Introduction
Although uterine leiomyosarcoma (Ut-LMS) is rare, it
is a highly aggressive malignancy arising from the uterine smooth
muscle and accounts for less than 1% of all uterine tumors
(1,2). Despite its low incidence, Ut-LMS is
associated with poor clinical outcomes due to rapid tumor
expansion, early hematogenous dissemination, and a high rate of
postoperative recurrence (3).
Current therapeutic options are limited, with hysterectomy serving
as the primary intervention (1).
Conventional chemotherapeutic regimens, including doxorubicin,
gemcitabine, and docetaxel, offer only modest benefits, and the
overall survival of patients with advanced disease has demonstrated
little improvement over the last decade (4,5). These
challenges highlight the urgent need for new molecular-targeted
therapies for Ut-LMS.
Recent genomic and transcriptomic studies have
indicated that Ut-LMS harbors substantial molecular heterogeneity
and frequently exhibits alterations in TP53, RB1, ATRX, and PTEN
with widespread chromosomal instability (6–8). These
molecular abnormalities suggest that Ut-LMS cells may rely on
stress-adaptive mechanisms that support DNA damage responses,
cell-cycle progression, transcriptional regulation, and protein
homeostasis. Therefore, targeting regulatory pathways that sustain
these adaptive survival mechanisms may provide a rational
therapeutic strategy for Ut-LMS.
TAK-981 (subasumstat) is a first-in-class inhibitor
of the small ubiquitin-like modifier (SUMO) activation pathway,
which functions by covalently inhibiting SUMO-activating enzyme
(SAE) complex composed of SAE1 and SAE2 (9,10).
SUMOylation is a crucial post-translational modification involved
in cell cycle regulation, DNA repair, transcription, and cellular
stress responses, and its aberrant regulation can promote tumor
progression and therapeutic resistance by supporting oncogenic
signaling, genome stability, and stress-adaptive survival
mechanisms in cancer cells (11–13).
These functions are particularly relevant to Ut-LMS, which is
characterized by frequent alterations in tumor suppressor pathways,
chromosomal instability, aggressive proliferation, and therapeutic
resistance. Thus, we hypothesized that Ut-LMS cells may be
vulnerable to SAE inhibition because blockade of the SUMO pathway
could disrupt stress-adaptive mechanisms required for tumor cell
survival. Preclinical studies have demonstrated that TAK-981 exerts
potent antitumor effects in several malignancies by inducing
interferon signaling, apoptosis, and cell cycle arrest (14–16).
In addition, our previous study showed that TAK-981 suppressed cell
viability and modulated apoptosis, cell-cycle arrest, and autophagy
in ELT3 uterine leiomyoma cells (17). Although uterine leiomyoma and Ut-LMS
both originate from uterine smooth muscle, they represent
biologically distinct benign and malignant tumors, respectively.
Therefore, it remains unclear whether the TAK-981-induced cellular
responses observed in benign uterine leiomyoma cells can be
extended to malignant Ut-LMS cells.
The effects of TAK-981 on cell growth in Ut-LMS
cells were investigated and TAK-981 was found to substantially
suppress SK-UT-1B cell growth. TAK-981 induced
G0/G1 cell cycle arrest, triggered reactive
oxygen species (ROS)-dependent mitochondrial dysfunction and
apoptosis, and inhibited SUMO2/3 conjugation while activating the
tumor suppressors, p21 and p53. These findings suggest that
targeting the SUMOylation pathway is a novel and promising
therapeutic strategy for Ut-LMS.
Materials and methods
Cell culture and reagents
Human Ut-LMS cell lines (SK-UT-1 and SK-UT-1B; ATCC,
Manassas, VA, USA) were cultured in the Minimal Essential Medium
(LM007-07; Welgene, Gyeongsan, Republic of Korea) supplemented with
10% fetal bovine serum (FBS; SH30919.03; Hyclone, Logan, UT, USA)
and 1% penicillin-streptomycin (15140122; Gibco, Waltham, MA, USA)
at 37°C in a humidified atmosphere containing 5% CO2.
TAK-981 (HY-111789; MedChemExpress, Monmouth Junction, NJ, USA) was
dissolved in dimethyl sulfoxide (DMSO; DMSO100; LPS Solution,
Daejeon, Republic of Korea) and used as vehicle control.
N-Acetyl-L-cysteine (NAC; A7250) and catalase (CAT; C1345) were
purchased from Sigma-Aldrich (St. Louis, MO, USA).
MTT and lactate dehydrogenase (LDH)
assays
Cell viability and cytotoxicity were determined
using MTT and LDH assays. Cells were seeded into 96-well plates at
a density of 5×103 cells per well and allowed to attach
for 24 h before TAK-981 treatment. After exposure to TAK-981 for 24
or 48 h, cell viability was assessed using MTT solution (5 mg/ml;
M2128, Sigma-Aldrich, St. Louis, MO, USA). Following a 2 h
incubation at 37°C, the resulting formazan crystals were dissolved
in 150 µl of DMSO, and absorbance was measured at 570 nm using a
microplate reader (INNO; L-Tek, Seongnam, Republic of Korea).
Cytotoxicity was evaluated using the LDH Plus Cytotoxicity Assay
Kit (GBL-P500; Dyne Bio, Seongnam, Republic of Korea) according to
the manufacturer's instructions, and absorbance was measured at 490
nm.
Flow cytometric analysis of
apoptosis
Apoptotic cell death was analyzed using a Muse
Annexin V & Dead Cell Kit (MCH100105; Cytek Biosciences,
Fremont, CA, USA) according to the manufacturer's guidelines. Cells
were seeded in six-well plates at a density of 5×104 cells/well and
allowed to adhere for 24 h before treatment with different
concentrations of TAK-981 for 48 h. After incubation, both floating
and adherent cells were collected by trypsinization, centrifuged,
and resuspended in 100 µl of assay medium. Subsequently, 100 µl of
Annexin V & Dead Cell Reagent containing 7-AAD was added, and
the mixture was incubated for 20 min at room temperature in the
dark. Stained samples were immediately analyzed using a Guava Muse
Cell Analyzer (0500-3115; Luminex, Austin, TX, USA).
Flow cytometric analysis of cell cycle
progression
The distribution of cells across different cell
cycle phases was determined using the Muse Cell Cycle Kit
(MCH100106; Cytek Biosciences, Fremont, CA, USA). Briefly, the
cells were harvested, washed with PBS, and fixed with 70% cold
ethanol at −20°C for approximately 3 h. After fixation, the samples
were centrifuged, resuspended in 200 µl of Muse Cell Cycle Reagent,
and incubated for 30 min at room temperature in the dark. DNA
content was analyzed using a Guava Muse Cell Analyzer to determine
the percentage of cells in the G0/G1, S and
G2/M phases.
Flow cytometric analysis of
Ki67-positive cells
Cell proliferation was assessed using the Muse Ki67
Proliferation Kit (MCH100114; Cytek Biosciences, Fremont, CA, USA).
After treatment, cells were collected and processed for fixation
and permeabilization according to the manufacturer's protocol. The
cells were then stained with either an anti-Ki67 antibody or the
corresponding isotype control antibody. Ki67-positive populations
were quantified using a Guava Muse Cell Analyzer.
Flow cytometric analysis of
intracellular ROS
Intracellular ROS levels were quantified using the
Muse Oxidative Stress Kit (MCH100111; Cytek Biosciences) following
the manufacturer's protocol. Cells were seeded in six-well plates
at a density of 5×104 cells per well and allowed to attach for 24 h
before exposure to TAK-981 at concentrations of 0, 0.5, 1, or 2 µM
for 48 h. After incubation, the cells were collected, washed, and
resuspended in 1X assay buffer. Afterward, cell suspensions were
stained with the Muse Oxidative Stress Reagent and incubated at
37°C for 30 min in the dark. ROS generation was analyzed using a
Guava Muse Cell Analyzer.
Measurement of mitochondrial membrane
potential (ΔΨm)
ΔΨm was evaluated using the Muse MitoPotential Kit
(MCH100110; Cytek Biosciences) according to the manufacturer's
instructions. After 48 h of TAK-981 treatment, the cells were
harvested, washed with PBS, and resuspended in 1X assay buffer at a
concentration of 1–2×106 cells/ml. Next, the cells were stained
with MitoPotential dye working solution for 20–30 min at room
temperature in the dark, followed by staining with 7-AAD for an
additional 5 min. The samples were analyzed using a Guava Muse Cell
Analyzer, and the percentages of live/depolarized, total
depolarized, and live/polarized cells were quantified.
Flow cytometric analysis of
autophagy
Autophagic activity was analyzed using the Muse
Autophagy LC3 Antibody-Based Kit (MCH200109; Cytek Biosciences) in
accordance with the manufacturer's instructions. Briefly, both
control and TAK-981-treated cells were collected, washed, and
incubated with anti-LC3 Alexa Fluor 555 antibody and 1X autophagy
reagent for 30 min in the dark. After staining, the cells were
resuspended in 200 µl of 1X assay buffer and subjected to flow
cytometric analysis using a Guava Muse Cell Analyzer. The degree of
autophagy induction was determined by comparing the fluorescence
signal intensities of treated and untreated samples.
Protein extraction and western
blotting
Total cellular proteins were isolated using
radio-immunoprecipitation assay lysis buffer (R2002; Biosesang,
Gyeonggi-do, Republic of Korea) supplemented with a protease
inhibitor cocktail (04693132001; Roche, Basel, Switzerland).
Protein samples were separated using 15% sodium dodecyl
sulphate-polyacrylamide gel electrophoresis and transferred onto
polyvinylidene fluoride membranes (IPVH00010; Merck Millipore,
Billerica, MA, USA). The membranes were blocked with 5% skim milk
(262100; BD Difco, Franklin Lakes, NJ, USA) for 1 h at room
temperature and incubated overnight at 4°C with primary antibodies.
The following primary antibodies were used: SUMO2/3 (4971T), SAE2
(8688T), poly (ADP-ribose) polymerase (PARP; 9542S), caspase-3
(9665S), cleaved caspase-3 (9664S), p53 (2527S), p27 (3686T), p21
(2947S), p62/SQSTM1 (5114T), LC3B (2775S), and β-actin (4967S) (all
from Cell Signaling Technology, Danvers, MA, USA), as well as SAE1
(sc-398080) from Santa Cruz Biotechnology (Dallas, TX, USA). After
washing, the membranes were incubated with horseradish peroxidase
(HRP)-conjugated secondary antibodies (7074S or 7076S; Cell
Signaling Technology) for 1 h. Immunoreactive bands were visualized
using the Western Pico ECL Kit (PICO-250; LPS Solution, Daejeon,
Republic of Korea) and detected using an Azure c280 imaging system
(Azure Biosystems; Dublin, CA, USA).
Statistical analysis
Statistical analyses were conducted using GraphPad
Prism software (version 8.0; GraphPad Software, La Jolla, CA, USA).
All data are expressed as the mean ± standard deviation obtained
from at least three independent experiments. Group comparisons were
conducted using one-way analysis of variance, followed by Tukey's
multiple comparisons test or Dunnett's post hoc test, when
appropriate. P<0.05 was considered to indicate a statistically
significant difference.
Results
TAK-981 exhibits stronger cytotoxic
effects in SK-UT-1B than in SK-UT-1 cells
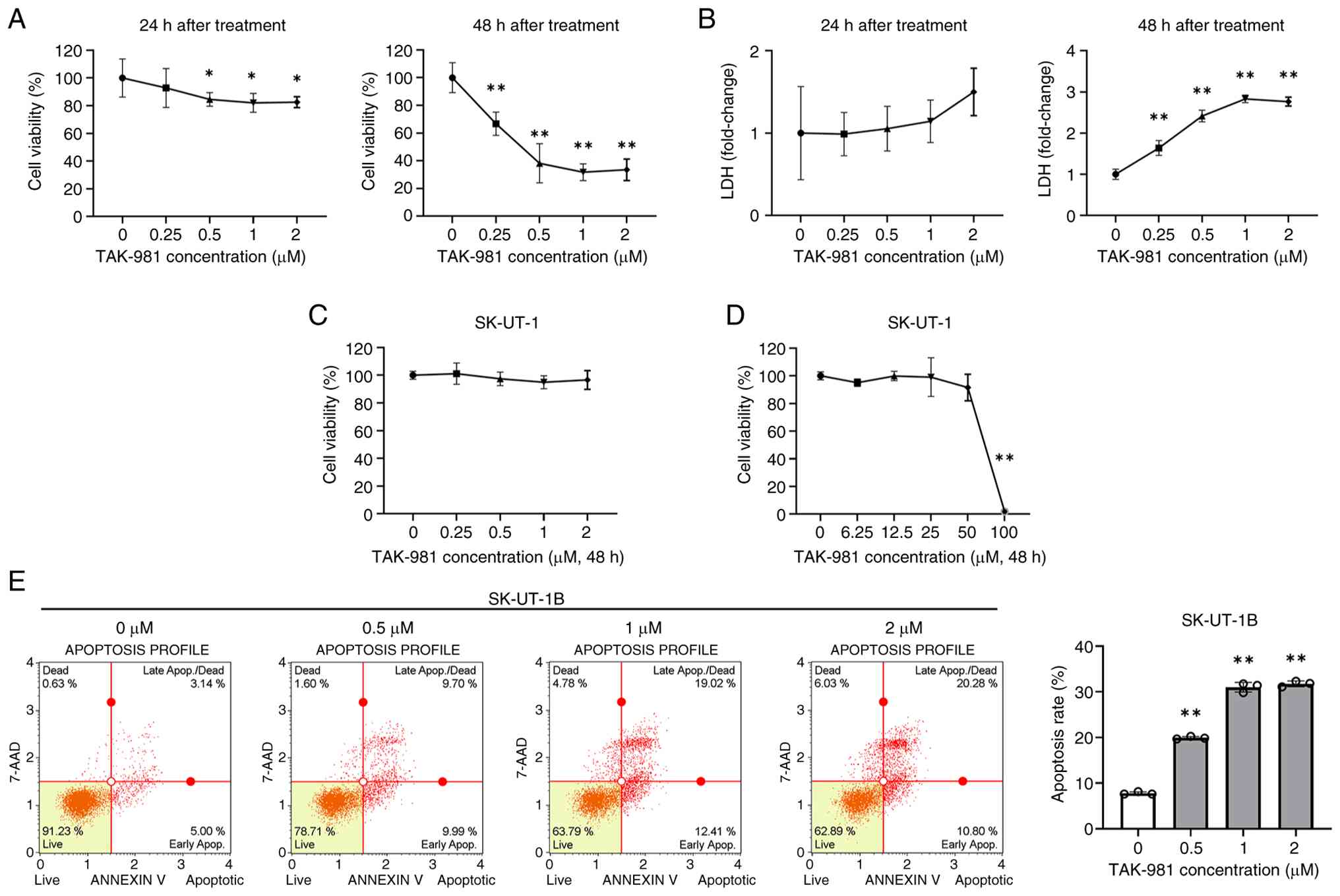
The cytotoxic effect of TAK-981 on Ut-LMS cells was
evaluated, in which SK-UT-1B cells were treated with increasing
concentrations of TAK-981 (0, 0.25, 0.5, 1, and 2 µM) for 24 and 48
h, and cell viability was measured using the MTT assay. After 24 h
of treatment, TAK-981 displayed minimal cytotoxicity, with 2 µM
treatment maintaining 82.5% cell viability compared to the control.
In contrast, prolonged exposure for 48 h markedly reduced cell
viability, showing 66.7, 38.1, 31.7, and 33.5% viability at 0.25,
0.5, 1, and 2 µM, respectively (Fig.
1A). Consistent with these findings, LDH release, an indicator
of membrane integrity loss, demonstrated no marked change at 24 h
but increased substantially after 48 h, with an approximately
2.8-fold elevation at 1 and 2 µM compared to the control (Fig. 1B).
Other Ut-LMS cell lines (SK-UT-1) exhibited no
marked changes in viability after 48 h of TAK-981 exposure at
similar concentrations (Fig. 1C).
SK-UT-1 cells exhibited <5% viability only at extremely high
concentrations (up to 100 µM), indicating nonspecific cytotoxicity
at supra-physiological levels (Fig.
1D). Collectively, these results demonstrate that TAK-981
exerted markedly greater cytotoxicity in SK-UT-1B cells than in
SK-UT-1 cells, particularly at low micromolar concentrations and
with prolonged exposure. Because normal uterine smooth muscle cells
have been reported to show reduced viability only at TAK-981
concentrations ≥50 µM (17),
SK-UT-1B cells were selected as the primary model for subsequent
mechanistic analyses.
TAK-981 induces apoptosis in SK-UT-1B
cells
Whether the cytotoxic effect of TAK-981 was
associated with apoptosis was determined using Annexin V staining
followed by flow cytometric analysis in SK-UT-1B cells after 48 h
of treatment. The proportion of apoptotic cells progressively
increased with the concentration of TAK-981, from 7.7% in control
cells to 19.9, 31.0, and 31.7% in cells treated with 0.5, 1, and 2
µM, respectively (Fig. 1E). TAK-981
induced apoptosis in SK-UT-1B cells and this apoptotic effect
became more prominent at higher drug concentrations.
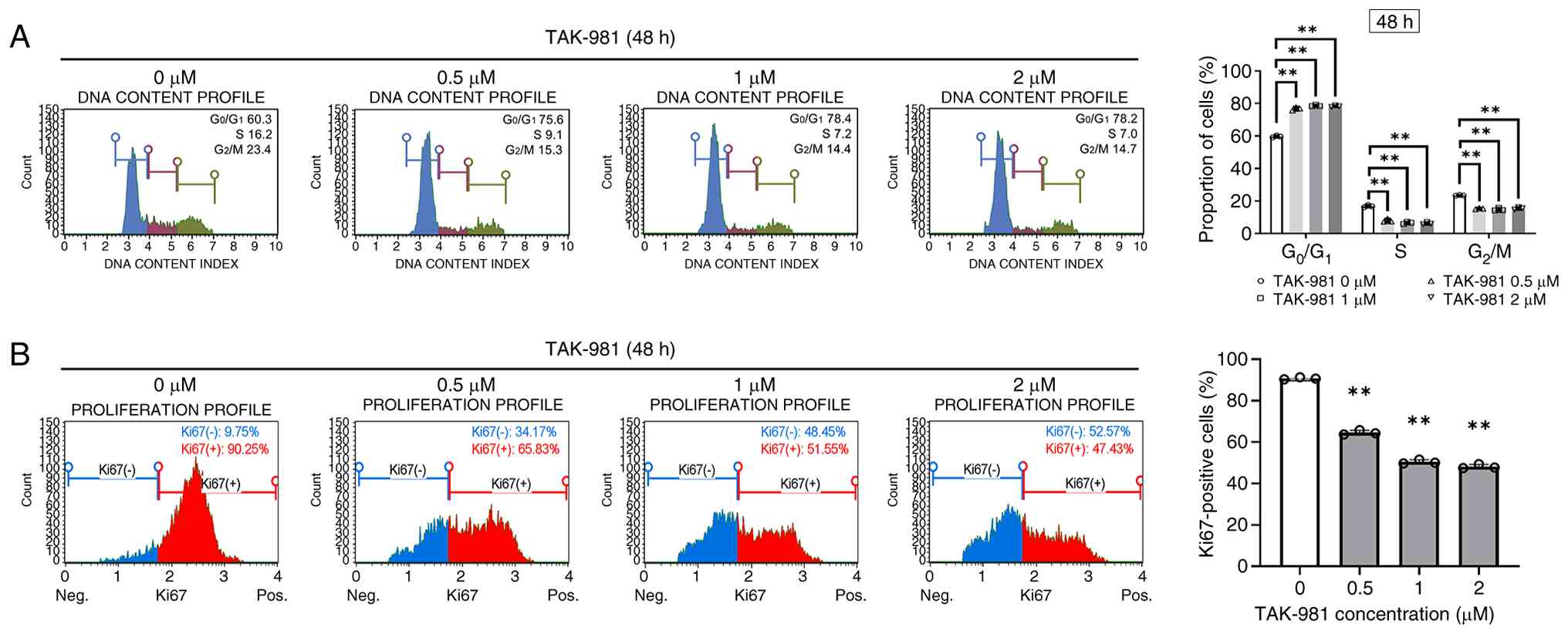
TAK-981 induces G0/G1-phase cell-cycle
arrest and suppresses proliferation in SK-UT-1B cells
The anti-proliferative effect of TAK-981 were
investigated by analyzing the cell cycle distribution and
proliferation markers in SK-UT-1B cells after 48 h of treatment.
Flow cytometry analysis indicated that TAK-981 caused a marked
accumulation of cells in the G0/G1 phase
(Fig. 2A). The proportion of
G0/G1-phase cells increased from 59.7% in
control cells to 76.7, 78.6, and 78.3% following exposure to 0.5,
1, and 2 µM TAK-981, respectively, whereas the proportion of
G2/M-phase cells decreased from 23.4% in control cells
to approximately 15% after treatment.
Cell proliferation was evaluated by assessing Ki67
expression using flow cytometry. The proportion of Ki67-positive
SK-UT-1B cells decreased markedly following TAK-981 exposure, from
90.7% in control cells to 64.5, 50.4, and 48.1% after treatment
with 0.5, 1, and 2 µM, respectively (Fig. 2B). Thus, TAK-981 suppressed DNA
synthesis and the proliferation of SK-UT-1B cells by inducing
G0/G1-phase cell-cycle arrest. Together with
the apoptosis data, these findings suggest that TAK-981 inhibited
SK-UT-1B cell growth through coordinated suppression of
proliferation and induction of apoptotic cell death.
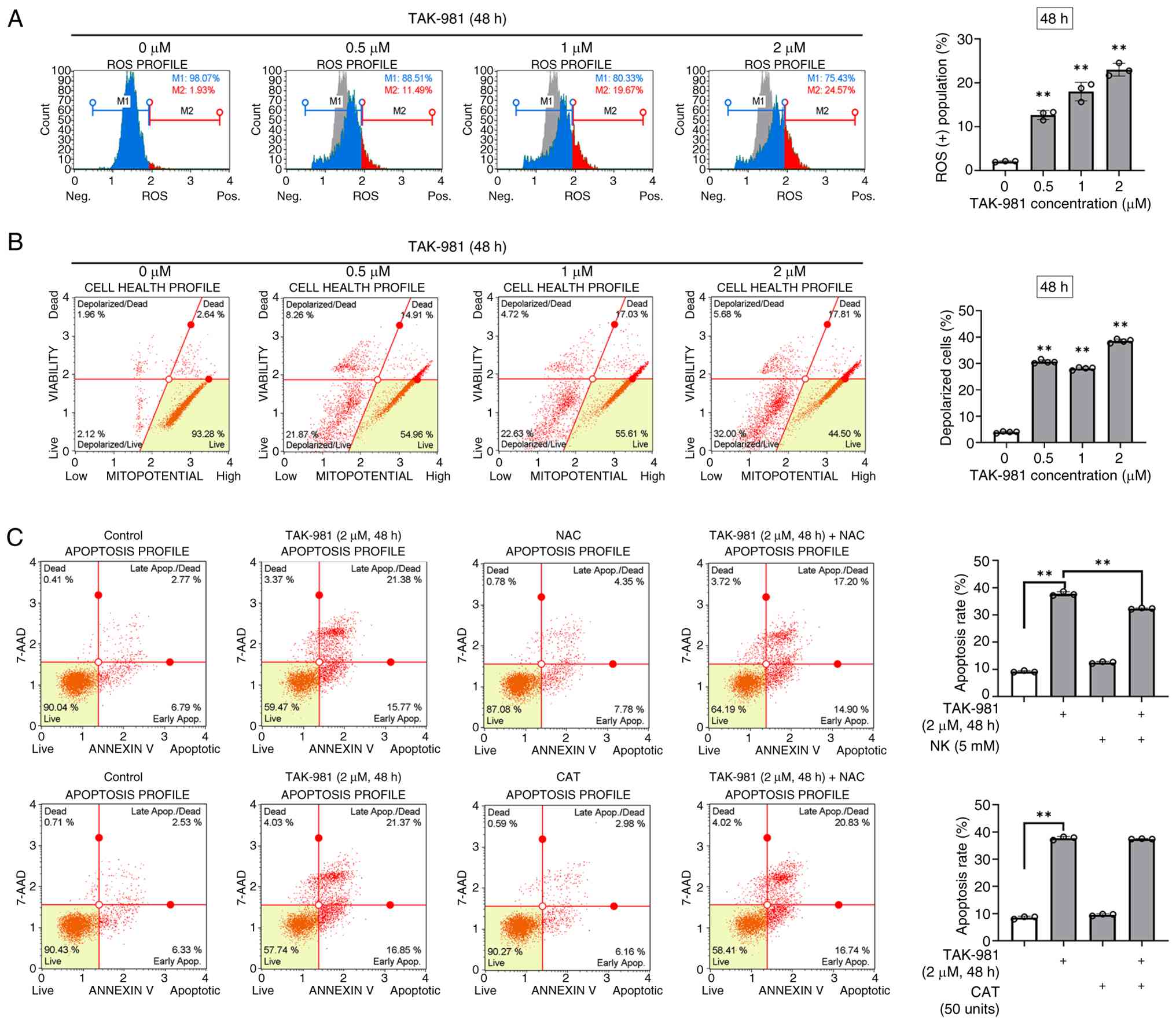
TAK-981-induced apoptosis is
associated with ROS accumulation and mitochondrial dysfunction in
SK-UT-1B cells
Whether oxidative stress contributed to
TAK-981-induced cytotoxicity was determined by treating SK-UT-1B
cells with TAK-981 (0.5–2 µM) for 48 h, and intracellular ROS
levels were measured using a fluorescent detection assay followed
by flow cytometric analysis. TAK-981 treatment markedly increased
the proportion of ROS-positive cells from 2.0% in the control group
to 12.6, 18.0, and 23.0% at 0.5, 1, and 2 µM, respectively
(Fig. 3A).
Mitochondrial membrane potential (ΔΨm) was evaluated
using the Muse MitoPotential assay after 48 h of TAK-981 exposure.
The proportion of total depolarized SK-UT-1B cells increased from
3.96% in the control group to 30.6, 28.1, and 38.5% after treatment
with 0.5, 1, and 2 µM TAK-981, respectively (Fig. 3B). SK-UT-1B cells were co-treated
with the antioxidants, NAC or CAT, for 48 h in the presence of 2 µM
TAK-981 to verify whether ROS accumulation mediated TAK-981-induced
apoptosis. NAC markedly reduced the TAK-981-induced apoptosis,
whereas CAT exerted little effect (Fig.
3C). These results indicate that TAK-981 increased ROS
accumulation and mitochondrial membrane depolarization in SK-UT-1B
cells; ROS contributed, at least in part, to TAK-981-induced
apoptotic cell death.
TAK-981 inhibits SUMOylation without
altering SAE1/SAE2 expression and modulates apoptotic and
cell-cycle regulators in SK-UT-1B cells
TAK-981 inhibits SUMOylation by blocking SAE2
activity without altering protein levels (10). The expression of SUMO2/3-conjugated
proteins, SAE1 and SAE2, was analyzed using western blotting to
examine whether TAK-981 affected SUMO pathway-related proteins in
SK-UT-1B cells. After 48 h of treatment with 0.5–2 µM TAK-981,
SUMO-conjugated proteins were markedly reduced, whereas SAE1 and
SAE2 protein levels were unchanged (Fig. 4A). These findings suggest that
TAK-981 suppressed SUMOylation primarily through functional
inhibition rather than through changes in SAE enzyme
expression.
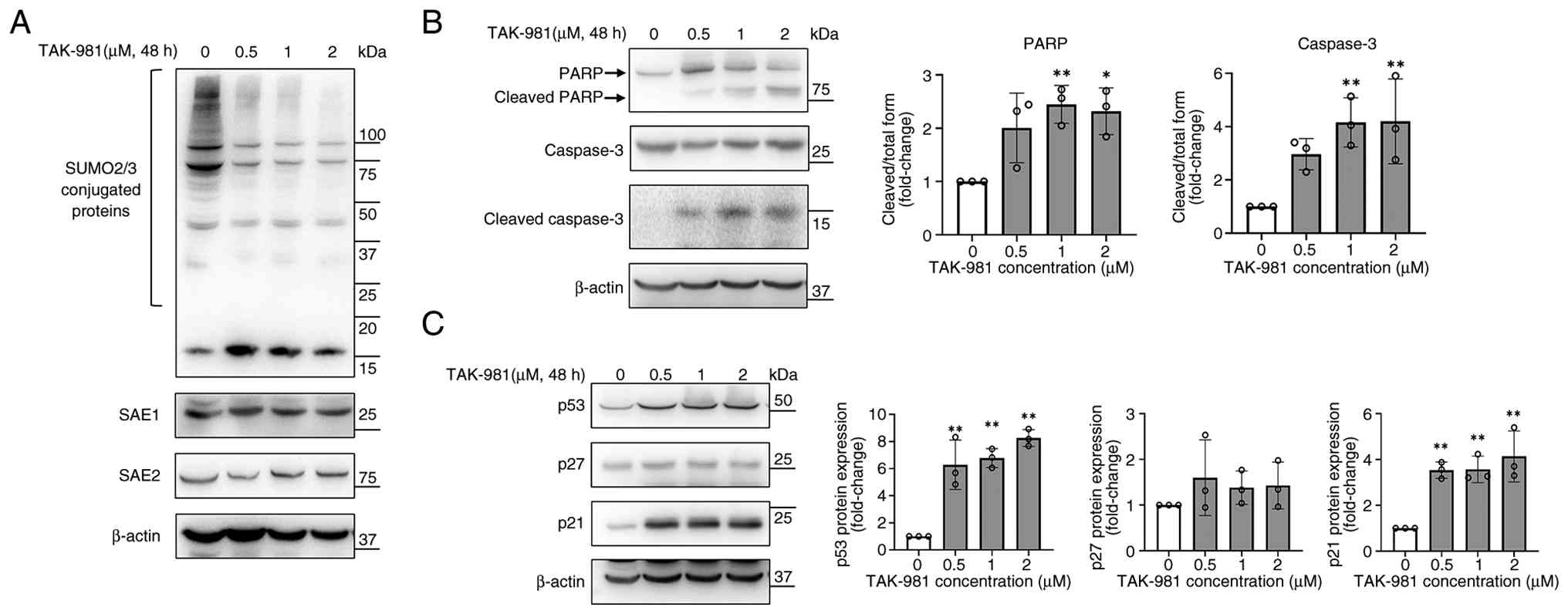 | Figure 4.TAK-981 inhibits SUMOylation and
modulates apoptotic- and cell-cycle-related pathways in SK-UT-1B
cells (A) SK-UT-1B cells were treated with TAK-981 (0.5, 1, and 2
µM) for 48 h, and the expression of SUMO2/3, SAE1, and SAE2 was
analyzed using western blotting with β-actin as a loading control.
(B) Apoptosis-associated proteins, including PARP, caspase-3 and
cleaved caspase-3, were analyzed using western blotting in SK-UT-1B
cells following 48 h of TAK-981 treatment. Semi-quantified band
intensities are shown in the right panels with β-actin serving as a
loading control. (C) Cell cycle-associated proteins, including p53,
p27 and p21, were analyzed using western blotting in SK-UT-1B cells
following 48 h of TAK-981 treatment. Semi-quantified band
intensities are shown in the right panels with β-actin serving as a
loading control. Data represent mean ± standard deviation from
three independent experiments. *P<0.05, **P<0.01 vs. control.
PARP, poly (ADP-ribose) polymerase; SAE, SUMO-activating enzyme;
SUMO, small ubiquitin-like modifier. |
The expression of caspase-3, cleaved caspase-3, and
PARP was analyzed to investigate the apoptosis-related signaling
events. TAK-981 treatment enhanced caspase-3 and PARP cleavage
after 48 h, indicating the activation of apoptotic signaling in
SK-UT-1B cells (Fig. 4B). The
downstream mechanisms associated with TAK-981-mediated growth
suppression were determined by examining the expression of key cell
cycle regulators. Western blotting indicated that TAK-981 markedly
increased p53 and p21 protein expression after 48 h of treatment,
whereas p27 levels were unchanged in SK-UT-1B cells (Fig. 4C). Collectively, these findings
indicate that TAK-981 inhibited SUMOylation, increased p53 and p21
protein expression, and triggered apoptotic signaling cascades in
SK-UT-1B cells.
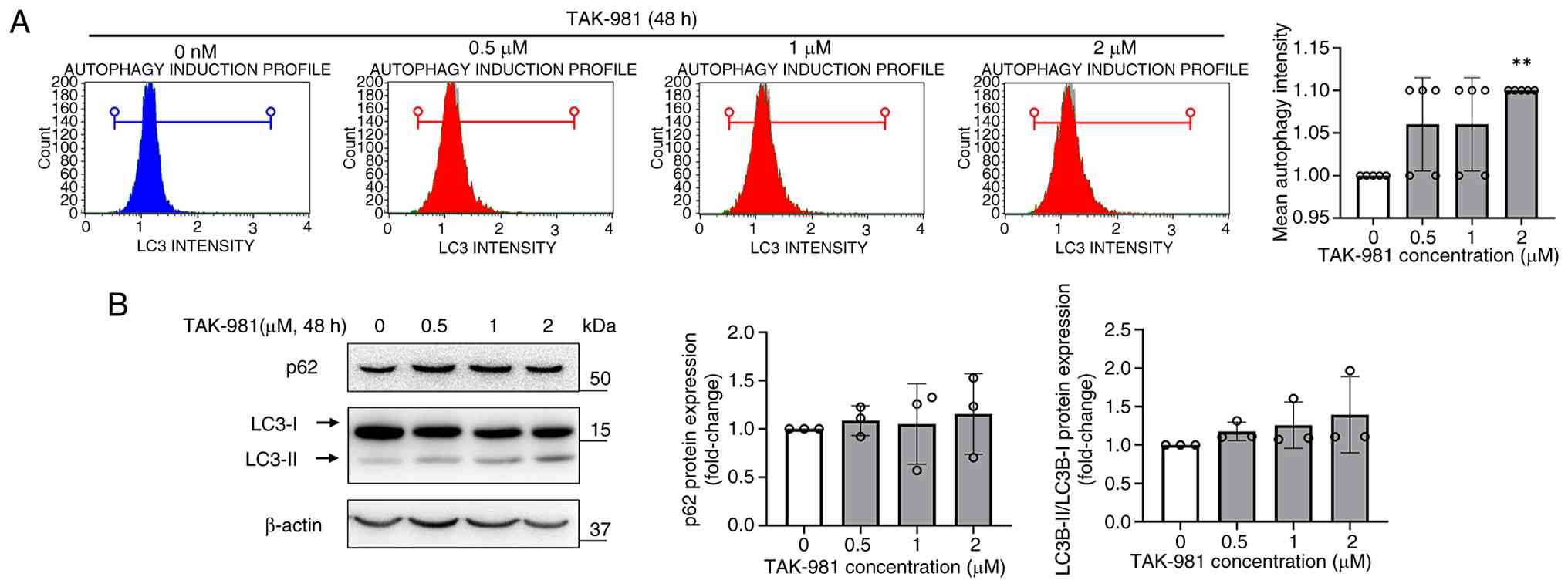
TAK-981 does not significantly
modulate autophagy in SK-UT-1B cells
Autophagy plays a key role in maintaining cell
survival and homeostasis under normal and stressful conditions.
Whether TAK-981 affected autophagy in SK-UT-1B cells was
investigated by measuring LC3B expression by flow cytometry after
treatment with 0.5, 1, or 2 µM TAK-981 for 48 h. TAK-981 treatment
did not alter LC3B fluorescence intensity markedly at most
concentrations, although a slight increase was observed at 2 µM
(Fig. 5A). Consistent with this,
western blotting indicated no marked changes in LC3B-I/II
conversion or p62 expression after 48 h of TAK-981 treatment
(Fig. 5B). TAK-981 therefore did
not modulate autophagy in SK-UT-1B cells, suggesting that its
cytotoxic effects were largely independent of the autophagic
pathway.
Discussion
This study provides the first evidence that the SUMO
inhibitor TAK-981 suppressed the viability of SK-UT-1B cells, even
at low concentrations, and was associated with ROS accumulation,
mitochondrial membrane depolarization, apoptosis, and
G0/G1 cell cycle arrest, accompanied by the
activation of p21 and p53. These results suggest that TAK-981
disrupts Ut-LMS cell survival by simultaneously engaging in
ROS-dependent apoptotic signaling and SUMO-inhibition-mediated
transcriptional pathways.
The present study extends our previous findings in
ELT3 uterine leiomyoma cells to a malignant Ut-LMS cell model
(17). Despite their shared uterine
smooth muscle origin, TAK-981-associated cellular responses
differed between benign leiomyoma and malignant Ut-LMS cells.
TAK-981 increased ROS accumulation in both ELT3 and SK-UT-1B cells;
however, apoptosis appeared to be ROS-independent in ELT3 cells,
whereas ROS-associated mitochondrial dysfunction was more closely
linked to apoptosis in SK-UT-1B cells. In addition, TAK-981 was
associated with G2/M arrest, MEK/ERK inhibition, reduced
extracellular matrix-related protein expression, and
autophagy-related changes in ELT3 cells, whereas it was associated
with G0/G1 arrest and p53/p21 activation in
SK-UT-1B cells. These differences suggest that cellular responses
to TAK-981 may depend on tumor type and cellular context, and
further studies are needed to clarify these model-dependent
effects.
Post-translational modification mechanisms have
emerged as critical determinants of tumor survival, proliferation,
and therapeutic resistance (18–20).
Of these, the SUMO pathway has gained particular attention as a key
regulatory axis that governs essential cellular processes, such as
DNA damage repair, cell-cycle progression, stress responses,
transcriptional regulation, and protein stability, which are
tightly linked to tumor growth and progression (13,21).
Aberrant SUMOylation has been reported to promote malignant
transformation, cancer stemness, drug resistance, and metastasis in
multiple tumor types (11,13,22).
Consistent with these determinations, the current study showed that
TAK-981 markedly inhibited global SUMOylation in SK-UT-1B cells and
affected multiple cellular processes, including reduced cell
viability, G0/G1 cell-cycle arrest, ROS
accumulation, mitochondrial dysfunction, and apoptosis.
Pharmacological inhibition of the SUMO cascade, particularly
through SAE inhibition by TAK-981, may therefore represent a
promising therapeutic strategy for Ut-LMS (14,15,17).
A notable aspect of the findings was the
differential sensitivity of Ut-LMS cell lines. SK-UT-1B cells
exhibited marked susceptibility to low micromolar concentrations of
TAK-981, whereas SK-UT-1 cells demonstrated limited responses at
similar concentrations. Several biological factors may be
responsible for this variability. Although SK-UT-1B is a sub-line
derived from the parental SK-UT-1 cell line, it displays distinct
molecular characteristics, including a potentially wild-type p53
background compared to the p53-mutant status reported for SK-UT-1
(23–25). As the p53 status influences cell
cycle regulation, DNA damage responses, and oxidative stress
sensitivity (26–28), TAK-981-induced p21/p53 activation
may exert greater biological effects on SK-UT-1B cells. Based on
our findings, the higher sensitivity of SK-UT-1B cells to TAK-981
may be related to their greater dependence on
SUMOylation-associated stress-adaptive mechanisms and their
susceptibility to p53/p21-mediated cell-cycle arrest and
ROS-associated mitochondrial apoptosis. In contrast, SK-UT-1 cells
may possess distinct molecular features that confer relative
resistance to TAK-981, such as altered p53 pathway activity,
reduced SUMO pathway dependency, or increased tolerance to
oxidative and mitochondrial stress. These intrinsic differences
highlight the significance of molecular heterogeneity in Ut-LMS
cells and underscore the need to tailor targeted therapies based on
the tumor genotype and pathway dependency.
Compared with conventional agents, such as
doxorubicin, gemcitabine, or docetaxel, which generally exert
nonspecific cytotoxic effects, TAK-981 is highly specific as it
directly inhibits the SUMOylation machinery (29). This provides a novel therapeutic
approach for Ut-LMS, particularly because SUMO-dependent
transcriptional and stress response pathways are increasingly
recognized as actionable vulnerabilities in high-grade sarcomas.
The robust cytotoxicity observed in SK-UT-1B cells at
pharmacologically-relevant concentrations supports the existence of
a therapeutic window that distinguishes TAK-981 from traditional
chemotherapeutics that often damage malignant and healthy
tissues.
From a translational perspective, the SUMO pathway
inhibition may offer opportunities for precision-guided Ut-LMS
therapy. The molecular diversity observed between SK-UT-1B and
SK-UT-1 cells suggests that biomarkers, such as p53 functionality,
SUMO pathway dependency, and ROS-related gene signatures, could
serve to stratify patients most likely to benefit from TAK-981.
Furthermore, combining SUMO inhibition with agents targeting DNA
damage repair, oxidative stress pathways, or immune activation may
enhance therapeutic responses due to the known role of TAK-981 in
amplifying type I interferon signaling (9,10).
This study had several limitations. These findings
are based solely on in vitro experiments, and additional
in vivo validation using animal models or patient-derived
xenografts is required to confirm the therapeutic potential of
TAK-981. Furthermore, the molecular factors underlying the
differential sensitivity among Ut-LMS cell lines warrant
comprehensive investigation through genomic, transcriptomic, and
proteomic analyses.
In conclusion, the findings provide the first
evidence that TAK-981 exerts strong anti-proliferative and
pro-apoptotic effects in SK-UT-1B cells. These effects were
associated with SUMOylation inhibition, ROS accumulation,
mitochondrial dysfunction, G0/G1 cell-cycle
arrest, and activation of p53/p21 and apoptotic signaling. These
data highlight TAK-981 as a promising candidate for future
therapeutic development and support the broader concept of
targeting SUMOylation as an emerging strategy for the treatment of
aggressive Ut-LMS.
Acknowledgements
No applicable.
Funding
This work was supported by a research fund from Chosun
University, 2025 (grant no. K208554004).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
HJ and HL designed the experiments and revised the
manuscript. HJ conducted experiments and wrote the manuscript. HJ
and HL conducted the data analysis. HJ and HL confirm the
authenticity of all the raw data. Both authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Ut-LMS
|
uterine leiomyosarcoma
|
|
SAE
|
SUMO-activating enzyme
|
|
DMSO
|
dimethyl sulfoxide
|
|
LDH
|
lactate dehydrogenase
|
|
ROS
|
reactive oxygen species
|
|
NAC
|
N-Acetyl-L-cysteine
|
|
CAT
|
catalase
|
|
PARP
|
poly (ADP-ribose) polymerase
|
|
SUMO
|
small ubiquitin-like modifier
|
References
|
1
|
Bogani G, Caruso G, Ray-Coquard I, Ramirez
PT, Concin N, Ngoi NY, Coleman RL, Mariani A, Cliby W, Leitao MM,
et al: Uterine leiomyosarcoma. Int J Gynecol Cancer. 35:1019922025.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Khamaiseh S, Koivisto-Korander R,
Schreiber N, Pitkanen E, Ahvenainen T, Butzow R, Mehine M and
Vahteristo P: Transcriptome profiling of uterine leiomyosarcomas
identifies a leiomyoma-like expression pattern that indicates
better survival. BJC Rep. 3:762025. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang Q, Madueke-Laveaux OS, Cun H,
Wlodarczyk M, Garcia N, Carvalho KC and Al-Hendy A: Comprehensive
review of uterine leiomyosarcoma: pathogenesis, diagnosis,
prognosis, and targeted therapy. Cells. 13:11062024. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Henry T, Fabre E, Baccar LS and Lamuraglia
M: Longer survival in patients with metastatic uterine
leiomyosarcoma treated with trabectedin: A case report. Mol Clin
Oncol. 10:387–390. 2019.PubMed/NCBI
|
|
5
|
Gupta AA, Yao X, Verma S, Mackay H and
Hopkins L; Sarcoma Disease Site Group and the Gynecology Cancer
Disease Site Group, : Systematic chemotherapy for inoperable,
locally advanced, recurrent, or metastatic uterine leiomyosarcoma:
A systematic review. Clin Oncol (R Coll Radiol). 25:346–355. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sparic R, Andjic M, Babovic I, Nejkovic L,
Mitrovic M, Stulic J, Pupovac M and Tinelli A: Molecular insights
in uterine leiomyosarcoma: A systematic review. Int J Mol Sci.
23:97282022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chudasama P, Mughal SS, Sanders MA,
Hubschmann D, Chung I, Deeg KI, Wong SH, Rabe S, Hlevnjak M,
Zapatka M, et al: Integrative genomic and transcriptomic analysis
of leiomyosarcoma. Nat Commun. 9:1442018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dall GV, Hamilton A, Ratnayake G, Scott C
and Barker H: Interrogating the genomic landscape of uterine
leiomyosarcoma: A potential for patient benefit. Cancers (Basel).
14:15612022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Langston SP, Grossman S, England D, Afroze
R, Bence N, Bowman D, Bump N, Chau R, Chuang BC, Claiborne C, et
al: Discovery of TAK-981, a first-in-class inhibitor of
SUMO-activating enzyme for the treatment of cancer. J Med Chem.
64:2501–2520. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lightcap ES, Yu P, Grossman S, Song K,
Khattar M, Xega K, He X, Gavin JM, Imaichi H, Garnsey JJ, et al: A
small-molecule SUMOylation inhibitor activates antitumor immune
responses and potentiates immune therapies in preclinical models.
Sci Transl Med. 13:eaba77912021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gu Y, Fang Y, Wu X, Xu T, Hu T, Xu Y, Ma
P, Wang Q and Shu Y: The emerging roles of SUMOylation in the tumor
microenvironment and therapeutic implications. Exp Hematol Oncol.
12:582023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eifler K and Vertegaal ACO:
SUMOylation-mediated regulation of cell cycle progression and
cancer. Trends Biochem Sci. 40:779–793. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Han ZJ, Feng YH, Gu BH, Li YM and Chen H:
The post-translational modification, SUMOylation, and cancer
(Review). Int J Oncol. 52:1081–1094. 2018.PubMed/NCBI
|
|
14
|
Kumar S, Schoonderwoerd MJA, Kroonen JS,
de Graaf IJ, Sluijter M, Ruano D, Gonzalez-Prieto R, Verlaan-de
Vries M, Rip J, Arens R, et al: Targeting pancreatic cancer by
TAK-981: A SUMOylation inhibitor that activates the immune system
and blocks cancer cell cycle progression in a preclinical model.
Gut. 71:2266–2283. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim HS, Kim BR, Dao TTP, Kim JM, Kim YJ,
Son H, Jo S, Kim D, Kim J, Suh YJ, et al: TAK-981, a SUMOylation
inhibitor, suppresses AML growth immune-independently. Blood Adv.
7:3155–3168. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou H, Deng N, Li Y, Hu X, Yu X, Jia S,
Zheng C, Gao S, Wu H and Li K: Distinctive tumorigenic significance
and innovative oncology targets of SUMOylation. Theranostics.
14:3127–3149. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu H, Seo S and Joung H: TAK-981 enhances
antitumor activity in ELT3 uterine leiomyoma cells through the
modulation of apoptosis, cell cycle arrest, and autophagy. Biochem
Biophys Res Commun. 770:1520002025. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bruno PS, Arshad A, Gogu MR, Waterman N,
Flack R, Dunn K, Darie CC and Neagu AN: Post-translational
modifications of proteins orchestrate all hallmarks of cancer. Life
(Basel). 15:1262025.PubMed/NCBI
|
|
19
|
Dutta H and Jain N: Post-translational
modifications and their implications in cancer. Front Oncol.
13:12401152023. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li W, Li F, Zhang X, Lin HK and Xu C:
Insights into the post-translational modification and its emerging
role in shaping the tumor microenvironment. Signal Transduct Target
Ther. 6:4222021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kroonen JS and Vertegaal ACO: Targeting
SUMO signaling to wrestle cancer. Trends Cancer. 7:496–510. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kukkula A, Ojala VK, Mendez LM, Sistonen
L, Elenius K and Sundvall M: Therapeutic potential of targeting the
SUMO pathway in cancer. Cancers (Basel). 13:44022021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Joung H, Seo S and Liu H: MG132 induces
cell type-specific anticancer effects in uterine leiomyosarcoma
cell lines. Mol Med Rep. 31:2025. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Smardova J, Pavlova S, Svitakova M,
Grochova D and Ravcukova B: Analysis of p53 status in human cell
lines using a functional assay in yeast: Detection of new non-sense
p53 mutation in codon 124. Oncol Rep. 14:901–907. 2005.PubMed/NCBI
|
|
25
|
Coley HM, Shotton CF, Kokkinos MI and
Thomas H: The effects of the CDK inhibitor seliciclib alone or in
combination with cisplatin in human uterine sarcoma cell lines.
Gynecol Oncol. 105:462–469. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pitolli C, Wang Y, Candi E, Shi Y, Melino
G and Amelio I: p53-mediated tumor suppression: DNA-damage response
and alternative mechanisms. Cancers (Basel). 11:19832019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Beyfuss K and Hood DA: A systematic review
of p53 regulation of oxidative stress in skeletal muscle. Redox
Rep. 23:100–117. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vousden KH and Prives C: Blinded by the
light: The growing complexity of p53. Cell. 137:413–431. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kroonen JS, de Graaf IJ, Kumar S, Remst
DFG, Wouters AK, Heemskerk MHM and Vertegaal ACO: Inhibition of
SUMOylation enhances DNA hypomethylating drug efficacy to reduce
outgrowth of hematopoietic malignancies. Leukemia. 37:864–876.
2023. View Article : Google Scholar : PubMed/NCBI
|