Introduction
Endometrial cancer (EC) has become the most
prevalent gynecologic malignancy, with ~420,000 new cases and
97,000 deaths worldwide in 2022 (1). Critically, EC is among the few
malignancies with rising incidence and mortality trends. Incidence
has increased by 0.7% annually overall and mortality has risen by
1.3–1.6% per year over the past decade (2). This dual increase underscores the
urgent need for improved biomarkers and therapeutic targets to
reverse these trends (3,4). Despite advancements in therapeutic
approaches, persistent challenges such as high recurrence rates and
metastatic potential have led to stagnant survival outcomes,
especially in cases of advanced-stage disease and aggressive
histological subtypes (5–7). These therapeutic challenges have
highlighted the urgent need for novel molecular biomarkers to
facilitate early detection, optimize risk stratification and
develop targeted treatment strategies (8).
Non-SMC condensin I complex subunit H (NCAPH),
localized at chromosome 2q11.2, functions as a key regulatory
component of the condensin I complex important in mitotic
chromosome condensation and faithful sister chromatid segregation
(9,10). Accumulating evidence has indicated
that dysregulation of NCAPH is involved in oncogenesis across
numerous malignancies (11–13). Its upregulation activates
proliferative signaling pathways (including MEK/ERK, β-catenin and
PI3K/Akt) and contributes to the acquisition of aggressive
phenotypes (14–16). NCAPH is also involved in regulating
apoptosis by modulating key signaling pathways, including Akt/mTOR,
checkpoint kinase (Chk)-1/Chk2 and MEK/ERK (14,17,18).
Mechanistically, NCAPH promotes chromosomal instability, DNA damage
tolerance and treatment resistance-features intrinsically
associated with poor prognosis in cancer types such as bladder
carcinoma, advanced colon adenocarcinoma and estrogen
receptor-positive breast cancer (14,19–21).
Recent molecular profiling of endometrial carcinomas has further
identified NCAPH amplification as a characteristic enriched in
high-risk subtypes, suggesting its potential role in the
pathogenesis of EC (22).
Despite these advances, comprehensive analyses
regarding expression patterns, mutational landscape, functional
networks and the clinical utility of NCAPH in endometrial
carcinogenesis remain insufficient. The molecular mechanisms
underlying NCAPH-driven progression of EC, especially its
interactions with cell cycle regulators and its contribution to
genomic instability, are poorly characterized. Furthermore, the
prognostic and diagnostic value of NCAPH in the clinical management
of EC remains unclear.
Subsequently, the present study conducted an
integrated multi-omics investigation of NCAPH in endometrial
cancer. By leveraging bioinformatics analyses supplemented with
experimental validation, the expression dynamics of NCAPH across
different histological subtypes and clinical stages was
systematically evaluated. The present study also characterized the
somatic mutation profiles and co-occurring genomic alterations of
NCAPH, delineated co-expression networks and enriched biological
pathways, investigated its correlation with immune infiltration and
established clinical correlations with survival outcomes and
diagnostic performance. Collectively, the present findings identify
NCAPH as a key molecular driver of EC progression and a promising
biomarker for precision oncology applications.
Materials and methods
Analysis of NCAPH expression
The EC-specific cohort (GDC project ID: TCGA-UCEC)
(23) was obtained from https://portal.gdc.cancer.gov/projects/TCGA-UCEC
and included 546 patients, all female, with a median age of 64
years (range: 31–90 years). Pan-cancer expression data were
obtained through Gene Expression Profiling Interactive Analysis 2
(GEPIA2; http://gepia2.cancer-pku.cn/) (24), which integrates all TCGA tumor
cohorts from the GDC portal (https://portal.gdc.cancer.gov/). The R software (Posit
Software, PBC) package ‘limma’ (version 3.54.0) (25) was employed to compare NCAPH
expression between EC and normal endometrial tissues. In addition,
the associations between NCAPH expression and clinicopathological
parameters (tumor stage and histological subtype) were evaluated
using UALCAN (http://ualcan.path.uab.edu/) (26,27).
Mutational landscape analysis
Somatic mutation data from TCGA-UCEC were processed
using the R package ‘maftools’ (version 2.14.0) (28). Samples were stratified into high/low
NCAPH expression groups based on median values. The top 10
significantly mutated genes (q<0.05) were visualized.
Co-expression and functional
enrichment
Co-expressed genes with NCAPH were identified using
Spearman correlation analysis (P<0.05). Functional enrichment
analysis of the gene sets was conducted using Metascape (http://metascape.org). A false discovery rate
(FDR)<0.05 was considered to indicate a statistically
significant difference for Gene Ontology (GO) terms and Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathways.
Immune cell infiltration analysis
The Tumor Immune Estimation Resource (TIMER) web
server (https://compbio.cn/timer3/) (29) was used to examine the correlations
of NCAPH expression with the abundance of tumor-infiltrating immune
cells, M2 macrophage markers, and programmed death-ligand 1
(PD-L1). Immune infiltration was estimated using the TIMER, XCELL,
CIBERSORT-absolute (ABS) and TIDE (Tracking of Indels by
Decomposition) algorithms, all accessed through the TIMER platform.
Purity-adjusted Spearman's rank correlation test was applied to
calculate correlation coefficients and corresponding P-values.
Survival analysis
Kaplan-Meier survival curves were generated using
the R packages ‘survival’ (version 3.5–7; http://CRAN.R-project.org/package=survival) and
‘survminer’ (version 0.4.9; http://cran.r-project.org/package=survminer), with
log-rank tests applied to assess differences in survival outcomes
between the high and low NCAPH expression groups. Time-dependent
ROC analysis was performed using the ‘timeROC’ package (version
0.4) (30) to evaluate the
diagnostic accuracy at 1, 3 and 5 years.
Single-sample Gene Set Enrichment
Analysis (ssGSEA)
Spliced Transcripts Alignment to a Reference-counts
data and corresponding clinical information from the TCGA-UCEC
cohort were extracted and normalized using the log2
(transcripts per million +1) transformation. Subsequently, the
genes included in the corresponding pathways were collected and
then analyzed using the ‘GSVA’ package in R software (31), with the parameter method ‘ssgsea’
for ssGSEA. Finally, Spearman's correlation analysis was used to
evaluate the correlation between gene expression and pathway
scores.
Immunohistochemical (IHC)
validation
A total of 108 patients (all female; median age 56
years; range: 30–79 years) who underwent surgical resection for
endometrial cancer at The Affiliated Huai'an No.1 People's Hospital
of Nanjing Medical University (Huai'an, China) between January 2018
and June 2023 were enrolled in the present study. The inclusion
criteria were as follows: i) Met EC diagnostic criteria, which was
confirmed by pathological examination; ii) diagnosed and treated at
The Affiliated Huai'an No. 1 People's Hospital of Nanjing Medical
University between 2018 and 2023; and iii) complete
clinicopathologic data were available. The exclusion criteria were
as follows: i) Pathological diagnosis of other uterine tumor types;
and ii) incomplete clinicopathologic data.
Formalin-fixed (10%, room temperature for 24 h)
paraffin-embedded tissue sections (4 µm) from 26 EC specimens and
their paired normal endometrial tissues were subjected to antigen
retrieval (citrate buffer; pH 6.0; 95°C; 15 min). Following
endogenous peroxidase inactivation with 3%
H2O2 at room temperature for 10 min, the
sections were blocked with 5% goat serum (cat. no. G1208; Wuhan
Servicebio Technology Co., Ltd.) at room temperature for 1 h. Then,
the sections were incubated with anti-NCAPH antibody (cat. no.
11515-1-AP; Proteintech Group, Inc.) at a dilution of 1:200
overnight at 4°C, followed by incubation with a HRP-conjugated
secondary antibody (1:5,000; cat. no. bs-0295G; BIOSS) at room
temperature for 1 h. DAB detection kit (cat. no. G1212; Wuhan
Servicebio Technology Co., Ltd.) was used for chromogenic
detection. Then, the slides were examined under a light microscope
(Olympus BX53; Olympus Corporation). The intensity of nuclear
staining was quantified as integrated optical density using ImageJ
software (version 2.1.0/1.53c; National Institutes of Health) and
statistical analyses were performed using GraphPad Prism (version
10.2.0; Dotmatics).
Western blotting analysis
Tissues were lysed by RIPA buffer (cat. no. G2002;
Wuhan Servicebio Technology Co., Ltd.) and quantified by BCA
protein quantitative kit (cat. no. G2026; Wuhan Servicebio
Technology Co., Ltd.). Equal amounts of protein (30 µg per lane)
were separated by 10% SDS-PAGE and transferred onto PVDF membranes.
After blocking with 5% non-fat milk in TBST (0.1% Tween-20) at room
temperature for 1 h, the membranes were incubated overnight at 4°C
with primary antibodies against NCAPH (1:2,000; cat. no.
11515-1-AP; Proteintech Group, Inc.) and GAPDH (1:8,000; cat. no.
60004-1-Ig; Proteintech Group, Inc.). After washing, membranes were
then incubated with HRP-conjugated secondary antibodies: Goat
anti-rabbit IgG (cat. no. SA00001-1; Proteintech Group, Inc.) and
goat anti-mouse IgG (cat. no. SA00001-2; Proteintech Group, Inc.)
for 1 h at room temperature. Finally, the immune complexes were
detected by an ECL kit (cat. no. G2014; Wuhan Servicebio Technology
Co., Ltd.).
Statistical analysis
Comparisons between NCAPH expression and
clinicopathological characteristics were evaluated using Student's
t-tests and logistic regression analysis. The Kaplan-Meier method
was employed to assess the prognostic impact of NCAPH expression.
P<0.05 was considered to indicate a statistically significant
difference.
Results
NCAPH expression is significantly
elevated in pan-cancer and EC
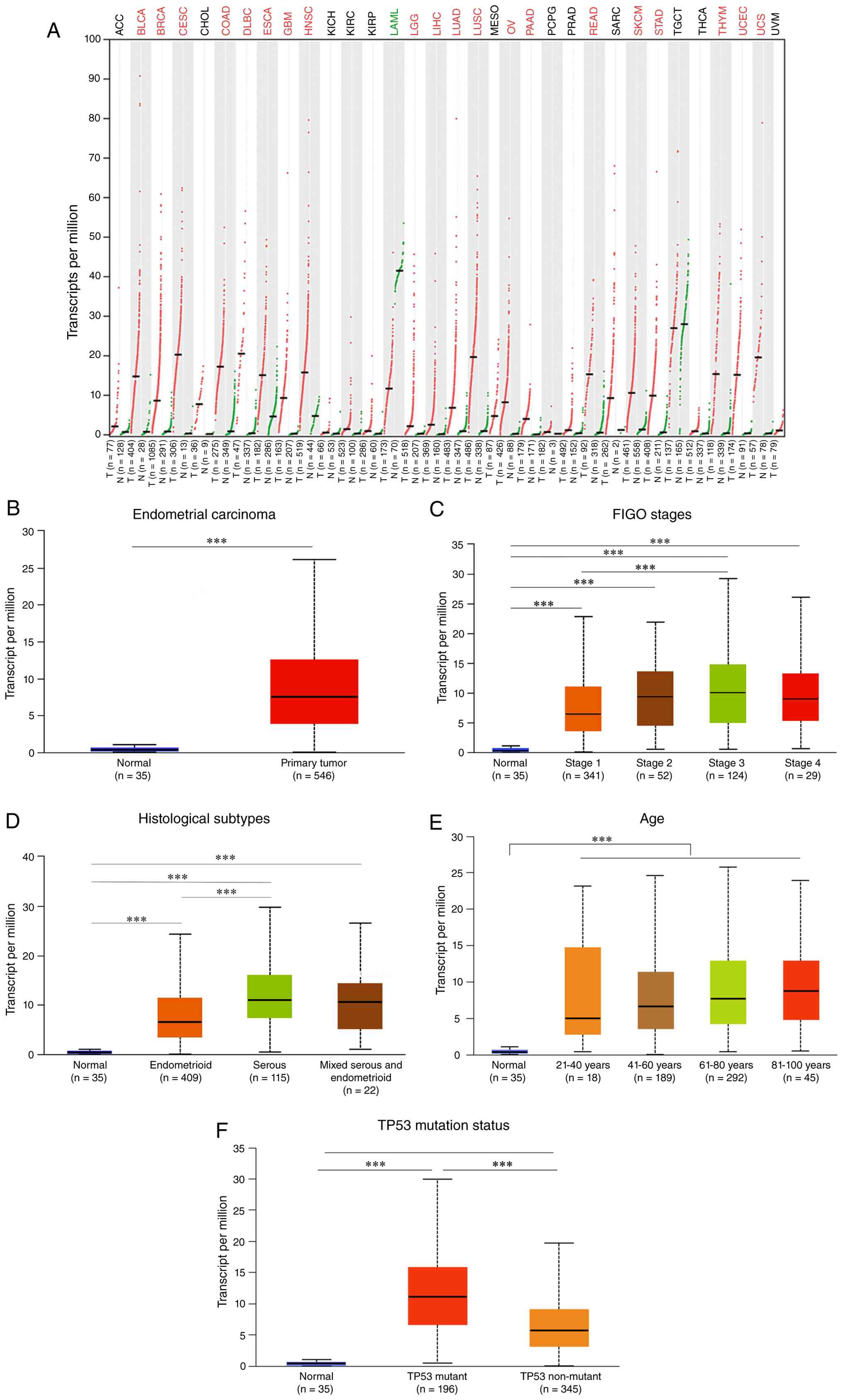
Pan-cancer analysis using GEPIA2 revealed
significant upregulation of NCAPH in numerous malignancies,
including bladder urothelial carcinoma, breast invasive carcinoma,
cervical squamous cell carcinoma, colon adenocarcinoma, diffuse
large B-cell lymphoma, esophageal carcinoma, glioblastoma, head and
neck squamous cell carcinoma, lower-grade glioma, liver
hepatocellular carcinoma, lung adenocarcinoma, lung squamous cell
carcinoma, ovarian cancer, pancreatic adenocarcinoma, rectal
adenocarcinoma, skin cutaneous melanoma, stomach adenocarcinoma,
thymoma, endometrial carcinoma and uterine carcinosarcoma. By
contrast, NCAPH was downregulated in acute myeloid leukemia
(Fig. 1A).
Analysis of the TCGA-UCEC cohort demonstrated
significantly elevated NCAPH expression in 546 endometrial
carcinoma samples compared with 35 normal endometrial samples
(Fig. 1B). Furthermore, high NCAPH
expression was significantly associated with more advanced
International Federation of Gynecology and Obstetrics (FIGO) stages
(Fig. 1C), serous histotype
(Fig. 1D) and older age (Fig. 1E). Given that TP53 mutation is
frequently observed in endometrial carcinoma and is associated with
poor prognosis (32), the
association between NCAPH and TP53 status was also evaluated. NCAPH
expression was found to be positively associated with TP53,
particularly in tumors carrying mutant TP53 (Fig. 1F). Collectively, these results
established NCAPH as a robust molecular marker in EC. Elevated
NCAPH expression was found to be associated with advanced disease
stage and adverse clinical outcomes, highlighting its potential
prognostic utility.
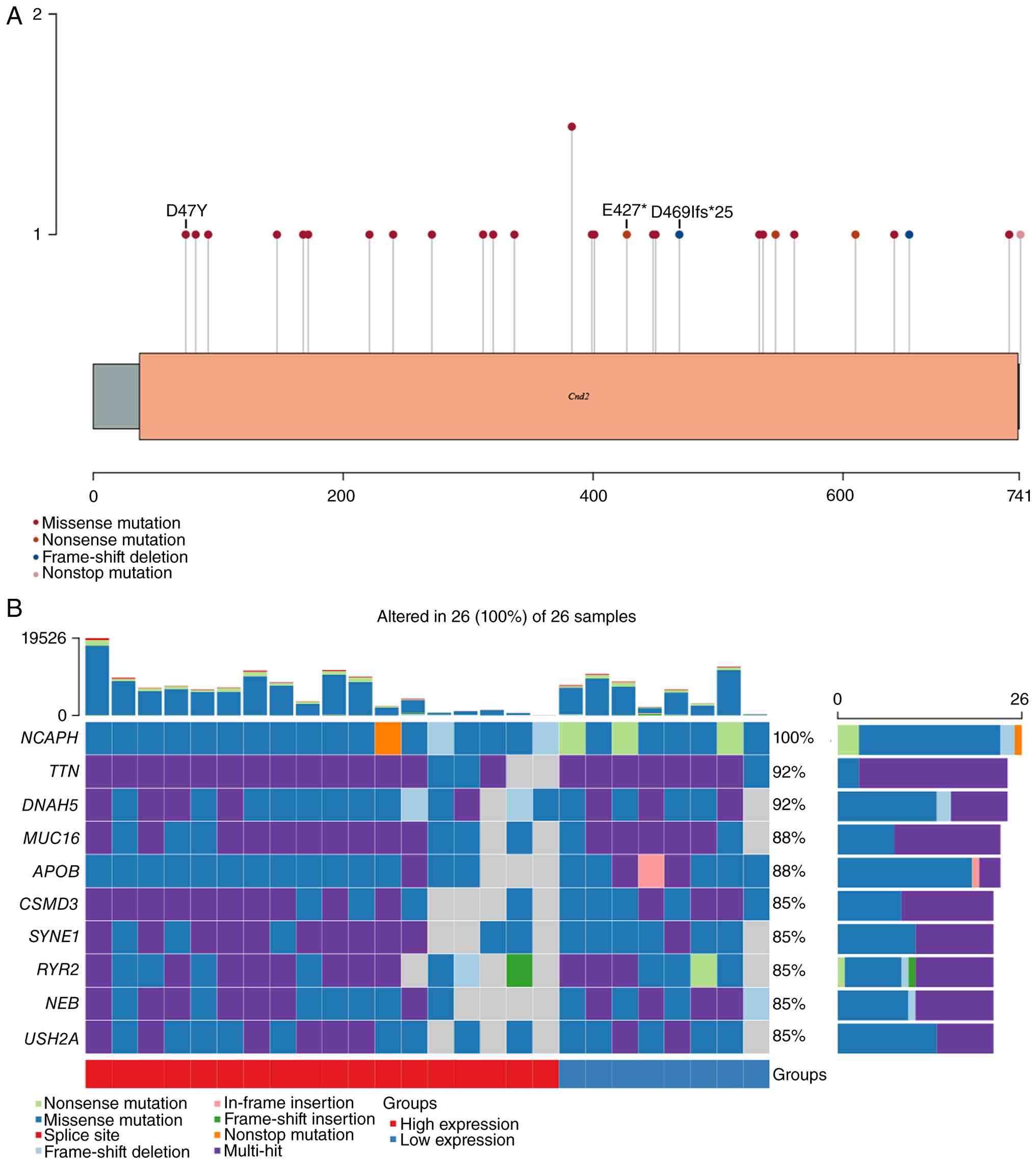
Missense mutations predominate the
NCAPH mutational landscape
Analysis of transcriptome data from 528 endometrial
cancer samples within TCGA-UCEC dataset identified 26 samples
harboring NCAPH mutations, accounting for 4.92% of all analyzed
cases. A total of 29 NCAPH mutations were detected among these 26
samples, with missense mutations being the predominant type,
representing 79.3% of all alterations (Fig. 2A). Besides NCAPH, other genes were
also frequently altered, demonstrating high mutation burdens within
this cohort. The ten most frequently mutated genes across all the
26 samples were NCAPH, TTN, DNAH5, MUC16, APOB, CSMD3, SYNE1, RYR2,
NEB and USH2A (Fig. 2B).
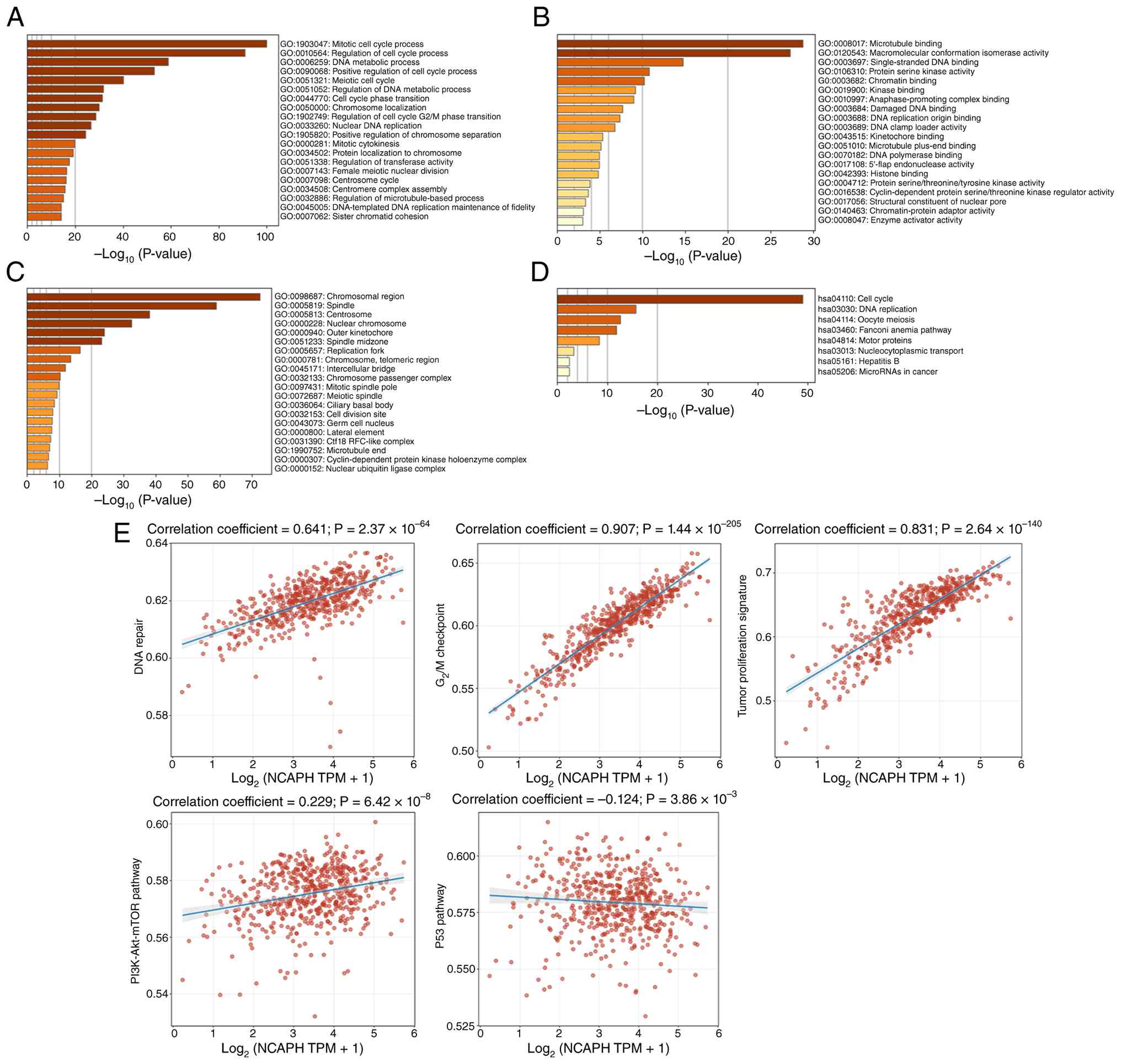
NCAPH co-expressed genes are enriched
in cell cycle and DNA repair pathways and correlate with
PI3K/Akt/mTOR activation and p53 attenuation
To investigate the potential biological functions of
NCAPH in endometrial cancer, the TCGA dataset was analyzed and 189
genes co-expressed with NCAPH were identified (correlation
coefficient ≥0.7). Functional enrichment analysis of these genes
was performed using Metascape. Biological processes were
significantly enriched in ‘mitotic cell cycle process’, ‘regulation
of cell cycle process’ and ‘DNA metabolic process’ (Fig. 3A). Molecular functions showed
primary enrichment in ‘microtubule binding’, ‘macromolecular
conformation isomerase activity’ and ‘single-stranded DNA binding’
(Fig. 3B). Cellular components were
predominantly enriched in the ‘chromosomal region’, ‘spindle’ and
‘centrosome’ (Fig. 3C). KEGG
pathway enrichment analysis further implicated NCAPH in endometrial
cancer pathogenesis through ‘cell cycle’, ‘DNA replication’ and
‘oocyte meiosis’ (Fig. 3D). ssGSEA
was then performed to evaluate correlation between NCAPH expression
and numerous signaling pathways in the TCGA-UCEC cohort. The
results revealed that NCAPH expression was positively correlated
with ‘DNA repair’, ‘G2M checkpoint’, ‘tumor
proliferation’ and the PI3K/AKT/mTOR pathway’ but negatively
associated with ‘p53 pathway’ (Fig.
3E). These findings suggested that NCAPH may promote
endometrial cancer progression through dysregulation of cell cycle
progression and DNA damage response, concomitant with PI3K/AKT/mTOR
pathway activation and p53 pathway attenuation.
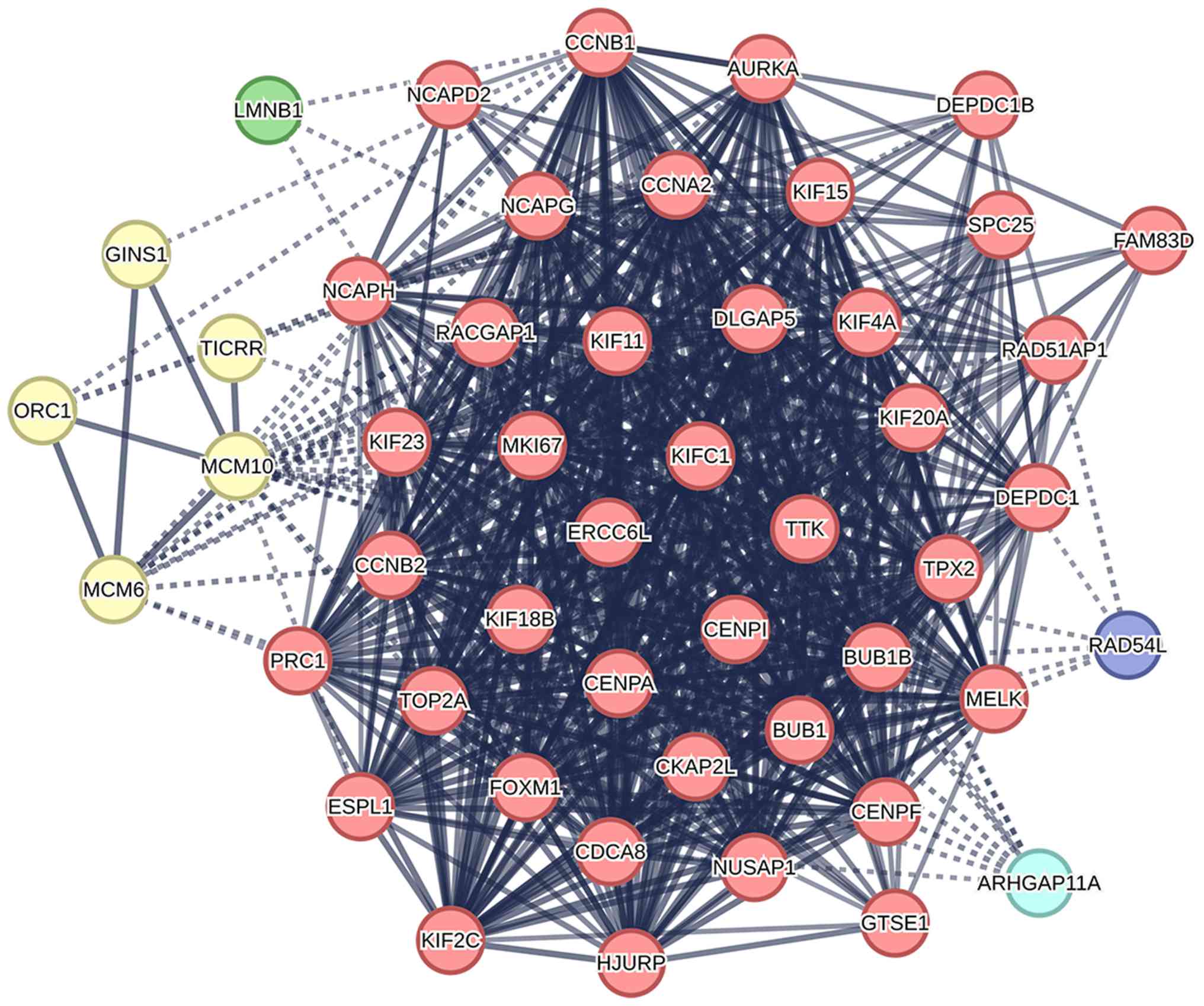
NCAPH interacts with key mitotic
regulators in the protein-protein interaction network
Based on the Search Tool for the Retrieval of
Interacting Genes/Proteins database, the top 50 proteins
co-expressed with NCAPH were selected for PPI network analysis. As
illustrated in Fig. 4, the
resulting network consisted of 50 nodes and 640 edges, with an
average local clustering coefficient of 0.822. The top 10 proteins
most associated with NCAPH were identified as non-SMC condensin I
complex subunit D (NCAPD)-2, non-SMC condensin I complex subunit G
(NCAPG), kinesin family member (KIF)-4A, budding uninhibited by
benzimidazoles (BUB)-1, BUB1B, CDCA8, KIF11, maternal embryonic
leucine zipper kinase (MELK), cellular communication network
(CCN)-B2 and KIF15. Functional clustering analysis revealed that
the majority of these proteins are involved in mitotic sister
chromatid segregation and the mitotic spindle checkpoint signaling
pathway. These findings suggested that dysregulation of NCAPH may
promote tumorigenesis in EC by disrupting mitotic processes.
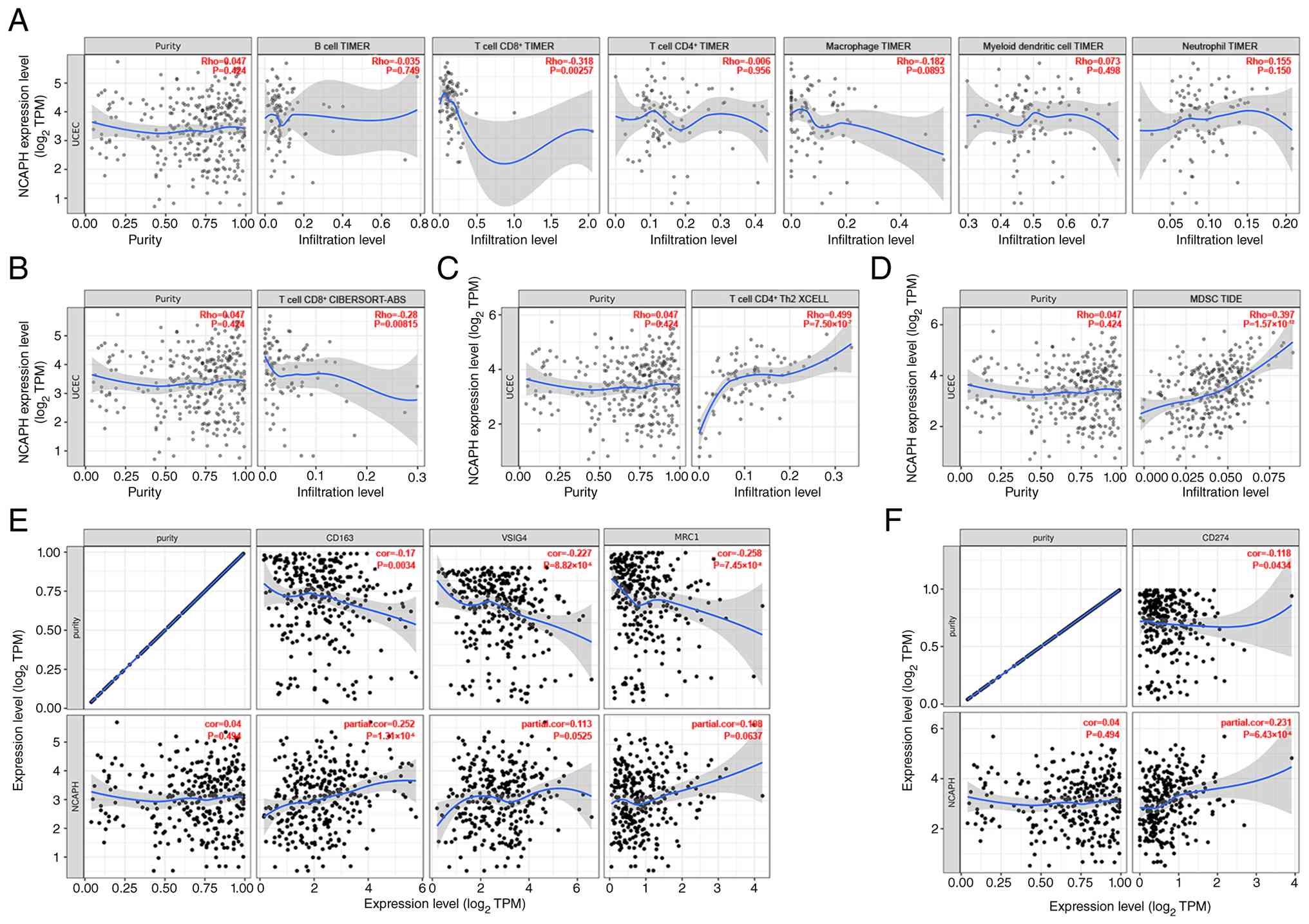
NCAPH expression correlates with an
immunosuppressive microenvironment in EC
To explore the correlation between NCAPH expression
and the abundance of tumor-infiltrating immune cells in EC, the
TIMER database (https://compbio.cn/timer3/) (29) was next used. Analysis revealed a
significant negative correlation between NCAPH expression and the
abundance of CD8+ T cells (R=−0.318; P<0.01; Fig. 5A). To validate these findings,
additional computational methods (XCELL, CIBERSORT-ABS and TIDE)
were employed for immune cell quantification. Consistent with the
TIMER results, the CIBERSORT-ABS method also demonstrated a
negative correlation between NCAPH expression and CD8+ T cell
infiltration (R=−0.28; P<0.01; Fig.
5B). Furthermore, NCAPH expression exhibited a marked positive
correlation with the infiltration of CD4+ T helper (Th)-2 cells
(R=0.499; P<0.001 by XCELL; Fig.
5C) and myeloid-derived suppressor cells (MDSCs; R=0.397;
P<0.001; TIDE; Fig. 5D). No
significant association was observed between NCAPH and regulatory T
cells (Tregs) by either CIBERSORT-ABS or XCELL (Fig. S1).
The present study subsequently investigated whether
NCAPH associates with M2 macrophage markers, since M2 polarization
is a well-recognized mechanism of tumor immune evasion (33). NCAPH exhibited a significant
positive correlation with CD163 (R=0.252; P<0.001); V-set and
immunoglobulin domain-containing 4 (VSIG4; R=0.113; P=0.053) and
mannose receptor C-type 1 (MRC1)/CD206 (R=0.108; P=0.064) trending
in the same direction but not reaching significance (Fig. 5E). This pattern suggested an M2-like
shift in tumor-associated macrophages in the context of high NCAPH
expression in EC.
PD-L1, a key immune checkpoint gene with established
relevance to immunotherapy response (34), was also examined. Results showed
that NCAPH was positively correlated with PD-L1 (CD274; R=0.231;
P<0.001; Fig. 5F). Collectively,
NCAPH-high EC showed fewer infiltrating CD8+ T cells, more CD4+ Th2
cells and MDSCs, M2-skewed macrophage marker expression and higher
PD-L1, collectively pointing to an immunosuppressive
microenvironment. In addition, these findings suggested that
patients with NCAPH-high EC may potentially benefit from immune
checkpoint therapy.
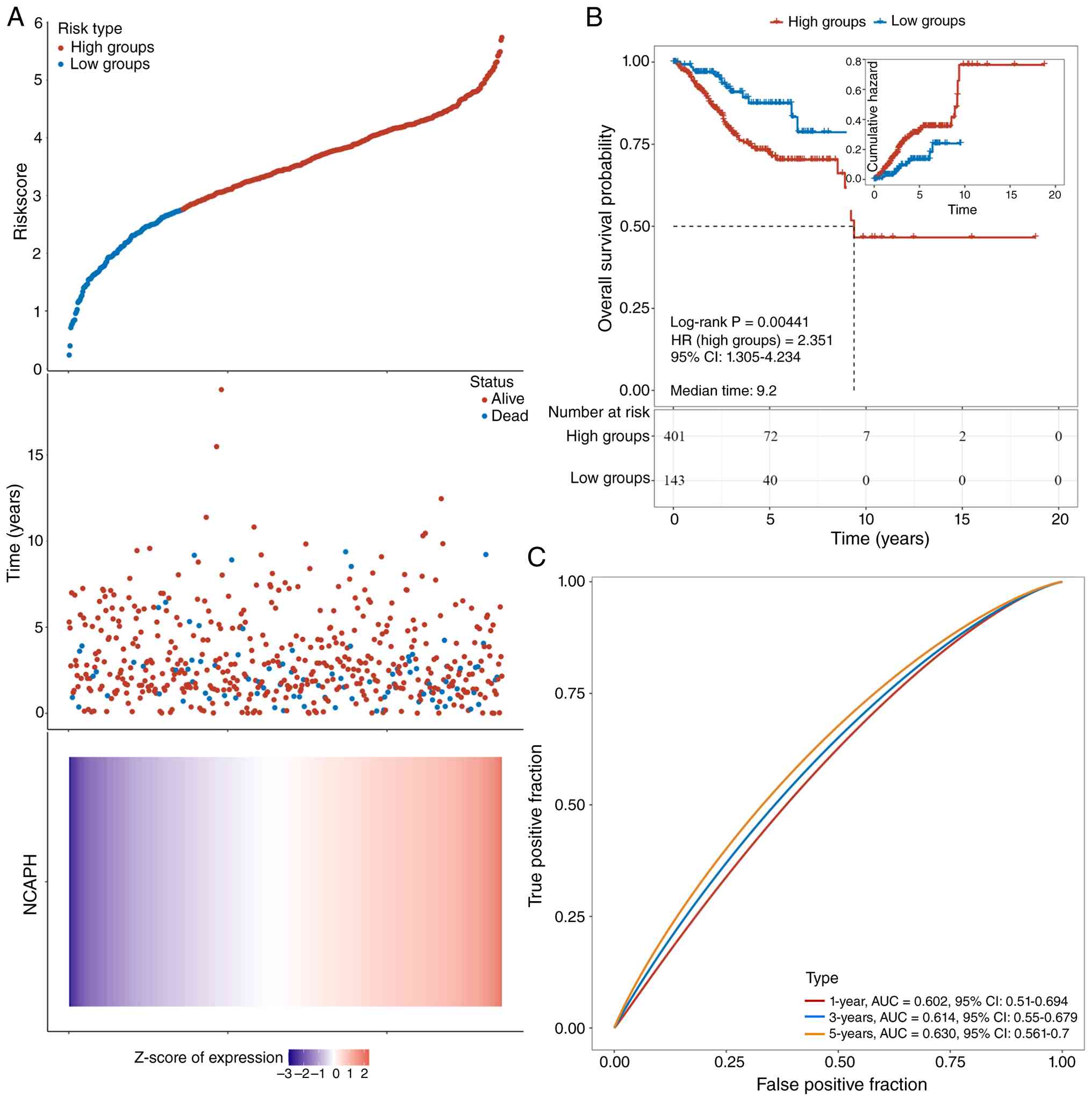
NCAPH serves as a prognostic biomarker
and predicts poor survival in EC
To investigate the association between NCAPH
expression and prognosis, a risk score was computed for each
individual. Using the ‘ggrisk’ and ‘survminer’ R packages, an
optimal cut-off value was determined to stratify patients into a
high-risk group (n=401) and a low-risk group (n=143) (Fig. 6A, upper panel). Fig. 6A (middle panel) shows the survival
status of all patients in the TGCA-UCEC cohort and Fig. 6A (lower panel) presents the
expression level of NCAPH. Kaplan-Meier survival analysis revealed
that patients in the high-risk group had significantly shorter
overall survival compared with those in the low-risk group
(Fig. 6B). To predict the
prognostic value of NCAPH in EC, diagnostic performance was
assessed through time-dependent ROC analysis at 1-, 3- and 5-year
intervals, yielding area under the curve (AUC) values of 0.602,
0.614 and 0.630 respectively (Fig.
6C), indicating moderate but consistent discriminatory capacity
for EC. These results demonstrate that elevated NCAPH expression is
associated with adverse prognosis in EC and may serve as a valuable
prognostic biomarker.
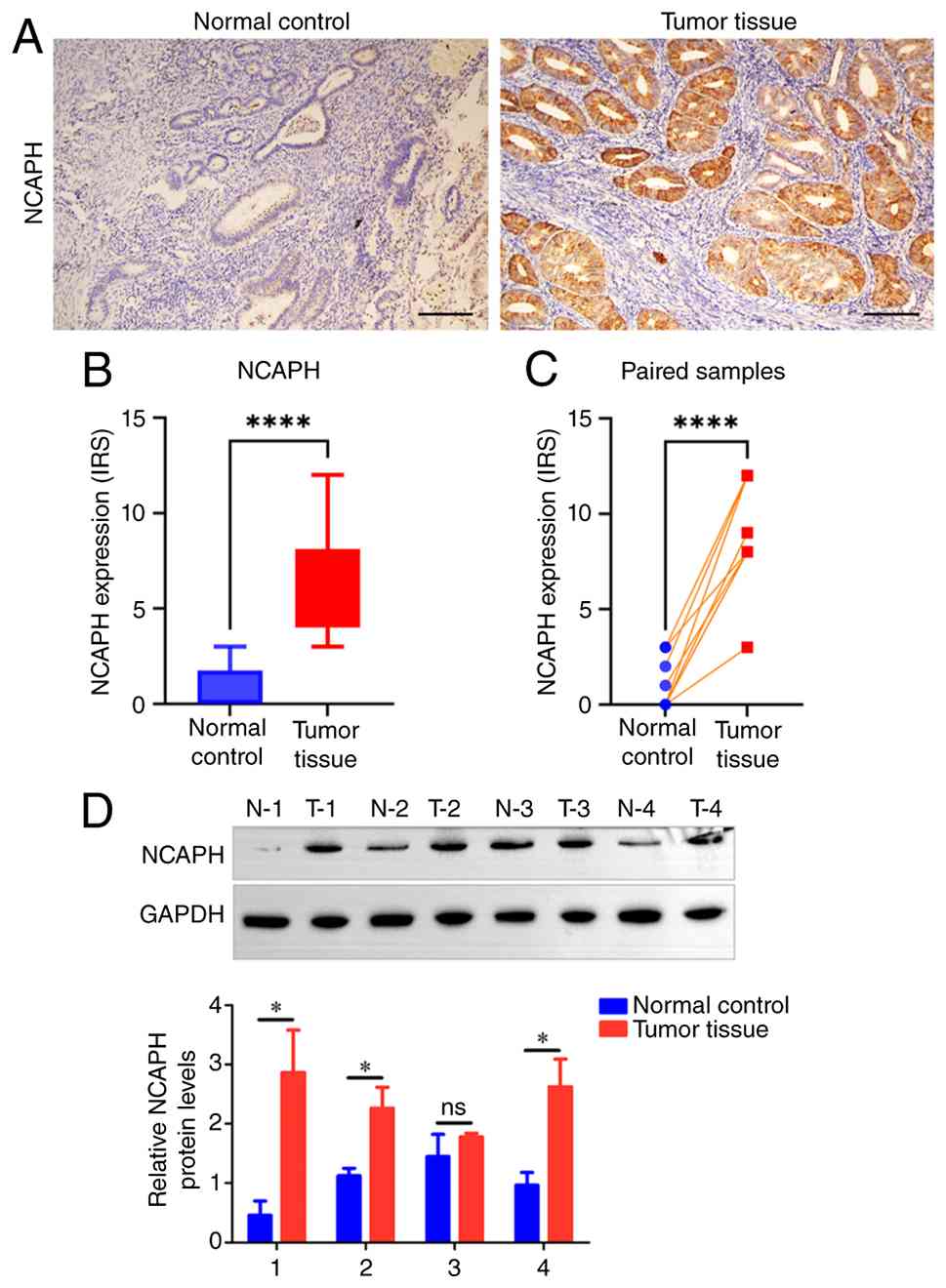
NCAPH protein upregulation in EC is
validated by IHC and western blotting and is associated with
aggressive clinicopathological features
To further validate NCAPH expression in clinical EC
samples, a cohort of 108 patients was retrospectively collected
(all female; median age 56 years; range: 30–79 years) who underwent
surgical resection for EC at The Affiliated Huai'an No. 1 People's
Hospital of Nanjing Medical University (Jiangsu, China) between
January 2018 and June 2023 (ethics approval no. KY-2025-195-01). As
shown in Fig. 7A, NCAPH was
predominantly localized in the cytoplasm of EC tissues. IHC
analysis revealed significantly higher NCAPH expression in EC
samples compared with normal endometrial controls (Fig. 7B). This upregulation was further
determined by paired analysis of tumor tissues and adjacent normal
endometrium (Fig. 7C). Consistent
with the IHC findings, the western blotting results demonstrated
that the NCAPH protein levels were significantly upregulated in EC
tumor tissues relative to normal endometrial controls in 3 of 4
paired samples (N-1 vs. T-1, N-2 vs. T-2 and N-4 vs. T-4;
P<0.05), with the remaining pair showing no significant
difference (N-3 vs. T-3) (Fig. 7D).
Correlations between NCAPH expression and clinicopathological
characteristics in patients with EC were then further evaluated. As
summarized in Table I, elevated
NCAPH levels were significantly correlated with aggressive
histological subtypes, higher tumor grade and advanced FIGO stages.
These results provided robust pathological validation of NCAPH
upregulation in EC and support its potential role in tumor
progression. The correlation with adverse clinicopathological
features underscores the utility of NCAPH as a diagnostic biomarker
and suggested its involvement in EC.
 | Table I.Association between
clinicopathological characteristics and NCAPH expression. |
Table I.
Association between
clinicopathological characteristics and NCAPH expression.
|
|
| NCAPH |
|
|---|
|
|
|
|
|
|---|
| Characteristic | n | Low | High | P-value |
|---|
| Age, years |
|
|
| 0.432 |
|
<45 | 75 | 31 | 44 |
|
|
≥60 | 33 | 11 | 22 |
|
| Histological
subtype |
|
|
| 0.025 |
|
Endometriod | 94 | 41 | 53 |
|
|
Serous | 9 | 0 | 9 |
|
|
Other | 5 | 1 | 4 |
|
| Grade |
|
|
| 0.020 |
| 1 | 29 | 18 | 11 |
|
| 2 | 49 | 18 | 31 |
|
| 3 | 11 | 2 | 9 |
|
| FIGO stage |
|
|
| 0.023 |
| Stage
I | 77 | 36 | 41 |
|
| Stage
II | 21 | 3 | 18 |
|
| Stage
III | 6 | 3 | 3 |
|
| Stage
IV | 3 | 0 | 3 |
|
| LVSI |
|
|
| 0.203 |
|
Negative | 94 | 39 | 55 |
|
|
Positive | 13 | 3 | 10 |
|
Discussion
As a core subunit of the condensin I complex, NCAPH
serves a key role in maintaining the structural integrity of
mitotic chromosomes and ensuring genome stability (35,36).
Previous studies have demonstrated its oncogenic functions across a
number of malignancies (11,12,37);
however, the role of NCAPH in EC remains incompletely
characterized.
The present study provided a comprehensive
multi-omics characterization of NCAPH in EC, integrating genomic,
transcriptomic, proteomic and clinicopathological data to establish
its role as a key oncogenic driver. The present findings
demonstrated that NCAPH expression was significantly upregulated in
a number of cancer types, including EC. Increased NCAPH expression
was associated with aggressive clinicopathological features,
including advanced age, TP53 mutation, higher tumor grade, advanced
FIGO stages and the serous histological subtype. These results
aligned with and extended previous reports of NCAPH dysregulation
in other malignancies such as breast, glioma, gastric, lung and
bladder cancers, supporting its broad oncogenic potential across
tissue types (13,14,35,38,39).
A notable finding of the present study was the
significant positive correlation between NCAPH and TP53,
particularly in tumors harboring mutant TP53. Furthermore, NCAPH
expression was negatively associated with p53 pathway activity,
consistent with the enrichment of TP53 mutations in the NCAPH-high
subgroup. This observation is of marked clinical relevance, given
the well-established role of TP53 mutations in defining high-risk
molecular subtypes of EC (23,40),
particularly serous and copy-number high endometrioid carcinomas,
which are characterized by aggressive clinical behavior,
chromosomal instability and poor prognosis (41,42).
The co-occurrence of elevated NCAPH expression and TP53 mutation
suggested a potential synergistic role in driving genomic
instability and tumor progression. TP53 loss or mutation impairs
cell cycle checkpoint control and DNA damage response, leading to
the accumulation of genetic alterations (43,44).
Meanwhile, NCAPH, as a core condensin subunit, is key in faithful
chromosome condensation and segregation (36,45,46).
Dysregulation of NCAPH may further exacerbate chromosomal
missegregation and aneuploidy, particularly in a TP53-deficient
background where cell cycle surveillance is compromised. This
mechanistic synergy might contribute to the more aggressive
phenotype observed in TP53-mutant EC.
At the genomic level, NCAPH mutations were
identified in 4.92% of EC cases, with a predominance of missense
mutations. Although the mutation rate was modest, the high
frequency of loss-of-function alterations suggested that NCAPH may
contribute to tumorigenesis through haploinsufficiency or
dominant-negative effects, warranting further functional
validation. Condensin I is a multiprotein complex that acts as a
core regulator of mitotic chromosome assembly and faithful
segregation in eukaryotes. It consists of the structural
maintenance of chromosomes (SMC)-2-SMC4 ATPase core, two
HEAT-repeat subunits (NCAPD2/CAP-D2 and NCAPG/CAP-G) and the
kleisin NCAPH that bridges the SMC core to the HEAT-repeat subunits
(47,48). Physical interactions between NCAPH
and the HEAT-repeat subunits are important in chromosome
condensation and DNA repair. Among the 29 NCAPH mutations from 26
samples, it was found that numerous mutations clustered in regions
important for interaction with HEAT-repeat subunits: The missense
mutation D74Y resides within the N-terminal short linear motifs
(~55-77 amino acids) that mediate the autoinhibitory interaction
with NCAPG (49) and truncating
mutations E427* and D469Ifs*25 abolish the entire C-terminal
NCAPG-interaction domain (motif IV; ~460-515 amino acids) (50). Given that the HEAT-repeat subunits
are central to DNA repair and cell cycle control, the
aforementioned mutations are likely to disrupt the physical
interaction between NCAPH and its HEAT-repeat partners, leading to
impaired DNA damage repair (DDR) and cell cycle control. Notably, a
single amino acid substitution in the N-terminal region of Cnd2
(the fission yeast ortholog of NCAPH) has been shown to disrupt
both UV-induced DDR and hydroxyurea-induced cells arrest recovery,
providing genetic evidence that kleisin N-terminal mutations
compromise condensin function in these processes (51). Systematic mutagenesis in fission
yeast has further demonstrated that single-amino-acid substitutions
in condensin subunits, including both the HEAT-repeat components
and the kleisin subunit, confer hypersensitivity to DNA-damaging
agents and cause ploidy maintenance defects (52). Collectively, these findings
suggested that disruption of HEAT-kleisin interactions represents a
potential key mechanism underlying NCAPH mutation-driven
chromosomal instability in EC.
Co-expression and functional enrichment analyses
revealed that NCAPH-associated genes were significantly enriched in
biological processes key in genome integrity, including mitotic
cell cycle regulation, DNA replication as well as DNA and
microtubule binding. These findings were consistent with the
established role of NCAPH in chromosomal condensation and
segregation (10) and further
suggest that its dysregulation may promote EC progression by
inducing chromosomal instability and impaired DDR. PPI analysis
identified strong associations between NCAPH and key mitotic
regulators such as NCAPD2, NCAPG, BUB1 and KIF family members,
reinforcing its central role in cell division. Notably, a number of
these interactors have been implicated in cancer pathogenesis
(53–55) and represent potential co-targets in
NCAPH-driven tumors.
An additional notable finding from the present PPI
network analysis was the enrichment of migration and
invasion-associated proteins within the NCAPH interactome: The
strongest interactor, NCAPG, directly enhances EC invasion through
PI3K/Akt signaling (56). KIF4A,
which competes with NCAPH for NCAPG binding (49), drives epithelial-mesenchymal
transition across numerous cancer types, including glioma and lung,
breast, gastric and colorectal cancer (57). Similarly, MELK, KIF11 and aurora
kinase A each contributes to migration and invasion through
distinct signaling routes. Furthermore, the present KEGG pathway
enrichment analysis determined enrichment of the motor protein
pathway, a key regulatory axis for cell migration and metastatic
progression. This pathway included 12 KIF genes, among which three
(KIF4A, KIF11 and KIF15) were identified as direct binding partners
of NCAPH in the present PPI network. These findings suggested that
NCAPH may drive the pro-invasive phenotype of EC by orchestrating a
prometastatic protein interaction network and modulating motor
protein-associated signaling. In addition, NCAPH has been shown to
activate a number of oncogenic signaling pathways that promote
cancer cell migration and invasion, including PI3K/Akt in glioma
(38) and cervical cancer (58), NF-κB/p65 in lung adenocarcinoma
(59) and Hippo-yes-associated
protein 1 in breast cancer (60).
To explore whether similar associations exist in EC, the present
study performed ssGSEA and observed a positive correlation between
NCAPH expression and PI3K/Akt/mTOR pathway activity, lending
independent support to this association. Collectively, these
results indicated that NCAPH upregulation may promote metastatic
potential in EC through both its network of invasion-associated
partners and directly PI3K/Akt pathway activation.
Accumulating evidence from numerous other cancer
types has demonstrated that NCAPH upregulation is associated with
resistance to platinum-based chemotherapy, including direct
experimental evidence in oral squamous cell carcinoma (61) and bioinformatic identification in
serous ovarian cancer (62). The
underlying mechanisms appear to involve DDR pathways: NCAPH
regulates the expression of key homologous recombination repair
proteins such as RAD54 and p95/nibrin 1 (35) and directly participates in DNA
damage resolution by stabilizing GEN1 for ultra-fine DNA bridge
repair (36). Consistent with these
findings, the present PPI network analysis revealed an enrichment
of DDR proteins within the NCAPH interactome, including direct
interactions with DNA topoisomerase II-α and RAD51AP1. Further
ssGSEA analysis also determined a positive correlation between
NCAPH expression and DNA repair pathway activity, indicating that
NCAPH-high EC possess elevated DNA repair capacity. Notably, the
present PPI network also identified marked overlap between the
NCAPH and RAD51AP1 interactomes, including BUB1, BUB1B, CCNA2,
CCNB2, DLG associated protein 5, KIF11, KIF15, KIF20A, KIF4A, MELK,
NCAPG and TTK protein kinase. These shared interactors are
predominantly involved in mitotic regulation and cell cycle
control, processes associated with DNA damage checkpoint signaling
and repair, suggesting that NCAPH and RAD51AP1 function within the
same biological module. Given that RAD51AP1 has been directly
implicated in chemoresistance in gynecological cancer types
(63,64), it is plausible that NCAPH
upregulation may similarly contribute to chemoresistance in EC.
Collectively, these findings imply that NCAPH upregulation may
enhance DNA repair capacity in EC through a multi-component DDR
axis, thereby promoting resistance to DNA-damaging agents such as
cisplatin and paclitaxel. This NCAPH-DDR axis represents a
promising therapeutic target for overcoming chemoresistance in EC
and warrants further experimental investigation.
A particularly novel aspect of the present study was
the investigation into the association between NCAPH expression and
the tumor immune microenvironment (TME). A significant negative
correlation was observed between NCAPH levels and CD8+ T cell
infiltration, along with a positive correlation with CD4+ Th2
cells, indicating an immunosuppressive TME in EC tumors. No
significant correlation was detected between NCAPH expression and
Treg infiltration using either the CIBERSORT-ABS or XCELL method,
indicating that the immunomodulatory effects of NCAPH in EC are
unlikely to be mediated by the direct expansion of Tregs. Further
analyses regarding canonical M2 macrophage markers (including
CD163, MRC1/CD206 and VSIG4) revealed that NCAPH was positively
correlated with CD163, while VSIG4 and MRC1/CD206 trended in the
same direction but did not reach statistical significance. This
expression profile suggested an M2-like shift in tumor-associated
macrophages in the context of high NCAPH expression in EC and
extends the pan-cancer observation previously reported in breast
cancer (37) to EC. The association
between NCAPH and PD-L1 (CD274) was further evaluated, given its
established role in predicting immunotherapy response (34). NCAPH exhibited a significant
positive correlation with PD-L1, consistent with prior mechanistic
evidence that NCAPH upregulated PD-L1 expression by inhibiting
β-catenin degradation in clear cell renal cell carcinoma (65). The NCAPH-PD-L1 correlation has
direct clinical relevance given recent Food and Drug Association
approval of PD-1/PD-L1 checkpoint inhibitors (pembrolizumab and
dostarlimab) for EC (66–68), suggesting that NCAPH expression
could help identify patients most likely to benefit from
immunotherapy. Furthermore, a strong immune correlation with MDSC
infiltration was also observed. MDSCs are central to tumor immune
evasion as they directly inhibit CD8+ T cell
cytotoxicity, deplete L-arginine through arginase-1 to starve T
cells and drive M2 macrophage polarization. The NCAPH-MDSC
correlation therefore provides a mechanistic association that may
explain numerous features of the NCAPH-associated immune landscape,
the reduced CD8+ T cell infiltration, elevated M2
markers and overall immunosuppression may all converge through
MDSC-mediated effects. Collectively, these analyses characterized
the immunosuppressive role of NCAPH in EC, supported its potential
utility as a candidate biomarker for immunotherapy patient
stratification and highlighted the NCAPH/MDSC/PD-L1 axis as a
promising target for future mechanistic and functional
investigations.
Lastly, the robust prognostic importance of NCAPH
upregulation corroborated by Kaplan-Meier survival analysis and
time-dependent ROC curves, supports its potential utility as a
clinical biomarker for risk stratification. The consistent
diagnostic performance of NCAPH across numerous timepoints
(AUC=0.602, 0.606, 0.628 at 1, 3, 5 years, respectively) further
highlights its reliability in predicting patient outcomes.
Despite these strengths, numerous limitations of the
present study should be acknowledged. The bioinformatics analyses
were conducted primarily on retrospective datasets and although IHC
validation was performed, the sample size was relatively limited.
in addition, the precise molecular mechanisms by which NCAPH
influences mitotic regulation, genomic instability and immune
modulation have not been fully elucidated. In particular, the
crosstalk between NCAPH and TP53 signaling remains poorly defined.
As such, these questions warrant further investigation using both
in vitro and in vivo models. Furthermore, the
potential implications of NCAPH in cancer immunotherapy, though
suggestive, remain largely speculative. Despite the present results
indicating an association between NCAPH expression and an
immunosuppressive microenvironment, it is still unclear whether
NCAPH contributes to primary or acquired resistance to immune
checkpoint inhibitors in EC.
In conclusion, the present integrated analysis
established NCAPH as a multifaceted oncoprotein in EC, contributing
to tumor progression through the dysregulation of mitotic
processes, induction of chromosomal instability and modulation of
the TME. Its significant association with advanced disease stage,
aggressive histologic subtypes and poor survival outcomes
underscores its value as a prognostic biomarker. Furthermore, the
strong immune correlations observed suggested that NCAPH may
influence response to immunotherapy, positioning it as a potential
target for future combination therapeutic strategies. The present
findings provide a compelling rationale for further mechanistic and
translational studies aimed at exploiting NCAPH as both a
predictive biomarker and a therapeutic target in EC.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Zhiyun Xu from
the Affiliated Huai'an No. 1 People's Hospital of Nanjing Medical
University (Nanjing, China) for his guidance on the bioinformatics
analysis.
Funding
The present study was supported by the Northern Jiangsu Clinical
Medicine Research Institute's 2024 Projects (grant no.
HAKY202400405).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
YH was responsible for conceptualization of the
present study, conducting formal analysis, performing the
investigation and writing, reviewing and editing the manuscript. KK
was responsible for conducting formal analysis, performing the
investigation and writing the original manuscript draft. QZ was
responsible for pathological diagnosis confirmation, histological
quality assessment and the resources used. QM was responsible for
IHC staining validation, results interpretation and conducting
formal analysis. YX performed tissue sample collection,
clinicopathological data acquisition and was responsible for the
resources used. YZ and MH conceptualized the present study,
provided supervision and were responsible for writing, reviewing
and editing the manuscript. All authors read and approved the final
version of the manuscript. YH and MH confirm the authenticity of
all the raw data.
Ethics approval and consent to
participate
Ethical approval was granted from the Medical Ethics
Committee of The Affiliated Huai'an No.1 People's Hospital of
Nanjing Medical University (Huai'an First People's Hospital,
Nanjing, China; approval no. KY-2025-195-01). All methods were
performed in accordance with the Declaration of Helsinki. Written
informed consent was waived for this retrospective
non-interventional study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of this work, artificial
intelligence tools were used to improve the readability and
language of the manuscript or to generate images, and subsequently,
the authors revised and edited the content produced by the
artificial intelligence tools as necessary, taking full
responsibility for the ultimate content of the present
manuscript.
References
|
1
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
2
|
Siegel RL, Giaquinto AN and Jemal A:
Cancer statistics, 2024. CA Cancer J Clin. 74:12–49.
2024.PubMed/NCBI
|
|
3
|
Cronin KA, Scott S, Firth AU, Sung H,
Henley SJ, Sherman RL, Siegel RL, Anderson RN, Kohler BA, Benard
VB, et al: Annual report to the nation on the status of cancer,
part 1: National cancer statistics. Cancer. 128:4251–4284. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nierengarten MB: Annual report to the
nation on the status of cancer: This latest report, a collaboration
from the American Cancer Society, the Centers for Disease Control
and Prevention, the National Cancer Institute, and the North
American Association of Central Cancer Registries, shows that
cancer mortality rates have accelerated, and incidence rates remain
stable, with pancreatic cancer showing an increase in both
incidence and mortality: This latest report, a collaboration from
the American Cancer Society, the Centers for Disease Control and
Prevention, the National Cancer Institute, and the North American
Association of Central Cancer Registries, shows that cancer
mortality rates have accelerated, and incidence rates remain
stable, with pancreatic cancer showing an increase in both
incidence and mortality. Cancer. 129:82023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Murali R, Soslow RA and Weigelt B:
Classification of endometrial carcinoma: More than two types.
Lancet Oncol. 15:e268–e278. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Makker V, MacKay H, Ray-Coquard I, Levine
DA, Westin SN, Aoki D and Oaknin A: Endometrial cancer. Nat Rev Dis
Primers. 7:882021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tronconi F, Nero C, Giudice E, Salutari V,
Musacchio L, Ricci C, Carbone MV, Ghizzoni V, Perri MT, Camarda F,
et al: Advanced and recurrent endometrial cancer: State of the art
and future perspectives. Crit Rev Oncol Hematol. 180:1038512022.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Corr B, Cosgrove C, Spinosa D and
Guntupalli S: Endometrial cancer: Molecular classification and
future treatments. BMJ Med. 1:e0001522022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou W, Hu J and Zhao J: Non-SMC condensin
I complex subunit H (NCAPH), a regulator of cell cycle, predicts
poor prognosis in lung adenocarcinoma patients: A study mainly
based on TCGA and GEO database. Transl Cancer Res. 9:7572–7587.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hirano T: Condensins: Organizing and
segregating the genome. Curr Biol. 15:R265–R275. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Song Z, Xia Y, Zhang G, Wang Z, Yang J and
Fan W: NCAPH as an onco-structural hub linking chromatin
architecture to metabolic reprogramming, immune evasion, and
therapy resistance. Crit Rev Oncol Hematol. 222:1052732026.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu C, Han X, Zhang S, Huang M, Guo B,
Zhao Z, Yang S, Jin J, Pu W and Yu H: The role of NCAPH in cancer
treatment. Cell Signal. 121:1112622024. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li C, Meng J and Zhang T: NCAPH is a
prognostic biomarker and associated with immune infiltrates in lung
adenocarcinoma. Sci Rep. 12:95782022. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li B, Xiao Q, Shan L and Song Y: NCAPH
promotes cell proliferation and inhibits cell apoptosis of bladder
cancer cells through MEK/ERK signaling pathway. Cell Cycle.
21:427–438. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang T, Li P, Guo W, Liu Q, Qiao W and
Deng M: NCAPH promotes proliferation as well as motility of breast
cancer cells by activating the PI3K/AKT pathway. Physiol Int.
109:334–347. 2022.PubMed/NCBI
|
|
16
|
Xiong Q, Jiang L, Liu K, Jiang X, Liu B,
Shi Y, Cheng D, Duan Y, Yang C and Chen Y: miR-133b targets NCAPH
to promote beta-catenin degradation and reduce cancer stem cell
maintenance in non-small cell lung cancer. Signal Transduct Target
Ther. 6:2522021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xiong YC, Wang J, Cheng Y, Zhang XY and Ye
XQ: Overexpression of MYBL2 promotes proliferation and migration of
non-small-cell lung cancer via upregulating NCAPH. Mol Cell
Biochem. 468:185–193. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim JH, Youn Y, Kim KT, Jang G and Hwang
JH: Non-SMC condensin I complex subunit H mediates mature
chromosome condensation and DNA damage in pancreatic cancer cells.
Sci Rep. 9:178892019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liao L, Cheng H and Liu S: Non-SMC
condensin I complex subunit H promotes the malignant progression
and cisplatin resistance of breast cancer MCF-7 cells. Oncol Lett.
24:3172022. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ogura T, Azuma K, Sato J, Kinowaki K,
Takayama KI, Takeiwa T, Kawabata H and Inoue S: OCT1 is a poor
prognostic factor for breast cancer patients and promotes cell
proliferation via inducing NCAPH. Int J Mol Sci. 22:115052021.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lei Y, Wang D, Chen W, Tian X and Wei J:
FOXM1/NCAPH activates glycolysis to promote colon adenocarcinoma
stemness and 5-FU resistance. Anticancer Drugs. 34:929–938. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qiu X, Gao Z, Shao J and Li H: NCAPH is
upregulated in endometrial cancer and associated with poor
clinicopathologic characteristics. Ann Hum Genet. 84:437–446. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Levine D; Cancer Genome Atlas Research
Network, : Integrated genomic characterization of endometrial
carcinoma. Nature. 497:67–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang Z, Kang B, Li C, Chen T and Zhang Z:
GEPIA2: An enhanced web server for large-scale expression profiling
and interactive analysis. Nucleic Acids Res. 47:W556–W560. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chandrashekar DS, Karthikeyan SK, Korla
PK, Patel H, Shovon AR, Athar M, Netto GJ, Qin ZS, Kumar S, Manne
U, et al: UALCAN: An update to the integrated cancer data analysis
platform. Neoplasia. 25:18–27. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mayakonda A, Lin DC, Assenov Y, Plass C
and Koeffler HP: Maftools: Efficient and comprehensive analysis of
somatic variants in cancer. Genome Res. 28:1747–1756. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cui H, Zhao G, Lu Y, Zuo S, Duan D, Luo X,
Zhao H, Li J, Zeng Z, Chen Q and Li T: TIMER3: An enhanced resource
for tumor immune analysis. Nucleic Acids Res. 53:W534–W541. 2025.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Blanche P, Dartigues JF and Jacqmin-Gadda
H: Estimating and comparing time-dependent areas under receiver
operating characteristic curves for censored event times with
competing risks. Stat Med. 32:5381–5397. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hanzelmann S, Castelo R and Guinney J:
GSVA: Gene set variation analysis for microarray and RNA-seq data.
BMC Bioinformatics. 14:72013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Moretti NR, Kamitani HZ, de Souza Wagner
PH, Matheus GTFU, Talah BAD, Tanimoto LE, de Oliveira Barretto IC
and de Moraes FCA: Impact of TP53 somatic mutations on prognosis in
endometrial cancer: A systematic review and meta-analysis. Clin
Transl Oncol. 28:1324–1339. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang W, Wang M, Ji C, Liu X, Gu B and
Dong T: Macrophage polarization in the tumor microenvironment:
Emerging roles and therapeutic potentials. Biomed Pharmacother.
177:1169302024. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Doroshow DB, Bhalla S, Beasley MB, Sholl
LM, Kerr KM, Gnjatic S, Wistuba II, Rimm DL, Tsao MS and Hirsch FR:
PD-L1 as a biomarker of response to immune-checkpoint inhibitors.
Nat Rev Clin Oncol. 18:345–362. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang Y, Li JQ, Yang ZL, Wang L, Zhang JC,
Sun YF, Li ZY and Qin L: NCAPH regulates gastric cancer progression
through DNA damage response. Neoplasma. 69:283–291. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim JH, Youn Y and Hwang JH: NCAPH
Stabilizes GEN1 in chromatin to resolve Ultra-fine DNA bridges and
maintain chromosome stability. Mol Cells. 45:792–805. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu Y, Ma X, Feng L, Lin Z and Zhou X: An
integrative pan-cancer analysis reveals the carcinogenic effects of
NCAPH in human cancer. Math Biosci Eng. 20:76–92. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liang J, Yun D, Jin W, Fan J, Wang X, Wang
X, Li Y, Yu S, Zhang C, Li T and Yang X: NCAPH serves as a
prognostic factor and promotes the tumor progression in glioma
through PI3K/AKT signaling pathway. Mol Cell Biochem. 480:589–605.
2025. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mendiburu-Elicabe M, Garcia-Sancha N,
Corchado-Cobos R, Martínez-López A, Chang H, Hua Mao J,
Blanco-Gómez A, García-Casas A, Castellanos-Martín A, Salvador N,
et al: NCAPH drives breast cancer progression and identifies a gene
signature that predicts luminal a tumour recurrence. Clin Transl
Med. 14:e15542024. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Anca-Stanciu MB, Manu A, Olinca MV,
Coroleucă C, Comandașu DE, Coroleuca CA, Maier C and Bratila E:
Comprehensive review of endometrial cancer: New molecular and FIGO
classification and recent treatment changes. J Clin Med.
14:13852025. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Espinosa I, D'Angelo E and Prat J:
Endometrial carcinoma: 10 years of TCGA (the cancer genome atlas):
A critical reappraisal with comments on FIGO 2023 staging. Gynecol
Oncol. 186:94–103. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bogani G, Ray-Coquard I, Concin N, Ngoi
NYL, Morice P, Enomoto T, Takehara K, Denys H, Nout RA, Lorusso D,
et al: Uterine serous carcinoma. Gynecol Oncol. 162:226–234. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tornesello ML: TP53 mutations in cancer:
Molecular features and therapeutic opportunities (Review). Int J
Mol Med. 55:72025. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang H, Guo M, Wei H and Chen Y: Targeting
p53 pathways: Mechanisms, structures, and advances in therapy.
Signal Transduct Target Ther. 8:922023. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hirano T: Condensins: Universal organizers
of chromosomes with diverse functions. Genes Dev. 26:1659–1678.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wood AJ, Severson AF and Meyer BJ:
Condensin and cohesin complexity: The expanding repertoire of
functions. Nat Rev Genet. 11:391–404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Onn I, Aono N, Hirano M and Hirano T:
Reconstitution and subunit geometry of human condensin complexes.
EMBO J. 26:1024–1034. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Schleiffer A, Kaitna S, Maurer-Stroh S,
Glotzer M, Nasmyth K and Eisenhaber F: Kleisins: A superfamily of
bacterial and eukaryotic SMC protein partners. Mol Cell.
11:571–575. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cutts EE, Tetiker D, Kim E and Aragon L:
Molecular mechanism of condensin I activation by KIF4A. EMBO J.
44:682–704. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hara K, Kinoshita K, Migita T, Murakami K,
Shimizu K, Takeuchi K, Hirano T and Hashimoto H: Structural basis
of HEAT-kleisin interactions in the human condensin I subcomplex.
EMBO Rep. 20:e471832019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Aono N, Sutani T, Tomonaga T, Mochida S
and Yanagida M: Cnd2 has dual roles in mitotic condensation and
interphase. Nature. 417:197–202. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xu X, Nakazawa N and Yanagida M: Condensin
HEAT subunits required for DNA repair, kinetochore/centromere
function and ploidy maintenance in fission yeast. PLoS One.
10:e01193472015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cai X, Gao J, Shi C, Guo WZ, Guo D and
Zhang S: The role of NCAPG in various of tumors. Biomed
Pharmacother. 155:1136352022. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kim T and Gartner A: Bub1 kinase in the
regulation of mitosis. Anim Cells Syst (Seoul). 25:1–10. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhong M, Gong L, Li N, Guan H, Gong K,
Zhong Y, Zhu E, Wang X, Jiang S, Li J, et al: Pan-cancer analysis
of kinesin family members with potential implications in prognosis
and immunological role in human cancer. Front Oncol.
13:11798972023. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ren Z, Zheng CY, Xiao X, Yang X, Li X, Cai
H and Fu C: Non-SMC Condensin I complex subunit G promotes the
proliferation, invasion and homologous recombination repair of
endometrial cancer through PI3K/AKT signaling pathway. J Biomed
Nanotechnol. 19:1422–1439. 2023. View Article : Google Scholar
|
|
57
|
Liu Y, Li Y, Tang C, Wen H, Tang J, Chen G
and Wu Y: KIF4A in disease pathogenesis and therapeutics: From
molecular mechanisms to clinical translation. Biol Direct.
20:1162025. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wang M, Qiao X, Cooper T, Pan W, Liu L,
Hayball J, Lin J, Cui X, Zhou Y, Zhang S, et al: HPV E7-mediated
NCAPH ectopic expression regulates the carcinogenesis of cervical
carcinoma via PI3K/AKT/SGK pathway. Cell Death Dis. 11:10492020.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Huang P, Zhao H, Sun R, Liu C, Wu L, Wang
Y, Gan Z, Yang X and Du J: MiR-1976/NCAPH/P65 axis inhibits the
malignant phenotypes of lung adenocarcinoma. Sci Rep. 14:112112024.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Qiu C, Wei Y and Li J: NCAPH-YAP1
interaction promotes breast cancer stemness and tumor progression.
Stem Cell Res Ther. 16:4942025. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shimomura H, Sasahira T, Nakashima C,
Kurihara-Shimomura M and Kirita T: Non-SMC condensin I complex
subunit H (NCAPH) is associated with lymphangiogenesis and drug
resistance in oral squamous cell carcinoma. J Clin Med. 9:722019.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zhan SJ, Liu B and Linghu H: Identifying
genes as potential prognostic indicators in patients with serous
ovarian cancer resistant to carboplatin using integrated
bioinformatics analysis. Oncol Rep. 39:2653–2663. 2018.PubMed/NCBI
|
|
63
|
Huang X, Wu Z, Xiao C and Chen XI:
E2F7/RAD51AP1 axis inhibits endometrial cancer sensitivity to 5-FU
via the fatty acid metabolic pathway. Anticancer Res. 43:4905–4914.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zeng SH, Yan ZQ, Ren Q, Lin LH and Chen Z:
Knocking down RAD51AP1 enhances chemosensitivity by inhibiting the
self-renewal of CD133 positive ovarian cancer stem-like cells.
Discov Oncol. 15:4102024. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chen Z, Ruan W, Guo C, Chen K, Li L, Tian
J, Hu Z, Peng D and Zeng X: Non-SMC condensin I complex subunit H
participates in anti-programmed cell death-1 resistance of clear
cell renal cell carcinomas. Cell Prolif. 56:e134002023. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
O'Malley DM, Bariani GM, Cassier PA,
Marabelle A, Hansen AR, De Jesus Acosta A, Miller WH Jr, Safra T,
Italiano A, Mileshkin L, et al: Pembrolizumab in patients with
microsatellite Instability-High advanced endometrial cancer:
Results from the KEYNOTE-158 study. J Clin Oncol. 40:752–761. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Mirza MR, Chase DM, Slomovitz BM, dePont
Christensen R, Novák Z, Black D, Gilbert L, Sharma S, Valabrega G,
Landrum LM, et al: Dostarlimab for primary advanced or recurrent
endometrial cancer. N Engl J Med. 388:2145–2158. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Powell MA, Bjorge L, Willmott L, Novák Z,
Black D, Gilbert L, Sharma S, Valabrega G, Landrum LM, Gropp-Meier
M, et al: Overall survival in patients with endometrial cancer
treated with dostarlimab plus carboplatin-paclitaxel in the
randomized ENGOT-EN6/GOG-3031/RUBY trial. Ann Oncol. 35:728–738.
2024. View Article : Google Scholar : PubMed/NCBI
|