Introduction
Colorectal cancer (CRC) is the third most common
malignancy and the second leading cause of cancer-related mortality
worldwide (1). In China, CRC has
the second-highest prevalence (after lung cancer) and is the fourth
leading cause of cancer-associated mortalities; of these CRC cases,
49.8% are rectal cancer (2).
Neoadjuvant chemoradiotherapy (nCRT) is a preoperative treatment
for rectal cancer, particularly middle and low locally advanced
rectal cancer (LARC) and is considered the standard neoadjuvant
treatment (3,4). Complete tumor regression following
nCRT is associated with favorable survival outcomes (5). However, the response to nCRT is
heterogeneous, with 12–18% of cases developing a pathological
complete response rate (6,7), whereas 11–20% of cases develop
progressive disease (PD) (8,9).
Therefore, the molecular signatures of nCRT-resistant LARC require
further investigation, providing insights for identifying potential
predictive biomarkers and candidate therapeutic targets to enhance
the precision treatment of LARC.
Owing to advancements in sequencing technologies,
numerous studies have reported the molecular features of nCRT
resistance in LARC. KRAS, neuroblastoma RAS viral oncogene homolog
and BRAF mutations are associated with poor tumor downstaging
(10–12). A genome-based model for adjusting
the radiotherapy dose has been the best-recognized predictive model
for radiosensitivity. The genomic-adjusted radiation dose (GARD)
model has been proposed as a genome-based approach for estimating
the biological effect of radiotherapy. GARD is derived by
integrating the gene-expression-based radiosensitivity index, which
is based on a 10-gene signature (androgen receptor, c-Jun, STAT1,
protein kinase C, RELA proto-oncogene, NF-κB subunit, cABL, small
ubiquitin-like modifier 1, CDK1, histone deacetylase 1 and
interferon regulatory factor 1), with the linear quadratic model
(13). In rectal cancer, GARD
values >17 are associated with improved therapeutic effects
(14). An additional RNA sequencing
study of 114 pre-treatment LARC biopsies revealed that insulin-like
growth factor 2 and L1 cell adhesion molecule (L1CAM) upregulation
was associated with a worse nCRT response (15).
With regard to protein levels, studies have examined
both circulating and tumor tissues to gain further insights
(16–24). Serum samples, collected before and
after nCRT, have been analyzed using techniques including
matrix-assisted laser desorption/ionization time-of-flight
(16) and isobaric tags for
relative and absolute quantitation (iTRAQ) (17), revealing activated immune pathways
in non-responders. Protein signatures in rectal cancer biopsies
were initially detected using phosphorylated protein microarrays
(18). Subsequent studies have used
label-free liquid chromatography mass spectrometry (LC-MS)
(19,20), isobaric tagging MS (21) and data-independent acquisition
(DIA)-MS to identify additional potential biomarkers (22). For rectal tumor tissues before and
after nCRT, a study analyzing samples using iTRAQ MS revealed that
acid ceramidase was associated with the nCRT response (23). An additional study analyzed samples
using label-free LC-MS, suggesting that ADAM metallopeptidase
domain 10 and carbamoyl-phosphate synthetase 2, aspartate
transcarbamylase and dihydroorotase were associated with nCRT
response (24).
Despite the presence of DNA, RNA and proteomic
studies predicting nCRT responses, the selected potential
biomarkers are distinct among studies. This could be due to the
inherent heterogeneity of human cancer types and variations in the
tumor stage at diagnosis. Therefore, investigating the protein
signatures of nCRT-resistant LARC is worthwhile. In the present
pilot study, biopsy and resection tumor tissues were collected from
four nCRT sensitive and four resistant patients with LARC and
subjected to DIA-MS and combined with results of GSE209746
(15) and PXD060201 (25) dataset analysis. Therefore, the aim
of the present study was to identify potential candidate biomarkers
and therapeutic targets for nCRT resistance.
Materials and methods
Tissue sample collection
Formalin-fixed paraffin-embedded (FFPE) tumor tissue
samples from 4 neoadjuvant treatment-sensitive and 4
treatment-resistant patients with LARC (T3-4N0 or any T, N+) who
were treated at The First Affiliated Hospital of Army Medical
University (Chongqing, China) between July 2018 and August 2022
were included in the present study. The cohort comprised of 7 men
and 1 woman, with a median age of 64 years (range: 42–81 years).
From retrospectively screened cases with pelvic MRI available both
before and after nCRT, cases with clear radiological response
categories according to Response Evaluation Criteria in Solid
Tumors (RECIST) version 1.1 (https://project.eortc.org/recist/wp-content/uploads/sites/4/2015/03/RECISTGuidelines.pdf)
and with adequate available FFPE material for pilot DIA-MS analysis
were identified. The tumor staging was recorded according to the
8th edition of the American Joint Committee on Cancer (and Union
for International Cancer Control Staging System for Colorectal
Cancer) (26). Tumor response was
evaluated 4–8 weeks after treatment according to the RECIST
guidelines. Complete response (CR) and partial response (PR) were
categorized as treatment-sensitive, whereas stable disease (SD) and
PD were categorized as treatment-resistant. Response categories in
the proteomic discovery set were defined primarily according to
RECIST guidelines, using pelvic MRI before and after nCRT.
Pathological tumor regression grade was recorded descriptively when
available but was not used for the primary response grouping.
Biopsy tissues were collected before neoadjuvant treatment and
tumor tissues were collected during surgery after neoadjuvant
therapy. The present study was conducted in accordance with the
principles of the Declaration of Helsinki and was approved by the
Ethics Committee of the First Affiliated Hospital of Army Medical
University, PLA [approval no. (A)KY2025105]. Written informed
consent was obtained from all participants or their legal
guardians, where applicable.
Pre-nCRT biopsy samples were obtained through
colonoscopic biopsy. Fresh samples were fixed in 10% neutral
buffered formalin at room temperature for 12–24 h, followed by
routine pathological examination whereby ~4 µm sections were cut
from FFPE blocks and stained with H&E for pathological
determination of adenocarcinoma.
Post-nCRT samples were fresh, unfixed and uncut.
After collection, images of gross samples were routinely captured
and sectioned. To accurately assess tumor regression and account
for intratumoral heterogeneity post-treatment, the entire tumor was
sampled. All specimens were fixed in 10% neutral buffered formalin
(Chongqing Boyi Chemical Reagent Co., Ltd.) at room temperature for
24–48 h, sectioned for routine histopathological examination and 4
µm FFPE sections were stained with H&E for analysis.
H&E staining of both pre-nCRT biopsy specimens
and post-nCRT resection specimens was performed using the same
staining program on an automated slide stainer (Ningbo Chiwell
Biotechnology Co., Ltd.) at room temperature. Briefly, FFPE
sections were deparaffinized in xylene, rehydrated through graded
ethanol and water, stained with hematoxylin for 5 min,
differentiated for 8 sec, blued for 15 sec and counterstained with
eosin for 7 sec. All H&E-stained sections from both pre-nCRT
biopsy specimens and post-nCRT resection specimens were visualized
using an Olympus BX43 microscope (Olympus Corporation).
Protein extraction from FFPE tissue
samples, digestion and preparation
For 15 FFPE rectal cancer tissues from 8 patients (8
before and 7 post-nCRT treatment), 8 sections (10-µm thick) were
cut using a microtome and transferred to 5 ml Eppendorf tubes.
Before protein extraction, FFPE sections were reviewed by an
experienced pathologist, but no macrodissection or laser-capture
microdissection was performed. Proteins were extracted using an
EasyPept FFPE micro sample preparation kit (cat. no. OSFP0006;
Shanghai Omicsolution Co., Ltd.). Briefly, tissue sections were
lysed using 50 µl kit-provided lysis buffer with steel balls and
subjected to 55 Hz grinding for 4 min at 4°C; the samples were then
incubated with agitation at 95°C and 1,000 rpm for 30 min. The
sample was left to stand and after it reached room temperature, the
supernatant was collected after centrifugation (4°C; 12,000 × g; 20
min) and the protein concentration was determined using the BCA
method. A total of 30 µg of protein was incubated at 95°C for 2 h
for reduction and alkylation and cooled to room temperature. The
samples were then digested with the kit-provided protease at 37°C
for 3 h, after which it was centrifuged (room temperature; 20,000 ×
g; 1 min) and the supernatant was collected. The peptide-containing
supernatant was loaded onto a desalting column. The column was then
washed twice and peptides were eluted with 100 µl kit-provided
elution buffer, which was then dried using a JM50-Plus vacuum
freeze dryer (Beijing JM Technology Co., Ltd.) for subsequent
analysis.
Nano-LC-MS/MS analysis
For each sample, 200 ng total peptide was mixed with
a kit containing 11 standard peptides (cat. no. Ki-3002-2;
Biognosys AG) at a 1:10 ratio, then separated using a nanoElute LC
system (Bruker Daltonics GmbH). Peptides were first trapped on a
thermo trap cartridge using mobile phase A (0.1% formic acid in
water) and then separated on a 15 cm reversed-phase C18
PepSep® column (Bruker Daltonics GmbH; particle size:
1.9 µm; inner diameter: 75 µm) maintained at 50°C. Gradient elution
was performed at a flow rate of 0.3 µl/min: Mobile phase B
proportion (0.1% formic acid in acetonitrile) increased from 2 to
24% in the first 30 min, then to 38% in the next 5 min and to 80%
in 7 min (total duration: 42 min) and finally maintained at 80% for
3 min.
Ultra-high-performance LC was coupled to a timsTOF
Pro2 mass spectrometer (Bruker Daltonics GmbH) equipped with a
nano-electrospray ion source operated in positive ion mode. The
nitrogen gas temperature was set to 180°C, the nebuliser pressure
was 2.90 psi and the dry gas flow rate was 3.0 l/min. Data were
acquired in Parallel Accumulation-Serial Fragmentation mode. The
electrospray ion source voltage was set to 1,400 V, precursor
scanning range was 100–1,700 m/z, ion mobility range (1/K0) was
0.6–1.6 Vs/cm2 and both trapped ion mobility
spectrometry ramp time and accumulation time were set to 100 ms,
with a duty cycle close to 100%. The secondary mass spectrometry
scanning range was 400–1,200 m/z and the collision energy increased
linearly with ion mobility (20 eV at 0.85 Vs/cm2 and 59
eV at 1.3 Vs/cm2). The isolation window was set to 25
Th, with a total of 32 windows.
Protein identification and
quantitation
Raw MS files were processed in library-free mode
using Spectronaut® software (version 18.2.230802.50606;
Biognosys AG) with its built-in Pulsar search engine. MS spectra
lists were searched against their species-level UniProt FASTA
database (uniprot_Homo sapiens_9606_reviewed_2024_05_fasta,
http://www.uniprot.org/proteomes/UP000005640), with
carbamidomethylation of cysteine as a fixed modification and
oxidation of methionine and acetylation of protein N-termini as
variable modifications. Trypsin was used as the protease. A maximum
of two missed cleavages was allowed. The false discovery rate at
both peptide and protein levels was set to 0.01. The maximum
allowable initial precursor mass deviation for peptide
identification was 20 ppm and the maximum allowable fragment mass
deviation was 20 ppm. The raw data screening criterion was that
each protein contained at least 1 unique peptide. Since samples
were collected both before and after treatment, protein abundance
may have been downregulated; thus, missing values were replaced
with half of the detected minimum value.
Bioinformatic analysis
Bioinformatic analyses were performed using R
software (version 3.6.3; R Foundation for Statistical Computing)
(27). Principal component analysis
(PCA) was performed using the R package ‘ropls’ (version 1.16.0)
(28) and visualized using
‘ggplot2’ (version 3.3.2) (29).
Proteins were considered differentially abundant proteins (DAPs) if
they met a nominal P<0.05 by Student's t-test and had a fold
change ≥1.2 or ≤0.83. DAPs were visualized using bar plots
generated with ‘ggplot2’. Hierarchical clustering heatmaps were
generated using ‘pheatmap’ (version 1.0.12) (30). Subcellular localization analysis was
performed using ‘loctree3’ (version 1.0.1) (31) and visualized using ‘ggplot2’. Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway analysis was
performed using the KEGG database (http://rest.kegg.jp) (32) and the results were visualized using
‘ggplot2’. The public transcriptomic dataset GSE209746 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE209746)
(15) and public proteomic dataset
PXD060201 (https://www.ebi.ac.uk/pride/archive/projects/PXD060201)
(25) were downloaded and analyzed
for cross-dataset prioritization of candidate proteins associated
with nCRT response.
Immunohistochemistry (IHC) and
assessment of immunostaining intensity
For IHC experiments, a tissue microarray (TMA)
containing 92 CRC and 87 matched adjacent non-tumor tissues was
acquired from Shanghai Outdo Biotech Co., Ltd. (cat. no.
HColA180Su24). IHC staining was conducted using an IHC SP Kit (cat.
no. SP-9001; Beijing Zhongshan Jinqiao Biotechnology Co., Ltd.)
following the manufacturer's instructions. The primary antibody
used was anti-branched-chain keto acid dehydrogenase E1 subunit α
(BCKDHA; cat. no. 30028-1-AP; 1:6,000; Proteintech Group, Inc.)
overnight at 4°C. Staining results were assessed independently by
two pathologists. Staining intensity was classified into four
levels: Negative, score 0; weak, score 1; medium, score 2; and
strong, score 3. The percentage of positively stained cells ranged
from 0–100%. The final IHC score was calculated by multiplying the
percentage of positive cells by the staining intensity. High and
low BCKDHA expression were defined using an IHC score cut-off of
2.375; scores ≤2.375 were classified as low expression, whereas
scores >2.375 were classified as high expression.
Cell culture
The human CRC SW480 cell line was obtained from
American Type Culture Collection (cat. no. CCL-228), grown in
medium supplemented with 10% FBS (Sangon Biotech Co., Ltd.) and
maintained in a humidified 5% CO2 incubator at 37°C,
with cells passaged using standard cell culture techniques.
Generation of stable cell lines
To construct the BCKDHA-overexpressing cell lines,
the BCKDHA coding sequence was synthesized by Beijing Tsingke
Biotech Co., Ltd., and then inserted into the XhoI-NotI sites of
the pLV10ltr-PGK-Hyg-CMV vector (Beijing Tsingke Biotech Co.,
Ltd.). The short hairpin (sh-)RNAs were integrated into the
pLKO.1-mCherry-puro vector (Beijing Tsingke Biotech Co., Ltd.). The
scrambled RNA sequence was
3′-CCGGGGTTCTCCGAACGTGTCACGTCTCGAGACGTGACACGTTCGGAGAACCTTTTTT-5′
and the shBCKDHA sequence was
3′-CCGGGGGTATGGCATCATGTCAATCCTCGAGGATTGACATGATGCCATACCCTTTTTT-5′.
For lentiviral packaging, 5 µg transfer plasmid was co-transfected
with 3.5 µg psPAX2 and 1.5 µg pMD2.G plasmids into 293T cells in
6-cm dishes using polyethylenimine (Yeasen Biotechnology Co.,
Ltd.). The plasmid-PEI complexes were formed at room temperature
for 15 min and then added to 293T cells. After incubation at 37°C
for 48 h, the supernatant was harvested and used to infect SW480
cells. SW480 cells were screened using 200 µg/ml hygromycin for 7
days or 3 µg/ml puromycin for 3 days. The construct efficiency was
determined using a western blotting.
X-ray irradiation
For the colony formation assay and western blotting
analyses after irradiation, cells were seeded into 6-well plates in
2 ml DMEM supplemented with 10% FBS (both Sangon Biotech Co.,
Ltd.). Irradiation was performed using a Varian 600CD machine
(Varian Medical Systems, Inc.) with 6 MV X-rays at a dose rate of 4
Gy/min. The source-to-surface distance was set to 100 cm with a
field size of 37.0×39.4 cm. For dose buildup, the plates were
covered with 1 cm solid water and placed on a 4 cm-thick solid
water slab beneath. Irradiation was delivered at a gantry angle of
0°.
Colony formation assay
SW480 cells were seeded at 1,000 or 2,000 cells per
well in 6-well plates. The following day, cells were exposed to 0,
1, 2, 3, 4 or 5 Gy radiation at a dose rate of 4 Gy/min at room
temperature, corresponding to irradiation durations of 0, 15, 30,
45, 60 and 75 sec, respectively. Cells were then cultured in a
humidified 5% CO2 incubator at 37°C for 10–14 days to
allow colony formation. Colonies were fixed with 4% formaldehyde at
room temperature for 20 min, stained with 0.05% crystal violet, and
analyzed using ImageJ (version 1.54 g; National Institutes of
Health). Colonies containing >50 cells were considered surviving
colonies. The survival fraction was calculated as follows: Planting
efficiency (PE) of untreated (UT) cells (PEUT)=(number
of colonies/number of seeded cells) ×100; survival fraction=number
of colonies/(number of seeded cells × PEUT/100).
Western blotting
Western blotting analysis was performed as
previously described (33). In
brief, cells were lysed in RIPA buffer (cat. no. C510006; Sangon
Biotech Co., Ltd.) containing a protease inhibitor (Roche
Diagnostics GmbH) and incubated on a rocker at 4°C for 15 min. The
protein concentration of the lysates was measured using a BCA
protein kit (Sangon Biotech Co., Ltd.), normalized to equal amounts
of protein (20 µg per lane), resolved on a 10–12% SDS-gel,
transferred to PVDF membranes and probed at 4°C overnight with one
of the following primary antibodies: BCKDHA (cat. no. A21588;
1:1,000), phosphorylated (p)-ataxia telangiectasia mutated (p-ATM;
Ser1981; cat. no. AP1030; 1:500), ATM (cat. no. A19650; 1:500),
p-checkpoint kinase 2 (p-Chk2; Thr68; cat. no. AP0590; 1:1,000),
Chk2 (cat. no. A19543; 1:500), DNA repair protein RAD51 homolog 1
(RAD51; cat. no. A26856; 1:1,000), H2A histone family member X
(γ-H2AX; cat. no. AP1555; 1:1,000), H2AX (cat. no. A11361; 1:1,000)
or β-actin (cat. no. AC026, 1:1,000). All primary antibodies were
purchased from ABclonal Biotech Co., Ltd. After probing with
HRP-conjugated goat anti-rabbit IgG (cat. no. D110058; 1:2,500) or
HRP-conjugated goat anti-mouse IgG (cat. no. D110087; 1:2,500)
secondary antibodies (both Sangon Biotech Co., Ltd.) at room
temperature for 1 h, signals were detected using an ECL reagent
(cat. no. C520045; Sangon Biotech Co., Ltd.).
Statistical analysis
A χ2 test was used to assess the
association between BCKDHA expression and clinicopathological
parameters. Kaplan-Meier analysis and the log-rank test were
performed to assess the association between BCKDHA expression and
overall survival (OS) in patients with CRC. Multivariate survival
analysis was performed using Cox proportional hazards regression
analysis. The survival fraction in the colony formation assay was
analyzed for significance using two-way ANOVA. P<0.05 was
considered to indicate a statistically significant difference.
Results
Clinical characteristics of patients
based on the proteomics analysis
To investigate proteins potentially associated with
nCRT response in LARC, four nCRT-sensitive and four nCRT-resistant
FFPE tumor tissue pairs collected before and after treatment were
analyzed using DIA-MS (Fig. 1A).
The clinical characteristics of the 8 patients are summarized in
Table I. Representative MRI images
of one nCRT-sensitive case and one nCRT-resistant case before and
after nCRT are shown in Fig. 1B.
The patients received intensity-modulated radiation therapy
(Fig. 1C) combined with concurrent
capecitabine chemotherapy. After nCRT, all patients underwent
surgery except one patient with PD, yielding 15 FFPE tumor
specimens for proteomic analysis. Representative H&E staining
is shown in Fig. 1D.
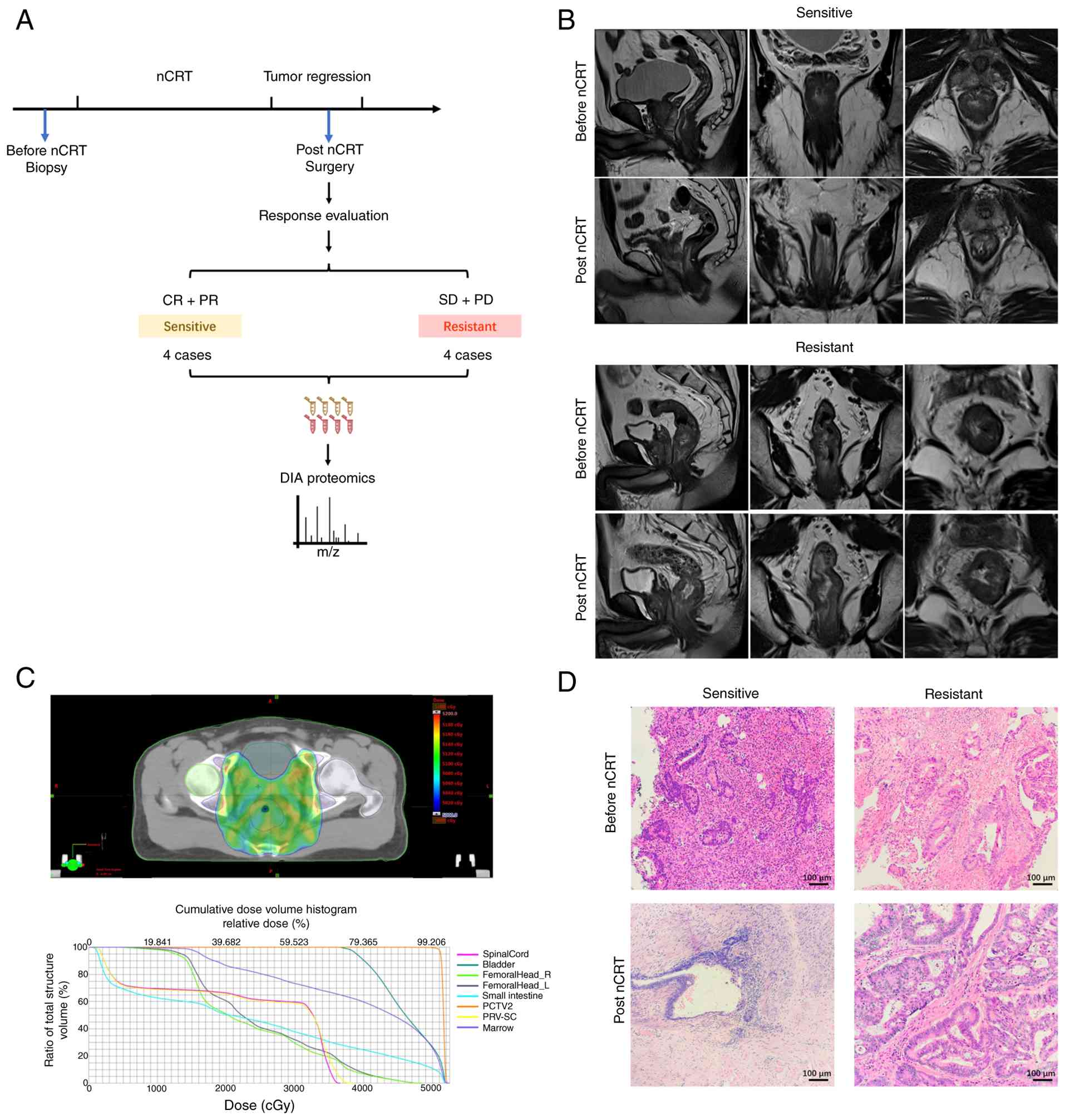 | Figure 1.Selection of nCRT-sensitive and
resistant patients with LARC for proteomic analysis. (A) Schematic
diagram of the present study design and FFPE sample collection
before and after nCRT, followed by DIA proteomics analysis. (B)
Pelvic MRI images of representative nCRT-sensitive (upper panel)
and resistant (lower panel) patients at baseline and post-nCRT.
Sagittal (left), coronal (middle) and transverse (right) sections
are shown. (C) Cumulative dose plan of intensity-modulated
radiation therapy fractions (upper panel) and associated dose
volume histogram (lower panel). (D) Representative H&E staining
images (scale bar, 100 µm). Biopsy specimens before nCRT (upper
panel) and resected tumor tissues after nCRT (lower panel; scale
bar, 100 µm). nCRT, neoadjuvant chemoradiotherapy; LARC, locally
advanced rectal cancer; FFPE, formalin-fixed paraffin-embedded;
DIA, data-independent acquisition; PCTV2, planning clinical target
volume 2; PRV-SC, planning organ at risk volume-spinal cord. |
 | Table I.Clinical characteristics. |
Table I.
Clinical characteristics.
| Sample no. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|
| Response
status | S | S | S | S | R | R | R | R |
| Response
evaluation | PR | PR | PR | PR | PD | PD | SD | SD |
| Mandard TRG | 1 | 2 | 1 | 1 | 3 | N/A | 3 | 3 |
| Sex | M | M | F | M | M | M | M | M |
| Age, years | 64 | 81 | 46 | 64 | 43 | 68 | 64 | 42 |
| Distance from anal
verge, cm | 3.8 | 8.3 | 4.2 | 4.5 | 5.4 | 7.2 | 1.6 | 4.2 |
| cTNM | T4aN0 | T4aN2b | T4aN2b | T4bN2b | T2N2a | T3bN2 | T3bN2 | T3bN2a |
| CRM | - | + | - | + | - | - | - | + |
| EMVI | - | + | + | + | - | + | + | - |
| ypTNM | T3N0 | T3N1a | T3N0 | T2N0 | T3N1c | T4aN0 | T3N0 | T3N1c |
| Radiation dose,
Gy | 50.0 | 50.0 | 50.4 | 50.0 | 50.4 | 50.4 | 50.4 | 50.4 |
| Fractions | 25 | 25 | 28 | 25 | 28 | 28 | 28 | 28 |
Proteomics analysis
DIA-MS identified 39,052 peptides and 6,006
proteins. Samples were labeled by treatment status and response:
Before_sensitive (BS), before_resistant (BR), post_sensitive (PS)
and post_resistant (PR). PCA revealed distinct separation among the
four groups (Fig. 2A). Bar plots of
DAPs are shown in Fig. 2B. To
characterize response-associated DAPs, comparisons between BS with
BR and PS with PR were performed. Under this denominator-first
notation, a higher abundance in the second group corresponded to
the resistant group in these two comparisons. Hierarchical
clustering using proteins meeting the predefined thresholds
separated sensitive and resistant samples both before treatment and
after nCRT (Fig. 2C and D). The
complete list of identified proteins and the DAP lists for the BS
vs. BR and PS vs. PR comparisons are provided in Table SI, Table SII, Table SIII. Given this was a small
proteomic discovery set, these analyses were intended for candidate
prioritization rather than definitive biomarker selection.
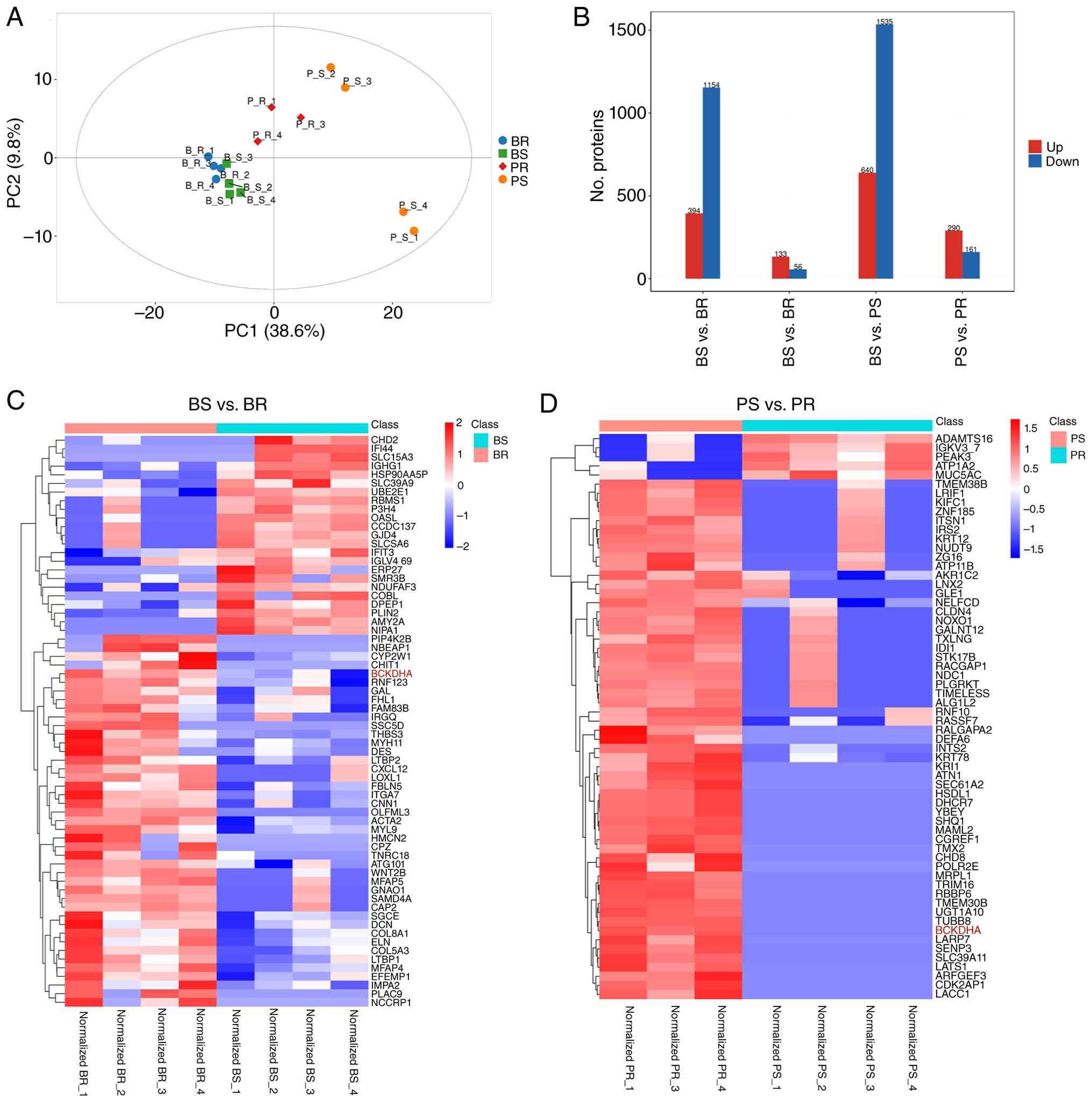 | Figure 2.Overview of the proteomic profiling
results. (A) Principal components analysis identified proteins in
each group. Green, BS; blue, BR; orange, PS; and red, PR. (B) Bar
plot showing the number of DAPs in the four indicated comparisons:
BR vs. PR, BS vs. BR, BS vs. PS and PS vs. PR. DAPs were defined as
those with P<0.05 (Student's t-test) and a fold change of ≥1.2
(upregulated, red) or ≤0.83 (downregulated, blue). In this
denominator-first notation, red indicates higher abundance in the
second group of each comparison and blue indicates higher abundance
in the first group. Hierarchical clustering analysis of DAPs in
comparison to (C) BS vs. BR and (D) PS vs. PR. Each row represents
a protein and each column represents a sample. The relative protein
abundance is shown as a Z-score, with red indicating upregulation
and blue indicating downregulation. BS, sensitive before nCRT; BR,
resistant before nCRT; PS, sensitive post-nCRT; PR, resistant
post-nCRT; DAP, differentially abundant protein; nCRT, neoadjuvant
chemoradiotherapy; Up, upregulated; Down, downregulated. |
Preliminary pathway-level
characterization of proteins more abundant in nCRT-resistant
tumors
To identify candidate proteins associated with nCRT
resistance, proteins that were more abundant in resistant than in
sensitive tumors before nCRT (BS vs. BR) and after nCRT (PS vs. PR)
were examined. Before nCRT, 133 proteins were found to be more
abundant in resistant tumors, primarily localized in the cytoplasm
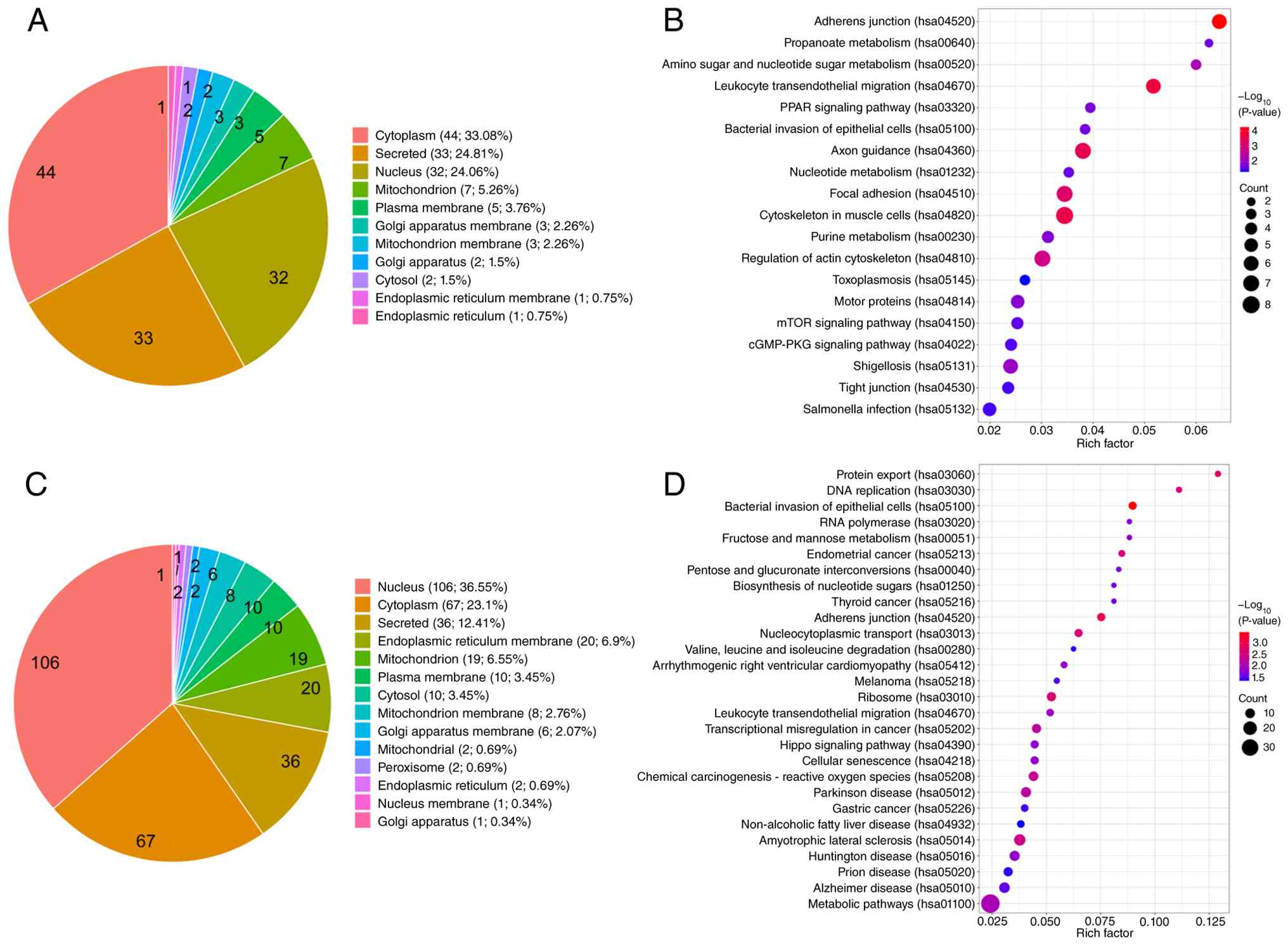
(33.08%), secretory fraction (24.81%) and nucleus (24.06%; Fig. 3A). Preliminary KEGG analysis
indicated enrichment in pathways that include ‘Adherens junction’,
‘Focal adhesion’, ‘Tight junction’, ‘Regulation of actin
cytoskeleton’, ‘Propanoate metabolism’ and ‘Amino sugar and
nucleotide sugar metabolism’ (Fig.
3B). After nCRT, 290 proteins were found to be more abundant in
resistant tumors, with nuclear localization (36.55%) being the most
prominent category (Fig. 3C).
Preliminary enrichment was observed in pathways that include ‘DNA
replication’, ‘RNA polymerase’, ‘Ribosome’, ‘Protein export’,
‘Nucleocytoplasmic transport’ and ‘Metabolic pathways’ (Fig. 3D). Representative proteins of
interest from these comparisons are listed in Tables II and III.
| Table II.Shortlisted upregulated proteins in
resistant tumor before therapy. |
Table II.
Shortlisted upregulated proteins in
resistant tumor before therapy.
| Gene | Protein
accession | P-value | Fold change |
|---|
| BCKDHA | P12694 | 0.008 | 2.159 |
| MIOS | Q9NXC5 | 0.008 | 1.571 |
| MYL9 | P24844 | 0.013 | 2.308 |
| L1CAM | P32004 | 0.049 | 2.397 |
| RRM2 | P31350 | 0.022 | 1.553 |
| GCDH | Q92947 | 0.007 | 1.648 |
| MACC1 | Q6ZN28 | 0.011 | 1.702 |
| PDPK1 | O15530 | 0.048 | 1.263 |
| Table III.Shortlisted upregulated proteins in
resistant tumor post therapy. |
Table III.
Shortlisted upregulated proteins in
resistant tumor post therapy.
| Gene | Protein
accession | P-value | Fold change |
|---|
| BCKDHA | P12694 | 0.008 | 70.848 |
| MIOS | Q9NXC5 | 0.033 | 1.546 |
| MCM2 | P49736 | 0.036 | 1.329 |
| MCM4 | P33991 | 0.009 | 1.429 |
| MCM6 | Q14566 | 0.043 | 1.610 |
| MCM7 | P33993 | 0.034 | 1.669 |
| POLR1E | Q9GZS1 | 0.047 | 3.752 |
| POLR2E | P19388 | 0.008 | 18.374 |
| POLR2I | P36954 | 0.047 | 2.876 |
| RPL5 | P46777 | 0.008 | 1.279 |
| MRPL1 | Q9BYD6 | 0.008 | 148.500 |
| MRPL24 | Q96A35 | 0.047 | 2.674 |
| MARS1 | P56192 | 0.003 | 1.385 |
| PROM1 | O43490 | 0.045 | 2.588 |
| HK2 | P52789 | 0.010 | 1.470 |
| PPAT | Q06203 | 0.027 | 2.754 |
Cross-dataset prioritization
identifies BCKDHA as a candidate resistance-associated protein and
CRC TMA supports its broader clinical relevance
To further prioritize candidate proteins associated
with nCRT response, the public proteomic dataset PXD060201
(25) and transcriptomic dataset
GSE209746 (15) were analyzed.
BCKDHA emerged as a shared candidate across all three datasets
(Fig. 4A). BCKDHA is the E1 subunit
of the branched-chain keto acid dehydrogenase complex, which
catalyzes the catabolism of branched-chain amino acids (34). In the present proteomic discovery
set, BCKDHA exhibited higher abundance in resistant tumors both
before and after nCRT (Fig. 4B and
C). A similar direction of change was observed in PXD060201
(Fig. 4D) and GSE209746 (Fig. 4E).
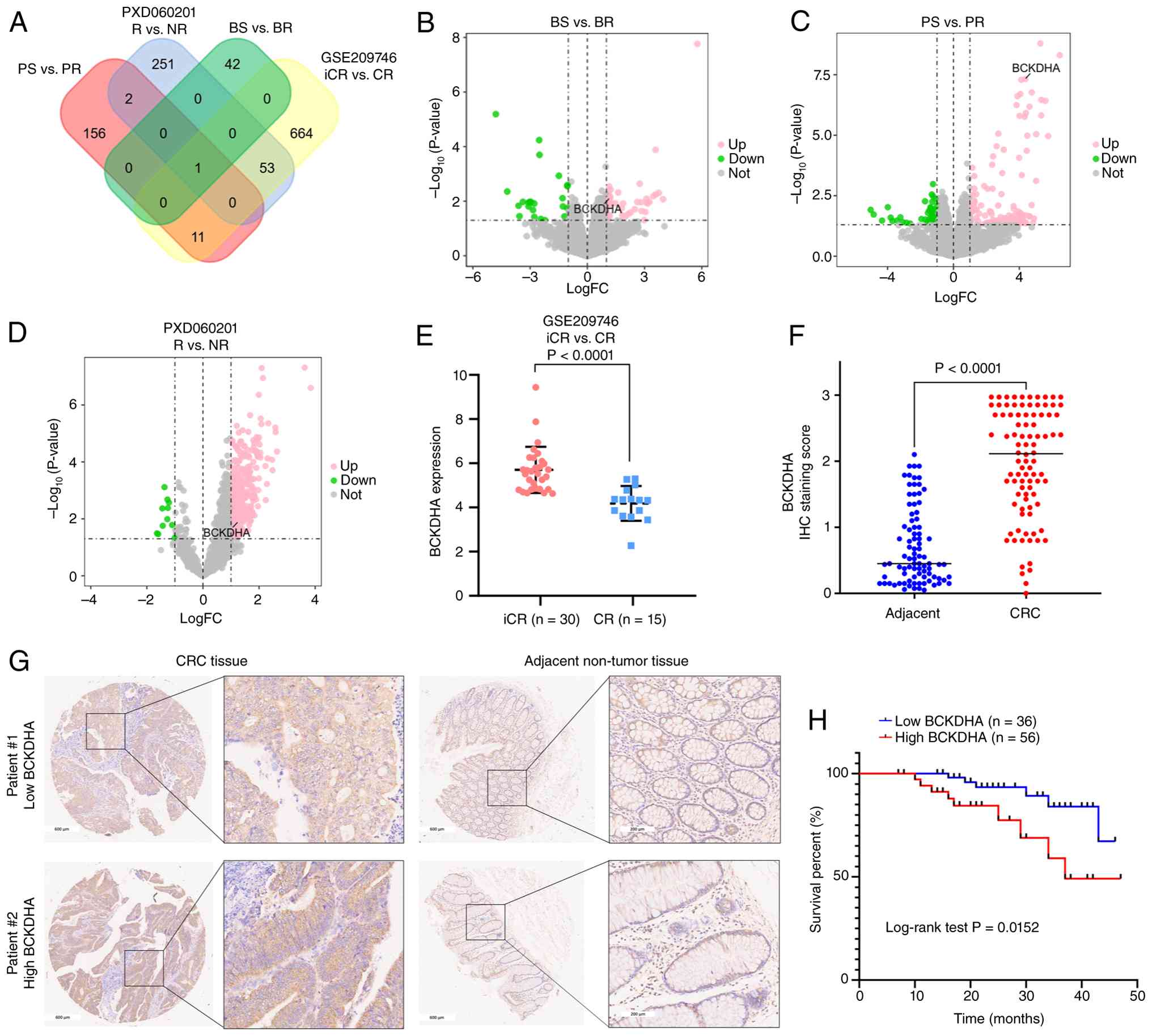 | Figure 4.Cross-dataset prioritization of
BCKDHA and its broader clinical relevance in CRC. (A) Venn diagram
of overlapping potentially associated proteins (P<0.05;
log2FC≥1) among the dataset (BS vs. BR and PS vs. PR),
PXD060201 (R vs. NR) and GSE209746 (iCR vs. CR). BCKDHA was
identified as a consistently upregulated candidate. Volcano plots
of the proteomic data: (B) BS vs. BR and (C) PS vs. PR with BCKDHA
labeled. (D) Volcano plot of PXD060201 with BCKDHA labeled. (E)
BCKDHA mRNA expression in GSE209746. iCR (n=30) and CR (n=15;
P<0.0001). (F) BCKDHA IHC scores in adjacent non-tumor and CRC
tissues (P<0.0001). (G) Representative IHC images: low BCKDHA in
CRC (left upper panel) and adjacent non-tumor tissue (right upper
panel); high BCKDHA in CRC (left lower panel) and adjacent
non-tumor tissue (right lower panel). Scale bars, 600 and 200 µm.
(H) Kaplan-Meier OS curve of patients with CRC stratified by high
BCKDHA expression (n=56) compared with low expression (n=36);
log-rank P=0.0152. BCKDHA, branched-chain keto acid dehydrogenase
E1 subunit α; CRC, colorectal cancer; nCRT, neoadjuvant
chemoradiotherapy; BS, sensitive before nCRT; BR, resistant before
nCRT; PS, sensitive post-nCRT; PR, resistant post-nCRT; R,
responders; NR, nonresponders; iCR, incomplete response; CR,
complete response; IHC, immunohistochemistry; OS, overall survival;
FC, fold change; Up, upregulated; Down, downregulated. |
To further assess its broader clinical relevance,
IHC staining was performed on a TMA containing samples from 92
patients with CRC, with 87 matched adjacent non-tumorous tissues.
BCKDHA localized to the cytoplasm (Fig.
4G) and IHC scores were significantly higher in CRC tissues
compared with adjacent non-tumor tissues (Fig. 4F). The χ2 test (Table IV) showed that high BCKDHA
expression was significantly associated with patient age.
Kaplan-Meier analysis showed that high BCKDHA expression was
associated with shorter OS in patients with CRC (Fig. 4H). Cox proportional hazards
regression analysis (Table V)
revealed that high BCKDHA expression, pathology grade and T stage
were significantly associated with worse OS in univariate analysis.
Multivariate Cox regression showed that high BCKDHA expression was
independently associated with a shorter OS in the CRC cohort.
Collectively, these data support BCKDHA as a candidate protein
associated with nCRT resistance and suggest broader clinical
relevance in CRC.
| Table IV.Correlation between BCKDHA expression
and clinicopathological parameters of patients with colorectal
cancer. |
Table IV.
Correlation between BCKDHA expression
and clinicopathological parameters of patients with colorectal
cancer.
|
|
| BCKDHA
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | All cases
(n=92) | Low (n=36) | High (n=56) | P-value |
|---|
| Pathology
grade |
|
|
| 0.386 |
|
I–II | 58 | 21 | 37 |
|
|
III | 34 | 15 | 19 |
|
| T stage |
|
|
| 0.139 |
|
T1-3 | 69 | 24 | 45 |
|
| T4 | 23 | 12 | 11 |
|
| N stage |
|
|
| 0.615 |
| N0 | 43 | 18 | 25 |
|
|
N1-2 | 49 | 18 | 31 |
|
| M stage |
|
|
| 0.510 |
| M0 | 84 | 32 | 52 |
|
| M1 | 8 | 4 | 4 |
|
| Sex |
|
|
| 0.603 |
|
Male | 48 | 20 | 28 |
|
|
Female | 44 | 16 | 28 |
|
| Age |
|
|
| 0.041a |
|
≤65 | 44 | 22 | 22 |
|
|
>65 | 48 | 14 | 34 |
|
| Table V.Univariate and multivariate Cox
proportional hazards regression analysis for overall survival in
patients with colorectal cancer. |
Table V.
Univariate and multivariate Cox
proportional hazards regression analysis for overall survival in
patients with colorectal cancer.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Variable | HR | 95% CI for HR | P-value | HR | 95% CI for HR | P-value |
|---|
| BCKDHA expression
high | 3.364 | 1.186–9.539 | 0.023a | 2.012 | 1.122–3.608 | 0.019a |
| Pathology
grade | 5.433 | 1.752–16.854 | 0.003b | 5.457 | 1.684–17.684 | 0.005b |
| T stage | 2.992 | 1.100–8.136 | 0.032a | 3.813 | 1.023–9.090 | 0.045a |
| N stage | 1.258 | 0.692–2.287 | 0.451 |
|
|
|
| Distant
metastasis | 0.823 | 0.108–6.299 | 0.851 |
|
|
|
| Sex | 0.493 | 0.178–1.365 | 0.173 |
|
|
|
| Age | 1.025 | 0.982–1.070 | 0.254 |
|
|
|
Preliminary functional data support an
association between BCKDHA and responses to radiotherapy and to DNA
damage response (DDR)-associated signaling in SW480 cells
To examine whether BCKDHA contributed to the
response to radiotherapy, SW480 cell lines with either stable
BCKDHA overexpression or stable BCKDHA knockdown were established.
In BCKDHA overexpressing SW480 cells, colony formation assays
exhibited significantly increased survival after irradiation
(Fig. 5A and B). In GSE209746,
BCKDHA mRNA expression was found to be positively correlated with a
number of DDR-associated genes, including ATM, ATR, BRCA1, BRCA2,
RAD51, RAD52, RPA1, RPA2, RPA3, MRE11 and H2AX (Fig. 5C), supporting a potential
association between BCKDHA and DDR. In BCKDHA-overexpressing SW480
cells, higher levels of ATM, Chk2, RAD51 and H2AX were observed,
together with increased phosphorylation of ATM and Chk2 at 30 min
to 1 h post-irradiation compared with the control cells (Fig. 5D). In addition, BCKDHA
overexpression was also accompanied by earlier induction of γ-H2AX
after irradiation.
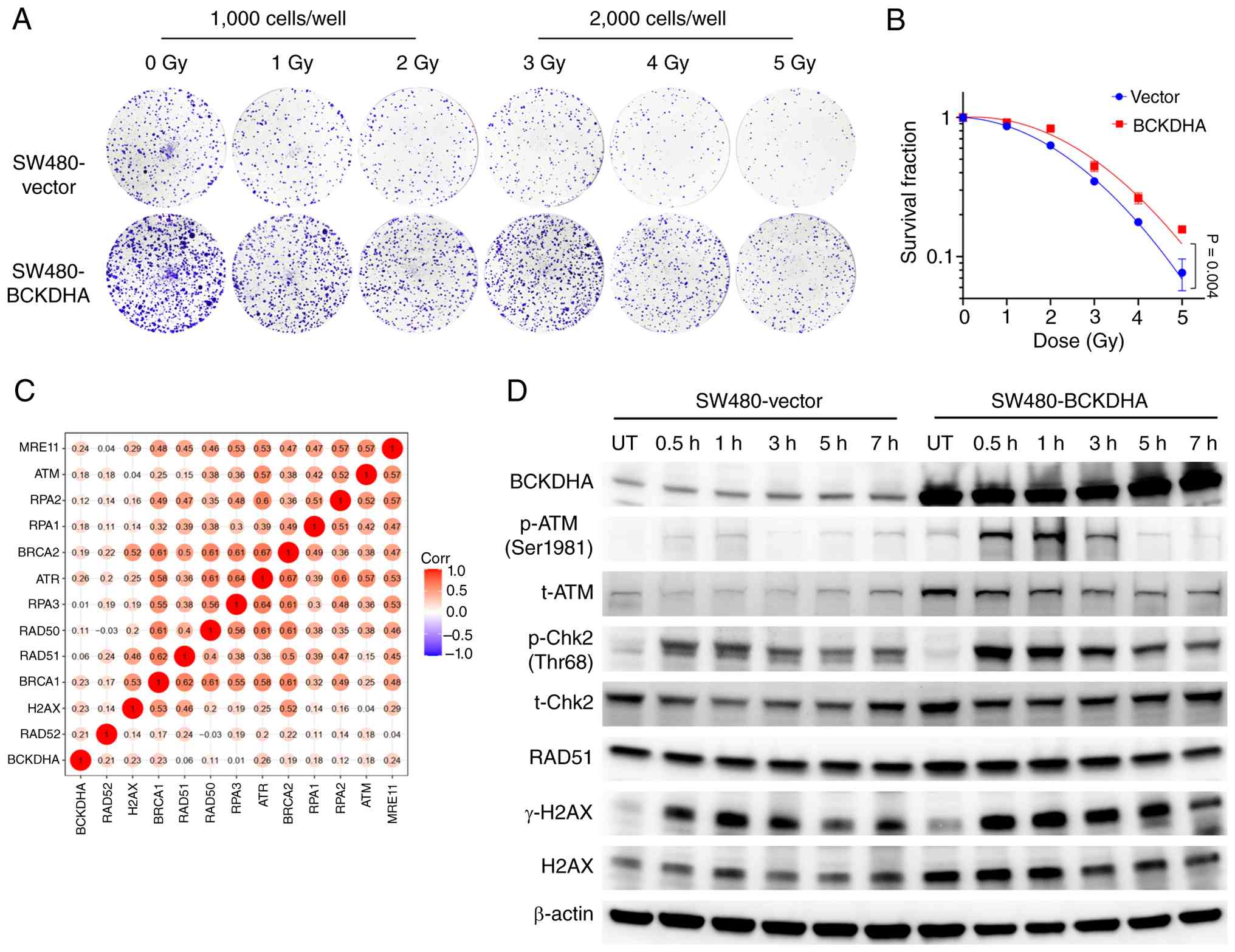 | Figure 5.Preliminary functional evidence
associating BCKDHA overexpression with the response to radiotherapy
and DDR-associated signaling in SW480 cells. Colony formation assay
of SW480 cells stably expressing vector control or BCKDHA. Cells
were seeded at different densities (1,000 or 2,000 cells per well)
to optimize colony formation across a range of irradiation doses
(0–5 Gy). (A) Representative images of crystal violet-stained
colonies. (B) Survival fraction curves plotted from colony counts.
Data are presented as the mean ± SD from three independent
experiments (P=0.004 using two-way ANOVA). (C) Spearman correlation
analysis between BCKDHA mRNA expression and DDR-associated genes in
the GSE209746 dataset. Color intensity indicates correlation
coefficient; values labeled on the heatmap represent Spearman's r
values. (D) Western blotting analysis of BCKDHA, p-ATM, t-ATM,
p-Chk2, t-Chk2, RAD51, γ-H2AX and H2AX in vector control and
BCKDHA-overexpressing SW480 cells, UT or at 0.5, 1, 3, 5 and 7 h
after 2 Gy X-ray irradiation. β-actin served as a loading control.
BCKDHA, branched-chain keto acid dehydrogenase E1 subunit α; DDR,
DNA damage response; p, phosphorylated; t, total; ATM, ataxia
telangiectasia mutated; Chk2, checkpoint kinase 2; H2AX, H2A
histone family member X; UT, untreated; Corr, correlation. |
By contrast, stable BCKDHA knockdown significantly
reduced colony formation after irradiation (Fig. 6A and B). BCKDHA knockdown was
accompanied by lower levels of ATM, Chk2, RAD51 and H2AX,
attenuated radiation-induced phosphorylation of ATM and Chk2 and
lower γ-H2AX levels across the examined time points
post-irradiation (Fig. 6C).
Collectively, these findings provide preliminary functional
evidence that BCKDHA overexpression is associated with increased
clonogenic survival and a more resistant phenotype after
irradiation, whereas BCKDHA knockdown is associated with reduced
clonogenic survival and enhanced radiosensitivity in SW480
cells.
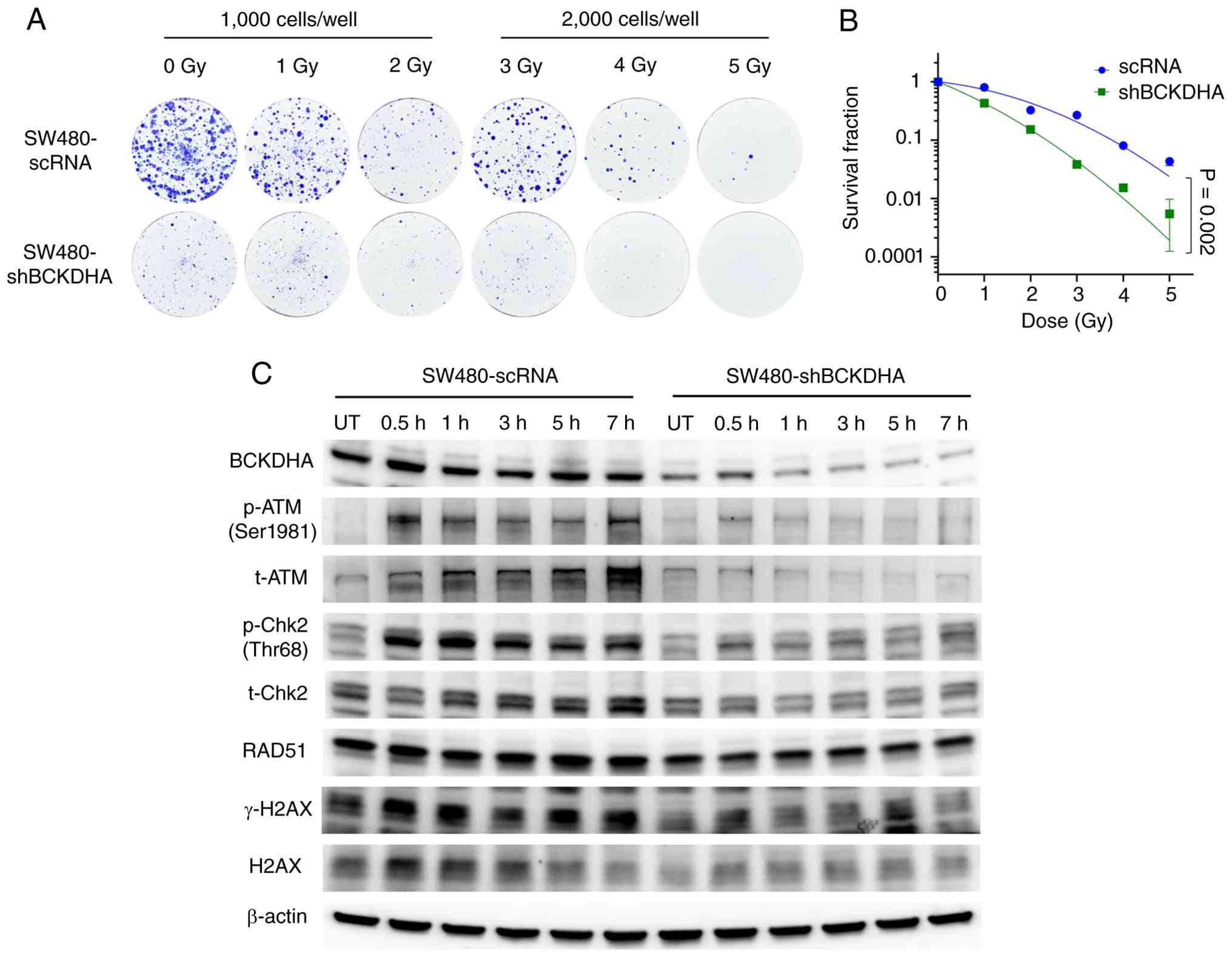 | Figure 6.Preliminary functional evidence
associating BCKDHA knockdown with increased radiosensitivity and
altered DNA damage response-associated signaling in SW480 cells.
(A) Representative images of crystal violet-stained colonies of
SW480-scRNA and SW480-shBCKDHA cells after 0, 1, 2, 3, 4 and 5 Gy
X-ray irradiation. (B) Survival fraction curves plotted from colony
counts. Data are presented as the mean ± SD from three independent
experiments (P=0.002 using two-way ANOVA). (C) Western blotting
analysis of BCKDHA, p-ATM, t-ATM, p-Chk2, t-Chk2, RAD51, γ-H2AX and
H2AX in SW480-scRNA and SW480-shBCKDHA cells, UT or at 0.5, 1, 3, 5
and 7 h after 2 Gy X-ray irradiation. β-actin served as a loading
control. BCKDHA, branched-chain keto acid dehydrogenase E1 subunit
α; scRNA, scrambled RNA; shRNA, short hairpin RNA; ATM, ataxia
telangiectasia mutated; Chk2, checkpoint kinase 2; H2AX, H2A
histone family member X; UT, untreated; t-, total; p-,
phosphorylated. |
Discussion
In the present pilot study, DIA-MS was used to
profile FFPE tumor specimens collected before and after nCRT from
four cases each of sensitive and resistant LARC. A total of 6,006
proteins were identified as potentially relevant and used in a
small proteomic discovery set to prioritize candidate proteins
associated with response categories. Before nCRT, proteins more
abundant in resistant tumors were preliminarily enriched in
pathways such as ‘Adherens junction’, ‘Focal adhesion’, ‘Tight
junction’, ‘Regulation of actin cytoskeleton’, ‘Propanoate
metabolism’ and ‘Amino sugar and nucleotide sugar metabolism’.
After nCRT, proteins more abundant in resistant tumors were
preliminarily enriched in pathways such as ‘DNA replication’, ‘RNA
polymerase’, ‘Ribosome’, ‘Protein export’, ‘Nucleocytoplasmic
transport’ and ‘Metabolic pathways’. Cross-dataset comparison
prioritized BCKDHA as a candidate resistance-associated protein and
CRC TMA analysis showed that high BCKDHA expression was associated
with shorter OS. In parallel, BCKDHA overexpression and knockdown
experiments in SW480 cells provided preliminary functional evidence
associating BCKDHA with the response to radiotherapy and to a
number of DDR-associated proteins and signaling changes.
Prior to nCRT, the pathways preliminarily enriched
in resistant tumors in the present small discovery set were
compatible with known intrinsic chemo- and radiotherapy-resistance
programs, particularly those related to cell-matrix, cell-cell
adhesion, cytoskeleton and metabolic adaptation, including
‘Adherens junction’, ‘Focal adhesion’, ‘Tight junction’,
‘Regulation of actin cytoskeleton’, ‘Propanoate metabolism’ and
‘Amino sugar and nucleotide sugar metabolism’. Myosin regulatory
light polypeptide 9 and L1CAM, both more abundant in resistant
tumors before nCRT, have previously been associated with worse
treatment response or aggressive behavior in CRC and LARC (15,19,35,36).
Ribonucleotide reductase regulatory subunit M2, a rate-limiting
enzyme for deoxyribonucleotide triphosphates synthesis, has been
associated with chemotherapy resistance in CRC (37–39).
Glutaryl-CoA dehydrogenase, metastasis-associated colon cancer-1
and 3-phosphoinositide-dependent protein kinase 1 further support a
role for pro-tumorigenic signaling and metabolic adaptation in
resistant tumors (40–46). Although these observations are
descriptive, they suggest that resistant tumors may harbor
pre-existing adhesion-associated and metabolic programs before
treatment.
Post-nCRT, proteins more abundant in resistant
tumors were preliminarily enriched in pathways related to
replication, transcription, translation and metabolism, including
‘DNA replication’, ‘RNA polymerase’, ‘Ribosome’, ‘Protein export’,
‘Nucleocytoplasmic transport’ and ‘Metabolic pathways’. This
pattern is compatible with retention of biosynthesis-, replication-
and DDR-associated programs in treatment-selected tumors. The
increased abundance of MCM family members (MCM2/4/6/7) and RNA
polymerase subunits (POLR1E, POLR2E and POLR2I) suggested
persistence of replication- and transcription-associated activity
(43–45). Elevated expression of the ribosomal
protein RPL5 and the mitochondrial ribosomal proteins, MRPL1 and
MRPL24, further indicated sustained translational capacity
(46,47). Methionyl-tRNA synthetase 1, which
has been implicated in DNA damage repair through AIMP3 release and
PROM1, a stem cell-associated marker implicated in
chemoradiotherapy resistance, were also more abundant in resistant
tumors (48–55). In parallel, hexokinase 2 and
phosphoribosyl pyrophosphate amidotransferase suggest continued
metabolic remodeling, including glycolytic and nucleotide
biosynthetic support for treatment-selected cells (56–61).
Collectively, these post-treatment data should be interpreted as
pathway-level signals consistent with treatment selection rather
than direct mechanistic proof of specific resistance processes. In
addition, these post-nCRT pathway signals should be interpreted as
bulk tissue readouts of treatment-selected residual lesions rather
than as purely tumor-cell-intrinsic changes.
BCKDHA (α subunit), together with BCKDHB (β
subunit), form the E1 component of the BCKDH complex. BCKDH
consists of E1, E2 (encoded by dihydrolipoamide branched-chain
transacylase E2) and E3 (encoded by dihydrolipoamide dehydrogenase)
and is the rate-limiting enzyme in branched-chain amino acid (BCAA)
catabolism, catalyzing the conversion of BCAAs (leucine, isoleucine
and valine) derived intermediates into acetyl-CoA and succinyl-CoA
(34). Accumulating evidence has
indicated altered BCAA metabolism in CRC (62,63).
For example, serum levels of leucine and valine were lower in
patients with CRC compared with healthy controls (62). A dynamic metabolomic study of 106
patients with LARC further suggested that BCAA metabolism was
associated with response to nCRT, identifying α-ketoisovaleric
acid, a key intermediate of BCAA catabolism, as a predictive
biomarker and reporting a marked decrease in nCRT resistance in
LARC (63). In this context, the
consistent direction of higher abundance or expression of BCKDHA
across datasets supports prioritizing BCKDHA as a candidate
resistance-associated protein. In addition, CRC TMA analysis showed
that higher BCKDHA expression was associated with a shorter OS,
supporting broader clinical relevance in CRC. However, this TMA
cohort addressed prognosis in CRC rather than the pretreatment
prediction of nCRT response in LARC; therefore, the TMA findings
should be interpreted as supportive rather than definitive evidence
of an association between BCKDHA and nCRT response in LARC.
BCAA catabolism can provide tumor cells with energy
through the tricarboxylic acid cycle and can also supply carbon and
nitrogen for biosynthetic processes, as well as metabolites
potentially relevant to epigenomic regulation (64). BCKDHA has been associated with
malignant phenotypes in a number of types of cancer, including
pancreatic cancer, melanoma and lung cancer; it has been reported
to provide carbon for pancreatic cancer cell proliferation
(65) and a previous study showed
that it promoted melanoma cell malignancy by activating lipogenic
enzymes fatty acid synthase and ATP citrate lyase (66). Bo et al (67) further demonstrated that
X-irradiation induced BCKDHA dephosphorylation in pancreatic and
lung cancer cells and that BCKDHA knockdown increased mitotic
catastrophe, increased DNA damage and lowered cellular ATP levels.
In the present study, BCKDHA overexpression increased clonogenic
survival after irradiation, whereas BCKDHA knockdown enhanced
radiosensitivity and was accompanied by altered
ATM-Chk2-RAD51-associated signaling. Notably, the relatively weak
γ-H2AX signal after BCKDHA knockdown did not necessarily exclude
persistent DNA damage, as γ-H2AX reflects upstream DDR kinase
activity and H2AX availability in addition to double-stranded break
burden and the immunoblot readout was not directly equivalent to
the γ-H2AX foci assay. A possible explanation lies in
BCKDHA-dependent metabolic changes limiting ATP availability,
thereby weakening phosphorylation-dependent DDR signaling. The
lower ATP levels reported by Bo et al (67) were biologically relevant, as ATP not
only serves as a cellular energy currency but also as the phosphate
donor used by protein kinases. Therefore, reduced ATP availability
may contribute to the blunted H2AX phosphorylation observed after
BCKDHA knockdown, although the direct molecular association remains
to be determined.
A number of limitations of the present study should
be noted. First, the proteomic discovery set was small,
retrospective, sex-imbalanced and composed of retrospectively
screened cases with pelvic MRI available both before and after nCRT
and adequate available FFPE material, selected to capture clear
RECIST (version 1.1)-defined radiological response categories
rather than matched baseline characteristics; this may have
increased selection bias and limited reproducibility and
generalizability. As the discovery set was markedly sex-imbalanced,
potential sex-associated differences in proteomic response patterns
could not be evaluated. Second, no proteins remained significant
after multiple-testing correction and candidate proteins were
screened using nominal P-value and fold-change thresholds. Third,
response grouping was based on RECIST rather than pathological
regression. Fourth, because no macrodissection or laser-capture
microdissection was performed before protein extraction, the
post-nCRT FFPE profiles should be interpreted as bulk tissue
readouts that may include tumor, stromal, inflammatory and
treatment-associated fibrotic components rather than as purely
tumor-cell-intrinsic changes. This issue is particularly relevant
in post-nCRT specimens, in which treatment can induce fibrosis,
inflammation and immune-cell infiltration. Fifth, the CRC tissue
microarray addressed broader prognostic relevance in CRC rather
than pretreatment predictive value for nCRT response in LARC.
Finally, the functional validation was performed primarily in a
single cell line and the DDR-associated western blotting data
should be interpreted as pathway-level association rather than
direct mechanistic proof of DNA repair regulation. Despite these
limitations, the present study provides a useful starting point for
future research by prioritizing BCKDHA and a number of additional
candidate proteins for validation in larger pretreatment LARC
cohorts and more direct mechanistic studies.
Collectively, the results of the present study
identified BCKDHA as a candidate nCRT resistance-associated protein
in LARC, providing preliminary functional evidence linking BCKDHA
to the response to radiotherapy and DNA damage response-associated
signaling. These findings are hypothesis-generating and warrant
validation in larger pretreatment LARC cohorts and in more direct
mechanistic studies.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Medical Scientific
Research Foundation of Chongqing, China (grant no. 2023MSXM040),
the Chongqing Natural Science Foundation General Project (grant no.
CSTB2022NSCQ-MSX0773) and the Clinical Study Projects of the First
Affiliated Hospital of Army Medical University (grant no.
2024IITZDB11).
Availability of data and materials
The data generated in the present study may be found
in the ProteomeXchange consortium via the iProX partner repository
under accession number PXD069647 or at the following URL:
https://www.iprox.cn/page/subproject.html?id=IPX0013849001.
Authors' contributions
JYH, YDo and CC contributed to the methodology, data
acquisition, sample collection, experiments, figure preparation,
and writing the original draft. FHL, FWR, QD, YDu and TRL
contributed to the experiments and project administration. YYZ, TW
and YHC reviewed and verified the clinical data, reviewed and
interpreted the experimental data, and designed and performed the
X-ray irradiation procedures. XZ, SLX and JJL contributed to
conceptualization of the present study, resources, funding
acquisition, writing, reviewing and editing the manuscript, and
provided supervision. JYH and JJL confirm the authenticity of all
the raw data. All authors read and approved the final version of
the manuscript.
Ethical approval and consent to
participate
The present study protocol was reviewed and approved
by the Ethics Committee of the First Affiliated Hospital of Army
Medical University, PLA. [approval no. (A)KY2025105]. Written
informed consent was obtained from all participants or their legal
guardians, where applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Use of artificial intelligence tools
During the preparation of this work, artificial
intelligence tools were used to improve the readability and
language of the manuscript or to generate images, and subsequently,
the authors revised and edited the content produced by the
artificial intelligence tools as necessary, taking full
responsibility for the ultimate content of the present
manuscript.
References
|
1
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI
|
|
2
|
Han B, Zheng R, Zeng H, Wang S, Sun K,
Chen R, Li L, Wei W and He J: Cancer incidence and mortality in
China, 2022. J Natl Cancer Cent. 4:47–53. 2024.PubMed/NCBI
|
|
3
|
Appelt AL, Pløen J, Harling H, Jensen FS,
Jensen LH, Jørgensen JC, Lindebjerg J, Rafaelsen SR and Jakobsen A:
High-dose chemoradiotherapy and watchful waiting for distal rectal
cancer: A prospective observational study. Lancet Oncol.
16:919–927. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scott AJ, Kennedy EB, Berlin J, Brown G,
Chalabi M, Cho MT, Cusnir M, Dorth J, George M, Kachnic LA, et al:
Management of locally advanced rectal cancer: ASCO guideline. J
Clin Oncol. 42:3355–3375. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fernandez LM, São Julião GP, Figueiredo
NL, Beets GL, van der Valk MJM, Bahadoer RR, Hilling DE,
Meershoek-Klein Kranenbarg E, Roodvoets AGH, Renehan AG, et al:
Conditional recurrence-free survival of clinical complete
responders managed by watch and wait after neoadjuvant
chemoradiotherapy for rectal cancer in the International Watch
& Wait Database: A retrospective, international, multicentre
registry study. Lancet Oncol. 22:43–50. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jin J, Tang Y, Hu C, Jiang LM, Jiang J, Li
N, Liu WY, Chen SL, Li S, Lu NN, et al: Multicenter, randomized,
phase III Trial of Short-term radiotherapy plus chemotherapy versus
long-term chemoradiotherapy in locally advanced rectal cancer
(STELLAR). J Clin Oncol. 40:1681–1692. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu J, Liu A, Sun X, Liu L, Zhu Y, Zhang
T, Jia J, Tan S, Wu J, Wang X, et al: Multicenter, randomized,
Phase III trial of neoadjuvant chemoradiation with capecitabine and
irinotecan guided by UGT1A1 status in patients with locally
advanced rectal cancer. J Clin Oncol. 38:4231–4239. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chang C, Bliggenstorfer JT, Liu J, Shearer
J, Dreher P, Bingmer K, Stein SL and Steinhagen E: Not all patients
with locally advanced rectal cancer benefit from neoadjuvant
therapy. Am Surg. 89:4327–4333. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Couwenberg AM, Burbach JPM, Berbee M,
Lacle MM, Arensman R, Raicu MG, Wessels FJ, Verdult J, Roodhart J,
Reerink O, et al: Efficacy of Dose-escalated chemoradiation on
complete tumor response in patients with locally advanced rectal
cancer (RECTAL-BOOST): A Phase 2 randomized controlled trial. Int J
Radiat Oncol Biol Phys. 108:1008–1018. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Peng J, Lv J and Peng J: KRAS mutation is
predictive for poor prognosis in rectal cancer patients with
neoadjuvant chemoradiotherapy: A systemic review and meta-analysis.
Int J Colorectal Dis. 36:1781–1790. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kamran SC, Lennerz JK, Margolis CA, Liu D,
Reardon B, Wankowicz SA, Van Seventer EE, Tracy A, Wo JY, Carter
SL, et al: Integrative molecular characterization of resistance to
neoadjuvant chemoradiation in rectal cancer. Clin Cancer Res.
25:5561–5571. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sanz-Garcia E, Argiles G, Elez E and
Tabernero J: BRAF mutant colorectal cancer: Prognosis, treatment,
and new perspectives. Ann Oncol. 28:2648–2457. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Scott JG, Berglund A, Schell MJ, Mihaylov
I, Fulp WJ, Yue B, Welsh E, Caudell JJ, Ahmed K, Strom TS, et al: A
genome-based model for adjusting radiotherapy dose (GARD): A
retrospective, cohort-based study. Lancet Oncol. 18:202–211. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xia H, Li Z, Lin Y, Lin Y, Zeng L, Xu B,
Yao Q and Zheng R: Validation of a genome-based model for adjusting
radiotherapy dose (GARD) in patients with locally advanced rectal
cancer. Sci Rep. 14:215722024. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chatila WK, Kim JK, Walch H, Marco MR,
Chen CT, Wu F, Omer DM, Khalil DN, Ganesh K, Qu X, et al: Genomic
and transcriptomic determinants of response to neoadjuvant therapy
in rectal cancer. Nat Med. 28:1646–1655. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Crotti S, Enzo MV, Bedin C, Pucciarelli S,
Maretto I, Del Bianco P, Traldi P, Tasciotti E, Ferrari M, Rizzolio
F, et al: Clinical predictive circulating peptides in rectal cancer
patients treated with neoadjuvant chemoradiotherapy. J Cell
Physiol. 230:1822–1828. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang H, Ji D, Tian H, Gao Z, Song C, Jia
J, Cui X, Zhong L, Shen J and Gu J: Predictive value of proteomic
markers for advanced rectal cancer with neoadjuvant
chemoradiotherapy. BMC Cancer. 22:8682022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mammano E, Galdi F, Pierobon M, Tessari E,
Deng J, Pucciarelli S, Agostini M, De Marchi F, Canzonieri V, De
Paoli A, et al: Multiplexed protein signal pathway mapping
identifies patients with rectal cancer that responds to neoadjuvant
treatment. Clin Colorectal Cancer. 11:268–274. 2012.PubMed/NCBI
|
|
19
|
Repetto O, De Re V, De Paoli A, Belluco C,
Alessandrini L, Canzonieri V and Cannizzaro R: Identification of
protein clusters predictive of tumor response in rectal cancer
patients receiving neoadjuvant chemo-radiotherapy. Oncotarget.
8:28328–28341. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chauvin A, Wang CS, Geha S, Garde-Granger
P, Mathieu AA, Lacasse V and Boisvert FM: The response to
neoadjuvant chemoradiotherapy with 5-fluorouracil in locally
advanced rectal cancer patients: A predictive proteomic signature.
Clin Proteomics. 15:162018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Croner RS, Sevim M, Metodiev MV, Jo P,
Ghadimi M, Schellerer V, Brunner M, Geppert C, Rau T, Stürzl M, et
al: Identification of predictive markers for response to
neoadjuvant chemoradiation in rectal carcinomas by proteomic
isotope coded protein label (ICPL) analysis. Int J Mol Sci.
17:2092016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stanojevic A, Samiotaki M, Lygirou V,
Marinkovic M, Nikolic V, Stojanovic-Rundic S, Jankovic R, Vlahou A,
Panayotou G, Fijneman RJA, et al: Data-Independent acquisition mass
spectrometry analysis of FFPE rectal cancer samples offers In-Depth
proteomics characterization of the response to neoadjuvant
chemoradiotherapy. Int J Mol Sci. 24:154122023. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bowden DL, Sutton PA, Wall MA, Jithesh PV,
Jenkins RE, Palmer DH, Goldring CE, Parsons JL, Park BK,
Kitteringham NR and Vimalachandran D: Proteomic profiling of rectal
cancer reveals acid ceramidase is implicated in radiation response.
J Proteomics. 179:53–60. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Babic T, Lygirou V, Rosic J, Miladinov M,
Rom AD, Baira E, Stroggilos R, Pappa E, Zoidakis J, Krivokapic Z
and Nikolic A: Pilot proteomic study of locally advanced rectal
cancer before and after neoadjuvant chemoradiotherapy indicates
high metabolic activity in non-responders' tumor tissue. Proteomics
Clin Appl. 17:e21001162023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zott T, Wolf M, Plessl-Walder G, Regele H,
Bergmann M, Meier-Menches SM, Gerner C, Silberhumer GR and Bileck
A: Proteomic analysis of FFPE tissue samples identifies potential
molecular mechanisms mediating resistance to radiotherapy in rectal
cancer. J Proteome Res. 24:3990–4001. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Weiser MR: AJCC 8th Edition: Colorectal
Cancer. Ann Surg Oncol. 25:1454–1455. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
R Core Team, . R: A language and
environment for statistical computing. Vienna, Austria: R
Foundation for Statistical Computing; 2012
|
|
28
|
Thévenot EA, Roux A, Xu Y, Ezan E and
Junot C: Analysis of the human adult urinary metabolome variations
with age, body mass index and gender by implementing a
comprehensive workflow for univariate and OPLS statistical
analyses. J Proteome Res. 14:3322–3335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wickham H: ggplot2: Elegant Graphics for
Data Analysis. Springer-Verlag New York; New York, NY: 2016
|
|
30
|
Kolde R: pheatmap: Pretty Heatmaps. R
package version 1.0.12. 2019.Available from:. https://CRAN.R-project.org/package=pheatmap
|
|
31
|
Goldberg T, Hecht M, Hamp T, Karl T,
Yachdav G, Ahmed N, Altermann U, Angerer P, Ansorge S, Balasz K, et
al: LocTree3 prediction of localization. Nucleic Acids Res.
42:W350–W355. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kanehisa M: Post-genome Informatics.
Oxford University Press; 2000, View Article : Google Scholar
|
|
33
|
Wang HB, Zhang H, Zhang JP, Li Y, Zhao B,
Feng GK, Liu W, Li MZ, Huang WL, Tsao SW, et al: Neuropilin 1 is an
entry factor that promotes EBV infection of nasopharyngeal
epithelial cells. Nat Commun. 6:62402015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Peng H, Wang Y and Luo W: Multifaceted
role of branched-chain amino acid metabolism in cancer. Oncogene.
39:6747–6756. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Deng S, Cheng D, Wang J, Gu J, Xue Y,
Jiang Z, Qin L, Mao F, Cao Y and Cai K: MYL9 expressed in
cancer-associated fibroblasts regulate the immune microenvironment
of colorectal cancer and promotes tumor progression in an autocrine
manner. J Exp Clin Cancer Res. 42:2942023. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ganesh K, Basnet H, Kaygusuz Y, Laughney
AM, He L, Sharma R, O'Rourke KP, Reuter VP, Huang YH, Turkekul M,
et al: L1CAM defines the regenerative origin of
metastasis-initiating cells in colorectal cancer. Nat Cancer.
1:28–45. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhan Y, Jiang L, Jin X, Ying S, Wu Z, Wang
L, Lou Y and Qiu Y: Inhibiting RRM2 to enhance the anticancer
activity of chemotherapy. Biomed Pharmacother. 133:1109962021.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yazdanian Z, Mobarra N, Fazel A, Fazeli
MS, Ghasemi S, Danesteh S, Khoshrou A, Pakzad R, Raji S, Rafiee M
and Akbar S: Ribonucleotide-diphosphate reductase subunit M2 (RRM2)
expression and colorectal cancer invasiveness: A potential
prognostic biomarker. Mol Biol Rep. 52:4472025. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lin ZP, Belcourt MF, Cory JG and
Sartorelli AC: Stable suppression of the R2 subunit of
ribonucleotide reductase by R2-targeted short interference RNA
sensitizes p53(−/-) HCT-116 colon cancer cells to DNA-damaging
agents and ribonucleotide reductase inhibitors. J Biol Chem.
279:27030–27038. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Verma S, Crawford D, Khateb A, Feng Y,
Sergienko E, Pathria G, Ma CT, Olson SH, Scott D, Murad R, et al:
NRF2 mediates melanoma addiction to GCDH by modulating apoptotic
signalling. Nature Cell Biology. 24:1422–1432. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Radhakrishnan H, Walther W, Zincke F,
Kobelt D, Imbastari F, Erdem M, Kortüm B, Dahlmann M and Stein U:
MACC1-the first decade of a key metastasis molecule from gene
discovery to clinical translation. Cancer Metastasis Rev.
37:805–820. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Han D, Wang W, Jeon JH, Shen T, Huang X,
Yi P, Dong B and Yang F: Cooperative activation of PDK1 and AKT by
MAPK4 enhances cancer growth and resistance to therapy. PLoS Biol.
21:e30022272023. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tian Y, Zhou Y, Chen F, Qian S, Hu X,
Zhang B and Liu Q: Research progress in MCM family: Focus on the
tumor treatment resistance. Biomed Pharmacother. 173:1164082024.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Malysa A, Zhang XM and Bepler G:
Minichromosome maintenance proteins: From DNA replication to the
DNA damage response. Cells. 14:122024. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Muste Sadurni M and Saponaro M:
Deregulations of RNA Pol II subunits in cancer. Appl Biosci.
2:459–476. 2023. View Article : Google Scholar
|
|
46
|
El Khoury W and Nasr Z: Deregulation of
ribosomal proteins in human cancers. Biosci Rep.
41:BSR202115772021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lin X, Guo L, Lin X, Wang Y and Zhang G:
Expression and prognosis analysis of mitochondrial ribosomal
protein family in breast cancer. Sci Rep. 12:106582022. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Khan K, Gogonea V and Fox PL:
Aminoacyl-tRNA synthetases of the multi-tRNA synthetase complex and
their role in tumorigenesis. Transl Oncol. 19:1013922022.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Jin Q, Liu G, Wang B, Li S, Ni K, Wang C,
Ren J, Zhang S and Dai Y: High methionyl-tRNA synthetase expression
predicts poor prognosis in patients with breast cancer. J Clin
Pathol. 73:803–812. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jang SI, Nahm JH, Lee SY, Cho JH, Do MY,
Park JS, Lee HS, Yang J, Kong J, Jung S, et al: Prediction of
prognosis in pancreatic cancer according to Methionyl-tRNA
synthetase 1 expression as determined by immunohistochemical
staining. Cancers (Basel). 15:54132023. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kwon NH, Kang T, Lee JY, Kim HH, Kim HR,
Hong J, Oh YS, Han JM, Ku MJ, Lee SY and Kim S: Dual role of
methionyl-tRNA synthetase in the regulation of translation and
tumor suppressor activity of aminoacyl-tRNA synthetase-interacting
multifunctional protein-3. Proc Natl Acad Sci USA. 108:19635–19640.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ren F, Sheng WQ and Du X: CD133: A cancer
stem cells marker, is used in colorectal cancers. World J
Gastroenterol. 19:2603–2611. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhang X, Yang L, Lei W, Hou Q, Huang M,
Zhou R, Enver T and Wu S: Single-cell sequencing reveals
CD133+CD44-originating evolution and novel stemness related
variants in human colorectal cancer. EBioMedicine. 82:1041252022.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yuan Z, Liang X, Zhan Y, Wang Z, Xu J, Qiu
Y, Wang J, Cao Y, Le VM, Ly HT, et al: Targeting CD133 reverses
drug-resistance via the AKT/NF-κB/MDR1 pathway in colorectal
cancer. Br J Cancer. 122:1342–1353. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Akbari M, Shomali N, Faraji A, Shanehbandi
D, Asadi M, Mokhtarzadeh A, Shabani A and Baradaran B: CD133: An
emerging prognostic factor and therapeutic target in colorectal
cancer. Cell Biol Int. 44:368–380. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liao M, Yao D, Wu L, Luo C, Wang Z, Zhang
J and Liu B: Targeting the Warburg effect: A revisited perspective
from molecular mechanisms to traditional and innovative therapeutic
strategies in cancer. Acta Pharm Sin B. 14:953–1008. 2024.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Huang X, Liu M, Sun H, Wang F, Xie X, Chen
X, Su J, He Y, Dai Y, Wu H and Shen L: HK2 is a radiation resistant
and independent negative prognostic factor for patients with
locally advanced cervical squamous cell carcinoma. Int J Clin Exp
Pathol. 8:4054–4063. 2015.PubMed/NCBI
|
|
58
|
Barba I, Carrillo-Bosch L and Seoane J:
Targeting the warburg effect in cancer: Where Do We Stand? Int J
Mol Sci. 25:31422024. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Pareek V, Pedley AM and Benkovic SJ: Human
de novo purine biosynthesis. Crit Rev Biochem Mol Biol. 56:1–16.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kodama M, Oshikawa K, Shimizu H, Yoshioka
S, Takahashi M, Izumi Y, Bamba T, Tateishi C, Tomonaga T, Matsumoto
M and Nakayama KI: A shift in glutamine nitrogen metabolism
contributes to the malignant progression of cancer. Nat Commun.
11:13202020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhou W and Wahl DR: Purine metabolism
promotes radioresistance and is a therapeutic target in
glioblastoma. Mol Cell Oncol. 7:18349022020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Qiu Y, Cai G, Su M, Chen T, Zheng X, Xu Y,
Ni Y, Zhao A, Xu LX, Cai S and Jia W: Serum metabolite profiling of
human colorectal cancer using GC-TOFMS and UPLC-QTOFMS. J Proteome
Res. 8:4844–4850. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lv J, Jia H, Mo M, Yuan J, Wu Z, Zhang S,
Zhe F, Gu B, Fan B, Li C, et al: Changes of serum metabolites
levels during neoadjuvant chemoradiation and prediction of the
pathological response in locally advanced rectal cancer.
Metabolomics. 18:992022. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Sivanand S and Vander Heiden MG: Emerging
roles for Branched-chain amino acid metabolism in cancer. Cancer
Cell. 37:147–156. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Lee JH, Cho YR, Kim JH, Kim J, Nam HY, Kim
SW and Son J: Branched-chain amino acids sustain pancreatic cancer
growth by regulating lipid metabolism. Exp Mol Med. 51:1–11. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Tian Y, Ma J, Wang M, Yi X, Guo S, Wang H,
Zhang H, Wang H, Yang Y, Zhang B, et al: BCKDHA contributes to
melanoma progression by promoting the expressions of lipogenic
enzymes FASN and ACLY. Exp Dermatol. 32:1633–1643. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Bo T, Osaki T and Fujii J:
Dephosphorylation of branched-chain α-keto acid dehydrogenase E1α
(BCKDHA) promotes branched-chain amino acid catabolism and renders
cancer cells resistant to X-rays by mitigating DNA damage. Biochem
Biophys Res Commun. 742:1511542025. View Article : Google Scholar : PubMed/NCBI
|