Introduction
Most cancer-related deaths (~90%) are caused by
metastatic cancer (1), which has
become a notable obstacle to effective cancer treatment. Metastasis
begins when malignant cells detach from the primary tumor,
disseminate through the lymphatic system, bloodstream or body
cavities and colonize distant organs (2,3). This
multistep process is influenced by the cancer microenvironment,
which can either restrain or promote the metastatic potential of
tumor cells (4). Once tumor cells
enter the circulation, they are referred to as circulating tumor
cells (CTCs), which serve a key role in the establishment of
distant metastases (5).
Although platelets are primarily responsible for
maintaining hemostasis, growing evidence indicates they actively
participate in cancer progression (6,7). CTCs
can activate platelets and form tumor cell-platelet aggregates,
which shield tumor cells from immune surveillance and enhance their
survival in the bloodstream. In addition, activated platelets
adhere to vascular endothelial cells, facilitating the attachment
of these aggregates to the vessel wall. This promotes tumor cell
extravasation through the endothelial barrier and basement
membrane, leading to the formation of secondary tumors (8). Consequently, interactions between CTCs
and platelets are a critical driving force in cancer metastasis
(7).
The vascular endothelium also serves a key role in
the process of tumor metastasis. During CTC adhesion, endothelial
cells upregulate surface adhesion molecules in response to
stimulation, thereby strengthening tumor cell attachment (9). This initiates a cascade of events such
as endothelial cytoskeletal rearrangement, increased endothelial
permeability and basement membrane degradation, all of which
promotes tumor cell transmigration and metastatic colonization. Our
previous studies demonstrated that adhesion of CTCs to the vascular
endothelium is a crucial early step in distant metastasis and that
disrupting this interaction can markedly suppress metastatic spread
(4,5,10).
Despite these findings, effective clinical strategies targeting CTC
adhesion and metastasis remain limited, highlighting the need to
identify novel therapeutic agents and targets.
Salvia miltiorrhiza, a traditional Chinese
herb first documented in Shennong's Classic of Herbal Medicine, has
been used to promote blood circulation (11). Clinically, it has therapeutic
benefits in the treatment of stroke (12), cardiovascular disease and
osteoporosis (13). Tanshinone IIA
(Tan IIA), a major bioactive compound derived from S.
miltiorrhiza, exhibits a wide range of pharmacological
activities, including anti-inflammatory, antioxidant and antitumor
effects (14–19). Tan IIA is applied in the management
of cardiovascular and cerebrovascular disorders (20) due to its vasodilatory,
anticoagulant, antithrombotic and endothelial-protective properties
(21–25).
Tan IIA can reduce vascular oxidative stress,
inhibit platelet aggregation and protect endothelial function
(26), suggesting its potential to
modulate platelet-endothelium interactions. Tan IIA has also
demonstrated therapeutic potential in metabolic disorders such as
diabetes (27,28). Tan IIA exerts antitumor effects
across multiple cancer types (29,30).
Tan IIA has been shown to suppress tumor cell proliferation,
migration and metastasis by regulating cytoskeletal dynamics
(31), inhibiting oncogenic
signaling pathways (32) and
inducing ferroptosis (33). These
effects have been reported in liver, lung, breast, prostate
(34) and colorectal cancer
(35–37). However, the role of Tan IIA in
regulating interactions between CTCs, platelets and endothelial
cells remains poorly understood.
The present study investigated the effects of Tan
IIA at low concentrations on platelets and endothelial cells using
both in vivo and in vitro models. The aim was to
elucidate how Tan IIA modulates platelet-tumor cell interactions
and tumor cell adhesion to the vascular endothelium, thereby
providing mechanistic insight into its potential to prevent
CTC-mediated metastasis.
Materials and methods
Cell culture
The human non-small cell lung cancer cell A549,
human breast cancer MCF-7 and mouse breast cancer 4T1 cells were
obtained from the Cell Bank of the Chinese Academy of Sciences.
A549 cells were cultured in RPMI-1640 medium (cat. no. SH30027.01;
Hyclone; Cytiva) supplemented with 10% fetal bovine serum (FBS;
cat. no. A5256701; Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 µg/ml streptomycin. MCF-7 and 4T1 cells were
cultured in high-glucose DMEM (cat. no. SH30243.01; Hyclone;
Cytiva) supplemented with 10% FBS, 100 U/ml penicillin and 100
µg/ml streptomycin. Human umbilical vein endothelial cells (HUVECs)
were isolated as previously described (38) and cultured in 1% gelatin-coated
flasks using endothelial cell medium (cat. no. #1001; ScienCell
Research Laboratories, Inc.) supplemented with 5% FBS, 100 µg/ml
endothelial cell growth supplement (cat. no. #1052; ScienCell
Research Laboratories, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin. All cells were maintained at 37°C in a humidified
incubator containing 5% CO2. Cells were harvested using
0.25% trypsin prior to experiments. HUVECs were used within six
passages.
Drug preparation
Tan IIA (purity, ≥98%, cat. no. S107694; lot no.
K2208491) was obtained from Shanghai Aladdin Biochemical Technology
Co., Ltd. For the in vitro experiments, Tan IIA was
dissolved in DMSO to a stock solution of 40 mM, filtered, stored at
4°C and diluted with normal saline when used.
Cell viability assay
The cytotoxicity of Tan IIA was evaluated using the
MTT assay as previously described (4). Briefly, MCF-7, A549 and HUVEC cells
were seeded into 96-well plates at a density of 1×104
cells per well and incubated at 37°C for 24 h. The cells were
treated with medium containing various concentrations (0, 5, 10 and
20 µM) of Tan IIA at 37°C for 24 h. MTT solution was added,
followed by incubation for 4 h at 37°C. The medium was removed and
formazan crystals were dissolved in 100 µl DMSO. Absorbance was
measured at 570 nm using a microplate reader (Tecan Group Ltd.;
M200 PRO). All absorbance values were normalized to blank wells
containing culture medium and MTT reagent only, which served as the
zero metabolic activity baseline. Cell viability was expressed as a
percentage relative to untreated controls.
Cell cycle and apoptosis assay
A549 and HUVEC cells in logarithmic growth phase
were treated with Tan IIA (0, 5, 10 and 20 µM) at 37°C for 24 h and
collected using trypsin without EDTA. For cell cycle analysis, A549
cells were fixed in 70% ethanol at 4°C for 12 h, treated with RNase
and stained with PI. For apoptosis analysis (early and late
apoptotic cells), treated cells (A549 and HUVEC cells) were
resuspended in binding buffer and stained with 5 µl of Annexin
V-FITC and 5 µl of PI for 15 min in the dark at room temperature,
followed by the addition of 0.5 ml PBS. Cell cycle and apoptosis
were detected by flow cytometry with a BD FACSAria (BD Biosciences)
and analyzed by FlowJo software (version number 10.8.1; BD
Biosciences).
Wound healing assay
Cell migration was evaluated using a wound healing
assay. A549 cells were seeded into 12-well plates (2×105
cells/well) and allowed to form a confluent (80–90%) monolayer. A
linear scratch was created using a pipette tip and detached cells
were removed with serum-free RPMI-1640. Cells were incubated in
RPMI-1640 containing 1% FBS and Tan IIA at the indicated
concentrations (0, 5, 10 and 20 µM). Images were captured at 0 and
24 h using a light microscope. The migration of the cells was
assessed by measuring the width of the wound area.
Cell invasion assay
Cell invasion assay was assessed using
Matrigel-coated Transwell chambers (Corning, Inc.; 8 µm pore size)
as previously described (39). The
membrane was coated with 0.5% Matrigel overnight at 37°C. A549
cells (4×104) were seeded into the upper chamber in serum-free
RPMI-1640 medium containing Tan IIA (0, 10 and 20 µM), while the
lower chamber contained RPMI-1640 medium with 10% FBS. After 24 h
of incubation at 37°C in a cell culture incubator, non-invading
cells were removed and invading cells were fixed with 4%
paraformaldehyde for 20 min at 4°C and stained with 0.1% crystal
violet at room temperature for 30 min. Images of the stained cells
were captured using a light microscope. The invasive ability was
quantified by counting the number of cells that had penetrated
through the membrane and analyzed using ImageJ software (version
number 1.54p; National Institutes of Health).
Flow cytometric analysis of cell
adhesion molecules
A549 cells were treated with different
concentrations of Tan IIA (0, 5, 10 and 20 µM) at 37°C for 24 h,
and HUVECs were treated with 0, 5 and 10 µM of Tan IIA at 37°C for
24 h. Prior to treatment, HUVECs were stimulated with TNF-α (10
ng/ml) at 37°C for 4 h (negative control, HUVECs without TNF-α).
Subsequently, the cells were harvested, resuspended in PBS and
incubated in the dark at room temperature for 10 min with 10 µl
antibodies as follows: Anti-CD29 (PE-labeled, cat. no. 303004) for
A549 cells and anti-ICAM (cat. no. 353112) or anti-E-selectin (both
APC-labeled, cat. no. 336011) (all 1:100, all Biolegend, Inc.) for
HUVECs. After washing with PBS buffer, cells were analyzed using a
BD FACSAria and FlowJo software.
Platelet activity assay
Blood was collected from 5 volunteers at the Blood
Bank of Fujian Medical University Union Hospital (Fuzhou, China)
between May and October 2023. The study included 2 men and 3 women,
aged 20–40 years. The inclusion criteria were as follows: i) Normal
platelet aggregation function; ii) normal coagulation parameters
and other complete blood count indices; and iii) no use of any
anticoagulants for 2 weeks prior to blood collection. All
participants provided written informed consent before sample
collection, explicitly agreeing to the use of their blood samples
for the present study. The study was approved by the Ethics
Committee of Fujian Medical University Union Hospital (Fuzhou,
China; approval no.2022KJT046) and was conducted in compliance with
the Declaration of Helsinki. All patients provided written informed
consent to participate.
Fresh blood (1.8 ml) was added to a centrifuge tube
containing 0.2 ml sodium citrate anticoagulant. After
centrifugation at 2,000 × g for 10 min at room temperature, the
upper layer of the supernatant was collected to obtain
platelet-rich plasma. The platelet-rich plasma was centrifuged at
15,00 × g for 15 min at room temperature and the lower pellet was
collected as the platelet precipitate, yielding a platelet
concentration of ~2×106 platelets/µl. The procedure was
completed rapidly to prevent platelet aggregation. To investigate
the effect of Tan IIA on platelet activity, the platelet pellet (10
µl) was resuspended in 1 ml PBS. Platelets were activated with ADP
(20 µM) while being treated with varying concentrations of Tan IIA
(0, 1, 5 and 10 µM). The mixture was incubated at 37°C for 5 min.
Thereafter, 20 µl P-selectin (1:100, PE-labeled; cat. no. 148306;
Biolegend, Inc.) antibody was added to each tube containing 200 µl
of platelet suspension, followed by incubation in the dark at 37°C
for 15 min. The effect of Tan IIA on platelet activity was analyzed
by flow cytometry with a BD FACSAria and the resulting data were
processed using FlowJo software.
Adhesion of cancer to endothelial
cells
Gelatin-coated 24-well plates (gelatin coating was
performed by incubation at 37°C for 1 h) were seeded with HUVECs at
a density of 2×105 cells per well to form confluent
(80–90%) monolayers, which were pretreated with TNF-α (10 ng/ml) at
37°C for 4 h. A group without TNF-α was set as the blank control.
A549 cells suspension at ~6×105 cells/ml treated with
Tan IIA (0, 5, 10, 20 µM) at 37°C and labeled with Rhodamine-123
(10 µl; 15 min) were added and co-cultured with HUVECs at 37°C for
1 h. After washing away non-adherent cells with PBS, fluorescence
microscopy images were captured in 12 randomly selected fields of
view/well. Adhesion was quantified as a percentage relative to
control.
To assess platelet-mediated adhesion, A549 cell
suspension (~6×105 cells/ml) were treated with Tan IIA
(0, 5, 10 and 20 µM) at 37°C and labeled with 10 µl Rhodamine-123
stain for 15 min. After adding 10 µl platelet-rich plasma, the cell
mixture was co-incubated with HUVECs at a density of
2×105 cells per well (pretreated with 10 ng/ml TNF-α for
4 h) at 37°C for 1 h, with the simultaneous addition of ADP (20 µM)
to activate platelets. A group without TNF-α stimulation was used
as the blank control. Adhesion was quantified as a percentage
relative to control.
Cancer cell-platelet aggregation
assay
The platelet cloaking of cancer cells was performed
as previously described (4).
Briefly, A549 cells were harvested and suspended in PBS at a
density of 2×106 cells/ml. Tan IIA (0, 5, 10 and 20 µM)
and A549 cells (2×105 cells) were added to 100 µl PBS
containing suspended platelets (2×108 cells/ml) in
centrifuge tubes, followed by the addition of ADP (20 µM) to
stimulate platelet activation. After 5 min incubation at 25°C, 20
µl each mouse anti-human CD61 (FITC-labeled, cat. no. 104306) and
CD326 (PE-labeled, cat. no. 324205) (both 1:100, Biolegend, Inc.)
were added to the tubes. The mixture was incubated for 20 min at
4°C in the dark, followed by the addition of 1 ml ice-cold 1%
paraformaldehyde for fixation at 4°C for 30 min. A549 cells cloaked
by platelets were identified by flow cytometry with a BD FACSAria;
CD61+CD326+ cancer cells were considered
platelet-cloaked cancer cells. The data were analyzed using FlowJo
software.
Fluorescence microscopy of
platelet-cancer cell interaction
The adhesion of platelets to tumor cells following
Tan IIA (0, 5, 10, and 20 µM) treatment was visualized using a
confocal fluorescence microscope (Leica GmbH; STELLARIS 5).
Suspended platelets (10 µl at a density of ~2×106
platelets/µl) were collected and stained with 5 µM
5-(and-6)-carboxyfluorescein diacetate N-succinimidyl ester (cat.
no. 21888; Sigma-Aldrich; Merck KGaA) in PBS for 10 min at 37°C.
The nuclei of A549 cells were stained with DAPI (1 µg/ml in PBS)
for 15 min at 25°C in the dark and the cell membranes were stained
with 2 µM PKH26 Red Fluorescent Cell Linker (cat. no. MINI26-1KT;
Sigma-Aldrich; Merck KGaA) for 15 min at 25°C in the dark.
Subsequently, the stained platelets and tumor cells were
co-cultured for 10 min at 25°C in the dark. After co-culture,
unattached platelets were removed by washing with PBS and the
platelet-tumor aggregates were transferred to confocal dishes for
incubation at 37°C for 4 h. Finally, 12 randomly selected fields of
view/dish were observed using a confocal microscope. The number of
platelets adhering to tumor cells was quantified by ImageJ
software.
Western blotting
Western blotting was performed to detect the effect
of Tan IIA at safe concentrations (0, 5 and 10 µM) at 37°C for 24 h
on the NF-κB signaling pathway in HUVECs. Whole-cell lysates were
from HUVECs prepared using RIPA Buffer (cat. no. WB3100; Suzhou
Xinsaimei Biotechnology Co., Ltd.) with 1% protease/phosphatase
inhibitors on ice for 30 min, followed by centrifugation at 12,000
× g for 20 min at 4°C. Protein concentration was determined using
the BCA method. Proteins (10 µg/lane) were separated using 4–12%
gradient precast gels according to their molecular sizes and
transferred to PVDF membranes. Membranes were blocked with 5%
skimmed milk and 5% BSA (cat. no. T50811010260; Beijing Dingguo
Changsheng Biotechnology Co., Ltd.) at room temperature for 90 min
and incubated with primary antibodies at 4°C overnight. After
washing with TBST (0.1% Tween) four times, 5 min each, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibodies (1:5,000; cat. no. 511203; Zenbio; Chengdu
Zhengneng Biotechnology Co., Ltd.) for 1 h at room temperature.
Signals were detected using an ECL substrate (NcmECL Ultra; cat.
no. P10300; Suzhou Xinsaimei Biotechnology Co., Ltd.) and a
chemiluminescence imaging system (ChemiDoc™ MP; Bio-Rad
Laboratories, Inc.). The following primary antibodies were used:
GAPDH (1:10,000; cat. no. A19056; ABclonal Biotech Co., Ltd.),
NF-κB p65 (cat. no. R25149), phosphorylated (p-)NF-κB p65 (cat. no.
310013), IκB-α (cat. no. R23322), p-IκB-α (cat. no. 340776),
IKK-α/β (cat. no. R24676) and p-IKK-α/β (all 1:1,000; cat. no.
530546; all Zenbio). Band intensities from three independent
experiments were semi-quantified using ImageJ software.
Molecular docking
The 3D structure of Tan IIA was downloaded from the
PubChem database (pubchem.ncbi.nlm.nih.gov/) in SDF format. The
compound was converted to MOL2 format using PyMOL (version number
3.1.6; Delano Scientific LLC), and its geometric conformation was
automatically optimized. The 3D structure of the target protein was
retrieved from the Research Collaboratory for Structural
Bioinformatics Protein Data Bank (rcsb.org/). The protein structure
was processed by removing water molecules, adding hydrogen atoms
and checking charge states. Both the ligand (Tan IIA) and the
receptor (CD29 and P-selectin) were converted to the PDBQT format
using the MGLTools software package (version number 1.5.7; The
Scripps Research Institute). Molecular docking was then performed
using AutoDock Vina (version number 1.1.2; The Scripps Research
Institute) to calculate the binding energies. The best docking pose
was selected and visualized using PyMOL.
In vivo assays
All animal procedures were conducted in accordance
with the Guide for the Care and Use of Laboratory Animals (National
Research Council, 2011) (40). The
Laboratory Animal Ethics Committee of Minjiang University (Fuzhou,
China) reviewed and approved all animal procedures (approval no.
IACUC-MJLAC-2023-020).
Female BALB/c and nude mice (n=110; weight, 20±2 g;
age, 6–8 weeks) were obtained from Shanghai SLAC Laboratory Animal
Co., Ltd. The animals were acclimatized to the laboratory
environment for ≥1 week prior to the start of the experiment. All
mice were housed in a specific-pathogen-free facility at the Animal
Experiment Center of Minjiang University, with 5 mice/cage under a
12/12-h light/dark cycle at a constant temperature of 23–25°C and
50–60% relative humidity. They were fed a standard rodent diet and
had free access to sterile water, which was replaced daily. The
experiment lasted 4–6 weeks.
Animal health and behavior were monitored daily for
clinical signs including activity levels, appetite and grooming.
Body weights were measured and recorded every 3 days throughout the
study. Humane endpoints were as follows: i) Maximum tumor volume
>750 mm3; ii) severe clinical manifestations such as
dyspnea or listlessness; and iii) body weight loss >20% of
initial body weight. In total, four mice were prematurely
euthanized based on tumor volume and clinical manifestations.
To prepare the vehicle, 0.1 g sodium carboxymethyl
cellulose (cat. no. C104984; Shanghai Aladdin Biochemical
Technology Co., Ltd.) was dissolved in 100 ml double-distilled
water to obtain a 0.1% solution. Tan IIA was dissolved in this 0.1%
sodium carboxymethyl cellulose solution to prepare Tan IIA
solutions at concentrations of 0.1, 1 and 2 mg/100 µl for oral
gavage. The anti-metastatic efficacy of Tan IIA was evaluated using
three independent mouse cancer models (n=10). For the A549 lung
metastasis model, A549 cells in logarithmic growth phase
(1×105 cells in 100 µl per mouse) were injected into the
tail vein of nude mice. For the hematogenous metastasis model, 4T1
cells in logarithmic growth phase (1×105 cells in 100 µl
per mouse) were injected via the tail vein of Balb/c mice. For the
orthotopic metastasis model, 4T1 cells (5×105 cells in
100 µl per mouse) were injected into the fourth mammary fat pad of
Balb/c mice to establish an orthotopic tumor model that simulates
the natural metastatic progression of breast cancer.
In the A549 lung metastasis model, a high-(1 mg/100
µl/day) and low-dose (0.1 mg/100 µl/day) and control group (100 µl
0.1% sodium carboxymethyl cellulose solution) were set up. In the
4T1 breast cancer metastasis model, a high-(2 mg/100 µl/day),
medium-(1 mg/100 µl/day) and low-dose (0.1 mg/100 µl/day) and
control group (100 µl 0.1% sodium carboxymethyl cellulose solution;
vehicle) were established. Mice continued to receive 100 µl Tan IIA
solution or vehicle by oral gavage daily for 4–6 weeks. During this
period, the physiological status of the mice was observed and
recorded. Body weight was measured and recorded for all surviving
mice at every 3 days. At the end of the experiment, mice were
anesthetized by inhalation of 3–5% isoflurane in oxygen for
induction, followed by maintenance anesthesia with 1–2% isoflurane.
Mice were euthanized with a CO2 displacement rate of 30%
of the container volume/min, followed by cervical dislocation.
Death was confirmed by cessation of heartbeat and respiration. The
lungs were removed fixed in 4% paraformaldehyde at 4°C overnight,
stained with picric acid at room temperature for 72 h and the
number of metastatic nodules was assessed under a dissecting
microscope. GraphPad Prism software (version number 8.0.2;
Dotmatics) was used for analysis.
Statistical analysis
Data are presented as the mean ± standard deviation
of ≥3 independent experimental repeats. Statistical analyses were
performed using GraphPad Prism. Differences were evaluated by
one-way ANOVA followed by Dunnett's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
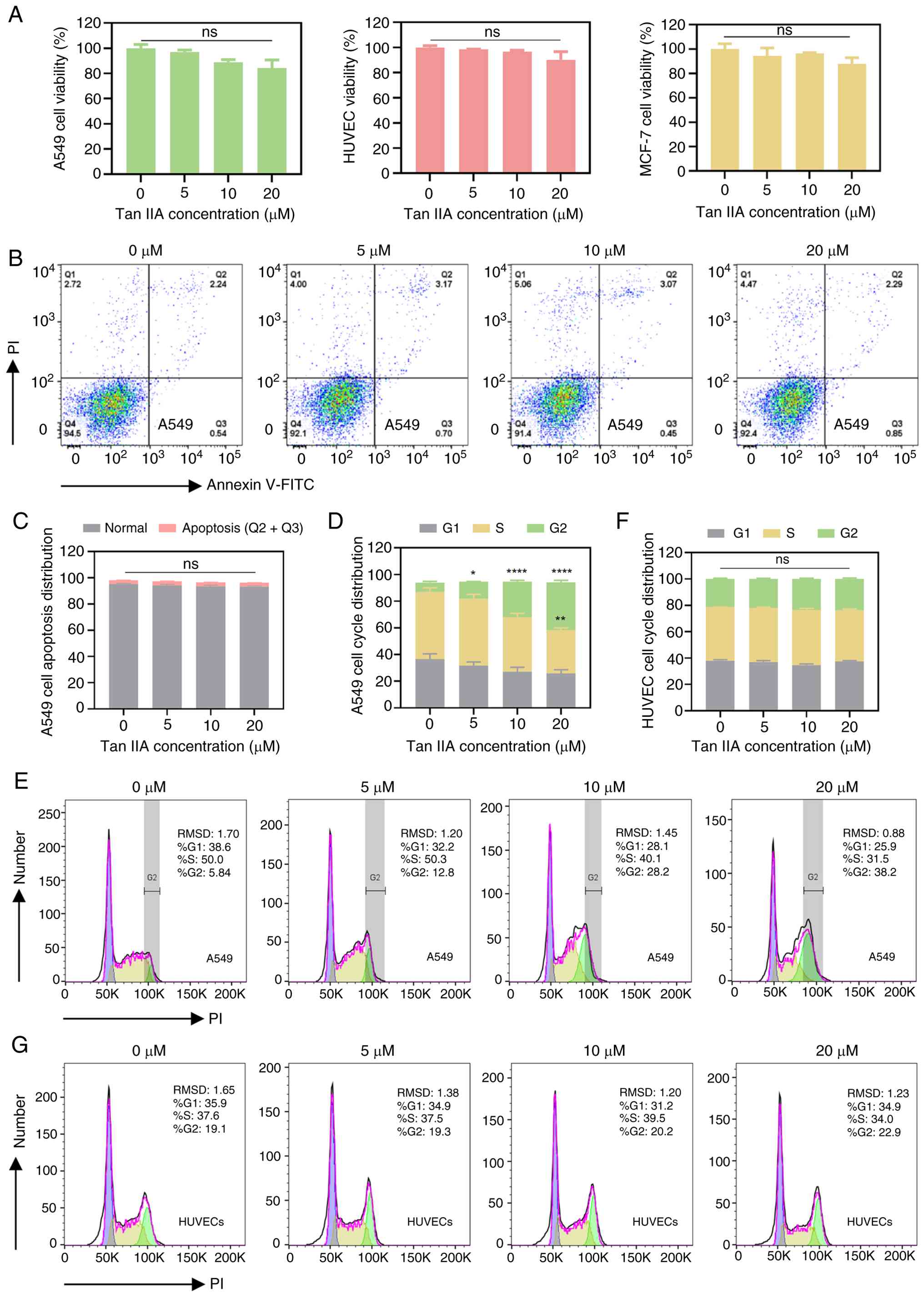
Tan IIA shows weak cytotoxicity toward
A549 cells and HUVECs
The present study aimed to identify Tan IIA
concentrations that exert minimal direct cytotoxic effects on tumor
and endothelial cells, thereby allowing investigation of its
effects on metastasis-associated interactions. Viability was
assessed in A549, HUVECs and MCF-7 cells following Tan IIA
treatment. Cell viability slightly decreased with increasing Tan
IIA concentration, however there was no significant difference in
any cell line (Fig. 1A). Consistent
with these findings, apoptosis analysis showed no significant
increase in apoptotic A549 cells following treatment with Tan IIA
(Fig. 1B and C). Cell cycle
analysis revealed A549 cell cycle arrest at the G2/M phase
following treatment with Tan IIA, while the cell cycle distribution
of HUVECs did not change significantly (Fig. 1D-G). This supports the selective
action of Tan IIA on carcinoma over normal endothelial cells, a key
point for its potential as an anti-metastatic agent with a
favorable safety profile regarding vascular toxicity.
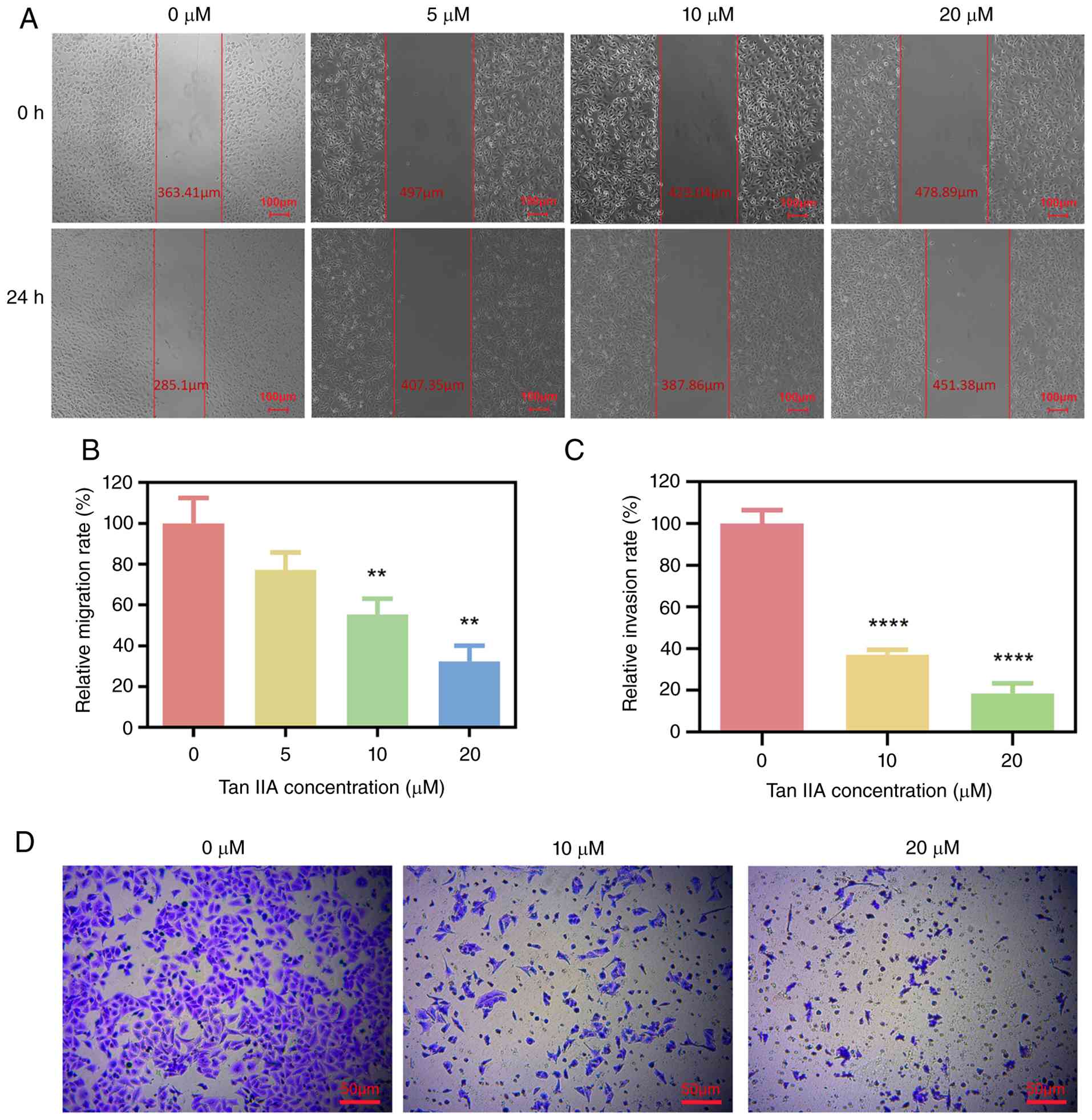
Tan IIA suppresses migration and
invasion of A549 cells
Tumor cell migration and invasion are key steps in
metastatic progression. To examine whether Tan IIA affects these
processes, wound healing and Transwell invasion assays were
performed. In the wound healing assay, A549 cells treated with
non-cytotoxic concentrations of Tan IIA (10 and 20 µM) showed
significantly decreased migration compared with untreated controls
after 24 h, however lower doses of Tan IIA had no significant
effect (Fig. 2A and B). Similarly,
Tan IIA inhibited A549 cell invasion in a concentration-dependent
(Fig. 2C and D). These findings
indicated that Tan IIA effectively suppressed both migration and
invasion of lung cancer cells independent of overt
cytotoxicity.
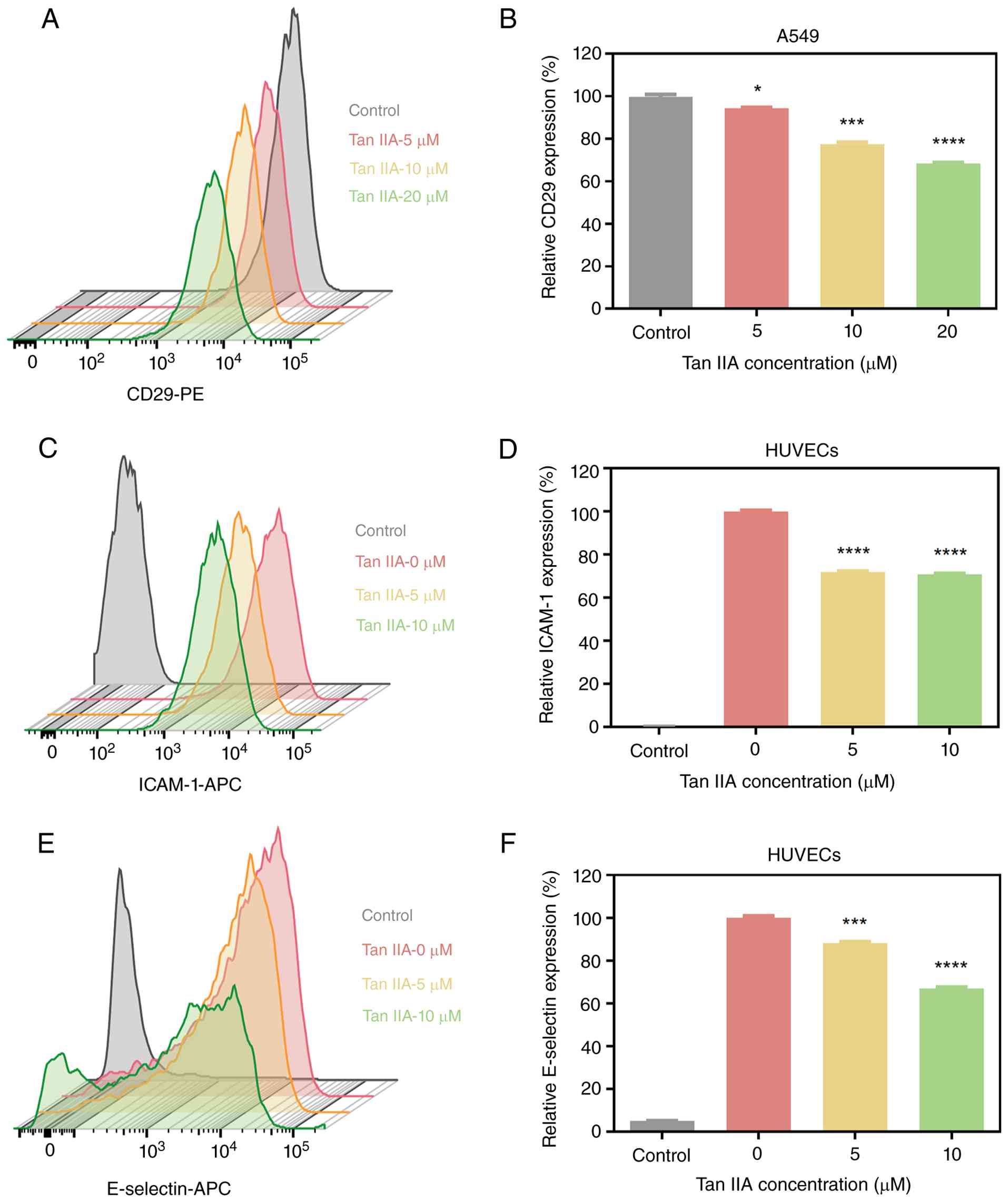
Tan IIA reduces adhesion molecules
expression on tumor cells and endothelial cells
Adhesion between tumor cells, platelets and
endothelial cells plays a central role in metastatic dissemination
(41–43). The present study examined whether
Tan IIA modulates adhesion molecule expression on tumor cells.
CD29, a key integrin involved in platelet binding and cell
adhesion, was assessed on A549 cells by flow cytometry (44,45).
Tan IIA resulted in a concentration-dependent reduction in CD29
expression on the A549 cell surface (Fig. 3A and B). The present study evaluated
the effects of Tan IIA on endothelial adhesion molecules.
E-selectin and ICAM are inducible adhesion proteins that facilitate
tumor cell attachment to activated endothelium (46). Flow cytometric analysis revealed
that Tan IIA significantly decreased the surface expression of
E-selectin on HUVECs in a concentration-dependent manner; the
expression of ICAM-1 on HUVECs was significantly reduced, while no
significant difference in ICAM-1 expression was observed between
higher Tan IIA doses (Fig. 3C-F).
These results suggested that Tan IIA disrupts a
metastasis-promoting microenvironment by decreasing adhesion
molecule expression on both tumor cells and endothelial cells.
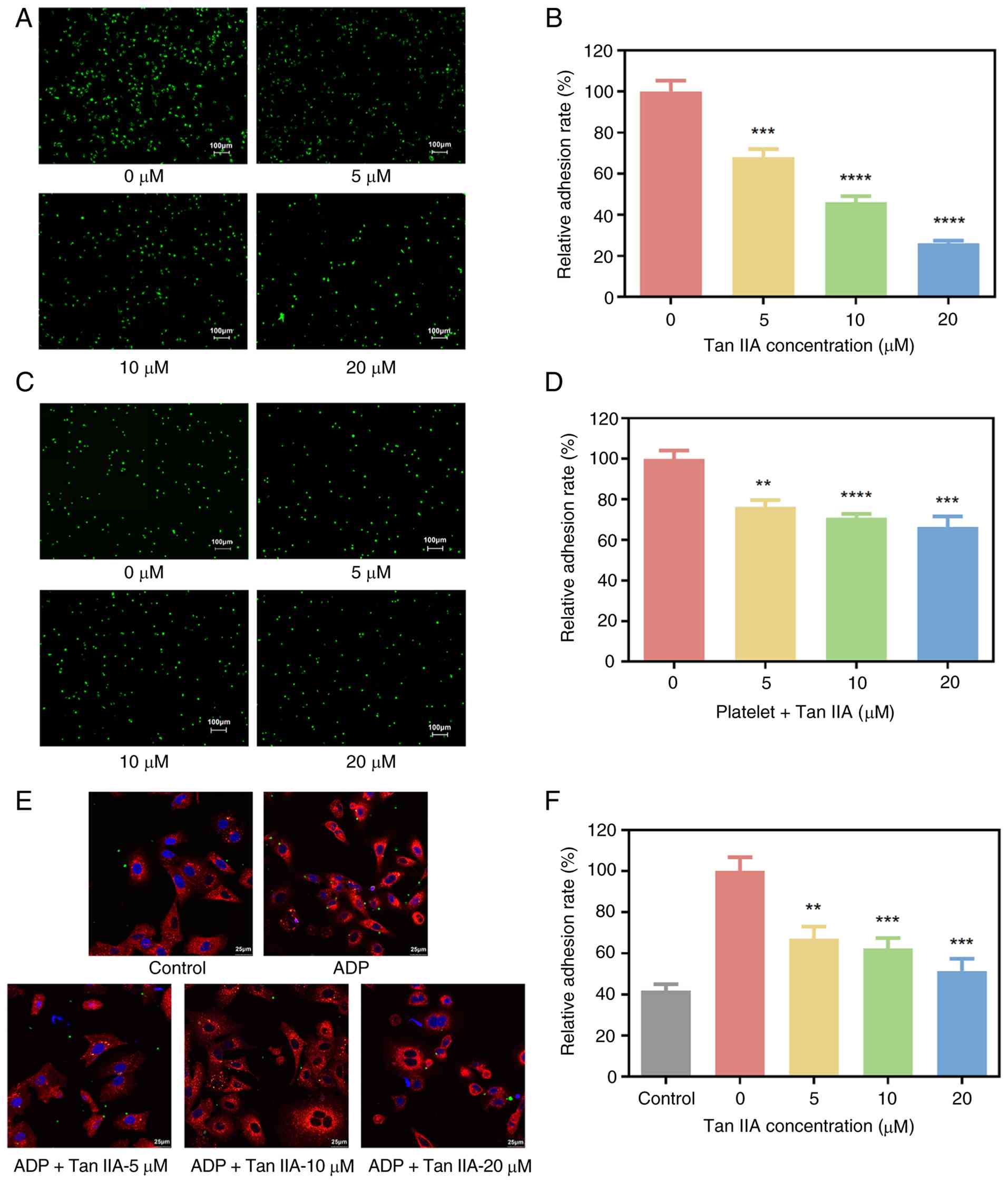
Tan IIA inhibits platelet activation
and tumor cell-platelet aggregate formation
Platelet activation and aggregation around CTCs
enhance tumor survival and metastatic potential (47). P-selectin is a member of the
selectin family of cell adhesion molecules. It is expressed on
stimulated endothelial cells and activated platelets, mediating
both heterotypic aggregation between activated platelets and cancer
cells, as well as the adhesion of cancer cells to stimulated
endothelial surfaces (48). To
evaluate the effect of Tan IIA on platelet activity, P-selectin
expression was measured by flow cytometry. ADP markedly increased
P-selectin expression; addition of Tan IIA significantly decreased
P-selectin levels in compared with the control (Fig. 4A and B), indicating inhibition of
platelet activation.
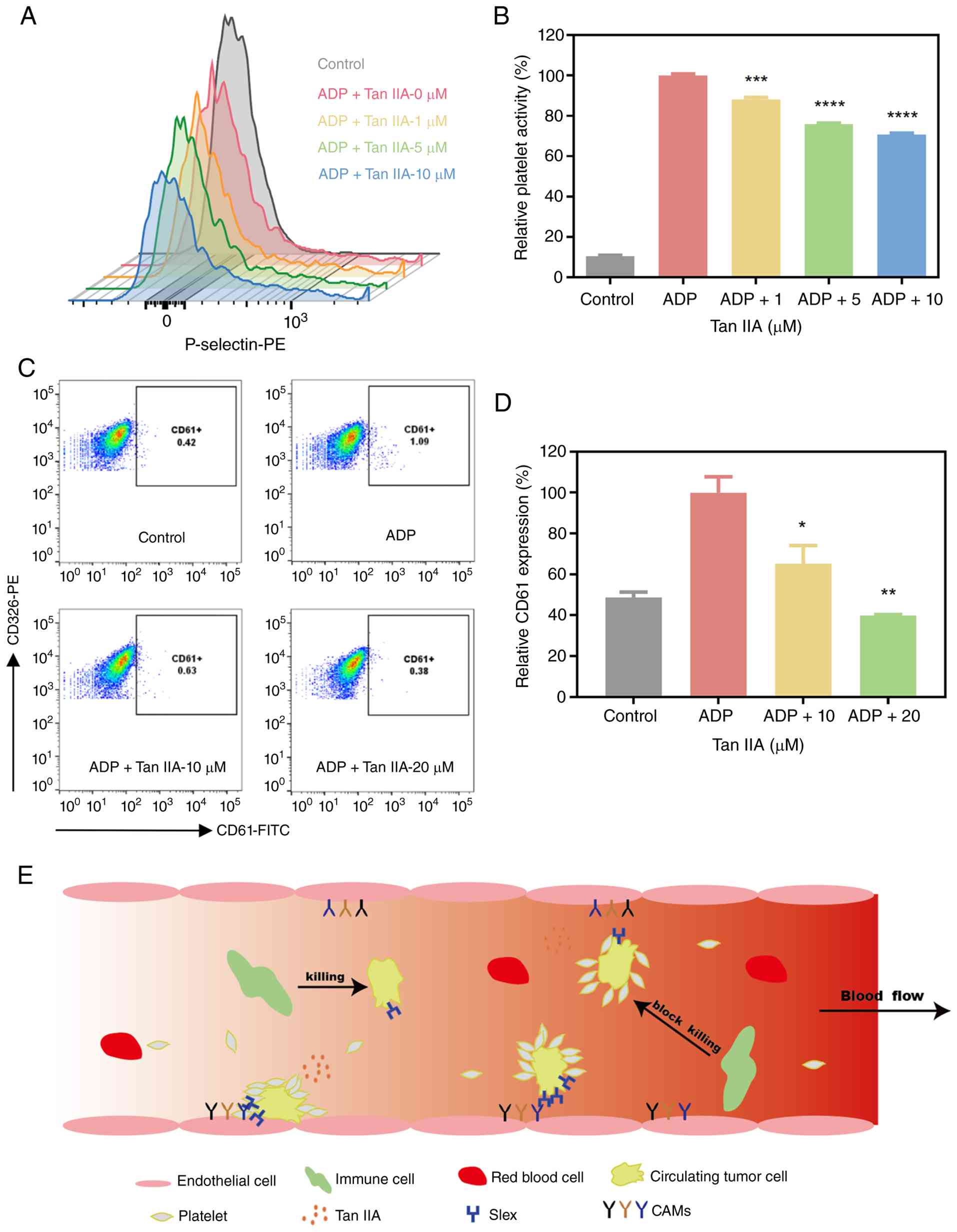 | Figure 4.Effects of Tan IIA on platelet
activation and tumor cell-platelet aggregation. (A) Tan IIA
significantly inhibited platelet activation, as indicated by
reduced expression of the platelet activation marker P-selectin.
(B) Quantitative results of platelet activity. (C) Flow cytometric
detection of tumor cell-platelet aggregation. (D) Quantitative
results showed that Tan IIA markedly decreased the formation of
platelet-tumor cell aggregates. *P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001 vs. ADP. (E) Schematic of the
inhibitory effects of Tan IIA on platelet-mediated interactions
within the circulating tumor cells microenvironment. Tumor cell
surface antigen sLex is recognized by P-selectin receptors on
platelets, forming platelet-tumor cell aggregates, which protect
tumor cells from immune recognition and attack. Following Tan IIA
treatment, the expression of P-selectin on platelets is decreased,
leading to disruption of platelet-tumor cell aggregates and partial
or complete exposure of tumor cells to the immune system. Binding
of tumor cell surface antigen sLex to selectins on endothelial
cells increases adhesion, and Tan IIA can also inhibit this process
by suppressing E-selectin expression on endothelial cells. Tan IIA
can inhibit the adhesion of tumor cells to endothelial cells by
decreasing expression of ICAM-1 on the endothelial cell surface and
CD29 on the tumor cell surface, a process that depends on the
binding of ICAM-1/VCAM between tumor and endothelial cells. Tan
IIA, tanshinone IIA; sLex, sialyl Lewis X; ICAM, intercellular
adhesion molecule; VCAM, vascular cell adhesion molecule. |
To assess tumor cell-platelet interactions, platelet
cloaking of A549 cells was examined by flow cytometry. Under
resting conditions, few A549 cells exhibited platelet binding.
Following ADP-induced platelet activation, the proportion of
platelet-coated tumor cells increased markedly. Tan IIA
significantly decreased platelet adhesion to A549 cells (Fig. 4C and D). These findings indicate
that Tan IIA interfered with platelet-tumor cell aggregate
formation by suppressing platelet activation. Collectively, these
results demonstrated that Tan IIA may limit key steps of
CTC-mediated metastasis by inhibiting platelet activation,
decreasing tumor cell-platelet aggregation and decreasing adhesion
molecule expression (Fig. 4E).
Tan IIA inhibits tumor cell adhesion
to endothelial cells
Adhesion of CTCs to the vascular endothelium is a
prerequisite for extravasation and metastatic colonization
(49). To evaluate whether Tan IIA
affected this process, A549 cells treated with Tan IIA were
co-cultured with TNF-α-stimulated HUVEC monolayers. Tan IIA
significantly decreased the number of tumor cells adhering to
endothelial cells in a concentration-dependent manner (Fig. 5A and B). To investigate the role of
platelets in this process, a co-culture system incorporating A549
cells, platelets and activated HUVECs was established. In the
presence of platelets, Tan IIA inhibited tumor cell adhesion to
endothelial cells (Fig. 5C and D).
Confocal microscopy confirmed that Tan IIA decreased platelet
coverage of tumor cells, resulting in fewer platelet-tumor cell
aggregates (Fig. 5E and F). These
findings indicate that Tan IIA inhibits platelet-mediated tumor
cell adhesion to the endothelium.
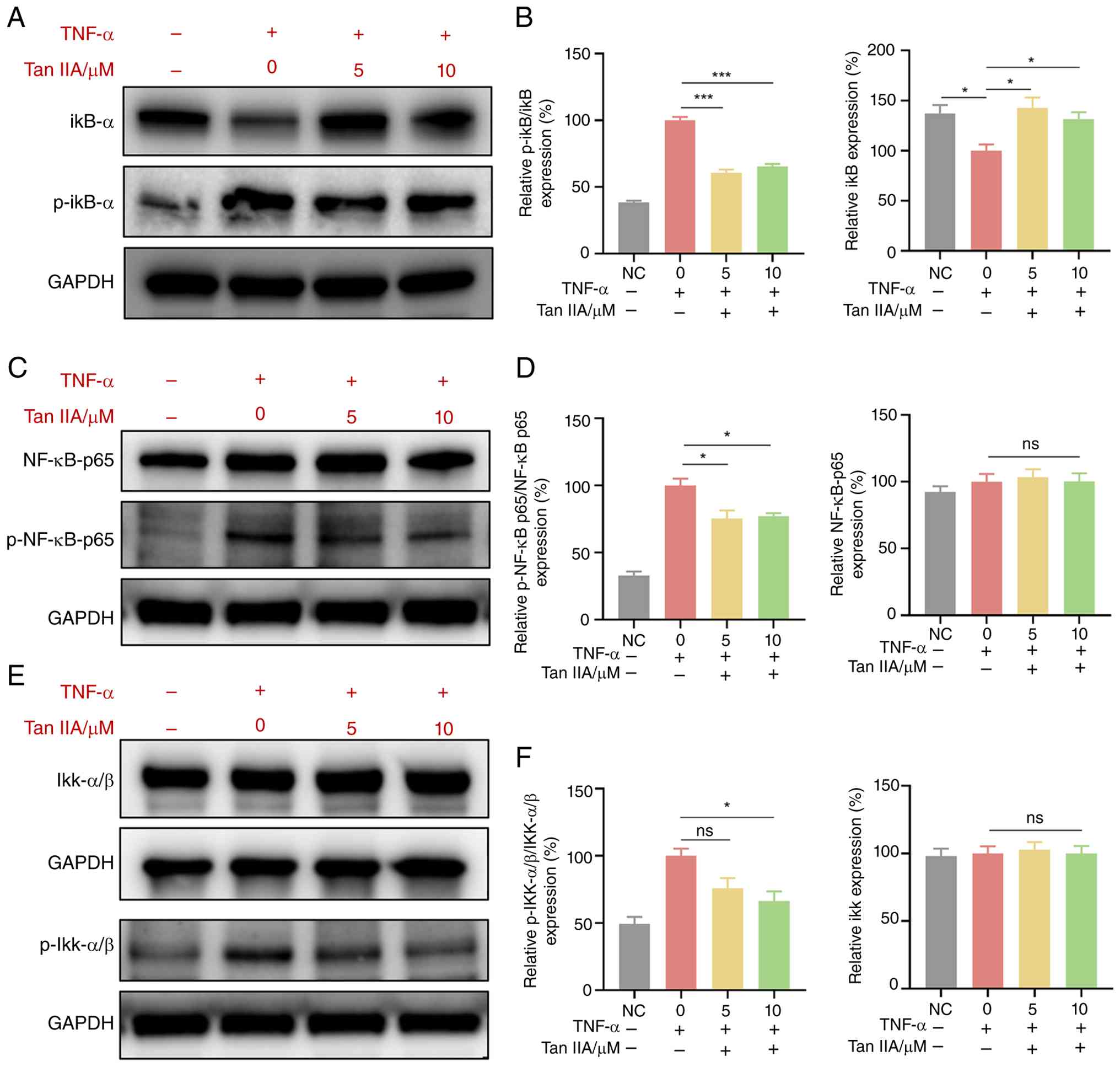
Tan IIA inhibits E-selectin and ICAM
expression on HUVECs by regulating the NF-κB signaling pathway
To investigate how Tan IIA affects the expression of
the endothelial surface proteins E-selectin and ICAM, the present
study examined whether Tan IIA influences the NF-κB signaling
pathway in endothelial cells. Endothelial cells were cultured in
the presence or absence of TNF-α stimulation. Western blotting was
performed to evaluate the effect of Tan IIA on the NF-κB signaling
pathway. TNF-α increased the normalized levels of NF-κB
pathway-related proteins, including p-IKKα/β, p-IκB-α and p-NF-κB
p65 (each normalized to its respective total protein), indicating
activation of this signaling pathway (Fig. 6). Compared with the 0 µM group, Tan
IIA at 10 µM significantly reduced the normalized levels of these
proteins (p-IKKα/β, p-IκB-α and p-NF-κB p65). These results
indicated that Tan IIA-mediated inhibition of expression of
E-selectin and ICAM on HUVECs is associated with NF-κB signaling
pathway.
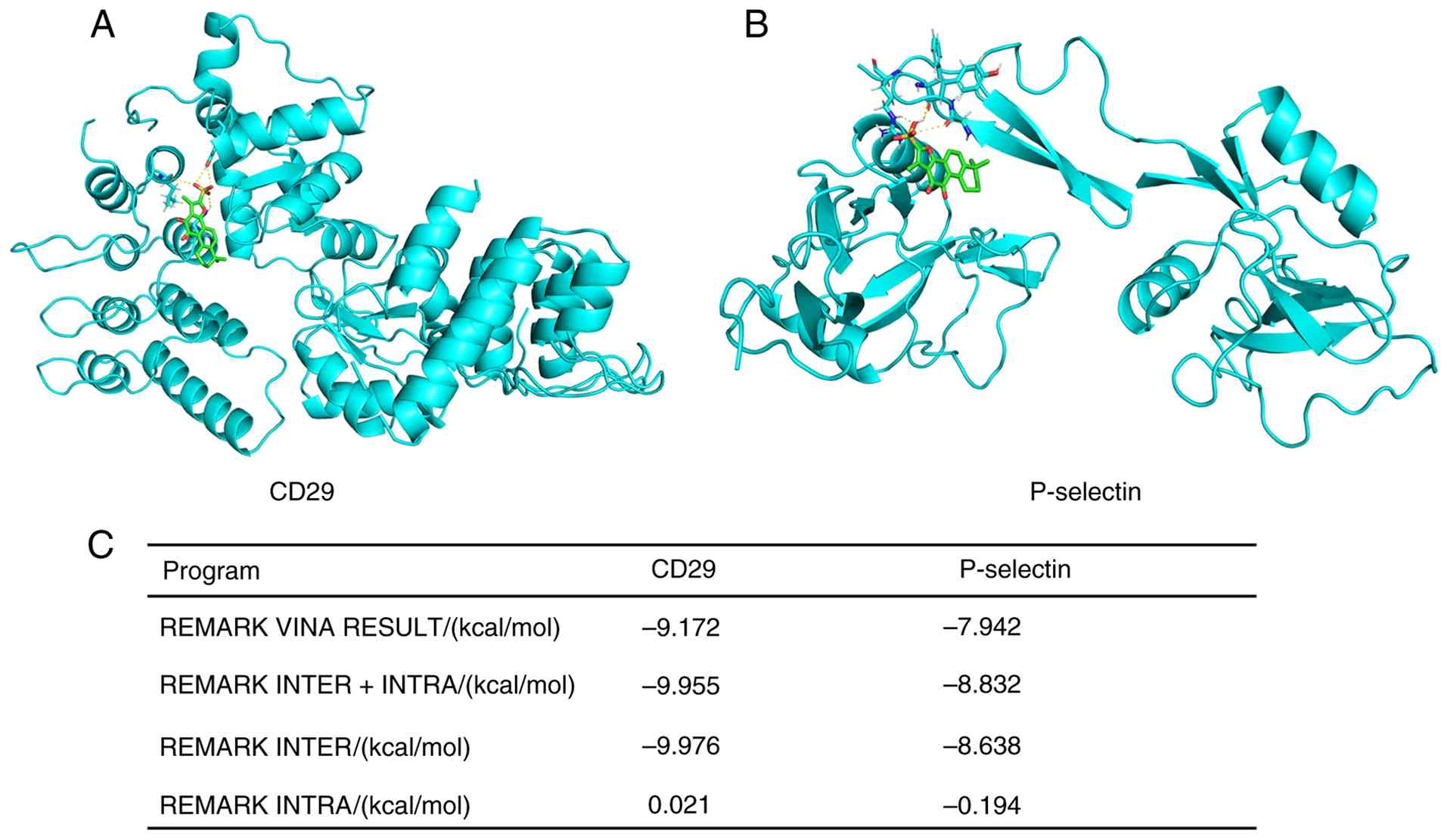
Molecular docking predicted that Tan IIA can
favorably bind to the extracellular domains of CD29(−9.976
kcal/mol) and P-selectin (−8.638 kcal/mol) with strong binding
affinity (Fig. 7). The docking
models suggested that Tan IIA potentially binds key functional
regions or pockets on these proteins, which may interfere with
their inter-molecular interactions.
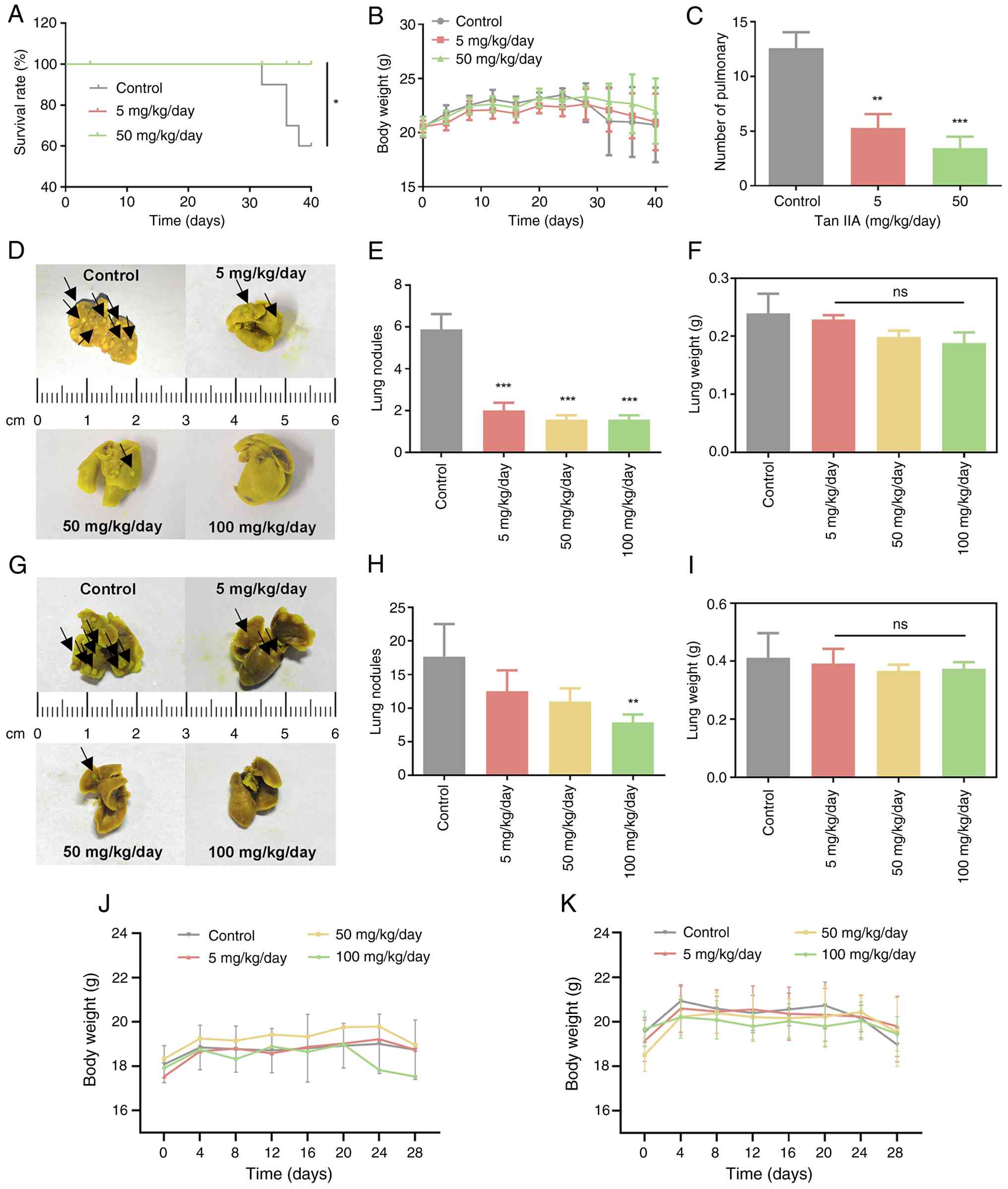
Tan IIA inhibits metastatic
progression and improves outcomes in vivo
To evaluate the anti-metastatic effects of Tan IIA
in vivo, mouse models were established. In the A549 tail
vein metastasis model, Tan IIA significantly prolonged survival
compared with controls (P=0.0169; Fig.
8A), but no significant body weight loss was observed during
treatment (Fig. 8B). The number of
lung metastatic nodules was significantly decreased in Tan
IIA-treated mice, particularly in the 50 mg/kg/day group (Fig. 8C).
Consistent results were observed in both 4T1 models.
In the hematogenous model, Tan IIA significantly decreased the
number of lung metastases (Fig. 8D and
E). Similarly, in the orthotopic model, Tan IIA led to fewer
lung metastatic nodules compared with controls (Fig. 8G and H). Lung weight was largely
unchanged across groups (Fig. 8F and
I) and no significant body weight loss was observed during
treatment (Fig. 8J and K),
indicating good tolerability. Taken together, these in vivo
findings demonstrated that Tan IIA effectively suppressed
metastatic tumor burden while improving overall physiological
condition and survival.
Discussion
Among 1,030,937 US (1992–2019) metastatic cancer
survivors, 82.6% died from their metastatic cancer (highest in
lung, pancreas, esophagus and stomach tumors) and 17.4% from
competing causes (50). Metastasis
poses clinical challenges such as therapeutic resistance, lesion
heterogeneity and plasticity (51).
The presence of CTCs in the bloodstream reflects the metastatic
potential of malignant tumors and is associated with disease
progression (52). In clinical
practice, the number and biological characteristics of CTCs are
increasingly (53,54) recognized as valuable indicators for
monitoring metastatic risk and therapeutic response (55). Because adhesion of CTCs to vascular
endothelial cells is a key early event in hematogenous metastasis,
strategies that eliminate CTCs or disrupt their adhesion and
extravasation represent effective approaches to suppress metastatic
spread (56). Studies have shown
that the hematogenous microenvironment of CTCs, particularly
platelet activation, platelet-tumor cell interactions and
endothelial adhesion molecule expression, is a potential target for
anti-metastatic intervention (57,58).
Traditional Chinese Medicine is characterized by
multi-target actions and low toxicity and is widely applied as an
adjunctive approach in cancer treatment, including metastatic
disease (59,60). Tan IIA, a bioactive compound derived
from S. miltiorrhiza, has demonstrated anti-tumor activity
in multiple cancer types (61).
Rather than exerting strong direct cytotoxic effects, Tan IIA
exerts regulatory effects on vascular function, platelet activity
and inflammatory responses (20).
The present study therefore focused on the effects of Tan IIA at
low, non-cytotoxic concentrations on metastasis-associated cellular
interactions, specifically those among tumor cells, platelets and
endothelial cells.
Once CTCs enter the circulation, they interact with
platelets, resulting in platelet activation and the formation of
tumor cell-platelet aggregates (57). Activated platelets express high
levels of P-selectin, which mediates platelet adhesion to CTCs and
contributes to platelet cloaking of CTCs. This cloaking protects
tumor cells from hemodynamic shear stress and immune-mediated
clearance and facilitates their arrest within the vasculature
(62). Platelet activation and
inflammatory stimulation promote endothelial activation, leading to
increased expression of adhesion molecules such as E-selectin and
ICAM-1 on endothelial cells (63).
These molecules support tumor cell rolling, firm adhesion and
stable attachment to the endothelial surface, which are
prerequisites for transendothelial migration and metastatic
colonization (64). Consistent with
this metastatic cascade, the present results demonstrated that Tan
IIA interferes with multiple steps of tumor dissemination. Tan IIA
reduced the expression of CD29 on A549 cells, thereby weakening
tumor cell adhesive capacity. Tan IIA inhibited platelet
activation, as indicated by decreased P-selectin expression, and
significantly decreased platelet adhesion to CTCs, limiting
platelet cloaking. Moreover, Tan IIA attenuated TNF-α-induced
upregulation of endothelial adhesion molecules ICAM-1 and
E-selectin on HUVECs. These effects resulted in reduced tumor cell
adhesion to endothelial cells and impaired platelet-mediated
enhancement of tumor cell-endothelial interaction.
Endothelial cell activation and aberrantly high
expression of adhesion molecules are key steps in the initiation of
inflammatory responses, as well as in tumor cell adhesion and
metastasis (65,66). The present study found that Tan IIA
inhibited the upregulation of ICAM-1 and E-selectin in
TNF-α-stimulated HUVEC cells. Tan IIA significantly decreased
TNF-α-induced phosphorylation of IκBα, IKK-α/β and p65 (normalized
to its respective total protein), indicating that Tan IIA inhibited
adhesion molecule expression by blocking the canonical NF-κB
pathway. This is consistent with anti-inflammatory effects of Tan
IIA in other cell types (67,68).
Furthermore, computational molecular docking predicted that Tan IIA
can favorably bind to the extracellular domains of CD29 and
P-selectin with strong binding affinity (binding energies <-6.8
kcal/mol). These results demonstrated that Tan IIA may exert its
inhibitory effects, at least in part, through direct binding to
these key adhesion molecules, potentially blocking their reciprocal
interactions.
The present in vitro findings were supported
by in vivo evidence from metastatic mouse models. Tan IIA
decreased lung metastatic burden and prolonged survival in the A549
tail vein model, while also suppressing pulmonary metastasis in
both tail vein and orthotopic 4T1 breast cancer models. Notably,
these anti-metastatic effects occurred without significant changes
in body or lung weight, suggesting that Tan IIA exerted its effects
primarily through modulation of metastatic processes rather than
overt toxicity. The concentrations of Tan IIA (5–20 µM) used to
inhibit platelet activation, endothelial adhesion and tumor cell
adhesion were selected based on prior mechanistic studies (33,69,70).
While direct measurement of plasma concentrations in the present
mouse model was not performed, pharmacokinetics data provide a
relevant context. For example, oral administration of 4.1 mg/kg Tan
IIA in rats results in a maximum concentration of 2.78±0.96 ng/ml
(~8.7 nM) and an area under the curve of 4.53 ng·h/ml (71). The effective concentrations in
vitro are therefore higher than reported systemic plasma
levels. This is a common scenario for natural compounds where in
vitro mechanistic studies employ concentrations that may not
reflect systemic bioavailability but are justified to elucidate
potential targets and pathways (72). Effective anti-metastatic
concentrations may result in significantly higher local
concentrations in key tissue such as the lung or liver due to
tissue distribution and accumulation, as suggested for other
lipophilic compounds (73,74). Furthermore, the primary metabolite
of Tan IIA, tanshinol, may also contribute to the overall
biological activity (75,76). Future studies measuring
tissue-specific pharmacokinetics and the activity of notable
metabolites are warranted to bridge this gap and optimize dosing
strategies.
The present study had limitations. First, while the
present study demonstrates a clear association between Tan IIA
treatment and decreased tumor cell-platelet-endothelial
interactions, the precise upstream molecular targets of Tan IIA
were not fully delineated and did not formally establish that this
axis is the primary mediator of the observed reduction in pulmonary
metastasis. Future studies employing conditional platelet
depletion, tumor cells with genetic ablation of specific adhesion
receptors (CD29) or neutralizing antibodies against key interaction
molecules (P-selectin) in metastatic models are required to
validate the causal role of the disrupted adhesion cascade. Second,
while HUVECs are a well-established and accessible model for
studies of endothelial behavior, organ-specific microvascular
endothelial cells (such as from the lung) may differ in their
response to stimuli (77–79). Future studies employing lung
microvascular endothelial cells are required to validate the
translational relevance of the present findings to pulmonary
metastasis. Finally, although the in vivo models provided
support for an anti-metastatic role of Tan IIA, additional
pharmacokinetic and dose-response studies are necessary to
facilitate clinical translation.
Taken together, the present findings supported the
hypothesis that Tan IIA suppresses hematogenous metastasis
primarily by modulating interactions between CTCs, platelets and
endothelial cells. Tan IIA may do this by attenuating platelet
activation, decreasing tumor cell-platelet aggregation and limiting
tumor cell adhesion to the vascular endothelium, and inhibiting the
NF-κB signaling pathway that leads to adhesion protein expression
in vascular endothelial cells. Tan IIA disrupts key early steps of
metastatic dissemination. The present results highlight Tan IIA as
a potential pharmacological agent targeting the metastatic
microenvironment rather than tumor cell viability.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant nos. 81703555 and 81961138017), the
Department of Science and Technology of Fujian Province (grant nos.
2025I0051, 2023Y9124 and 2021D043) and the Research Project of
Fashu Foundation (grant no. MFK24021).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
JX analyzed data, designed and performed
experiments, constructed figures and wrote the manuscript. LL
analyzed data, designed and performed experiments and wrote the
manuscript. FL analyzed data, designed and performed experiments
and edited the manuscript. SW analyzed data, designed and performed
experiments and supervised the study. CF designed the study. GW
designed and performed experiments and constructed figures. YW and
YL conceived the study and edited the manuscript. YL analyzed data,
constructed figures, supervised the study and edited the
manuscript. JX and FL confirm the authenticity of all the raw data.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The animal study was reviewed and approved by the
institutional animal care and use committee of Minjiang University,
Fuzhou, China (approval no. IACUC-MJLAC-2023-020). The collection
and use of human blood samples in this study was reviewed and
approved by the Ethics Committee of Fujian Medical University Union
Hospital, Fuzhou, China (approval no. 2022KJT046). All patients
provided written informed consent to participate.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Garemilla SSS, Kadambala MC, Gampa SC,
Chinthala S and Garimella SV: Cancer metastasis: Therapeutic
challenges and opportunities. Med Oncol. 42:5182025. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Steeg PS: Tumor metastasis: Mechanistic
insights and clinical challenges. Nat Med. 12:895–904. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shen MM: Cancer: The complex seeds of
metastasis. Nature. 520:298–299. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu Y, Lian S, Ye Y, Yu T, Liang H, Cheng
Y, Xie J, Zhu Y, Xie X, Yu S, et al: S-Nitrosocaptopril prevents
cancer metastasis in vivo by creating the hostile bloodstream
microenvironment against circulating tumor cells. Pharmacol Res.
139:535–549. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu Y, Liang H, Yu T, Xie J, Chen S, Dong
H, Sinko PJ, Lian S, Xu J, Wang J, et al: Isolation and
characterization of living circulating tumor cells in patients by
immunomagnetic negative enrichment coupled with flow cytometry.
Cancer. 121:3036–3045. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou L, Zhang Z, Tian Y, Li Z, Liu Z and
Zhu S: The critical role of platelet in cancer progression and
metastasis. Eur J Med Res. 28:3852023. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Morris K, Schnoor B and Papa AL: Platelet
cancer cell interplay as a new therapeutic target. Biochim Biophys
Acta Rev Cancer. 1877:1887702022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rejniak KA: Circulating tumor cells: When
a solid tumor meets a fluid microenvironment. Adv Exp Med Biol.
936:93–106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kong DH, Kim YK, Kim MR, Jang JH and Lee
S: Emerging roles of vascular cell adhesion Molecule-1 (VCAM-1) in
immunological disorders and cancer. Int J Mol Sci. 19:10572018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lu Y, Lian S, Cheng Y, Ye Y, Xie X, Fu C,
Zhang C, Zhu Y, Iqbal Parker M and Jia L: Circulation patterns and
seed-soil compatibility factors cooperate to cause cancer
organ-specific metastasis. Exp Cell Res. 375:62–72. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wei B, Sun C, Wan H, Shou Q, Han B, Sheng
M, Li L and Kai G: Bioactive components and molecular mechanisms of
Salvia miltiorrhiza Bunge in promoting blood circulation to
remove blood stasis. J Ethnopharmacol. 317:1166972023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Y, Liao J, Xiong L, Xiao Z, Ye F, Wang
Y, Chen T, Huang L, Chen M, Chen ZS, et al: Stepwise targeted
strategies for improving neurological function by inhibiting
oxidative stress levels and inflammation following ischemic stroke.
J Control Release. 368:607–622. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Su CY, Ming QL, Rahman K, Han T and Qin
LP: Salvia miltiorrhiza: Traditional medicinal uses,
chemistry, and pharmacology. Chin J Nat Med. 13:163–182.
2015.PubMed/NCBI
|
|
14
|
Zhang W, Feng J, Liu R, Xiang T and Wu X:
Tanshinone IIA regulates NRF2/NLRP3 signal pathway to restrain
oxidative stress and inflammation in Uric Acid-induced HK-2
fibrotic models. Endocr Metab Immune Disord Drug Targets.
25:721–731. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu CH, Chen Y, Jin TY, Wang Z, Jin B, Liao
J, Ding CY, Zhang A, Tang WY, Zhang LX, et al: A derivative of
tanshinone IIA and salviadione, 15a, inhibits inflammation and
alleviates DSS-induced colitis in mice by direct binding and
inhibition of RIPK2. Acta Pharmacol Sin. 46:672–686. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Q, Li X and Luo Y: Tanshinone IIA
delays liver aging by modulating oxidative stress. Front Pharmacol.
15:14340242024. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li Y, Guo Z, Li P, Guo J, Wang H, Pan W,
Wu F, Li J, Zhou J and Ma Z: Tanshinone T1/T2A inhibits non-small
cell lung cancer through Lin28B-let-7-BORA/MYC regulatory network.
Gene. 935:1490582024. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fang ZY, Zhang M, Liu JN, Zhao X, Zhang YQ
and Fang L: Tanshinone IIA: A review of its anticancer effects.
Front Pharmacol. 11:6110872020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang W, Feng J, Liu R, Xiang T and Wu X:
Tanshinone IIA regulates NRF2/NLRP3 signal pathway to restrain
oxidative stress and inflammation in uric acid-induced HK-2
fibrotic models. Endocr Metab Immune Disord Drug Targets.
25:721–731. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu T, Chen J, Zhang M, Tang Z, Tong J,
Hao X, Li H, Xu J and Yang J: Tanshinone IIA exerts
cardioprotective effects through improving Gut-Brain axis
post-myocardial infarction. Cardiovasc Toxicol. 24:1317–1334. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mao D, Wang H, Guo H, Che X, Chen M, Li X,
Liu Y, Huo J and Chen Y: Tanshinone IIA normalized hepatocellular
carcinoma vessels and enhanced PD-1 inhibitor efficacy by
inhibiting ELTD1. Phytomedicine. 123:1551912024. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang P, Gu Y, Lu J, Song M, Hou W, Li P,
Sun Y, Wang J and Chen X: Endothelial TRPV4 channel mediates the
vasodilation induced by Tanshinone IIA. Chem Biol Interact.
402:1111812024. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu HH, Wei W, Wu FF, Cao L, Yang BJ, Fu
JN, Li JX, Liang XY, Dong HY, Heng YY and Zhang PF: Sodium
tanshinone IIA sulfonate protects vascular relaxation in
ApoE-knockout mice by inhibiting the SYK-NLRP3 inflammasome-MMP2/9
pathway. BMC Cardiovasc Disord. 24:3542024. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Xin G, Zhou Q, Yu X, Feng L, Wen
A, Zhang K, Wen T, Zhou X, Wu Q, et al: Elucidating the distinctive
regulatory effects and mechanisms of active compounds in Salvia
miltiorrhiza Bunge via network pharmacology: Unveiling their
roles in the modulation of platelet activation and thrombus
formation. Toxicol Appl Pharmacol. 484:1168712024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu JR, Liu YY, Gao YY, Qian LH, Qiu JL,
Wang PP and Zhang GJ: Diterpenoid tanshinones inhibit gastric
cancer angiogenesis through the PI3K/Akt/mTOR signaling pathway. J
Ethnopharmacol. 324:1177912024. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xuan Y, Yu C, Ni K, Congcong L, Lixin Q
and Qingxian L: Protective effects of tanshinone IIA on
Porphyromonas gingivalis-induced atherosclerosis via the
downregulation of the NOX2/NOX4-ROS mediation of NF-κB signaling
pathway. Microbes Infect. 25:1051772023. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zeng JY, Wang Y, Hong FY, Miao M, Jiang
YY, Qiao ZX, Wang YT and Bao XR: Tanshinone IIA is superior to
paricalcitol in ameliorating tubulointerstitial fibrosis through
regulation of VDR/Wnt/β-catenin pathway in rats with diabetic
nephropathy. Naunyn Schmiedebergs Arch Pharmacol. 397:3959–3977.
2024. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei W, Heng YY, Wu FF, Dong HY, Zhang PF,
Li JX, Liu CY, Yang BJ, Fu JN and Liang XY: Sodium Tanshinone IIA
Sulfonate alleviates vascular senescence in diabetic mice by
modulating the A20-NFκB-NLRP3 inflammasome-catalase pathway. Sci
Rep. 14:176652024. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li H, Hu P, Zou Y, Yuan L, Xu Y, Zhang X,
Luo X and Zhang Z: Tanshinone IIA and hepatocellular carcinoma: A
potential therapeutic drug. Front Oncol. 13:10714152023. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang P, Liu W and Wang Y: The mechanisms
of tanshinone in the treatment of tumors. Front Pharmacol.
14:12822032023. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liang EY, Huang MH, Chen YT, Zhang PW,
Shen Y, Tu XX, Chen WY, Wang Y, Yan J, Wang HY, et al: Tanshinone
IIA modulates cancer cell morphology and movement via Rho
GTPases-mediated actin cytoskeleton remodeling. Toxicol Appl
Pharmacol. 483:1168392024. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang Y, Bi Y, Zhou L, Zheng S, Jian T and
Chen J: Tanshinone IIA inhibits proliferation and migration by
downregulation of the PI3K/Akt pathway in small cell lung cancer
cells. BMC Complement Med Ther. 24:682024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ge T, Li H, Xiang P, Yang D, Zhou J and
Zhang Y: Tanshinone IIA induces ferroptosis in colorectal cancer
cells through the suppression of SLC7A11 expression via the
PI3K/AKT/mTOR pathway. Eur J Med Res. 30:5762025. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ji XZ, Qin X, Wang W and Wang L: A review
of tanshinone compounds in prostate cancer treatment. Transl Androl
Urol. 13:1278–1287. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu L, Gao H, Wen T, Gu T, Zhang S and
Yuan Z: Tanshinone IIA attenuates AOM/DSS-induced colorectal
tumorigenesis in mice via inhibition of intestinal inflammation.
Pharm Biol. 59:89–96. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qian J, Cao Y, Zhang J, Li L, Wu J, Wei G,
Yu J and Huo J: Tanshinone IIA induces autophagy in colon cancer
cells through MEK/ERK/mTOR pathway. Transl Cancer Res. 9:6919–6928.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ma S, Lei Y, Zhang L and Wang J: Research
on the inhibiting effect of tanshinone IIA on colon cancer cell
growth via COX-2-Wnt/β-catenin signaling pathway. J BUON.
23:1337–1342. 2018.PubMed/NCBI
|
|
38
|
Lu Y, Yu T, Liang H, Wang J, Xie J, Shao
J, Gao Y, Yu S, Chen S, Wang L and Jia L: Nitric oxide inhibits
hetero-adhesion of cancer cells to endothelial cells: Restraining
circulating tumor cells from initiating metastatic cascade. Sci
Rep. 4:43442014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Petpiroon N, Sritularak B and
Chanvorachote P: Phoyunnanin E inhibits migration of non-small cell
lung cancer cells via suppression of epithelial-to-mesenchymal
transition and integrin αv and integrin β3. BMC Complement Altern
Med. 17:5532017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
National Research Council Committee for
the Update of the Guide for the C and Use of Laboratory A, . The
National Academies Collection: Reports funded by National
Institutes of Health. Guide for the Care and Use of Laboratory
Animals National Academies Press US. Copyright© 2011, National
Academy of Sciences. Washington, DC: 2011
|
|
41
|
Alessio M, Petiti J, Basile R, Poggio P,
Acquarone D, Scalera A, Avalle L, Orso F, Bertoni A, Porporato PE,
et al: Mitochondrial IκBα fuels cancer progression through
metabolic rewiring, endothelial activation, and thrombotic spread.
Cell Death Discov. Mar 27–2026.(Epub ahead of print). doi:
10.1038/s41420-026-03022-0. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rab SO, Altalbawy FMA, Bishoyi AK, Ballal
S, Kareem M, Singh A, Kubaev A, Soleimani Samarkhazan H and Bagheri
S: Targeting platelet-tumor cell interactions: A novel approach to
cancer therapy. Medical Oncology. 42:2322025. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Souchak J, Mohammed NBB, Lau LS and
Dimitroff CJ: The role of galectins in mediating the adhesion of
circulating cells to vascular endothelium. Front Immunol.
15:13957142024. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jung SM and Moroi M: Activation of the
platelet collagen receptor integrin alpha(2)beta(1): Its mechanism
and participation in the physiological functions of platelets.
Trends Cardiovasc Med. 10:285–292. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen Y, Yang S, Tavormina J, Tampe D,
Zeisberg M, Wang H, Mahadevan KK, Wu CJ, Sugimoto H, Chang CC, et
al: Oncogenic collagen I homotrimers from cancer cells bind to α3β1
integrin and impact tumor microbiome and immunity to promote
pancreatic cancer. Cancer Cell. 40:818–834.e9. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chriqui LE, Cavin S and Perentes JY: Dual
implication of endothelial adhesion molecules in tumor progression
and cancer immunity. Cell Adh Migr. 19:24723082025. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Labelle M and Hynes RO: The initial hours
of metastasis: The importance of cooperative host-tumor cell
interactions during hematogenous dissemination. Cancer Discov.
2:1091–1099. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Geng JG, Chen M and Chou KC: P-selectin
cell adhesion molecule in inflammation, thrombosis, cancer growth
and metastasis. Curr Med Chem. 11:2153–2160. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Barbazán J, Alonso-Alconada L, Elkhatib N,
Geraldo S, Gurchenkov V, Glentis A, van Niel G, Palmulli R,
Fernández B, Viaño P, et al: Liver metastasis is facilitated by the
adherence of circulating tumor cells to vascular fibronectin
deposits. Cancer Res. 77:3431–3441. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mani K, Deng D, Lin C, Wang M, Hsu ML and
Zaorsky NG: Causes of death among people living with metastatic
cancer. Nat Commun. 15:15192024. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ma B, Wells A and Clark AM: The
pan-therapeutic resistance of disseminated tumor cells: Role of
phenotypic plasticity and the metastatic microenvironment. Semin
Cancer Biol. 60:138–147. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lawrence R, Watters M, Davies CR, Pantel K
and Lu YJ: Circulating tumour cells for early detection of
clinically relevant cancer. Nat Rev Clin Oncol. 20:487–500. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jiang L, Yang H, Cheng W, Ni Z and Xiang
N: Droplet microfluidics for CTC-based liquid biopsy: A review.
Analyst. 148:203–221. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Alix-Panabières C, Marchetti D and Lang
JE: Liquid biopsy: From concept to clinical application. Sci Rep.
13:216852023. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Castro-Giner F and Aceto N: Tracking
cancer progression: From circulating tumor cells to metastasis.
Genome Med. 12:312020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chen S, Yang X, Zhang Y, Liu Y, Lu H, Qiu
Y, Cheng L, Li C and Wang C: Inhalable porous microspheres loaded
with metformin and docosahexaenoic acid suppress tumor metastasis
by modulating premetastatic niche. Mol Pharm. 18:2622–2633. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liu Y, Zhang Y, Ding Y and Zhuang R:
Platelet-mediated tumor metastasis mechanism and the role of cell
adhesion molecules. Crit Rev Oncol Hematol. 167:1035022021.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Reymond N, d'Água BB and Ridley AJ:
Crossing the endothelial barrier during metastasis. Nat Rev Cancer.
13:858–870. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shi XY, Bao X, Li Y and Yin CL: Theanine
combined with cisplatin inhibits the proliferation and metastasis
of TNBC cells through Akt signaling pathway. Tradit Med Res.
8:25–30. 2023. View Article : Google Scholar
|
|
60
|
Wang SF, Dong SQ, Dong Q, Lin WX, Dong M
and Liu D: Natural product-induced oxidative stress-synergistic
anti-tumor effects of chemotherapeutic agents. Tradit Med Res.
9:13–18. 2024. View Article : Google Scholar
|
|
61
|
Zhang W, Liu C, Li J, Lu Y, Li H, Zhuang
J, Ren X, Wang M and Sun C: Tanshinone IIA: New perspective on the
Anti-Tumor mechanism of A Traditional Natural Medicine. Am J Chin
Med. 50:209–239. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Luo G, Jin K, Deng S, Cheng H, Fan Z, Gong
Y, Qian Y, Huang Q, Ni Q, Liu C and Yu X: Roles of CA19-9 in
pancreatic cancer: Biomarker, predictor and promoter. Biochim
Biophys Acta Rev Cancer. 1875:1884092021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Schmidt EP, Kuebler WM, Lee WL and Downey
GP: Adhesion molecules: Master controllers of the circulatory
system. Compr Physiol. 6:945–973. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Osmani N, Follain G, García León MJ,
Lefebvre O, Busnelli I, Larnicol A, Harlepp S and Goetz JG:
Metastatic tumor cells exploit their adhesion repertoire to
counteract shear forces during intravascular arrest. Cell Rep.
28:2491–2500.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Maltseva D, Nersisyan A and Tonevitsky A:
Interplay of integrins and selectins in metastasis. Mol Oncol.
19:1582–1611. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang K, Dong S, Higazy D, Jin L, Zou Q,
Chen H, Inayat A, Hu S and Cui M: Inflammatory environment promotes
the adhesion of tumor cells to brain microvascular endothelial
cells. Front Oncol. 11:6917712021. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang Y, Wang J, Yang S, Kou H and Liu P:
Tanshinone IIA alleviate atherosclerosis and hepatic steatosis via
down-regulation of MAPKs/NF-κB signaling pathway. Int
Immunopharmacol. 152:1144652025. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhou C, Yu Z, Chen T, Chen Q, Zhang Y, Cai
J, Xu C and Yu L: Tanshinone IIA attenuates
cerebral-ischemia-reperfusion-induced neuroinflammation by
inhibiting the TLR4/NF-κB signaling cascade: A study integrating
network pharmacology, bioinformatics, and experimental validation.
Phytomedicine. 149:1575482025. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Chen H, Shu H, Su W, Li B, Zhang H, Li L,
Lin C, Yi W, Zhan XY, Chen C, et al: Tanshinone IIA has a potential
therapeutic effect on kawasaki disease and suppresses
megakaryocytes in rabbits with immune vasculitis. Front Cardiovasc
Med. 9:8738512022. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Fang J, Chen Q, He B, Cai J, Yao Y, Cai Y,
Xu S, Rengasamy KRR, Gowrishankar S, Pandian SK and Cao T:
Tanshinone IIA attenuates TNF-α induced PTX3 expression and
monocyte adhesion to endothelial cells through the p38/NF-κB
pathway. Food Chem Toxicol. 121:622–630. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Park EJ, Ji HY, Kim NJ, Song WY, Kim YH,
Kim YC, Sohn DH and Lee HS: Simultaneous determination of
tanshinone I, dihydrotanshinone I, tanshinone IIA and
cryptotanshinone in rat plasma by liquid chromatography-tandem mass
spectrometry: Application to a pharmacokinetic study of a
standardized fraction of Salvia miltiorrhiza, PF2401-SF.
Biomed Chromatogr. 22:548–555. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lyu C, Zhang Y, Zhou W, Zhang S, Kou F,
Wei H, Zhang N and Zuo Z: Gender-Dependent pharmacokinetics of
veratramine in rats: In vivo and in vitro evidence. AAPS J.
18:432–444. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Liu Y, Wang L, Li X, Lv C, Feng D and Luo
Z: Tanshinone IIA improves impaired nerve functions in experimental
diabetic rats. Biochem Biophys Res Commun. 399:49–54. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Li F, Han G and Wu K: Tanshinone IIA
alleviates the AD phenotypes in APP and PS1 transgenic mice. Biomed
Res Int. 2016:76318012016.PubMed/NCBI
|
|
75
|
Jia W, Du F, Liu X, Jiang R, Xu F, Yang J,
Li L, Wang F, Olaleye OE, Dong J and Li C: Renal tubular secretion
of tanshinol: Molecular mechanisms, impact on its systemic
exposure, and propensity for dose-related nephrotoxicity and for
renal herb-drug interactions. Drug Metab Dispos. 43:669–678. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zhu P, Liu Z, Zhou J and Chen Y: Tanshinol
inhibits the growth, migration and invasion of hepatocellular
carcinoma cells via regulating the PI3K-AKT signaling pathway. Onco
Targets Ther. 12:87–99. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Urbanczyk M, Abuhelou A, Köninger M,
Jeyagaran A, Carvajal-Berrio D, Kim E, Marzi J, Loskill P, Layland
SL and Schenke-Layland K: Heterogeneity of endothelial cells
impacts the functionality of human pancreatic in vitro models.
Tissue Eng Part A. 31:988–1004. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Hennigs JK, Matuszcak C, Trepel M and
Körbelin J: Vascular endothelial cells: Heterogeneity and targeting
approaches. Cells. 10:27122021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Uwamori H, Ono Y, Yamashita T, Arai K and
Sudo R: Comparison of organ-specific endothelial cells in terms of
microvascular formation and endothelial barrier functions.
Microvasc Res. 122:60–70. 2019. View Article : Google Scholar : PubMed/NCBI
|