
(|log2FC|>1), followed by functional enrichment
analysis of DEGs using Gene Ontology (GO) and Kyoto Encyclopedia of
Genes and Genomes (KEGG) (Metascape; National Institutes of
Health). Heatmap visualization was performed using the pheatmap
function from the ‘pheatmap’ package (version 1.0.12).
Bioinformatics analyses were performed using R software (version
4.2.1; R Foundation for Statistical Computing). The raw sequencing
data are provided in ScienceDB database (https://doi.org/10.57760/sciencedb.32976).
Western blotting
Western blot analysis was performed following
established protocols as previously described (26). Briefly, proteins derived from cell
samples were separated using 10% SDS-PAGE and transferred to PVDF
membranes, which were then blocked with 5% BSA at room temperature
for 1 h, followed by incubation with corresponding primary
antibodies at 4°C overnight and secondary antibodies at room
temperature for 1 h. The intensities of the protein bands were
analyzed using an Fiji software (version 2.9.0; National Institutes
of Health). The following antibodies were used: p16-INK4A (1:1,000;
cat. no. 80772; Cell Signaling Technology, Inc.), p21 (1:1,000;
cat. no. 10355-1-AP; Proteintech Group, Inc.), p38 MAPK (1:2,000;
cat. no. 14064-1-AP; Proteintech Group, Inc.), phosphorylated-p38
MAPK (p-p38; 1:1,000; cat. no. 28796-1-AP; Proteintech Group,
Inc.), β-tubulin (1:5,000; cat. no. ab179513 Abcam) and
HRP-conjugated anti-Rabbit IgG (1:5,000; cat. no. SA00001-2;
Proteintech Group, Inc.).
ELISA
SA-secretory phenotype (SASP) factors in U251 cells
were quantified using the IL-1β ELISA Kit (cat. no. CSB-E08053h;
Cusabio Technology, LLC) and IL-6 ELISA Kit (cat. no. CSB-E04638h;
Cusabio Technology, LLC), following the manufacturer's
protocols.
Statistical analysis
Data are presented as the mean ± standard deviation.
GraphPad Prism software (version 9.0; Dotmatics) was used to
perform the statistical analyses. For two-group comparisons, an
unpaired two-tailed Student's t-test was employed, whilst one-way
ANOVA with the Bonferroni method was applied to compare multi-group
differences. P<0.05 was considered to indicate a statistically
significant difference.
Results
IL-37 modulates the sensitivity of GBM
cells to TMZ
The role of IL-37 in modulating TMZ resistance in
GBM is unclear; therefore, to address this, the present study first
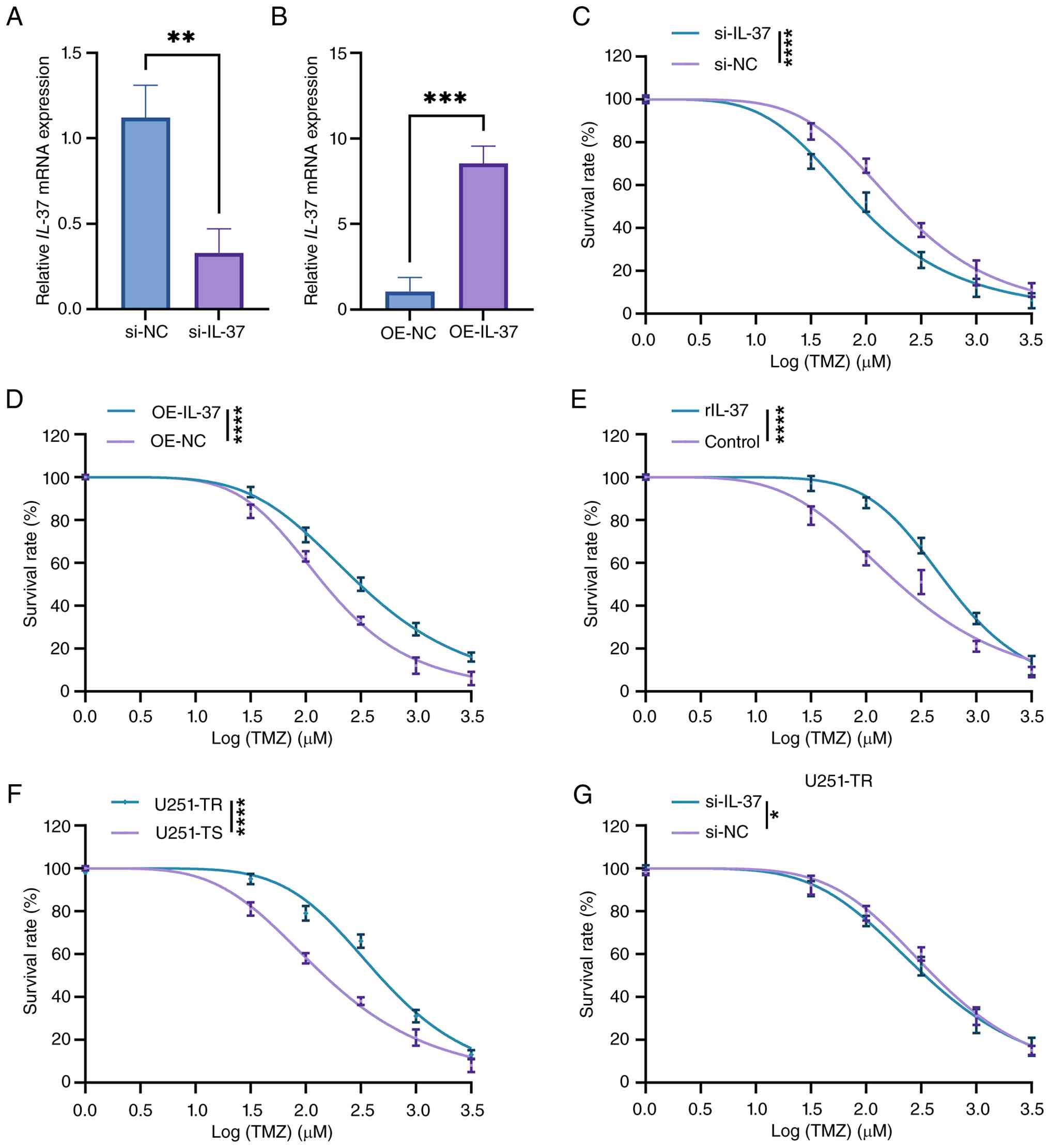
validated the efficiency of IL-37 knockdown and overexpression in
U251 cells using RT-qPCR, which demonstrated ~70% silencing and
7-fold upregulation of IL-37 expression (Fig. 1A and B). The subsequent CCK-8 assays
revealed that IL-37 overexpression enhanced the survival of U251
cells treated with log concentrations of 0, 1.5, 2, 2.5, 3 and 3.5
µM TMZ treatment for 24 h, whilst IL-37 knockdown markedly reduced
cell viability (Fig. 1C and D). As
extracellular IL-37 has been reported to aid in forming an
immunosuppressive microenvironment for immune escape (27,28),
rIL-37 was co-incubated with U251 cells which further diminished
cellular TMZ sensitivity (Fig. 1E).
Moreover, to explore the role of IL-37 in the established
TMZ-resistance, the U251-TR cell line was first generated using
prolonged and elevated TMZ exposure. This cell line exhibited
markedly higher survival rates than the TMZ-sensitive U251 cells
under TMZ exposure (Fig. 1F).
Notably, IL-37 knockdown potentiated TMZ sensitivity in U251-TR
cells (Fig. 1G), indicating a
partial restoration of drug responsiveness. These data collectively
establish IL-37 as a critical mediator of TMZ resistance in
GBM.
 | Figure 1.IL-37 modulates the sensitivity of
glioblastoma cells to TMZ. (A) qPCR analysis of the expression of
IL-37 in U251 cells treated with si-NC, and si-IL-37 (n=3). (B)
qPCR analysis of the expression of IL-37 in U251 cells treated with
OE-NC and OE-IL-37 (n=3). (C) CCK-8 assay of the survival rates of
U251 cells receiving si-IL-37 under TMZ exposure (n=3). (D) CCK-8
assay of the survival rates of U251 cells receiving OE-IL-37 under
TMZ exposure (n=3). (E) CCK-8 assay of the survival rates of U251
cells receiving rIL-37 under TMZ exposure (n=3). (F) The
establishment of U251-TR and U251-TS cells confirmed by CCK-8 assay
(n=3). (G) CCK-8 assay of the survival rates of U251-TR cells
receiving si-IL-37 or si-NC under TMZ exposure (n=3). Data are
expressed as mean ± SD. Statistical significance was set at
*P<0.05. **P<0.01; ***P<0.001; ****P<0.0001. TMZ,
temozolomide; si, small interfering RNA; NC, negative control; OE,
overexpression; U251-TR, temozolomide-resistant U251; U251-TS,
temozolomide-sensitive U251; qPCR, quantitative PCR; CCK-8, Cell
Counting Kit-8; rIL-37, recombinant human IL-37. |
IL-37 suppresses TMZ efficacy by
promoting proliferation and inhibiting apoptosis of GBM cells
To elucidate the role of IL-37 in TMZ-induced
cytotoxicity and chemoresistance in GBM, the present study assessed
the modulatory effects of IL-37 on the proliferation and apoptotic
responses of both U251 and U251-TR cells. Colony formation assays
revealed that rIL-37 co-incubation significantly mitigated
TMZ-induced suppression of U251 cell proliferation, with a
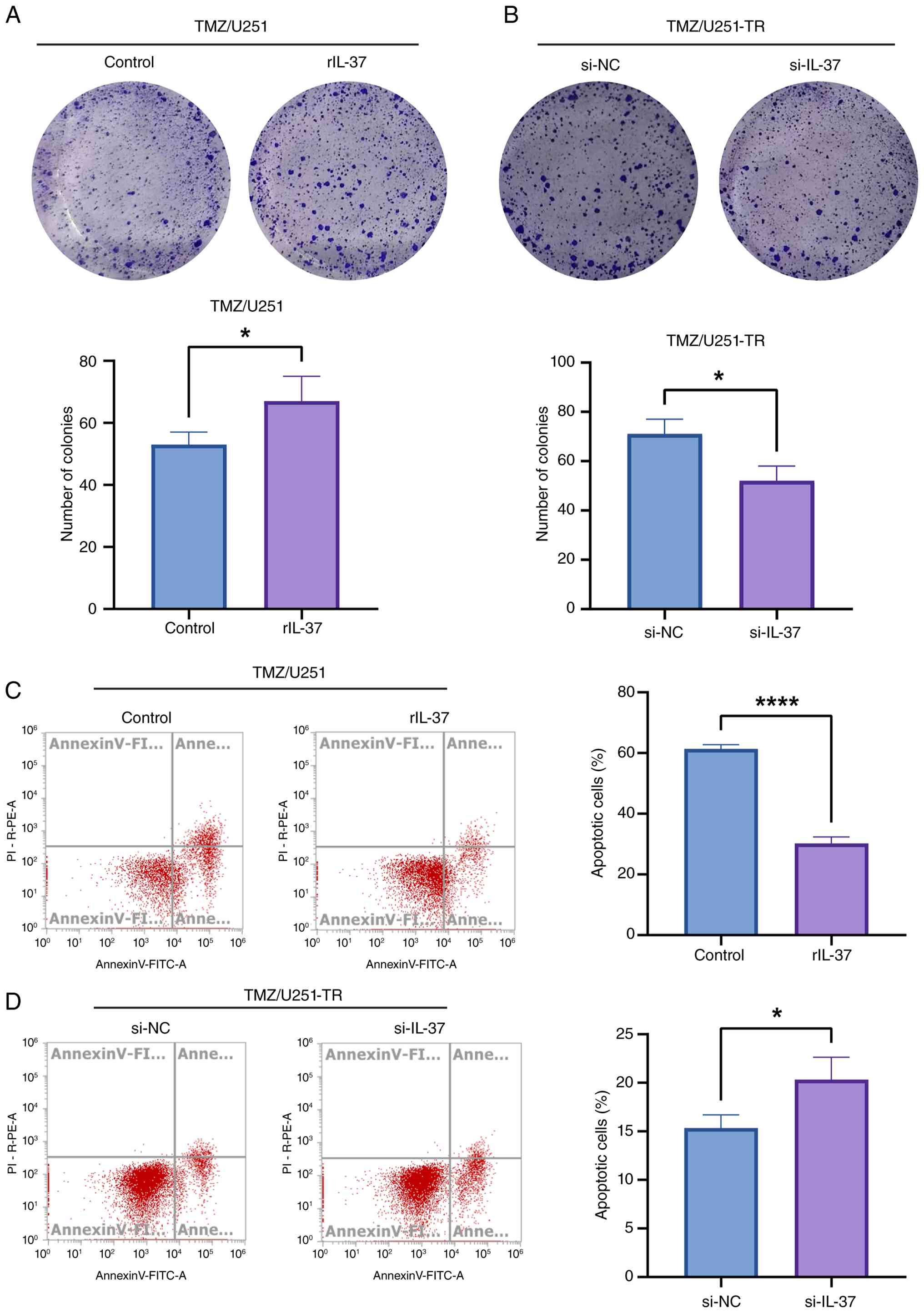
~1.4-fold increase in the colony number (53 vs. 67; Fig. 2A). Conversely, IL-37 knockdown in
U251-TR cells potentiated TMZ-induced inhibitory effects on
cellular proliferation and reduced colony formation (71 vs. 52;
Fig. 2B). Furthermore, flow
cytometry demonstrated that rIL-37 significantly inhibited
TMZ-induced apoptosis in U251 cells (61 vs. 30%; Fig. 2C), whereas si-IL-37 augmented
apoptosis in U251-TR cells (15 vs. 20%; Fig. 2D). These findings suggest that IL-37
sustains GBM cell survival under TMZ pressure by promoting
proliferation and blunting apoptosis.
RNA-seq reveals the activated MAPK
signaling and senescence-related pathways in rIL-37 induce TMZ
resistance
To assess the molecular mechanisms underlying
rIL-37-mediated TMZ resistance in U251 cells, RNA-seq was performed
to identify DEGs and their functional enrichment pathways. The
violin plots comparing RNA-seq data of the control group (TMZ
exposure without rIL-37; n=3) and rIL-37 group (TMZ exposure with
rIL-37 treatment; n=3) exhibited nearly identical shape symmetry
and median alignment, suggesting a comparable global transcript
abundance between the two groups. The high consistency in violin
shapes within each group further validated experimental
reproducibility (Fig. 3A).
Moreover, the principal component analysis 3D score plot of RNA-seq
data revealed a pronounced segregation between the control and
rIL-37 groups along the PC1 axis, indicating the remodeling of
genome-wide expression profiles induced by rIL-37 (Fig. 3B). The pheatmap of RNA-seq data also
displayed clear clustering separation between groups along
principal components, confirming rIL-37-induced profound
alterations in global gene expression, whilst high consistency
among biological replicates within each group again underscored
reproducibility (Fig. 3C). The DEGs
were identified with thresholds set at |log2FC|>1 and
FDR<0.05, and the volcano plot showed 2,075 upregulated and 444
downregulated genes respectively (Fig.
3D). The heatmap of DEGs also demonstrated that there were more
DEGs upregulated in U251 cells undergoing TMZ exposure and rIL-37
treatment, and good repeatability was evidenced by highly
consistent gene expression patterns among samples within each group
(Fig. 3E).
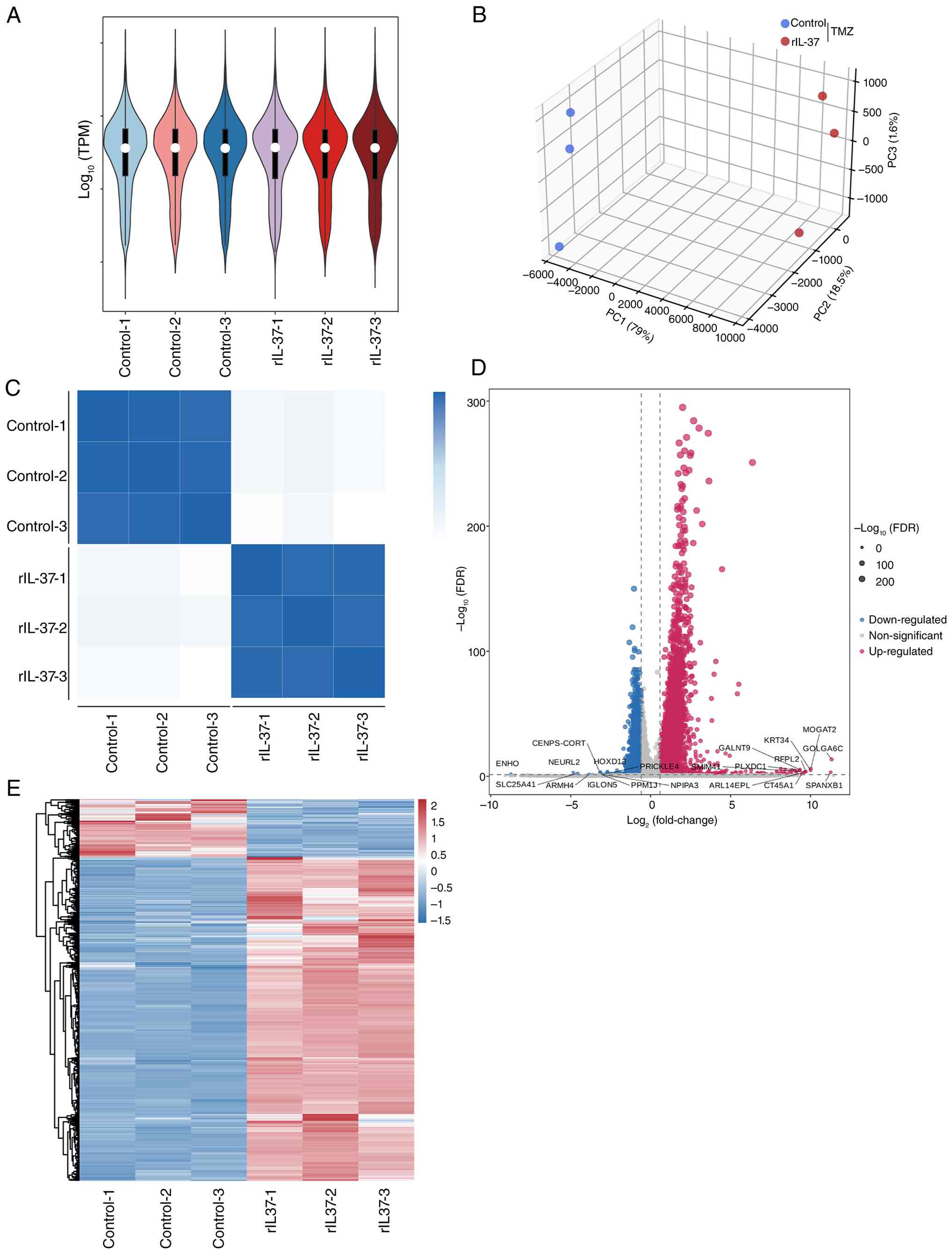 | Figure 3.RNA-seq analysis indicates greater
upregulated DEGs in rIL-37-treated U251 cells exposed to TMZ. (A)
Violin plot of RNA-seq data of control group (TMZ exposure alone)
and rIL-37 group (TMZ exposure and rIL-37 treatment). (B) Principal
components analysis 3D score plot of RNA-seq data of control group
and rIL-37 group. (C) The heatmap of RNA-seq data of control group
and rIL-37 group. The DEGs were identified with the threshold set
at |log2FC|>1 and FDR <0.05. (D) The distribution of DEGs
were shown by the volcano plot . (E) The distribution of DEGs were
shown as a heatmap. Data was expressed as mean ± SD. Statistical
significance was set at P<0.05. DEGs, differentially expressed
genes; FC, fold-change; FDR, false discovery rate, PC1, principal
component 1; PC2, principal component 2; PC3, principal component
3; RNA-seq, RNA sequencing; TMZ, temozolomide; TPM, transcripts per
million; rIL-37, recombinant human IL-37. |
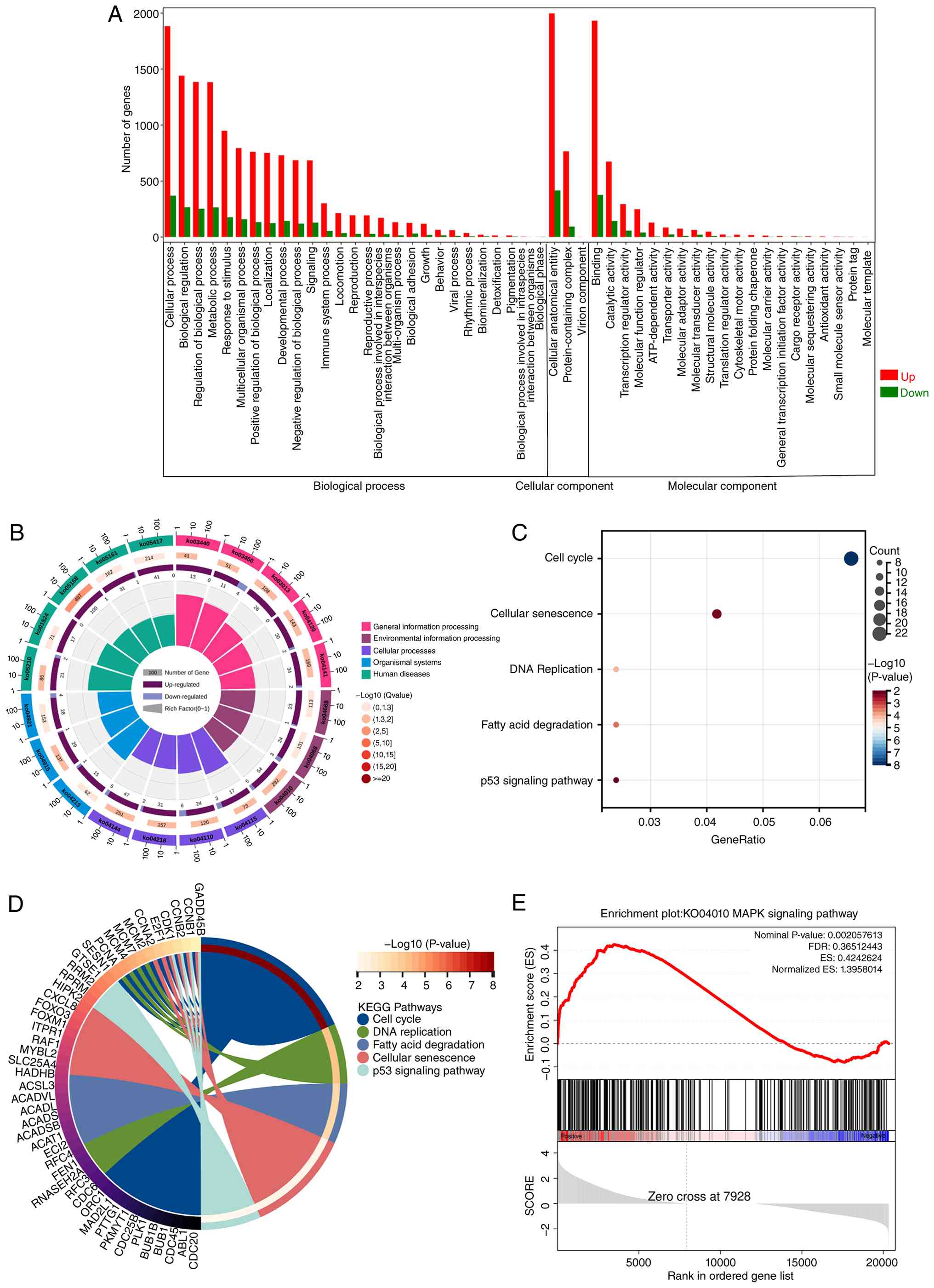
GO enrichment analysis revealed notable enrichment
of DEGs in cellular processes and biological regulation, suggesting
that rIL-37 treatment evoked multiple biological processes to
establish TMZ resistance in U251 cells (Fig. 4A). KEGG enrichment analysis further
indicated that upregulated DEGs were mainly clustered in the ‘cell
cycle’, ‘cellular senescence’, ‘DNA replication’, ‘fatty acid
degradation’ and the ‘p53 signaling pathway’. This implies the
potential involvement of senescence in rIL-37-induced TMZ
resistance in U251 cells (Fig. 4B and
C). Moreover, the chord diagram illustrated the specific DEGs
related to the top five enriched KEGG pathways (Fig. 4D). Gene set enrichment analysis
further demonstrated that the MAPK signaling pathway (ID, KO04010)
was markedly enriched between the rIL-37 and control groups
(Fig. 4E). Overall, the RNA-seq
results indicate the activation of MAPK signaling pathway and
senescence-related pathways in U251 cells undergoing TMZ exposure
and rIL-37 treatment, suggesting involvement in rIL-37-induced
TMZ.
rIL-37 activates the p38 MAPK pathway
and SA pathways in GBM cells under TMZ exposure
To validate the generalizability of the sequencing
findings, rIL-37 treatment also suppressed TMZ-induced apoptosis in
both U87 and A172 cell lines, as evidenced by a downregulation of
BAX and an upregulation of BCL-2 expression (Fig. 5A). RT-qPCR and ELISA assays
demonstrated that SASP components of U251/U87/A172 cells exposed to
TMZ, including IL-1β and IL-6, were induced by rIL-37 treatment
(Fig. 5B and C).
Senescence-associated DNA damage was identified by
immunofluorescence staining of γ-H2AX, and rIL-37 increased γ-H2AX
signals in the nuclei of rIL-37-treated U251/U87/A172 cells exposed
to TMZ (Fig. 5D and E). Western
blot analysis indicated that rIL-37 upregulated the expression of
senescence-related p21 and p16 (Fig.
5F). Meanwhile, rIL-37 treatment significantly activated the
MAPK pathway in TMZ-exposed U251/U87/A172 cells, which was marked
by increased phosphorylation of p38 MAPK (p-p38 MAPK) (Fig. 5G). To verify the direct activating
effect of rIL-37, siRNA was used to knock down the IL-37 receptor
IL-1R8 (SIGIRR) (Fig. 5H) and found
that SIGIRR knockdown effectively reversed rIL-37-induced MAPK
pathway activation (Fig. 5I). These
findings indicate that IL-37 potentiates TMZ resistance by
synchronously activating MAPK-driven survival signals and
senescence-associated secretory reprogramming.
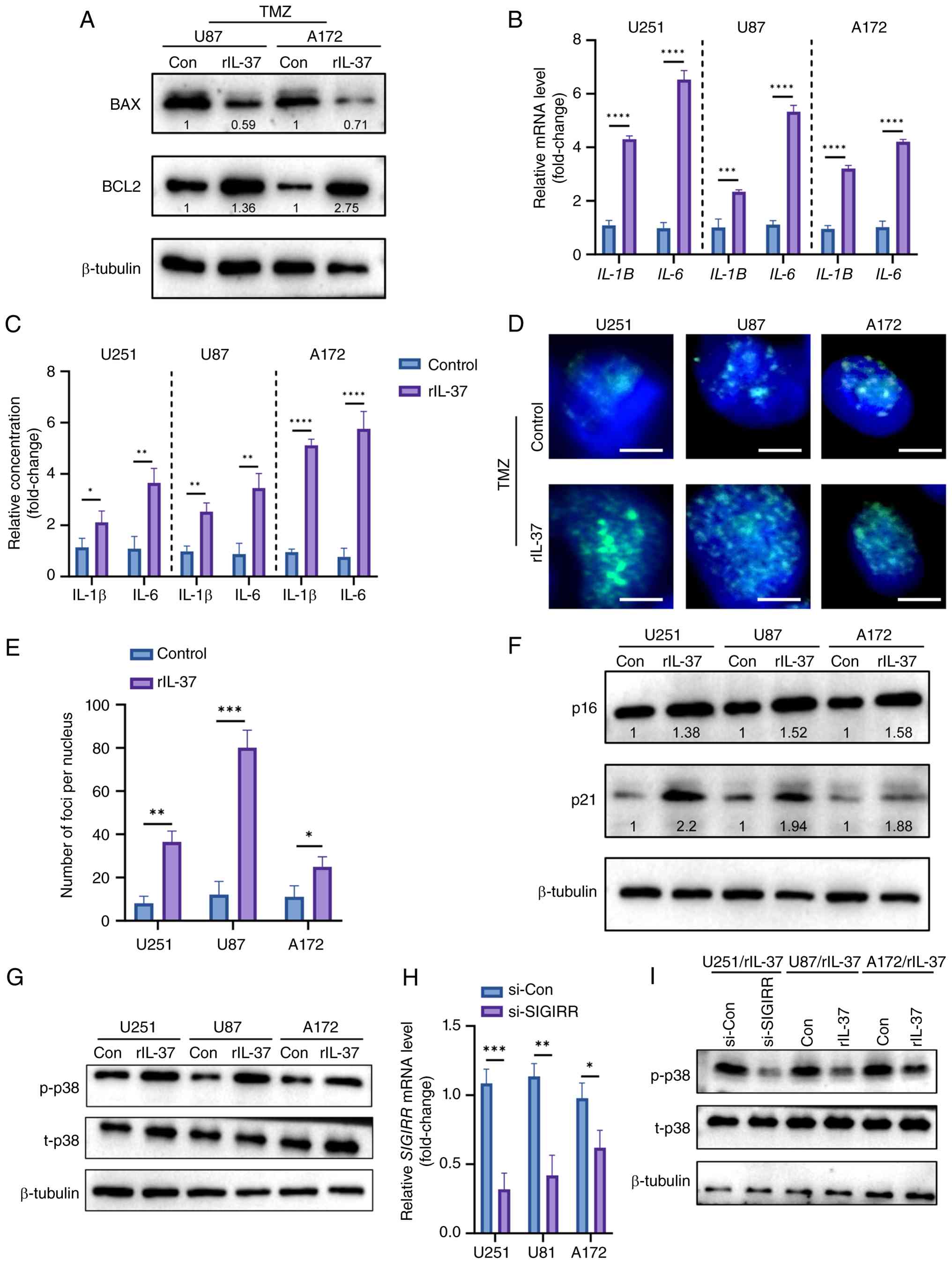 | Figure 5.rIL-37 activated p38 MAPK pathway and
senescence-associated pathways in glioblastoma cells. (A) The
expression of BAX and BCL2 in U87 and A172 cells exposed to TMZ
with or without rIL-37. (B) Quantitative-PCR analysis of SASP
factors IL-1β and IL-6 in TMZ-stimulated U251, U87 and A172 cells
with or without rIL-37 treatment (n=3). (C) ELISA analysis of SASP
factor IL-1β and IL-6 in TMZ-stimulated U251, U87 and A172 cells
with or without rIL-37 treatment (n=3). (D) γ-H2AX immunostaining
(scale bar, 5 µm) and (E) corresponding statistical data of
TMZ-stimulated U251, U87 and A172 cells with or without rIL-37
treatment. White bar represents 5 µm. (F) The expression of p21 and
p16 in U251, U87 and A172 cells. (G) p-p38 MAPK and t-p38 MAPK in
U251, U87 and A172 cells. (H) The mRNA expression of SIGIRR in
U251, U87 and A172 cells treated with si-SIGIRR. (I) The expression
of p-p38 MAPK and t-p38 MAPK in U251, U87 and A172 cells exposed to
TMZ with or without si-SIGIRR. Con, control; p-p38,
phosphorylated-p38; SASP, senescence-associated secretory
phenotype; TMZ, temozolomide; t-p38, total p38; MAPK,
mitogen-activated protein kinase; si, small interfering RNA;
rIL-37, recombinant human IL-37; γ-H2AX, γ-H2A histone family
member X. *P<0.05; **P<0.01; ***P<0.001;
****P<0.0001. |
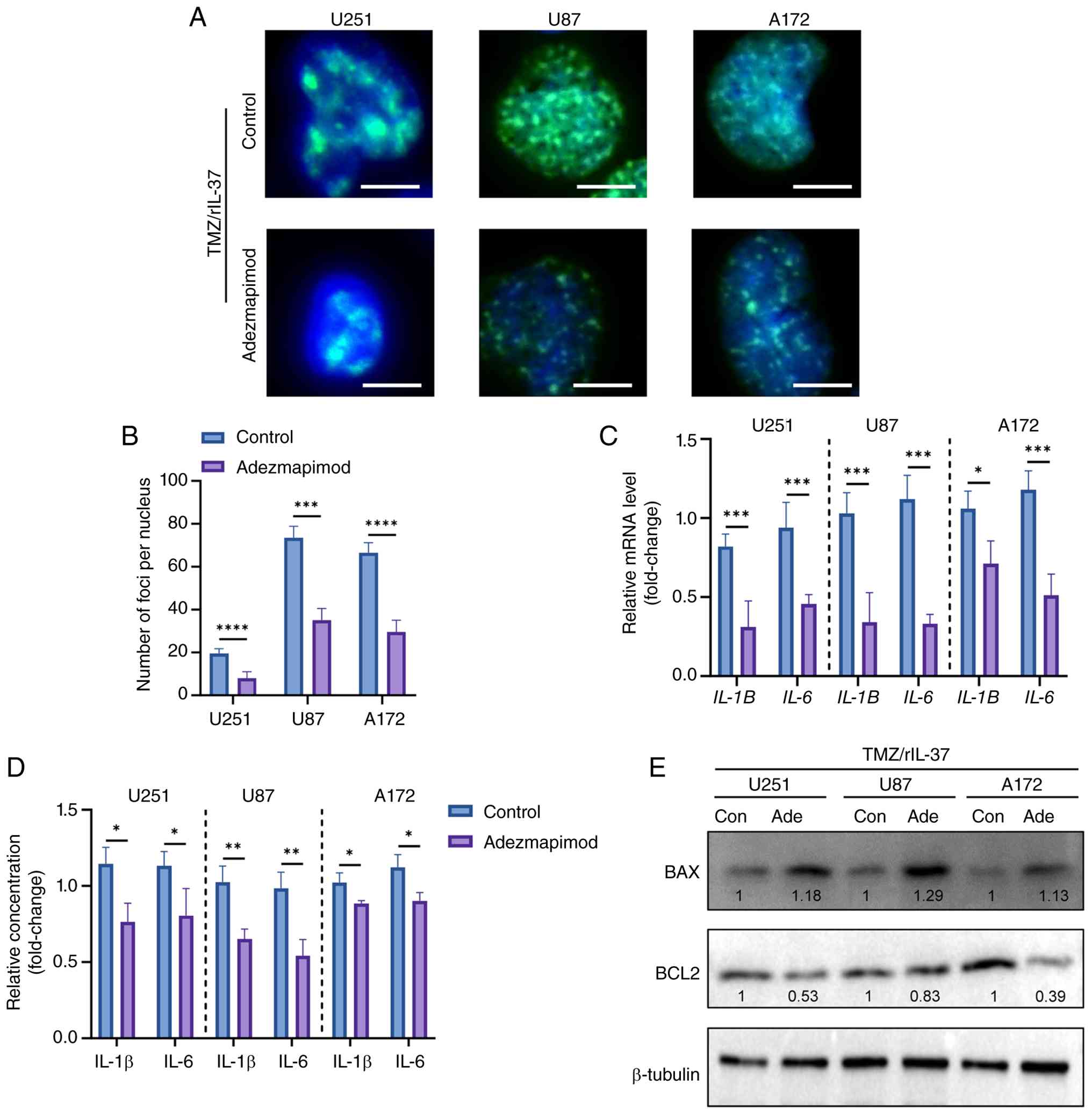
Inhibition of the MAPK pathway
reverses senescence and TMZ resistance in rIL-37-treated GBM
cells
To assess the importance of the MAPK signaling
pathway in rIL-37-induced TMZ resistance and senescence, the MAPK
signaling pathway was blocked using the inhibitor Adezmapimod.
γ-H2AX immunostaining (Fig. 6A and
B) and SASP factors (Fig. 6C and
D) were markedly attenuated by Adezmapimod in U251/U87/A172
cells. Consistently, the suppression of TMZ-induced apoptosis by
rIL-37 was reversed by Adezmapimod, as evidenced by the restoration
of pro-apoptotic BAX upregulation and the attenuation of
anti-apoptotic BCL2 expression in U251/U87/A172 cells (Fig. 6E). These findings demonstrate that
MAPK activation is indispensable for rIL-37-mediated TMZ resistance
and senescence reprogramming, providing a rationale for targeting
IL-37/MAPK axis to overcome therapeutic evasion.
Discussion
The results of the present study illustrate the
pro-survival roles of IL-37b, the current primary research target
among IL-37 splice variants (IL-37a-e), in GBM and explored the
underlying mechanisms. The major findings are summarized as
follows: First, the application of rIL-37 significantly increased
the survival rates of GBM U251 cells exposed to TMZ. Notably, the
present study also evaluated the functions of endogenous IL-37 and
revealed that IL-37 was negatively associated with the sensitivity
of U251 cells to TMZ; IL-37 overexpression significantly promoted
the survival of U251 cells upon TMZ exposure, whereas IL-37
knockdown enhanced TMZ sensitivity. Consistently, IL-37 knockdown
potentiated the sensitivity of U251-TR cells to TMZ. Secondly,
rIL-37 promoted the proliferative capacity of U251 cells, as
demonstrated by the increased colony formation of U251 cells
treated with rIL-37 and TMZ. Moreover, rIL-37 suppressed
TMZ-induced apoptotic responses in U251 cells by decreasing the
total apoptosis rate. Conversely, silencing IL-37 expression
significantly inhibited cellular proliferation and promoted the
activation of apoptotic responses in U251-TR cells upon TMZ
exposure. Thirdly, RNA sequencing results revealed significant
activation of the MAPK signaling pathway and senescence-related
pathways in TMZ-stimulated U251 cells treated with rIL-37 when
compared with those treated with TMZ alone. Finally, TMZ resistance
of U251 cells driven by rIL-37 depended on activation of the MAPK
signaling pathway, as the p38 MAPK inhibitor Adezmapimod markedly
reversed both rIL-37-induced TMZ resistance and cellular
senescence, and rIL-37-suppressed apoptosis of U251 cells under TMZ
exposure. In summary, the results of present study highlight the
pro-tumorigenic roles of IL-37 in GBM against TMZ cytotoxicity by
promoting proliferation and inhibiting apoptosis by activating MAPK
signaling.
Although TMZ is a well-recognized cornerstone of GBM
therapy, the features of high heterogeneity and aggressiveness of
GBM make it easy to develop chemoresistance to TMZ, contributing to
the poor prognosis of patients (29). The acquisition of TMZ resistance in
GBM cells involves multifaceted mechanisms, including enhanced DNA
repair capabilities, abnormal activation of pro-survival pathways
and formation of an immunosuppressive microenvironment (30). Research further highlights the roles
of nano strategy-enhanced drug delivery (31), oxygen-stimulated phenotypic
plasticity (32) and intercellular
adhesion molecule-1-mediated ATP binding cassette subfamily B
member 1 membrane assembly (33) in
modulating TMZ sensitivity. Therefore, the strategy of combining
TMZ and inhibitors targeting the pathways modulating the DNA damage
response (34), inflammatory
response (35) and immune
microenvironment (36) for GBM
therapy has become an important and promising method to improve
efficacy and overcome drug resistance.
Cytokines are a diverse group of proteins secreted
by immune or non-immune cells, including lymphocytes, macrophages
and tumor cells, to orchestrate intercellular communication to
regulate immune responses, cellular proliferation, differentiation
and inflammation by binding to specific receptors (37–39).
Cytokine family proteins serve crucial roles in regulating the
proliferation, invasion and treatment resistance of tumor cells
through complex signaling networks (40). For instance, IL-1β derived from
cancer-associated fibroblasts (CAFs) not only acts on CAFs to form
the fibrotic tumor microenvironment but also stimulates pancreatic
ductal adenocarcinoma cells to proliferate, survive and develop
chemoresistance (41). Similarly,
cytokine family members are closely involved in gliomagenesis,
progression and TMZ resistance through multifaceted signaling
pathways (9,42,43).
The pro-inflammatory cytokine IL-17 has been reported to have
pro-tumorigenic functions, as it enhances the proliferative and
migratory capabilities of glioma cells through the PI3K/Akt1/NF-κB
pathway (44). IL-10 promotes the
immune escape of gliomas as IL-10 released by myeloid cells results
in the dysfunction of T cells and the final formation of an
immunosuppressive microenvironment (13). Additionally, TMZ-resistant glioma
cells have also been reported to be capable of releasing IL-10,
which together with TGF-β, drive the formation of an
immunosuppressive microenvironment to assist in the invasion of
glioma cells (36).
The present study focused on the biological
functions of IL-37 in the sensitivity of GBM cells to TMZ. IL-37,
an anti-inflammatory cytokine of the IL-1 family, serves an
important role in shaping the tumor microenvironment and displays
several functions in different tumors. IL-37 can drive tumor
progression by fostering an immunosuppressive microenvironment and
suppressing CD8+ T cell activity in colorectal cancer
(27). Conversely, IL-37 can
inhibit tumorigenesis by inhibiting pro-inflammatory signals and
enhance antitumor immunity by inducing the M1 macrophage phenotype
and dendritic cell maturation in non-small cell lung cancer
(45). The present study revealed
that rIL-37 treatment markedly reduced the sensitivity of U251
cells to TMZ, which may be explained by the enhanced proliferation
and suppressed apoptosis of GBM cells mediated by rIL-37. As the
complexity of the microenvironment consisting of tumor cells,
fibroblasts and immune cells renders the sources of IL-37 diverse,
IL-37 in the immune microenvironment may come from tumor cells
themselves, in addition to classic sources such as TAMs or Tregs
(46,47). The present study then explored the
roles of endogenous IL-37 in GBM development and chemoresistance
and demonstrated that IL-37 knockdown significantly enhanced the
sensitivity of both U251 cells and U251-TR cells to TMZ, as shown
by decreased survival rates of the tumor cells exposed to TMZ. By
contrast, IL-37 overexpression rendered U251 cells more resistant
to TMZ. Furthermore, IL-37 knockdown markedly attenuated
proliferation and exacerbated TMZ-induced apoptosis in U251-TR
cells. These results highlight the pro-survival effects of both
intracellular and extracellular IL-37 on GBM cells.
RNA-seq analysis was used to explore the mechanisms
underlying the pro-survival role of rIL-37 in GBM. Enrichment
analysis of DEGs revealed that rIL-37 treatment notably activated
MAPK and senescence-related pathways in U251 cells exposed to TMZ.
MAPK signaling has been reported to serve a central role in
malignant progression and drug resistance in several tumors
(48,49); for instance, the mutated and
persistently activated NRAS/MAPK signaling pathway in melanoma
serves as the critical target of long non-coding RNA T-RECS to
inhibit apoptosis and tumor growth (50). Therefore, combined interventions
targeting this pathway, such as synergistic treatment with MEK
inhibitors and immunotherapy (51–53),
may become a new strategy to improve the prognosis of patients.
After revealing the activation of the MAPK signaling pathway with
elevated p-p38 MAPK, the present study then evaluated the necessity
of MAPK cascades in rIL-37-induced TMZ-resistance in U251 cells and
demonstrated that inhibiting MAPK pathways reversed drug
resistance, as shown by reduced survival rates and enhanced
apoptotic responses of U251 cells under TMZ exposure. This
indicates that activated MAPK pathways may be essential for
rIL-37-induced therapeutic resistance. The induced
senescence-related pathways primarily manifest as activation of p21
and p16-Rb signaling axes, leading to permanent cell cycle arrest,
which serves complex roles in tumor progression, including limiting
malignant proliferation and facilitating immune evasion and
acquired drug resistance (54). The
present study observed significant upregulation of p21 and p16
proteins and elevation of SASP factors, including IL-1β and IL-6,
and increased SA-β-galactosidase staining in U251 cells treated
with TMZ and rIL-37. Cellular SA DNA damage was confirmed by γ-H2AX
staining. Although blocking cellular senescence was previously
proposed to inhibit tumor proliferation (55,56),
senescent cells have also been reported to promote prostate cancer
progression through SASP reprogramming induced by deletion of the
metalloproteinase inhibitor, tissue inhibitor of metalloproteinases
1 (57,58). The present study also demonstrated
that Adezmapimod, a p38 MAPK inhibitor, abolished the activation of
senescence markers p16 and p21 and SASP factors, as well as
enhanced positive rates of both SA-β-galactosidase and γ-H2AX
staining of U251 cells receiving TMZ and rIL-37. This suggests that
cellular senescence may be involved in rIL-37-induced TMZ
resistance in GBM.
It should be noted that the present study has
certain limitations. First, although the results demonstrated that
activated MAPK cascade signaling is indispensable for
rIL-37-induced TMZ resistance, the pleiotropic and
context-dependent nature of cytokines complicates therapeutic
targets, and a heterogeneous tumor microenvironment may also
critically limit cytokine efficacy. This makes the presence of
other potential targets for IL-37 undeniable. Second, given that
the diverse receptors on cell membranes are the core basis of
cytokines to exert different biological functions, and the highly
heterogeneous feature of gliomas inevitably leads to spatially
varied expression of cytokine receptors across glioma subclones,
elucidating the well-identified receptors of rIL-37 such as IL-18Rα
(59) and IL-1R8 (60) in other tumors to guarantee treatment
predictability of GBM should be the focus of future work. Third,
the redundancy of cytokine signaling networks and the activation of
compensatory pathways often compromise therapeutic outcomes. For
instance, compensatory activation of NF-κB downstream pathways may
undermine the therapeutic efficacy of C-X-C motif chemokine
ligand-12/C-X-C chemokine receptor type 4 in enhancing TMZ
sensitivity (61). The activated
senescence-related pathways in TMZ-stimulated U251 cells with
rIL-37 treatment may be a compensatory downstream pathway of MAPK
signaling activation, and its roles in TMZ resistance or functional
associations with MAPK cascades in GBM cells should also be the
focus of subsequent studies.
In conclusion, the results of the present study
indicate that the anti-inflammatory cytokine IL-37 enhanced TMZ
resistance in GBM cells by activating MAPK signaling to maintain
cellular anti-apoptotic capacity and induce cellular
senescence-related pathways. This highlights the IL-37-MAPK axis as
a critical mediator of tumorigenesis and therapeutic resistance in
GBM. Therefore, the strategy of integrating TMZ with targeted
inhibition of MAPK cascades is promising for GBM therapy in the
clinic.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the Qiqihar
Science and Technology Tackling Project (grant no.
LSFGG-2023013).
Availability of data and materials
The data generated in the present study may be found
in the ScienceDB database using the following URL: https://doi.org/10.57760/sciencedb.32976.
Authors' contributions
YW, YL and JW performed the formal analysis. WR and
XS performed data curation (organising, cleaning and formatting),
analysis and interpretation of data. YW, YL and XS wrote the
original draft. YL was responsible for validation, investigation,
project administration and funding acquisition. YW and YL confirm
the authenticity of all the raw data. All authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lapointe S, Perry A and Butowski NA:
Primary brain tumours in adults. Lancet. 392:432–446. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Le Calvez K, Mauricaite R, Treasure P,
Booth TC, Price SJ, Brodbelt A, Gregory JJ, Dadhania S,
Pakzad-Shahabi L, Dumba M, et al: Adult glioblastoma in England:
Incidence, treatment, and outcomes with novel population-based
strata. Cancer Epidemiol. 97:1028112025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang M, Shen S, Hou F and Yan Y:
Pathophysiological roles of integrins in gliomas from the
perspective of glioma stem cells. Front Cell Dev Biol.
10:9624812022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krivosheya D, Prabhu SS, Weinberg JS and
Sawaya R: Technical principles in glioma surgery and preoperative
considerations. J Neurooncol. 130:243–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karachi A, Dastmalchi F, Mitchell DA and
Rahman M: Temozolomide for immunomodulation in the treatment of
glioblastoma. Neuro Oncol. 20:1566–1572. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iturrioz-Rodríguez N, Sampron N and Matheu
A: Current advances in temozolomide encapsulation for the
enhancement of glioblastoma treatment. Theranostics. 13:2734–2756.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Vieito M, Simonelli M, de Vos F, Moreno V,
Geurts M, Lorenzi E, Macchini M, van den Bent MJ, Del Conte G, de
Jonge M, et al: Trotabresib (CC-90010) in combination with adjuvant
temozolomide or concomitant temozolomide plus radiotherapy in
patients with newly diagnosed glioblastoma. Neurooncol Adv.
14:vdac1462022.PubMed/NCBI
|
|
8
|
Tomar MS, Kumar A, Srivastava C and
Shrivastava A: Elucidating the mechanisms of Temozolomide
resistance in gliomas and the strategies to overcome the
resistance. Biochim Biophys Acta Rev Cancer. 1876:1886162021.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee GA, Hsu JB, Chang YW, Hsieh LC, Li YT,
Wu YC, Chu CY, Chiang YH, Guo WY, Wu CC, et al: IL-19 as a
promising theranostic target to reprogram the glioblastoma
immunosuppressive microenvironment. J Biomed Sci. 32:342025.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ji H, Lan Y, Xing P, Wang Z, Zhong X, Tang
W, Wei Q, Chen H, Liu B and Guo H: IL-18, a therapeutic target for
immunotherapy boosting, promotes temozolomide chemoresistance via
the PI3K/AKT pathway in glioma. J Transl Med. 22:9512024.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu S, Ba Y, Li C, Xing M, Zhang T, Liu Y,
Gao Y and Xu G: Interleukin 37 inhibits the migration and invasion
of Glioma cells. Biotechnol Genet Eng Rev. 40:926–942. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Luo C, Shu Y, Luo J, Liu D, Huang DS, Han
Y, Chen C, Li YC, Zou JM, Qin J, et al: Intracellular IL-37b
interacts with Smad3 to suppress multiple signaling pathways and
the metastatic phenotype of tumor cells. Oncogene. 36:2889–2899.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ravi VM, Neidert N, Will P, Joseph K,
Maier JP, Kückelhaus J, Vollmer L, Goeldner JM, Behringer SP,
Scherer F, et al: T-cell dysfunction in the glioblastoma
microenvironment is mediated by myeloid cells releasing
interleukin-10. Nat Commun. 13:9252022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barthel L, Hadamitzky M, Dammann P,
Schedlowski M, Sure U, Thakur BK and Hetze S: Glioma: Molecular
signature and crossroads with tumor microenvironment. Cancer
Metastasis Rev. 41:53–75. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang M, Zhao Y, Yu ZY, Zhang RD, Li SA,
Zhang P, Shan TK, Liu XY, Wang ZM, Zhao PC and Sun HW: Glioma
exosomal microRNA-148a-3p promotes tumor angiogenesis through
activating the EGFR/MAPK signaling pathway via inhibiting ERRFI1.
Cancer Cell Int. 20:5182020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang W and Liu HT: MAPK signal pathways
in the regulation of cell proliferation in mammalian cells. Cell
Res. 12:9–18. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hagemann C and Blank JL: The ups and downs
of MEK kinase interactions. Cell Signal. 13:863–875. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu YR and Lei CQ: TAK1-TABs Complex: A
central signalosome in inflammatory responses. Front Immunol.
11:6089762020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qi XM and Chen G: p38γ mapk inflammatory
and metabolic signaling in physiology and disease. Cells.
12:16742023. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Simoes AE, Rodrigues CM and Borralho PM:
The MEK5/ERK5 signalling pathway in cancer: A promising novel
therapeutic target. Drug Discov Today. 21:1654–1663. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ammar UM, Abdel-Maksoud MS and Oh CH:
Recent advances of RAF (rapidly accelerated fibrosarcoma)
inhibitors as anti-cancer agents. Eur J Med Chem. 158:144–166.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Houles T, Lavoie G, Nourreddine S, Cheung
W, Vaillancourt-Jean É, Guérin CM, Bouttier M, Grondin B, Lin S,
Saba-El-Leil MK, et al: CDK12 is hyperactivated and a
synthetic-lethal target in BRAF-mutated melanoma. Nat Commun.
13:64572022. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sevin M, Debeurme F, Laplane L, Badel S,
Morabito M, Newman HL, Torres-Martin M, Yang Q, Badaoui B,
Wagner-Ballon O, et al: Cytokine-like protein 1-induced survival of
monocytes suggests a combined strategy targeting MCL1 and MAPK in
CMML. Blood. 137:3390–3402. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu P, Zhang G, Hou S and Sha LG: MAPK8
mediates resistance to temozolomide and apoptosis of glioblastoma
cells through MAPK signaling pathway. Biomed Pharmacother.
106:1419–1427. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zou Y, Xu L, Wang W, Zhu X, Lin J, Li H,
Chen J, Xu W, Gao H, Wu X, et al: Muscone restores anoikis
sensitivity in TMZ-resistant glioblastoma cells by suppressing
TOP2A via the EGFR/Integrin beta1/FAK signaling pathway.
Phytomedicine. 129:1557142024. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Z, Zeng FL, Hu YW, Wang XY, Zhao FL,
Zhou P, Hu J, Xiao YY, Hu ZL, Guo MF, et al: Interleukin-37
promotes colitis-associated carcinogenesis via SIGIRR-mediated
cytotoxic T cells dysfunction. Signal Transduct Target Ther.
7:192022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gu M, Jin Y, Gao X, Xia W, Xu T and Pan S:
Novel insights into IL-37: An anti-inflammatory cytokine with
emerging roles in anti-cancer process. Front Immunol.
14:12785212023. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lan Z, Li X and Zhang X: Glioblastoma: An
update in pathology, molecular mechanisms and biomarkers. Int J Mol
Sci. 25:30402024. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang Q, Ren T, Bai P, Wang X, Zhao L,
Zhong R and Sun G: Novel strategies to overcome chemoresistance in
human glioblastoma. Biochem Pharmacol. 230:1165882024. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lan Y, Li X, Liu B, Lu J, Zuo B, Wang Y,
Cao S, Fu X, Yue Q, Luo X, et al: Framework nucleic acid-based
nanoparticles enhance temozolomide sensitivity in glioblastoma.
Drug Resist Updat. 76:1011222024. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ma K, Wang S, Ma Y, Zeng L, Xu K, Mu N,
Lai Y, Shi Y, Yang C, Chen B, et al: Increased oxygen stimulation
promotes chemoresistance and phenotype shifting through PLCB1 in
gliomas. Drug Resist Updat. 76:1011132024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang X, Tan Y, Li T, Tan D, Fu B, Yang M,
Chen Y, Cao M, Xuan C, Du Q, et al: Intercellular adhesion
molecule-1 suppresses TMZ chemosensitivity in acquired
TMZ-resistant gliomas by increasing assembly of ABCB1 on the
membrane. Drug Resist Updat. 76:1011122024. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hanisch D, Krumm A, Diehl T, Stork CM,
Dejung M, Butter F, Kim E, Brenner W, Fritz G, Hofmann TG and Roos
WP: Class I HDAC overexpression promotes temozolomide resistance in
glioma cells by regulating RAD18 expression. Cell Death Dis.
13:2932022. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang W, Zhong Z, Luo C, Xiao Y, Li L,
Zhang X, Yang L, Xiao K, Ning Y, Chen L, et al: The
miR-26a/AP-2α/Nanog signaling axis mediates stem cell self-renewal
and temozolomide resistance in glioma. Theranostics. 9:5497–5516.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li X, Cheng Y, Yang Z, Ji Q, Huan M, Ye W,
Liu M, Zhang B, Liu D and Zhou S: Glioma-targeted
oxaliplatin/ferritin clathrate reversing the immunosuppressive
microenvironment through hijacking Fe2+ and boosting Fenton
reaction. J Nanobiotechnology. 22:932024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li L, Yu R, Cai T, Chen Z, Lan M, Zou T,
Wang B, Wang Q, Zhao Y and Cai Y: Effects of immune cells and
cytokines on inflammation and immunosuppression in the tumor
microenvironment. Int Immunopharmacol. 88:1069392020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kaminska P, Tempes A, Scholz E and Malik
AR: Cytokines on the way to secretion. Cytokine Growth Factor Rev.
79:52–65. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Opal SM and DePalo VA: Anti-inflammatory
cytokines. Chest. 117:1162–1172. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kureshi CT and Dougan SK: Cytokines in
cancer. Cancer Cell. 43:15–35. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang D, Li L, Jiang H, Li Q, Wang-Gillam
A, Yu J, Head R, Liu J, Ruzinova MB and Lim KH: Tumor-stroma
IL1β-IRAK4 feedforward circuitry drives tumor fibrosis,
chemoresistance, and poor prognosis in pancreatic cancer. Cancer
Res. 78:1700–1712. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zuo M, Zhang S, Chen S, He Y, Li J, Xiang
Y, Yuan Y, Li T, Yang W, Wang Z, et al: Cytokine CCL2 secreted by
cancer-associated fibroblasts augments temozolomide resistance in
glioblastoma through ERK1/2 signaling. Oncogene. 44:4657–4670.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wan W, Wang P, Liao B, Zhao L, Gong S, Wei
L and Wu N: Single-cell mapping of TMZ-resistant glioma reveals
CXCL2/3-CXCR2-associated neutrophil communication. Biochem Biophys
Res Commun. 785:1527062025. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang B, Zhao CH, Sun G, Zhang ZW, Qian BM,
Zhu YF, Cai MY, Pandey S, Zhao D, Wang YW, et al: IL-17 induces the
proliferation and migration of glioma cells through the activation
of PI3K/Akt1/NF-κB-p65. Cancer Lett. 447:93–104. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang J, Wise SG, Zuo S, Bao S and Zhang
X: The distinct roles of IL-37 and IL-38 in non-small cell lung
carcinoma and their clinical implications. Front Immunol.
16:15643572025. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mei Y, Zhu Y, Yong KSM, Hanafi ZB, Gong H,
Liu Y, Teo HY, Hussain M, Song Y, Chen Q and Liu H: IL-37 dampens
immunosuppressive functions of MDSCs via metabolic reprogramming in
the tumor microenvironment. Cell Rep. 43:1138352024. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mei Y, Zhu Y, Teo HY, Liu Y, Song Y, Lim
HY, Binte Hanafi Z, Angeli V and Liu H: The indirect antiangiogenic
effect of IL-37 in the tumor microenvironment. J Leukoc Biol.
107:783–796. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee S, Rauch J and Kolch W: Targeting MAPK
signaling in cancer: Mechanisms of drug resistance and sensitivity.
Int J Mol Sci. 21:11022020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nussinov R, Tsai CJ and Jang H: Anticancer
drug resistance: An update and perspective. Drug Resist Updat.
59:1007962021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Feichtenschlager V, Chen L, Zheng YJ, Ho
W, Sanlorenzo M, Vujic I, Fewings E, Lee A, Chen C, Callanan C, et
al: The therapeutically actionable long non-coding RNA ‘T-RECS’ is
essential to cancer cells' survival in NRAS/MAPK-driven melanoma.
Mol Cancer. 23:402024. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhao K, Dai Q, Wu J, Wei Z, Duan Y and
Chen B: Morusin enhances the antitumor activity of MAPK pathway
inhibitors in BRAF-mutant melanoma by inhibiting the feedback
activation of STAT3. Eur J Cancer. 165:58–70. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhao K, Lu Y, Chen Y, Cheng J and Zhang W:
Dual inhibition of MAPK and JAK2/STAT3 pathways is critical for the
treatment of BRAF mutant melanoma. Mol Ther Oncolytics. 18:100–108.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Khaliq M, Manikkam M, Martinez ED and
Fallahi-Sichani M: Epigenetic modulation reveals differentiation
state specificity of oncogene addiction. Nat Commun. 12:15362021.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wagner KD and Wagner N: The senescence
markers p16INK4A, p14ARF/p19ARF, and p21 in organ development and
homeostasis. Cells. 11:19662022. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen Q, Sun X, Luo X, Wang J, Hu J and
Feng Y: PIK3R3 inhibits cell senescence through p53/p21 signaling.
Cell Death Dis. 11:7982020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wu T, Yang Z, Chen W, Jiang M, Xiao Z, Su
X, Jiao Z, Yu Y, Chen S, Song M and Yang A: miR-30e-5p-mediated
FOXD1 promotes cell proliferation by blocking cellular senescence
and apoptosis through p21/CDK2/Rb signaling in head and neck
carcinoma. Cell Death Discov. 9:2952023. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Guccini I, Revandkar A, D'Ambrosio M,
Colucci M, Pasquini E, Mosole S, Troiani M, Brina D,
Sheibani-Tezerji R, Elia AR, et al: Senescence reprogramming by
TIMP1 deficiency promotes prostate cancer metastasis. Cancer Cell.
39:68–82.e9. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Da Silva-Alvarez S and Collado M: The
Jekyll and Hyde of senescence in cancer: TIMP1 controls the switch
from Tumor-controlling to tumor-promoting senescence. Cancer Cell.
39:13–15. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Wang D, Zhang B, Liu X, Kan LL, Leung PC
and Wong CK: Agree to disagree: The contradiction between IL-18 and
IL-37 reveals shared targets in cancer. Pharmacol Res.
200:1070722024. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Landolina N, Mariotti FR, Pelosi A, D'Oria
V, Ingegnere T, Alicata C, Vacca P, Moretta L and Maggi E: The
anti-inflammatory cytokine IL-37 improves the NK cell-mediated
anti-tumor response. Oncoimmunology. 13:22975042024. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Chiang IT, Liu YC, Liu HS, Ali AAA, Chou
SY, Hsu TI and Hsu FT: Regorafenib reverses Temozolomide-induced
CXCL12/CXCR4 signaling and triggers apoptosis mechanism in
glioblastoma. Neurotherapeutics. 19:616–634. 2022. View Article : Google Scholar : PubMed/NCBI
|