Introduction
Osteosarcoma (OS) is a common primary malignant bone
tumor predominantly affecting adolescents and young adults. A
population-based study using the Surveillance, Epidemiology, and
End Results (SEER) database reported that the 5-year survival rate
for patients with metastatic OS ranges from 10 to 30% (1). Despite advancements in multimodal
therapy for patients with localized OS, treatment outcomes for
patients with metastatic or recurrent disease remain limited, with
5-year survival rates ranging from 10–30% (2,3). OS is
highly metastatic, especially to the lungs (4), and is characterized by resistance to
conventional chemotherapy, with the underlying mechanisms remaining
complex and poorly understood (5,6).
Although genomic studies have revealed alterations in tumor
suppressor genes, such as tumor protein 53 and retinoblastoma 1,
existing targeted therapies have not effectively addressed the
challenges in OS treatment (6,7). This
suggests that, in addition to classical genetic mutations,
mechanisms such as post-translational modifications, epigenetic
regulation and protein degradation serves critical roles in the
onset and progression of OS, highlighting the need for further
research to uncover novel therapeutic targets.
The E3 ubiquitin ligase family is a core component
of the ubiquitin-proteasome system, serving crucial roles in
regulating protein stability, the cell cycle, signal transduction
and other processes. F-box and leucine-rich repeat protein 5
(FBXL5), a member of the Skp1-Cullin-F-box protein complex, has
been recognized as an important sensor of oxygen and iron
homeostasis, participating in the regulation of cellular iron
homeostasis by controlling the degradation of iron regulatory
protein 2 (IRP2) (8,9). FBXL5 not only serves a pivotal role in
iron metabolism but also mediates the ubiquitin-mediated
degradation of various substrates, thereby regulating cellular
processes such as cell cycle progression, DNA damage response and
apoptosis (10). These distinct
functions position FBXL5 as a potential regulatory factor with
significant roles in multiple biological processes.
Although the role of FBXL5 in various tumors has
been preliminarily investigated, to the best of our knowledge, its
specific function in OS remains underexplored. Studies have
demonstrated that FBXL5 exerts different roles in various types of
malignant tumors. For example, in gastric cancer, FBXL5 inhibits
the epithelial-mesenchymal transition (EMT) and tumor metastasis
through the ubiquitin-mediated degradation of the transcription
factor Snail1 (11). By contrast,
in hepatocellular carcinoma (12,13)
and clear cell renal cell carcinoma (ccRCC), the reduced expression
of FBXL5 is associated with a poor prognosis, suggesting that FBXL5
may function as a tumor suppressor in these cancers. However, the
role of FBXL5 in OS has not been extensively examined.
The mitogen-activated protein kinase
(MAPK)/extracellular signal-regulated kinase (ERK) signaling
pathway is a critical regulator of fundamental cellular processes
such as proliferation, migration, differentiation and apoptosis
(14), and serves a pivotal role in
the initiation and progression of various malignant tumors.
Activation of this pathway is mediated through a cascade of
signaling molecules, including Ras, Raf, MAPKK (MEK) and ERK,
ultimately regulating the activity of downstream transcription
factors and protein kinases to promote tumor cell proliferation and
migration (15). In OS, aberrant
activation of the MAPK/ERK signaling pathway is closely linked to
tumor metastasis and resistance (16,17).
Therefore, investigating the upstream regulatory mechanisms of this
pathway is crucial for understanding the progression of OS.
The present study investigated the potential role of
FBXL5 in the progression of OS. Using RNA sequencing (RNA-seq), we
analyzed the impact of FBXL5 knockdown on the transcriptome of OS
cells. Therefore, the aim of the present study was to investigate
the role of FBXL5 in OS and its potential interaction with the
MAPK/ERK signaling pathway. To address this aim, the present study
examined the effect of FBXL5 depletion on the expression of
MAPK/ERK pathway-related genes, as well as on OS cell
proliferation, apoptosis, migration and invasion. These findings
highlight the need to identify novel therapeutic targets and
understand their clinical relevance.
Materials and methods
Clinical samples
The present study collected tumor and paired normal
bone tissues from 6 patients with OS at The Second Hospital of
Lanzhou University (Lanzhou, China) between October 2021 and
December 2023. The cohort consisted of 6 patients (5 men; 1 woman),
aged 14–21 years, with pathological diagnoses of conventional OS
(n=5) and parosteal OS (n=1). The inclusion criteria were: i)
Histopathologically confirmed diagnosis of conventional or
parosteal OS; ii) aged between 10 and 25 years; iii) no prior
neoadjuvant chemotherapy or radiotherapy before surgery; iv)
availability of both tumor and paired normal bone tissue. The
exclusion criteria were: i) Recurrent or metastatic OS at
diagnosis; ii) history of other malignancies; iii) insufficient
tissue quality or quantity for downstream analysis. All samples
were immediately frozen in liquid nitrogen after surgery and stored
at −80°C. The study was approved by The Second Hospital of Lanzhou
University ethics committee (approval no. 2024A-504), and written
informed consent was obtained from all patients or their
guardians.
Cell culture and reagents
The human OS cell lines MG63 and 143B, along with
the normal osteoblast cell line hFOB1.19, were purchased from
Shanghai Yilei information Technology Co., Ltd. The cells were
cultured in high-glucose Dulbecco's Modified Eagle's Medium
(HyClone™; Cytiva) supplemented with 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.). MG63 and 143B cells were incubated
at 37°C, whereas hFOB1.19 cells were maintained at 34°C.
Lentivirus-mediated gene
knockdown
FBXL5-targeting short hairpin RNA (shRNA) lentiviral
constructs (cat. no. GXDL0418586; FBXL5 Human GV493) were obtained
from Shanghai GeneChem Co., Ltd. The target sequences were as
follows: FBXL5-RNAi (118431–1): GTACAGGAACAGCTTTAAGAA; FBXL5-RNAi
(118432–1): CCTGATGATGAATGGGTGAAA; FBXL5-RNAi (118433–1):
GCACTTTACTCATAGCACATT. A second-generation lentiviral packaging
system was used, including the shuttle vector GV493
(hU6-MCS-CBh-gcGFP-IRES-puromycin) and packaging plasmids psPAX2
and pMD2.G. Lentiviral particles were produced in 293T cells
(Shanghai GeneChem Co., Ltd.) according to the manufacturer's
protocol. Plasmids were transfected at a ratio of 20 (GV493), 15
(psPAX2) and 10 µg (pMD2.G), followed by incubation at 37°C with 5%
CO2 for 48 h. Viral supernatants were collected and
concentrated by ultracentrifugation (13,680 × g; 2 h; 4°C). For
transduction, 143B and MG63 cells were infected with lentivirus at
multiplicities of infection (MOI) of 15 and 30, respectively, in
the presence of polybrene (8 µg/ml). After 48 h, stable clones were
selected using puromycin (2 µg/ml), and selection was maintained
for 7 days to establish stable knockdown pools. Knockdown
efficiency was confirmed by western blotting (WB) analysis and
reverse transcription-quantitative PCR (RT-qPCR). All subsequent
experiments were performed 10 days after the initial
transduction.
RNA extraction and RT-qPCR
Total RNA was extracted from cells using Trizol
(total RNA extraction reagent; cat. no. R0016; Beyotime
Biotechnology). RT was performed using the Evo M-MLV One-step
RT-PCR kit (with gDNA removal, for qPCR; cat. no. AG11705; AGbio)
according to the manufacturer's protocol. The reaction conditions
for RT were: 37°C for 15 min (gDNA removal), 42°C for 15 min (RT),
followed by 85°C for 5 sec (enzyme inactivation). For qPCR, the
reaction was performed using SYBR Green Master Mix (cat. no.
AG11732; AGbio), which contains SYBR Green as the fluorophore. The
following thermocycling conditions were applied: initial
denaturation at 95°C for 30 sec, followed by 40 cycles of 95°C for
5 sec and 60°C for 30 sec. GAPDH was used as the reference gene.
The primer sequences used are provided in Table I. Relative expression levels were
calculated using the 2−ΔΔCq method (18).
 | Table I.Primer sequences used in the
quantitative PCR experiment. |
Table I.
Primer sequences used in the
quantitative PCR experiment.
| Primer | Primer sequence
(5′-3′) |
|---|
| FBXL5 forward |
TTGTCCTAACCTGGAGCATCTGG |
| FBXL5 reverse |
GCAACCAAGCCAAGACCAACTG |
| GAPDH forward |
AGGTGAAGGTCGGAGTCAACG |
| GAPDH reverse |
GCTCCTGGAAGATGGTGATGG |
Protein extraction and western blot
analysis
Total proteins were extracted from cells using RIPA
lysis buffer (cat. no. P0013B; Beyotime Biotechnology) and
quantified by the BCA method. Proteins (30 µg/lane) were separated
by 10% SDS-PAGE and electrotransferred to a PVDF membrane
(MilliporeSigma). After blocking with 5% non-fat milk (cat. no.
MA0406; Dalian Meilun Biology Technology Co., Ltd.cat. no.) at 4°C
for 1 h, the membrane was incubated with primary antibodies at 4°C
overnight. The primary antibodies and their cat. nos. were as
follows: FBXL5 (1:1,000; cat. no. BD-PT6983; Biodragon
Immunotechnologies Co., Ltd.), PCNA (1:2,000; cat. no. 10205-2-AP;
Proteintech Group, Inc.), CDK4 (1:1,000; cat. no. 11026-1-AP
Proteintech Group, Inc.), cyclin D1 (1:1,000; cat. no. 60186-1-Ig;
Proteintech Group, Inc.), Bcl-2 (1:1,000; cat. no. AF6139; Affinity
Biosciences, Ltd.), Bax (1:1,000; cat. no. AF0120; Affinity
Biosciences, Ltd.), cleaved caspase-3 (1:1,000; cat. no. AF7022;
Affinity Biosciences, Ltd.), total caspase-3 (1:1,000; cat. no.
db12058; Hangzhou Diagbio Technology Co., Ltd.), MMP2 (1:1,000;
cat. no. RM8377; Biodragon Immunotechnologies Co., Ltd.), MMP9
(1:1,000; cat. no. RM3763; Biodragon Immunotechnologies Co., Ltd.),
MEK1/2 (1:1,000; HUABIO; cat. no. ET1602-3), ERK (1:1,000; cat. no.
ET1601-29; HUABIO), Ras (1:1,000; cat. no. RM7373; Biodragon
Immunotechnologies Co., Ltd.), Raf (1:1,000; cat. no. RM4242;
Biodragon Immunotechnologies Co., Ltd.), and β-actin (1:8,000; cat.
no. HA722023; HUABIO). After washing, the membrane was incubated
with horseradish peroxidase-conjugated goat anti-rabbit/mouse IgG
(1:10,000; cat. no. ZB-2306; Beijing Zhongshan Jinqiao
Biotechnology Co., Ltd.) at 4°C for 1 h. Proteins were detected
using enhanced chemiluminescence substrate (cat. no. G2020-500ML;
Wuhan Servicebio Technology Co., Ltd.). Densitometric analysis was
performed using ImageJ software (version 1.52a; National Institutes
of Health).
Immunofluorescence
143B and MG63 cells were seeded on coverslips, fixed
with 4% paraformaldehyde in PBS at room temperature for 15 min,
permeabilized with 0.1% Triton X-100 in PBS for 10 min at room
temperature and blocked with 5% normal goat serum (cat. no. SL038;
Beijing Solarbio Science & Technology Co., Ltd.) in PBS at room
temperature for 1 h. After blocking, cells were incubated overnight
at 4°C with the following primary antibodies: FBXL5 (1:100; Santa
Cruz Biotechnology, Inc.; cat. no. sc-390102), PCNA (1:100; cat.
no. 10205-2-AP; Proteintech Group, Inc.cat. no.), CDK4 (1:100; cat.
no. 11026-1-AP; Proteintech Group, Inc.), and cyclin D1 (1:100;
cat. no. 60186-1-Ig; Proteintech Group, Inc.cat. no.). After
washing, cells were incubated with Alexa Fluor 488-conjugated
secondary antibody (1:200; Biodragon Immunotechnologies Co., Ltd.;
cat. no. BD9010) at room temperature for 1 h. Nuclei were
counterstained with DAPI (1 µg/ml) at room temperature for 5 min.
Fluorescence images were captured using a fluorescence
microscope.
Functional assays
Proliferation: Cell viability was assessed using the
Cell Counting Kit-8 (CCK-8) assay (cat. no. C0038; Beyotime
Biotechnology). Cells were seeded in 96-well plates and cultured
for 24, 48 and 72 h. At each time point, CCK-8 reagent was added
and incubated for 2 h at 37°C, and the absorbance was measured at
450 nm using a microplate reader. Proliferation rates were also
determined using 5-ethynyl-2′-deoxyuridine (EdU) staining with the
EdU-488 Cell Proliferation Kit (cat. no. C0071S; Beyotime
Biotechnology) according to the manufacturer's instructions.
Briefly, cells were incubated with EdU at 37°C for 2 h, then fixed
with 4% paraformaldehyde in PBS at room temperature for 15 min,
permeabilized with 0.3% Triton X-100 in PBS and stained with the
Click-It reaction mixture. Nuclei were counterstained with DAPI
(cat. no. C1006; Beyotime Biotechnology). Images were captured
using a fluorescence microscope.
Apoptosis: Apoptotic cells were identified
using the TUNEL staining. Cells were fixed with 4% paraformaldehyde
in PBS at room temperature for 15 min, permeabilized with 0.1%
Triton X-100 in PBS for 5 min and then incubated with TUNEL
reaction mixture (cat. no. C1088; Beyotime Biotechnology) at 37°C
for 60 min according to the manufacturer's protocol. Nuclei were
counterstained with DAPI (1 µg/ml) at room temperature for 5 min.
After rinsing with PBS, the slides were mounted using anti-fade
mounting medium (Beyotime Biotechnology China; cat. no. P0126). At
least five random fields of view per sample were observed under a
fluorescence microscope, and TUNEL-positive cells were counted.
Migration: A wound healing assay was
performed to measure cell migration. No substrate coating was used.
143B and MG63 cells were seeded in 6-well plates and cultured until
reaching 90–100% confluence. A linear scratch was created across
the cell monolayer using a sterile 200 µl pipette tip. Debris was
removed by washing with PBS, and cells were then cultured in Gibco
(Thermo Fisher Scientific, Inc.) basal medium to exclude the effect
of proliferation. Wound closure was monitored and imaged at 0, 24,
48, 72 and 96 h under a light microscope. The wound area was
measured using ImageJ software (version 1.52a, National Institutes
of Health), and the percentage of wound closure was calculated as:
[(area at 0 h-area at time point)/area at 0 h] ×100%. No drugs were
used in this assay.
Animal experiment
A total of 36 BALB/c nude mice (4–6 weeks old;
female; weighing 16–18 g) were obtained from Lanzhou Veterinary
Research Institute, Chinese Academy of Agricultural Sciences. Mice
were housed under specific pathogen-free conditions with
temperature 22±2°C, humidity 50±10%, a 12 h light/dark cycle, and
ad libitum access to food and water. Mice were randomly assigned to
six groups (n=6 per group): Group 1: 143B + control; group 2: 143B
+ sh-NC; group 3: 143B + sh-FBXL5; group 4: MG63 + control; group
5: MG63 + sh-NC; group 6: MG63 + sh-FBXL5. A xenograft tumor model
was established by subcutaneously injecting 2×106 cells
into the right axilla of each mouse. Tumor volume was measured
every 3 days using a caliper and calculated as volume=(length ×
width2)/2. The ethical endpoint was defined as tumor
volume >1,500 mm3, tumor ulceration, substantial
weight loss (>20% of initial body weight) or signs of
pain/distress. Tumor volume and body weight were measured every 3
days from day 0 until the endpoint for each group. The 143B
xenografts grew more rapidly and reached the ethical endpoint
(tumor volume >1,500 mm3) by day 30, whereas the MG63
×enografts grew more slowly and did not reach the endpoint until
day 34. Therefore, mice in the 143B and MG63 groups were euthanized
on day 30 and day 34, respectively, to prevent exceeding the
predefined ethical endpoints. No mice were euthanized before the
scheduled endpoints. Mice in the 143B and MG63 groups were
euthanized by CO2 inhalation (30–50% vol/min) on day 30
and day 34, respectively. The tumor tissues were then processed for
subsequent analysis. The animal study was approved by the Animal
Ethics Committee of Lanzhou University Second Hospital (approval
no. D2024-417).
RNA-seq
Total RNA was extracted from cells using TRIzol
reagent (cat. no. R0016; Beyotime Biotechnology). RNA integrity was
assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies,
USA). Sequencing libraries were prepared using the TruSeq RNA
Sample Prep Kit v2 (Illumina, Inc.; cat. no. RS-122-2001) according
to the manufacturer's protocol. The libraries were sequenced on the
Illumina NovaSeq 6000 platform (Illumina, Inc.) using the NovaSeq
6000 S4 Reagent Kit v1.5 (300 cycles; Illumina, Inc.; cat. no.
20028312) with 150 bp paired-end sequencing. The final library was
loaded at a concentration of 1.8 pM, as measured by Qubit 2.0
Fluorometer (Thermo Fisher Scientific, Inc.). For data analysis,
Trimmomatic (version 0.39; http://www.usadellab.org/cms/index.php?page=trimmomatic)
was used for quality control, HISAT2 (version 2.2.1; http://ccb.jhu.edu/software/hisat2/) was
used for alignment to the reference genome, and featureCounts
(version 2.0.1; http://subread.sourceforge.net/) was used to quantify
gene expression levels. edgeR (version 3.40.0) was applied to
identify differentially expressed genes (DEGs) with criteria of
|log2FC|≥1 and FDR ≤0.05. Functional enrichment analysis was
conducted using GOATools (version 1.0.0; http://github.com/tanghaibao/GOatools) for Gene
Ontology (GO) enrichment and KOBAS (version 3.0; http://bioinfo.org/kobas3/) for KEGG pathway
enrichment. DAVID (Database for Annotation, Visualization and
Integrated Discovery; http://david.ncifcrf.gov) and GSEA (Gene Set
Enrichment Analysis; http://www.gsea-msigdb.org/gsea/index.jsp) were also
used for functional annotation and enrichment analysis.
Immunofluorescence (IF)
After deparaffinization in xylene and rehydration
through a graded ethanol series, antigen retrieval was performed by
heating the paraffin-embedded tissue sections in citrate buffer (pH
6.0) at 95°C for 15 min. Sections were incubated with primary
antibodies against Raf (1:100; cat. no. RM4242; Suzhou Botron
Immunotherapy Technology Co., Ltd.) and MEK1/2 (1:100; HUABIO; cat.
no. ET1602-3) at 4°C overnight. After washing, sections were
incubated with Alexa Fluor 488-conjugated secondary antibody
(1:200; Biodragon Immunotechnologies Co., Ltd.; cat. no. BD9010) at
room temperature for 1 h. Nuclei were counterstained with DAPI (1
µg/ml) at room temperature for 10 min. Fluorescence images were
captured using a fluorescence microscope.
H&E staining
Paraffin-embedded tissue sections were
deparaffinized in xylene and rehydrated through a graded ethanol
series (100, 95, 70 and 50%) for 2 min each, followed by rinsing in
distilled water. Sections were stained with hematoxylin at room
temperature for 5 min, followed by eosin staining at room
temperature for 2 min. Sections were then dehydrated through a
graded ethanol series (70, 95 and 100%), cleared in xylene, and
mounted with a coverslip using a resinous mounting medium. Images
were captured using a light microscope (Olympus Corporation) and
analyzed using ImageJ software (version 1.52a; National Institutes
of Health).
Statistical analysis
All experimental data were statistically analyzed
using GraphPad Prism 8.0.2 software (GraphPad; Dotmatics). A paired
Student's t-test was used when comparing two measurements from the
same subject, whereas an unpaired Student's t-test was used when
comparing data from different groups of subjects. For multiple
groups, one-way ANOVA was used, followed by Tukey's multiple
comparisons test. Data are expressed as the mean ± standard
deviation from six independent repeats. Fluorescence images and WB
bands were analyzed and measured using ImageJ 1.52a software
(National Institutes of Health). P<0.05 was considered to
indicate a statistically significant difference.
Results
FBXL5 expression in OS tissues and
cells is significantly higher compared with normal controls, and a
FBXL5 knockdown model was successfully established in 143B and MG63
cells
qPCR and WB analyses revealed that FBXL5 protein
(Fig. 1A) and mRNA (Fig. 1B) expression levels were
significantly elevated in OS tissues compared to normal tissues. In
OS cell lines 143B and MG63, FBXL5 mRNA (Fig. 1D and F) and protein (Fig. 1C and E) expression was also
significantly upregulated compared with the normal osteoblast cell
line hFOB1.19.
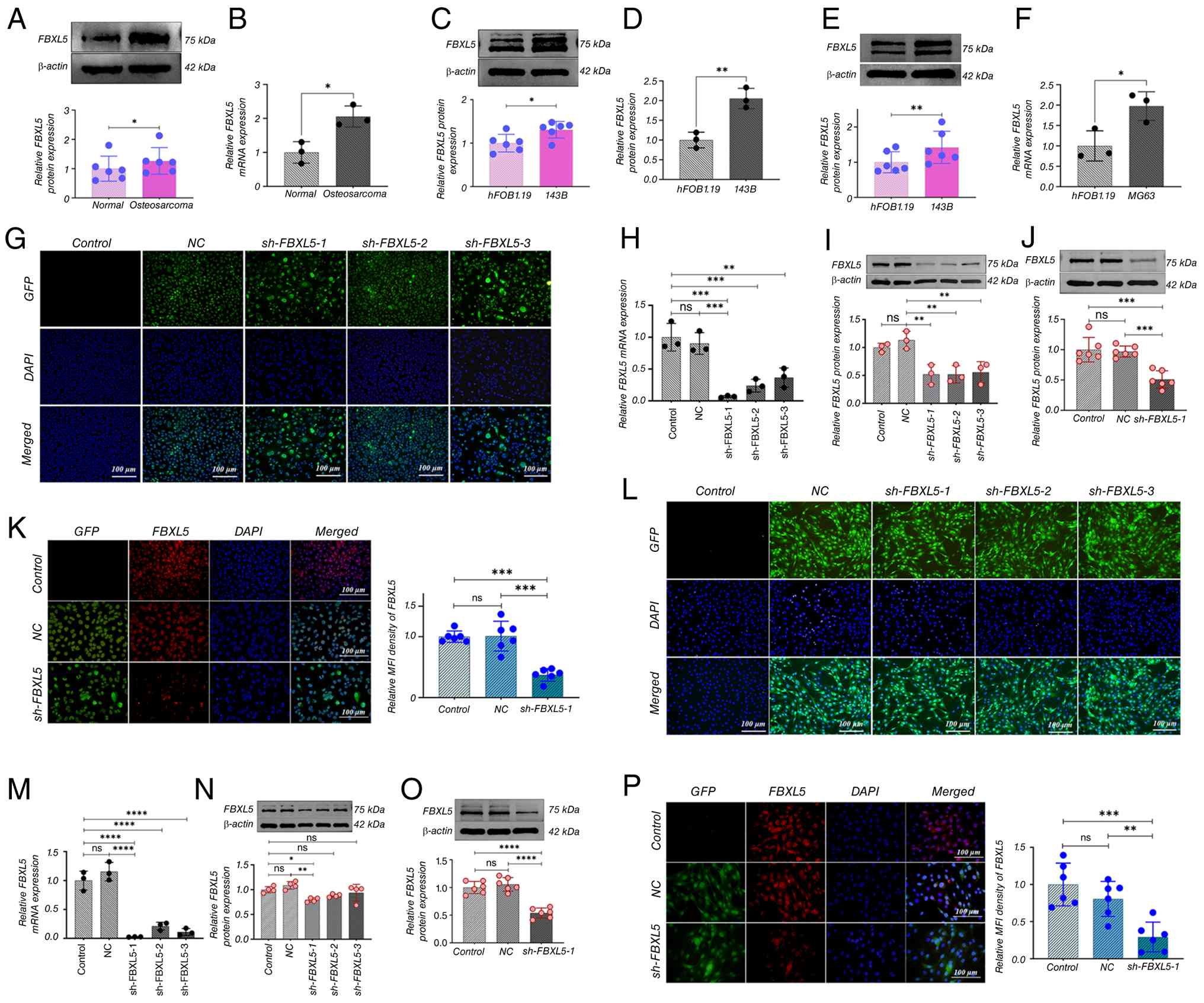 | Figure 1.FBXL5 expression in OS tissues and
cells and establishment of stable knockdown cell lines. (A) Western
blot analysis of FBXL5 protein expression in OS cell lines;
statistical analyses shown. (B) qPCR measurement of FBXL5 mRNA
expression levels (n=3). (C) Western blot of FBXL5 in 143B cells.
(D) qPCR of FBXL5 mRNA in 143B cells. (E) Western blot of FBXL5 in
MG63 cells. (F) qPCR of FBXL5 mRNA in MG63 cells (n=3). (G)
Lentiviral infection efficiency in 143B cells. (H) FBXL5 mRNA
levels in 143B knockdown groups (n=3). (I) FBXL5 protein expression
in 143B groups. (J) FBXL5 protein quantification in 143B groups.
(K) Immunofluorescence staining of FBXL5 in 143B cells (scale bar,
100 µm). (L) Lentiviral infection, (M) mRNA, (N) protein expression
and (O) quantification, and (P) immunofluorescence in MG63 cells
(scale bar, 100 µm). n=3; *P<0.05, **P<0.01, ***P<0.001,
****P<0.0001. FBXL5, F-box and leucine-rich repeat protein 5;
OS, osteosarcoma; WB, Western blot; qPCR, quantitative PCR; sh-,
short hairpin; NC, negative control; GFP, green fluorescent
protein. |
Lentivirus-mediated RNA interference successfully
established stable FBXL5 knockdown cell lines, with infection rates
exceeding 99% in 143B (Fig. 1G) and
MG63 (Fig. 1L) cells. qPCR results
showed significantly lower FBXL5 mRNA levels in the sh-FBXL5 group
compared with the control and NC groups in both 143B (Fig. 1H) and MG63 (Fig. 1M) cells. WB analysis further
confirmed a marked reduction in FBXL5 protein expression in the
sh-FBXL5-1 group of 143B (Fig. 1I and
J) and MG63 (Fig. 1N and O)
cells compared with the control group. Immunofluorescence analysis
also demonstrated significantly decreased FBXL5 expression in the
sh-FBXL5-1 group in both 143B (Fig.
1K) and MG63 (Fig. 1P) cells
compared with the control and NC groups.
These results demonstrate that FBXL5 is highly
expressed in OS tissues and cell lines, providing experimental
support for investigating its functional role in OS.
Lentivirus-mediated RNA interference successfully knocked down
FBXL5 expression, with the sh-FBXL5-1 sequence reducing both mRNA
and protein expression levels. Therefore, subsequent experiments
were conducted using the sh-FBXL5-1 lentivirus.
Knockdown of FBXL5 inhibits the
proliferation of 143B and MG63 cells
WB analysis and immunofluorescence were used to
detect changes in expression of the proliferation-related proteins
PCNA, CDK4 and cyclin D1 after FBXL5 knockdown, with EdU and CCK-8
assays assessing the impact on cell proliferation.
WB analysis showed a significant downregulation of
PCNA, CDK4 and cyclin D1 protein expression levels in the sh-FBXL5
group compared with the control and NC groups in 143B (Fig. 2A-C) and MG63 (Fig. 2I-K) cells. Immunofluorescence
analysis further confirmed a significant reduction in the mean
fluorescence intensity of these proteins in the sh-FBXL5 group in
both 143B (Fig. 2D-F) and MG63
(Fig. 2L-N) cells.
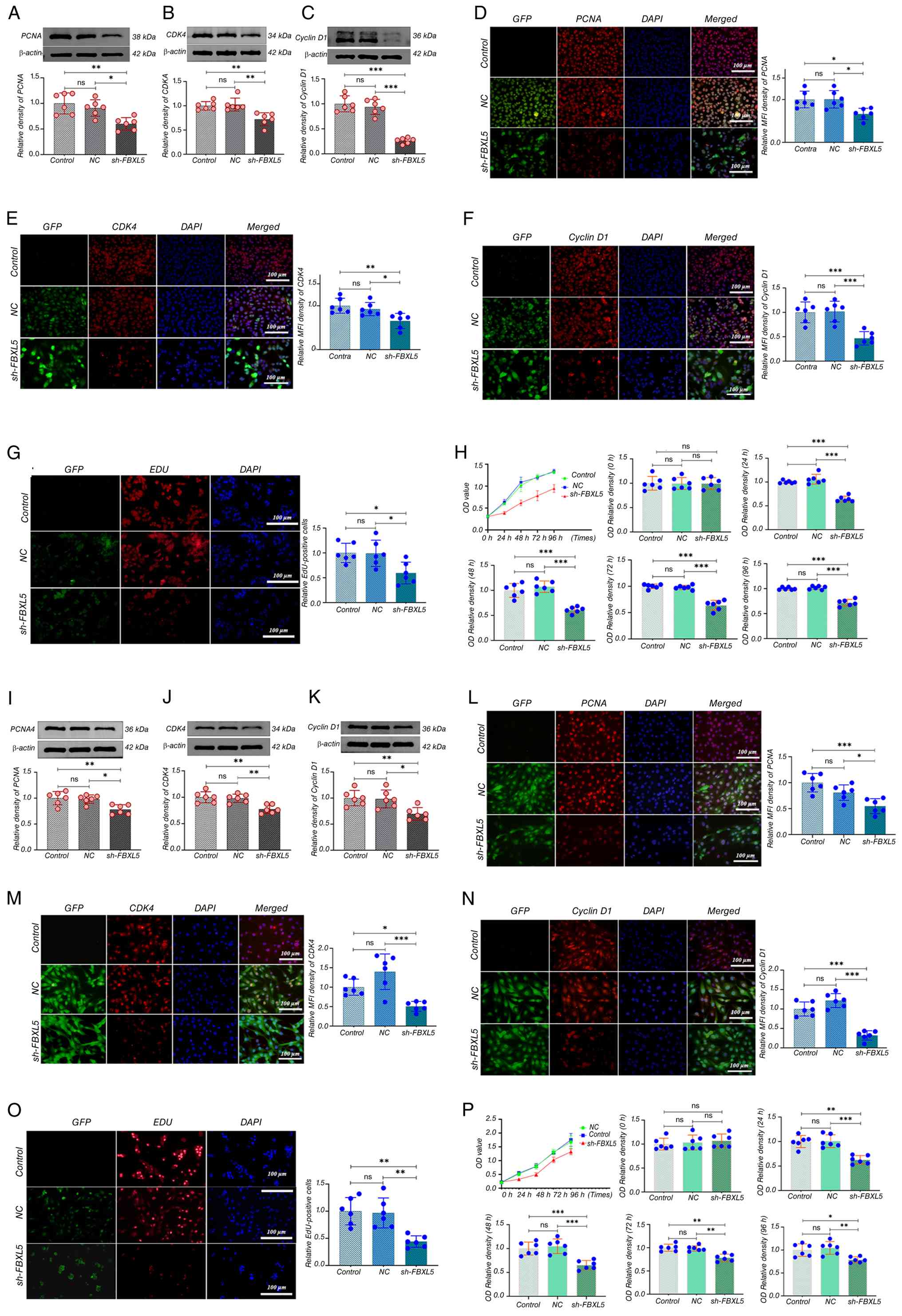 | Figure 2.FBXL5 knockdown inhibits OS cell
proliferation and downregulates cell cycle-related proteins. (A) WB
analysis of PCNA protein expression in 143B cells after FBXL5
knockdown. (B) CDK4 expression by WB in 143B cells. (C) Cyclin D1
expression by WB in 143B cells. Immunofluorescence staining (red)
of (D) PCNA, (E) CDK4 and (F) cyclin D1 in 143B cells; quantified
using MFI (scale bar, 100 µm). (G) EdU assay in 143B cells showing
proliferation; EdU-positive cells in red; quantification shown on
the right. (H) CCK-8 cell proliferation assay in 143B cells with
quantification of cell viability at different time points. WB
analysis of (I) PCNA, (J) CDK4 and (K) cyclin D1 in MG63 cells
after FBXL5 knockdown. Immunofluorescence staining (red) of (L)
PCNA, (M) CDK4 and (N) cyclin D1 in MG63 cells; quantified by MFI
(scale bar, 100 µm). (O) EdU assay showing proliferation in MG63
cells. (P) CCK-8 proliferation curve with quantification in MG63
cells. n=6; *P<0.05, **P<0.01, ***P<0.001. MFI, mean
fluorescence intensity; PCNA, proliferating cell nuclear antigen;
CDK4, cyclin-dependent kinase 4; GFP, green fluorescent protein;
NC, negative control; FBXL5, F-box and leucine-rich repeat protein
5; OS, osteosarcoma; OD, optical density; sh-, short hairpin; WB,
western blot; CCK-8, Cell Counting Kit-8. |
The EdU assay revealed a significant decrease in the
number and proportion of EdU-positive cells in the sh-FBXL5 group
in 143B (Fig. 2G) and MG63
(Fig. 2O) cells compared to the
control and NC groups.
CCK-8 proliferation assays showed that sh-FBXL5
significantly inhibited the proliferation of 143B (Fig. 2H) and MG63 (Fig. 2P) cells, with notably lower optical
density values compared to controls starting at 24 h; no
significant differences were observed between the control and NC
groups. These findings suggest that FBXL5 serves a key role in
promoting OS cell proliferation, and its depletion suppresses tumor
cell growth through downregulation of PCNA, CDK4 and cyclin D1.
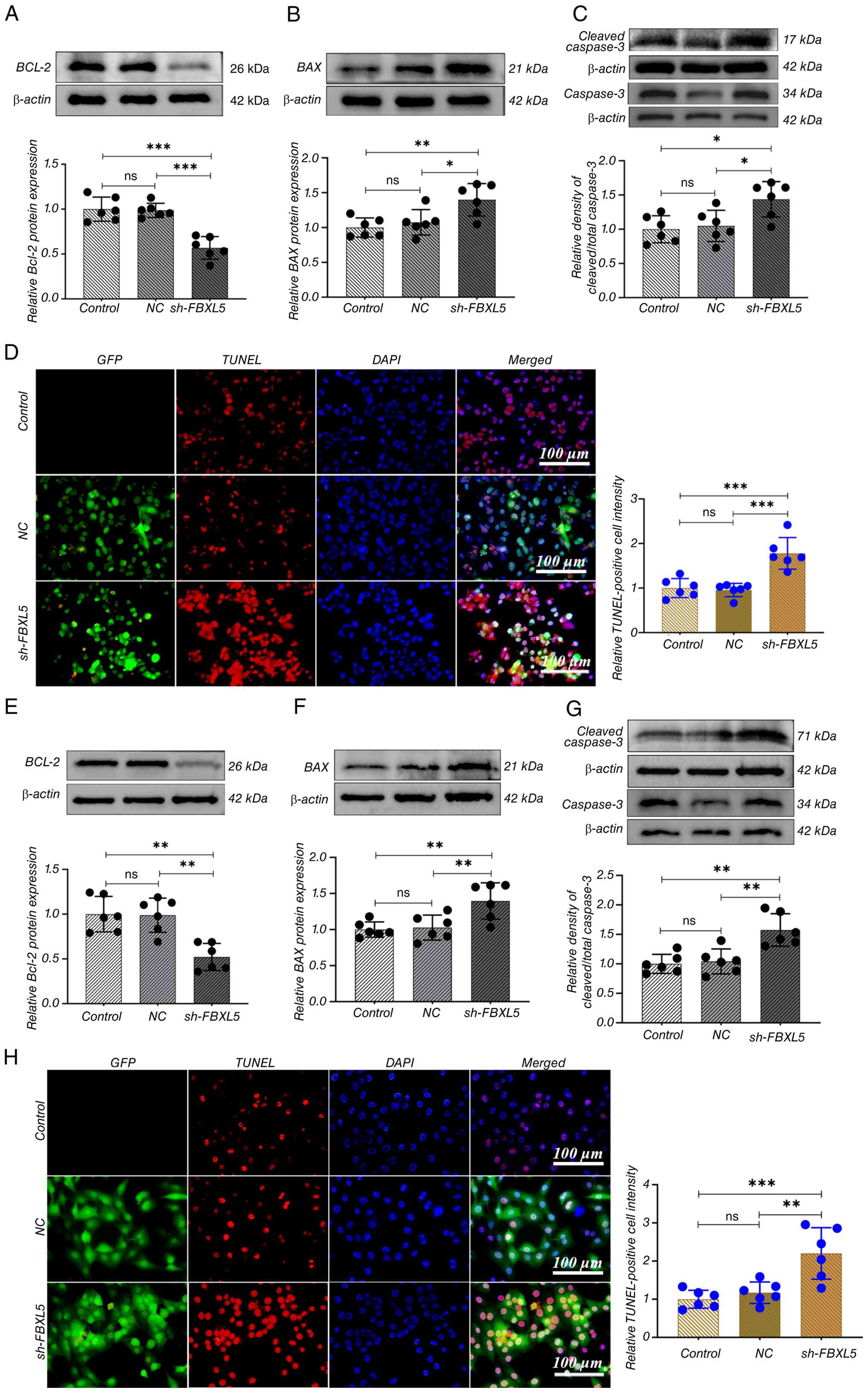
FBXL5 knockdown promotes apoptosis in
143B and MG63 cells
WB analysis was used to analyze the expression
changes of apoptosis-related proteins following FBXL5 knockdown,
while the TUNEL assay assessed cell apoptosis.
The results showed that in the sh-FBXL5 group,
expression of the anti-apoptotic protein Bcl-2 was significantly
reduced, while the pro-apoptotic proteins BAX and caspase-3 were
significantly increased in both 143B (Fig. 3A-C) and MG63 (Fig. 3E-G) cells. TUNEL staining revealed
nuclear fragmentation and morphological abnormalities in the
sh-FBXL5 group, with intense red fluorescence signals (Fig. 3D and H). The proportion of
TUNEL-positive cells was significantly higher in the sh-FBXL5 group
compared with the control and NC groups, whereas there was no
statistical difference observed between the control and NC
groups.
This indicates that FBXL5 knockdown induces
apoptosis in OS cells, suggesting that FBXL5 may function as an
anti-apoptotic factor in osteosarcoma by upregulating Bcl-2 and
downregulating BAX and caspase-3.
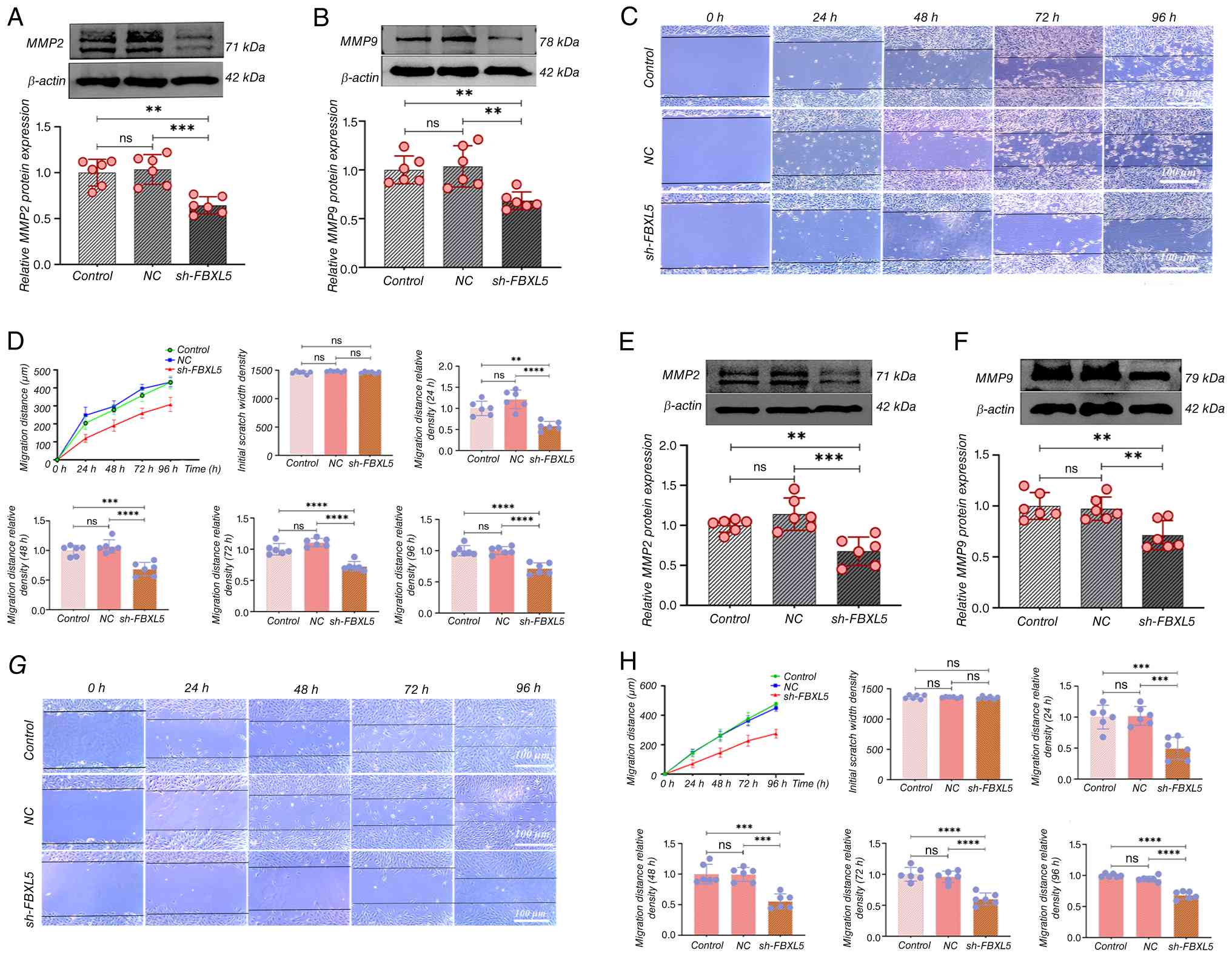
Knockdown of FBXL5 inhibits the
migration and invasion abilities of 143B and MG63 cells
WB analysis was performed to detect changes in the
expression of migration and invasion markers MMP2 and MMP9
following FBXL5 knockdown, and a wound healing assay was used to
evaluate cell migration ability.
The results showed that MMP2 and MMP9 protein
expression were significantly reduced in both 143B (Fig. 4A and B) and MG63 (Fig. 4E and F) cells in the sh-FBXL5 group
compared with both the control and NC groups. The wound healing
assay demonstrated that 143B (Fig.
4C) and MG63 (Fig. 4G) cells in
the control and NC groups migrated markedly more quickly when
compared with the sh-FBXL5 group; wound healing was significantly
delayed at all time points in the sh-FBXL5 group, with incomplete
wound closure at 96 h. The migration distance curve revealed that
the migration distance of 143B (Fig.
4D) and MG63 (Fig. 4H) cells in
the sh-FBXL5 group was significantly lower than that of the control
group at all time points from 24 to 96 h.
These results imply that FBXL5 is involved in
regulating OS cell migration and invasion, likely through the
downregulation of MMP2 and MMP9, and targeting FBXL5 may limit
metastatic potential.
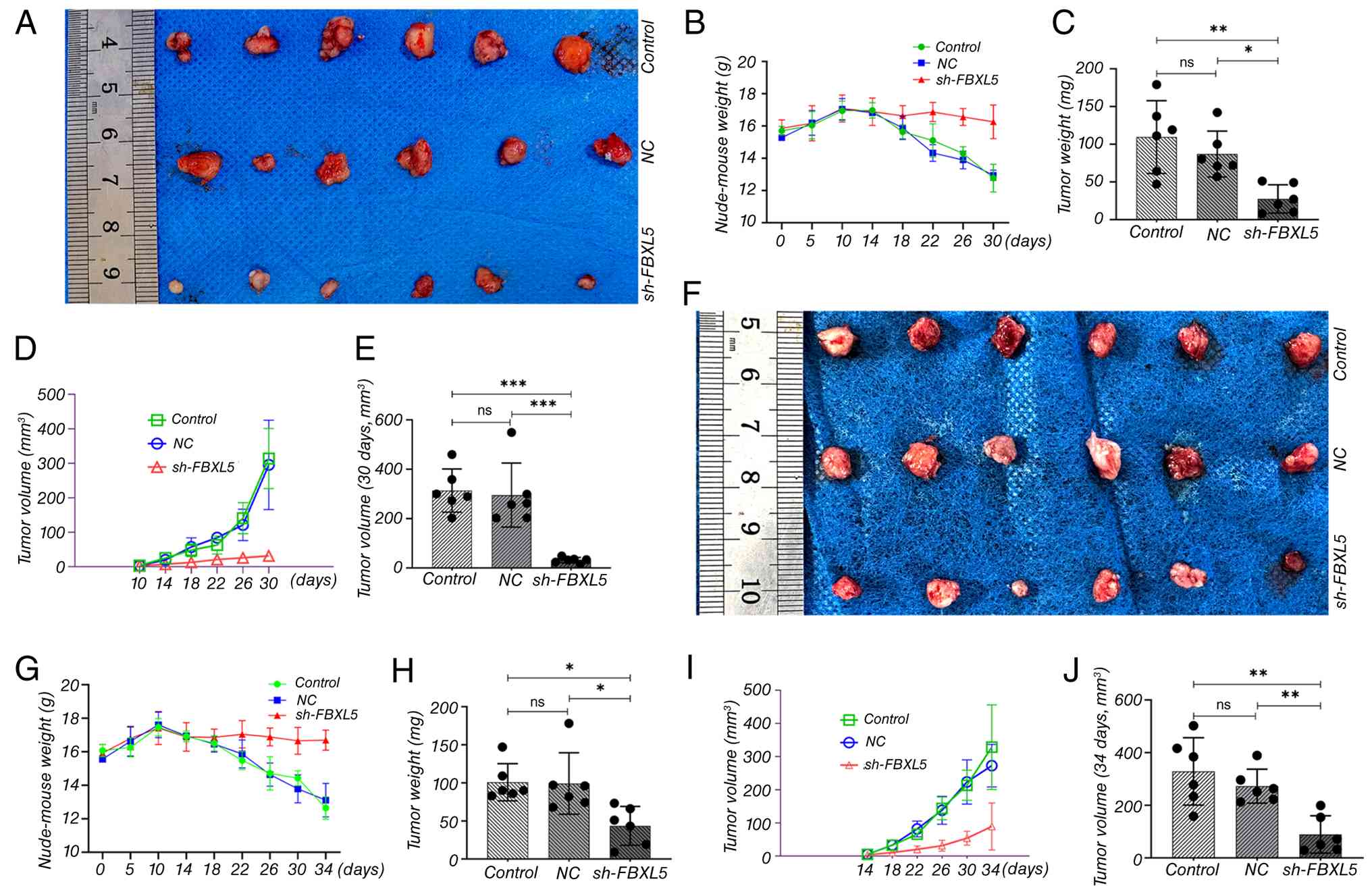
Subcutaneous tumor xenograft model in
nude mice
A subcutaneous tumor implantation model in nude mice
was used to systematically assess the impact of FBXL5 knockdown on
tumor growth, thereby investigating the role of FBXL5 in OS
progression.
FBXL5 knockdown inhibits tumor
growth
i) Body weight changes. In the sh-FBXL5 group, the
body weight of nude mice in 143B (Fig.
5B) and MG63 (Fig. 5G)
xenograft models remained stable, with a slight increase. By
contrast, mice in the control and NC groups showed a slight weight
increase during the first 10 days, followed by a continuous decline
from day 14 onward.
ii) Tumor volume. Tumor formation was observed in
all groups by day 10 post-implantation. Tumor volumes of 143B
(Fig. 5D) and MG63 (Fig. 5I) in the sh-FBXL5 group were
significantly smaller than those in the control group, with a
marked reduction in growth rate at each time point.
iii) Tumor weight and morphology. At the study
endpoint, gross tumor images showed that tumors in both the control
and NC groups exhibited tight adhesion to the ribcage, whereas the
sh-FBXL5 group displayed well-encapsulated tumors with no apparent
invasion in both the 143B and MG63 ×enograft models (Fig. 5A and F). Tumor weights and volumes
of 143B (Fig. 5C and E) and MG63
(Fig. 5H and J) in the sh-FBXL5
group were significantly lower than those in the control group.
Histological analysis of transplanted
tumors
Immunofluorescence analysis confirmed that in both
the 143B (Fig. 6A and B) and MG63
(Fig. 6D and E) tumor models, FBXL5
expression was significantly reduced in the sh-FBXL5 group compared
to the control and NC groups, confirming that lentivirus-mediated
FBXL5 knockdown remained stable in vivo.
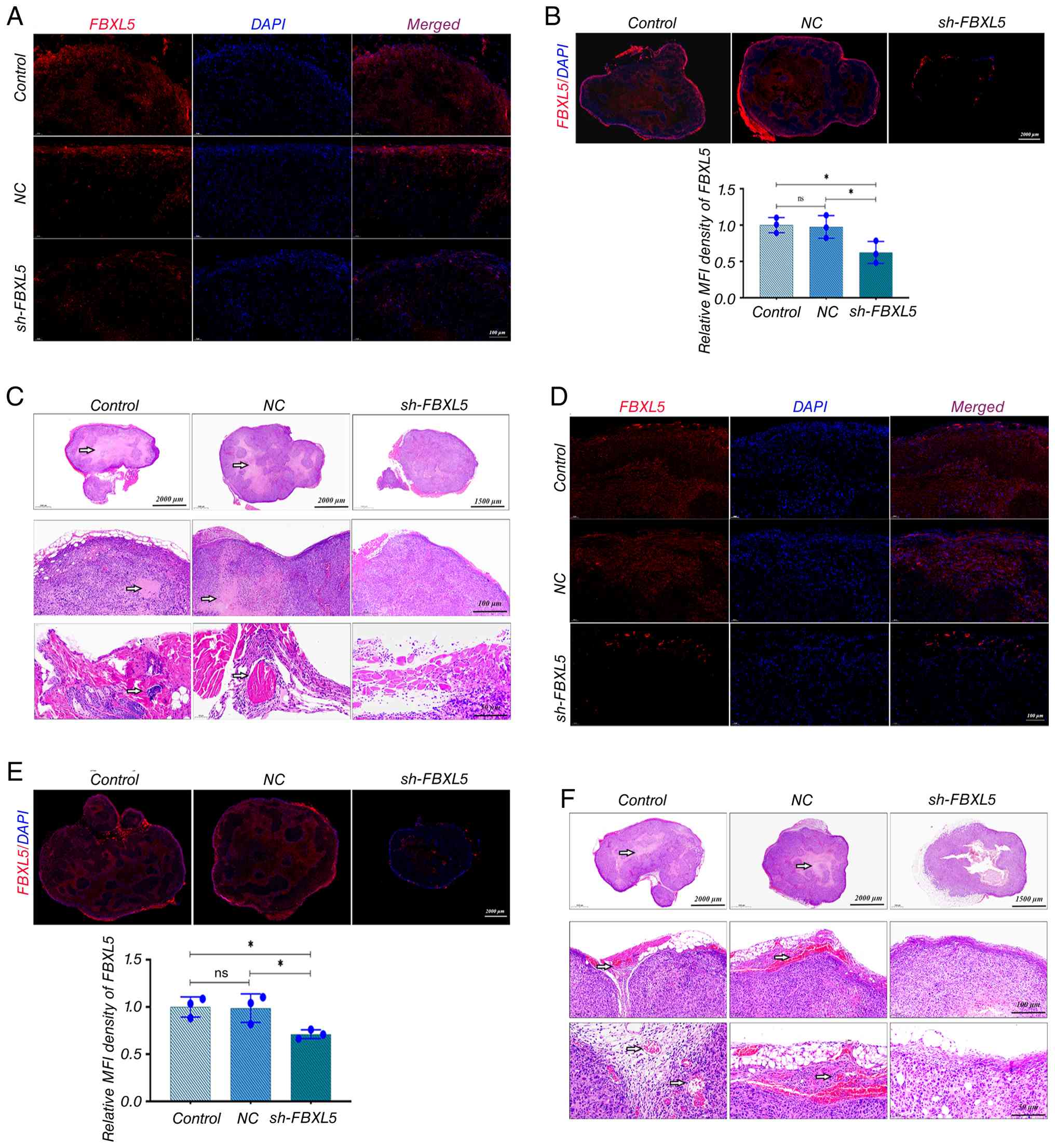 | Figure 6.In vivo stability verification
of FBXL5 knockdown and H&E staining of xenograft tumors. (A) IF
staining of FBXL5 protein in tumor tissues from each group in the
143B nude mouse model. (B) Quantification of FBXL5 fluorescence
intensity in 143B tumors; y-axis represents relative MFI density of
FBXL5. (C) H&E staining of 143B tumor tissues from each group;
top row, low magnification; middle row, medium magnification;
bottom row, high magnification. (D) IF staining of FBXL5 protein in
tumor tissues from each group in the MG63 nude mouse model. (E)
Quantification of FBXL5 fluorescence intensity in MG63 tumors;
y-axis represents relative MFI density of FBXL5. (F) H&E
staining of MG63 tumor tissues from each group; top row, low
magnification; middle row, medium magnification; bottom row, high
magnification. Arrows indicate cells with high FBXL5 expression.
n=6; *P<0.05. FBXL5, F-box and leucine-rich repeat protein 5;
HE, hematoxylin and eosin; NC, negative control; sh-, short
hairpin; IF, immunofluorescence. |
Hematoxylin and eosin staining showed that 143B
(Fig. 6C) and MG63 (Fig. 6F) tumors in the control and NC
groups exhibited characteristic malignant features, including
disorganized cell arrangement, nest-like or diffuse distribution,
variable morphology, uneven nuclear staining, frequent mitotic
figures, marked atypia, unclear tissue boundaries, and extensive
areas of necrosis and neovascularization. By contrast, tumors in
the sh-FBXL5 group displayed more organized cell arrangement,
relatively uniform morphology, even nuclear staining, fewer mitotic
figures, reduced atypia, clear tissue boundaries, and significantly
less necrosis and neovascularization.
Histological analysis confirmed that FBXL5 knockdown
reduces malignant features of OS tumors in vivo, including
disorganized cell arrangement, necrosis and neovascularization,
further supporting its tumor-suppressive role.
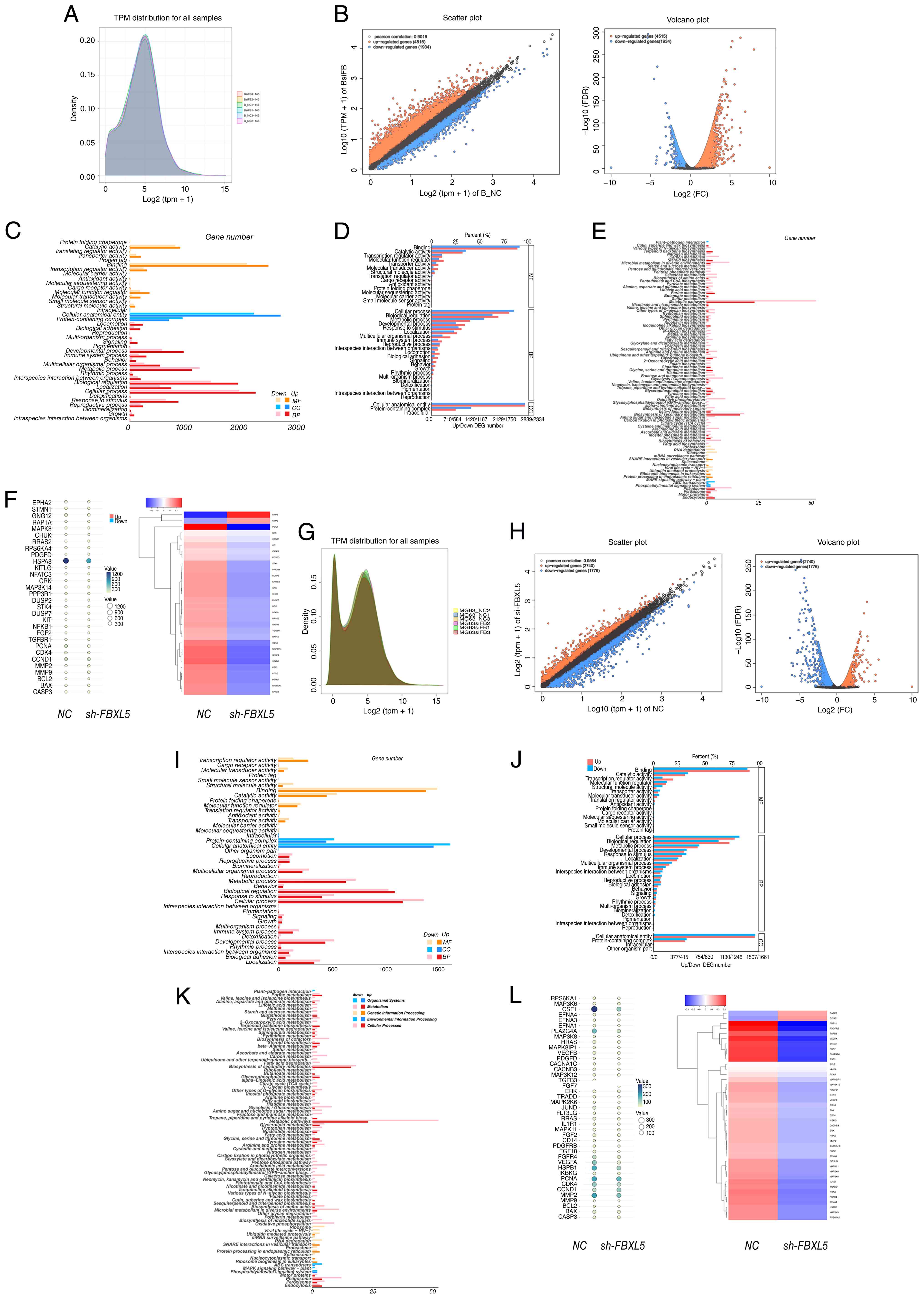
Transcriptome sequencing analysis to
explore the molecular mechanisms by which FBXL5 knockdown inhibits
OS progression
RNA-seq was employed to analyze the transcriptomic
changes in 143B and MG63 cell lines following FBXL5 knockdown to
further investigate the molecular mechanisms through which FBXL5
knockdown inhibits OS progression.
Gene expression distribution and
differential analysis
Following transcripts per million normalization, the
gene expression distributions in 143B (Fig. 7A) and MG63 (Fig. 7G) cells exhibited normal
distributions with high overlap, indicating high data quality.
Volcano plot analysis revealed that, following FBXL5 knockdown,
3,039 genes were upregulated and 2,472 genes were downregulated in
143B cells (Fig. 7B), while in MG63
cells, 2,740 genes were upregulated and 1,776 genes were
downregulated (Fig. 7H) (P<0.05,
|log2FC|>1). These GO enrichment results indicate
that FBXL5 primarily affects metabolic and proliferation-related
biological processes in OS cells.
GO enrichment analysis
GO enrichment analysis revealed that the
differentially expressed genes were primarily enriched in
biological processes such as ‘metabolic process’, ‘cellular
process’, ‘immune system process’ and ‘growth’. In the cellular
component category, the genes were mainly associated with ‘protein
complex’ and ‘intracellular anatomical structure’, while in the
molecular function category, they were enriched in ‘catalytic
activity’ and ‘binding’ (Fig. 7C and
I). Enrichment ratio analysis indicated that metabolism- and
proliferation-related pathways were the most prominent (Fig. 7D and J), suggesting that FBXL5
regulates OS progression by modulating these pathways.
KEGG enrichment analysis
KEGG enrichment analysis revealed that the
differentially expressed genes were significantly enriched in the
‘MAPK signaling pathway-plant’ (Fig. 7E
and K). In the sh-FBXL5 cells, the expression levels of
proliferation-related genes (PCNA, cyclin D1 and CDK4) were
significantly downregulated, and alterations in the expression of
MAPK pathway members [MAPK8, dual specificity phosphatase 2
(DUSP2)] were observed. Notably, the downregulation of DUSP2 may be
associated with altered ERK activity and suppressed proliferative
signaling; however, the direct regulatory relationship requires
further investigation (Fig. 7F and
L).
KEGG pathway analysis identified the MAPK signaling
pathway as significantly enriched following FBXL5 knockdown,
highlighting this pathway as a key mediator of FBXL5′s oncogenic
functions in OS (Fig. 7E and
K).
MEK/ERK signaling pathway
validation
WB analysis showed that after FBXL5 knockdown, the
expression levels of Ras, Raf, MEK1/2 and ERK1/2 proteins were
significantly reduced in 143B (Fig.
8A-D) and MG63 (Fig. 8I-L)
cells. Immunofluorescence further confirmed that the expression of
MEK1/2 and ERK1/2 was markedly decreased in 143B (Fig. 8E and F) and MG63 (Fig. 8M and N) cells. Immunohistochemistry
of xenograft tumors revealed that Raf and MEK1/2 expression levels
were significantly reduced in the sh-FBXL5 group in both 143B
(Fig. 8G and H) and MG63 (Fig. 8O and P) models.
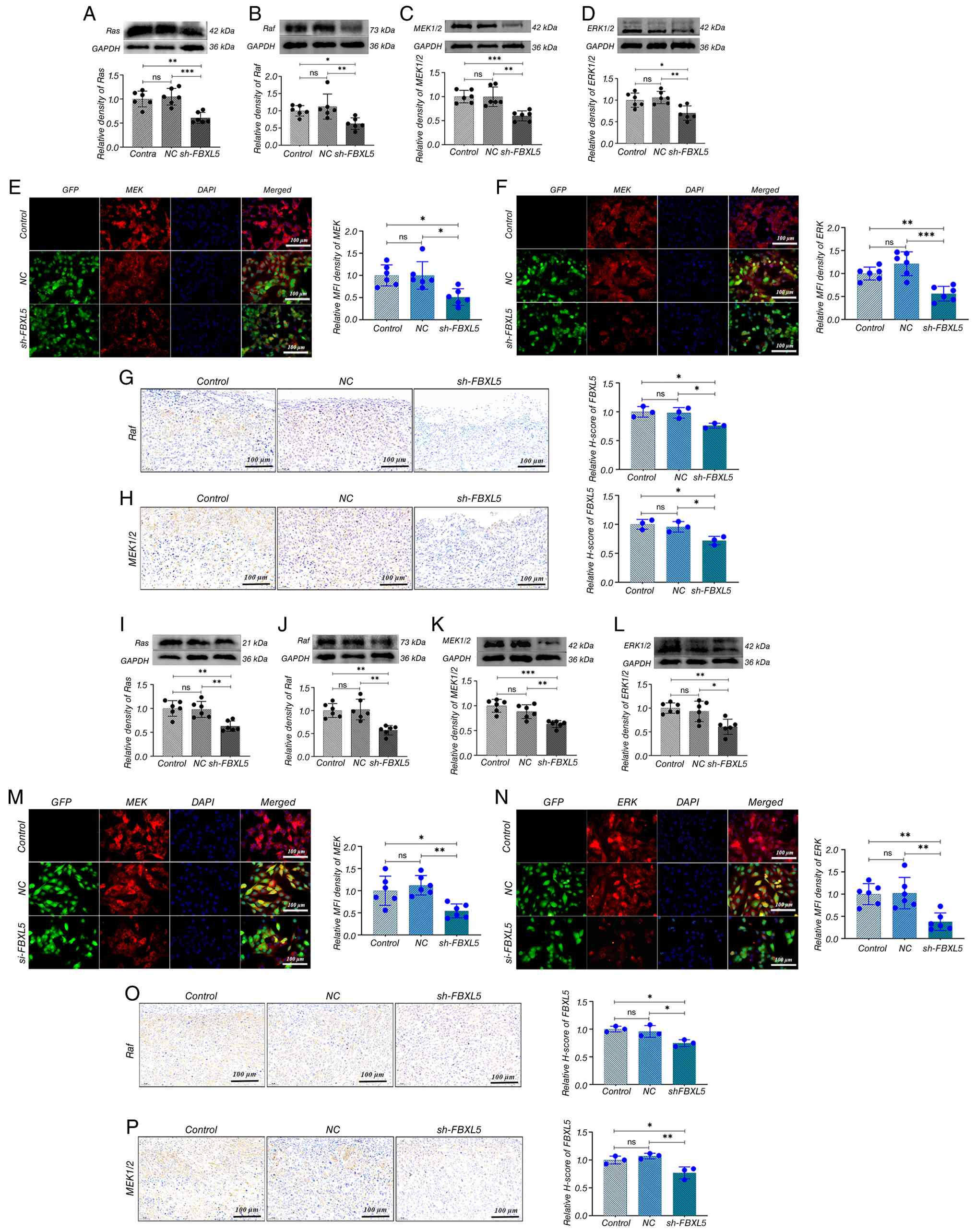 | Figure 8.FBXL5 knockdown suppresses the
Ras/Raf/MEK/ERK signaling cascade in OS cells. Immunoblot analysis
and quantification of (A) Ras, (B) Raf, (C) MEK1/2 and (D) ERK1/2
proteins in 143B cells; y-axis represents relative density. IF
staining of (E) MEK1/2 and (F) ERK1/2 in 143B cells (green,
GFP-positive transfected cells; red, target protein; blue,
DAPI-stained nuclei; scale bar, 100 µm). IHC staining and
semi-quantitative analysis of (G) Raf and (H) MEK1/2 in 143B
xenograft tumors (scale bar, 100 µm). Immunoblot analysis and
quantification of (I) Ras, (J) Raf, (K) MEK1/2 and (L) ERK1/2 in
MG63 cells; y-axis represents relative density. IF staining of (M)
MEK1/2 and (N) ERK1/2 in MG63 cells (scale bar, 100 µm). IHC
staining of (O) Raf and (P) MEK1/2 in MG63 ×enograft tumors (scale
bar, 100 µm). n=6; *P<0.05, **P<0.01, ***P<0.001. OS,
osteosarcoma; GFP, green fluorescent protein; NC, negative control;
sh-, short hairpin; FBXL5, F-box and leucine-rich repeat protein 5;
ns, not significant; IF, immunofluorescence; IHC,
immunohistochemical. |
Collectively, these data demonstrate that FBXL5
knockdown suppresses OS progression by inhibiting the MAPK/ERK
signaling pathway at multiple levels (Ras, Raf, MEK and ERK),
providing a mechanistic basis for its tumor-suppressive
effects.
Discussion
Surgical resection combined with chemotherapy
remains the standard treatment for OS, but metastasis, drug
resistance and poor prognosis continue to present significant
clinical challenges (19–21). The 5-year survival rate for
localized OS is ~60%, while it remains under 20% for metastatic
cases (22,23). Current chemotherapy regimens offer
limited efficacy, and although targeted therapies are recommended
by the National Comprehensive Cancer Network, they have not
substantially improved the overall survival rate of patients
(24,25). Thus, identifying novel molecular
mechanisms and therapeutic targets is essential. The present study
investigated the role of the E3 ubiquitin ligase FBXL5 in OS,
specifically focusing on its signaling pathways and regulatory
effects on OS cancer cell behavior.
As an E3 ubiquitin ligase that regulates iron
homeostasis, FBXL5 serves a complex role in various cancers,
functioning as either an oncogene or tumor suppressor (26). In gastric cancer, overexpression of
FBXL5 significantly reduces metastatic potential, while its
knockdown enhances Snail1 stability, promoting cell migration
(27–30). By contrast, the present study
revealed that FBXL5 expression was significantly elevated in OS. In
a lung cancer model, FBXL5 knockout induced iron accumulation,
which inhibited p27^Kip1 degradation and caused G1/S-phase cell
cycle arrest, ultimately suppressing tumor growth (10,31).
Clinical studies indicate that ~19% of patients with hepatocellular
carcinoma show low FBXL5 expression, which correlates with a poor
prognosis (12,32). In advanced-stage ccRCC, ~30% of
patients exhibit reduced FBXL5 expression, which is significantly
associated with poorer survival (hazard ratio ~2.0) (33). These findings highlight the variable
expression of FBXL5 between normal and tumor tissues across
malignancies, suggesting its potential as a prognostic biomarker
for various cancers, including OS.
FBXL5 functions at the intersection of iron
metabolism and cellular signaling pathways (34). In gastric cancer, FBXL5 inhibits the
EMT by mediating the ubiquitination and degradation of Snail1 and
cortactin, while also reducing the stability of human
single-stranded DNA-binding protein 1 (27,28,35).
In OS, FBXL5 predominantly regulates cell cycle progression and
survival. The present study demonstrated that FBXL5 knockdown
suppressed cell proliferation by downregulating the expression of
PCNA, cyclin D1 and CDK4. FBXL5 deficiency increases IRP2 activity,
disrupts iron homeostasis and causes reactive oxygen species (ROS)
accumulation, stabilizing p27 and inducing G1-phase arrest
(12,31,36).
Furthermore, FBXL5 knockdown reduced the expression of MMP2 and
MMP9, significantly impairing cell migration. Consequently, FBXL5
can act as an oncogene or tumor suppressor, depending on the
specific molecular pathways it regulates in different cancers.
The present study found that FBXL5 knockdown was
associated with a significant reduction in MAPK/ERK signaling
activity in OS models. RNA-seq and experimental validation revealed
that FBXL5 silencing led to a marked decrease in the
phosphorylation levels of ERK1/2 (37,38).
Although FBXL5 may modulate negative regulators of the MAPK
pathway, such as sprouty RTK signaling antagonist 2 or DUSP family
members, this proposed mechanism is speculative and requires direct
experimental validation (39). The
disruption of iron homeostasis and increased ROS levels induced by
FBXL5 knockdown may further suppress ERK pathway activity by
activating stress pathways such as p38/c-Jun N-terminal kinase
(38). While the present study does
not explicitly confirm that p21 influences the ERK signaling in OS
cells through this mechanism, further research is warranted to
clarify the specific role of FBXL5 in regulating MAPK
signaling.
Research suggests that FBXL5 modulates ferroptosis
and other signaling pathways, such as iron homeostasis and
ROS-mediated signaling (40,41).
As an iron-dependent form of programmed cell death characterized by
lipid peroxidation, ferroptosis represents a potential therapeutic
target for malignant tumors such as OS (42). High FBXL5 expression limits iron and
ROS accumulation, inhibiting ferroptosis. Conversely, inducing
ferroptosis has been shown to effectively eradicate drug-resistant
cells and OS stem cells (42–44).
Additionally, FBXL5 may modulate the crosstalk between EMT and
nuclear factor κB (NF-κB) signaling by inducing Snail1 degradation
(28,45,46) or
regulating β-transducin repeat-containing protein to influence
NF-kB activity (47).
Targeted strategies involving FBXL5 in OS may
leverage insights gained from successful mouse double minute 2-p53
inhibitor therapies (48,49). One potential approach is to simulate
a pseudo-iron deficiency by utilizing iron chelators to activate
FBXL5′s iron-responsive domain, triggering its self-ubiquitination
and degradation (36). This mimics
the effects of FBXL5 knockdown, thereby inhibiting tumor cell
proliferation or inducing ferroptosis (40,50).
Iron chelators such as deferoxamine are established clinical
treatments for iron overload diseases (51,52),
and are currently being investigated as adjunctive therapies for
cancer (53). Similarly,
ferroptosis-inducing agents such as RSL3 and erastin can be
combined with chemotherapy (42,54).
FBXL5 expression levels may serve as a predictor of therapeutic
responses, with higher expression potentially indicating increased
resistance to chemo- and radiotherapy. Modulating the FBXL5-iron
homeostasis axis holds promise for enhancing radiosensitivity
(55–57).
Overall, FBXL5 serves a multifaceted role in the
progression of OS by promoting cell proliferation and migration
through both direct substrate degradation and indirect modulation
of iron homeostasis and ROS levels. The present study reveals a
previously unrecognized link between FBXL5 and the MAPK/ERK
signaling pathway, broadening the understanding of its function and
suggesting novel therapeutic targets. To more fully understand the
molecular mechanisms by which FBXL5 regulates OS progression,
future research should involve screening interacting proteins via
co-immunoprecipitation mass spectrometry and performing gene rescue
experiments.
In conclusion, the present study suggests that FBXL5
promotes tumor growth in OS by activating the MAPK/ERK signaling
pathway. The results indicated that FBXL5 promoted tumor cell
proliferation and migration by maintaining iron homeostasis and
activating the MAPK/ERK signaling pathway. This tissue-specific
function contradicts the anti-cancer role observed in epithelial
tumors. The reveal that FBXL5 modulates the MAPK/ERK pathway not
only broadens the functional knowledge of this protein but also
offers a novel potential therapeutic avenue for treating OS.
Leveraging the iron-responsive properties of FBXL5, iron
chelator-induced degradation is a promising treatment strategy.
Future studies should validate FBXL5 expression in larger cohorts
to confirm its prognostic and therapeutic value in OS. Furthermore,
an investigation into combining FBXL5 inhibition with ferroptosis
inducers or MEK inhibitors is warranted. This study provides key
evidence for understanding the molecular mechanisms of OS and
developing new therapeutic strategies.
The present study demonstrated that FBXL5 knockdown
inhibited the progression of OS through multiple mechanisms. In
vitro experiments confirmed that FBXL5 knockdown suppressed
cell proliferation, induced apoptosis and reduced migration and
invasion. In the xenograft model, FBXL5 knockdown significantly
inhibited tumor growth, reduced tumor volume and weight, and
improved the physiological condition of tumor-bearing mice.
Histopathological analysis showed that FBXL5 knockdown reduced
tumor malignancy, as evidenced by decreased cellular atypia, clear
tissue boundaries and reduced necrosis and neovascularization. The
in vitro and in vivo data consistently indicate that
FBXL5 serves a functional role in regulating the malignant
biological behaviors of OS.
Limitations of the present study include limited
clinical sample size (n=6), warranting validation in larger
cohorts. The exact molecular mechanism by which FBXL5 regulates the
MAPK/ERK pathway remains incompletely understood. In vivo
experiments were limited to subcutaneous xenograft models, which
may not fully represent OS metastasis. Additionally, shRNA-mediated
knockdown was the primary method used; alternative approaches are
needed to confirm the results.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science
Foundation of China (grant no. 82360435), the Lanzhou University
Second Hospital Cuiying Youth Fund Project (grant no.
CY2022-QN-A03), the Cuiying Science and Technology Innovation
Program Project (grant no. 2022-MS-A10), the Lanzhou Youth Science
and Technology Talents Innovation Project (grant no. 2023-2-39) and
the Natural Science Foundation of Gansu Province (grant no.
24JRRA1101).
Availability of data and materials
The RNA sequencing data generated in this study have
been deposited in the NCBI BioProject database under BioProject
accession number PRJNA1440046 and are publicly available at the
following URL: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1440046.
All other data supporting the findings of the present study are
available from the corresponding author upon reasonable
request.
Author's contributions
YCM and WMZ contributed equally to this work as
first authors. YCM, WMZ and WN conceived and designed the study.
YCM, WMZ, YQS and YBD performed the experiments, including cell
culture, lentiviral transduction, qPCR, western blotting,
immunofluorescence and animal studies. YCM, WMZ and HHZ analyzed
and interpreted the data, including RNA-seq bioinformatics
analysis. YCM and WMZ drafted the manuscript. HHZ and WN critically
revised the manuscript for important intellectual content. WN and
HHZ are co-corresponding authors. All authors read and approved the
final manuscript, and agree to be accountable for all aspects of
the work, ensuring that questions related to the accuracy or
integrity of any part are appropriately investigated and resolved.
YCM and WMZ confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
This study was conducted in accordance with the
Declaration of Helsinki and approved by the Ethics Committee of the
Second Hospital of Lanzhou University. Human tissue samples
collection was approved under ethics approval no. 2024A-504.
Written informed consent was obtained from all adult patients or
from the parents/legal guardians of patients under 18 years of age
prior to sample collection and participation in the study. All
animal experiments were approved by the Institutional Animal Care
and Use Committee of Lanzhou University (approval no. D2024-417)
and performed in accordance with the Guide for the Care and Use of
Laboratory Animals.
Patient consent for publication
Written informed consent for publication of clinical
details and clinical images was obtained from all patients or their
parents/legal guardians for patients under 18 years of age. All
patient information has been de-identified to protect patient
privacy.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Song K, Song J, Lin K, Chen F, Ma X, Jiang
J and Li F: Survival analysis of patients with metastatic
osteosarcoma: A surveillance, epidemiology, and end results
population-based study. Int Orthop. 43:1983–1991. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mettmann VL, Blattmann C, Friedel G,
Harrabi S, von Kalle T, Kager L, Kevric M, Kühne T, Nathrath M,
Sorg B, et al: Primary Multi-Systemic metastases in osteosarcoma:
Presentation, treatment, and survival of 83 patients of the
cooperative osteosarcoma study group. Cancers. 16:2752024.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mao P, Feng Z, Liu Y, Zhang K, Zhao G, Lei
Z, Di T and Zhang H: The role of ubiquitination in osteosarcoma
development and therapies. Biomolecules. 14:7912024. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xin S and Wei G: Prognostic factors in
osteosarcoma: A study level meta-analysis and systematic review of
current practice. J Bone Oncol. 21:1002812020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bishop MW, Janeway KA and Gorlick R:
Future directions in the treatment of osteosarcoma. Curr Opin
Pediatr. 28:26–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zoumpoulidou G, Alvarez-Mendoza C, Mancusi
C, Ahmed RM, Denman M, Steele CD, Tarabichi M, Roy E, Davies LR,
Manji J, et al: Therapeutic vulnerability to PARP1,2 inhibition in
RB1-mutant osteosarcoma. Nat Commun. 12:70642021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sayles LC, Breese MR, Koehne AL, Leung SG,
Lee AG, Liu HY, Spillinger A, Shah AT, Tanasa B, Straessler K, et
al: Genome-informed targeted therapy for osteosarcoma. Cancer
Discov. 9:46–63. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Muto Y, Nishiyama M, Nita A, Moroishi T
and Nakayama KI: Essential role of FBXL5-mediated cellular iron
homeostasis in maintenance of hematopoietic stem cells. Nat Commun.
8:161142017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang H, Shi H, Rajan M, Canarie ER, Hong
S, Simoneschi D, Pagano M, Bush MF, Stoll S, Leibold EA and Zheng
N: FBXL5 Regulates IRP2 stability in iron homeostasis via an
Oxygen-responsive (2Fe2S) cluster. Mol Cell. 78:31–41.e5. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen ZW, Liu B, Tang NW, Xu YH, Ye XY, Li
ZM, Niu XM, Shen SP, Lu S and Xu L: FBXL5-mediated degradation of
single-stranded DNA-binding protein hSSB1 controls DNA damage
response. Nucleic Acids Res. 42:11560–11569. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bian X, Yin S, Yin X, Fang T, Wang Y, Yang
S, Jiang X, Xue Y, Ye F and Zhang L: Clinical and biological
significances of FBLN5 in gastric cancer. Cancers (Basel).
15:5532023. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Muto Y, Moroishi T, Ichihara K, Nishiyama
M, Shimizu H, Eguchi H, Moriya K, Koike K, Mimori K, Mori M, et al:
Disruption of FBXL5-mediated cellular iron homeostasis promotes
liver carcinogenesis. J Exp Med. 216:950–965. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He ZJ, Li W, Chen H, Wen J, Gao YF and Liu
YJ: miR-1306-3p targets FBXL5 to promote metastasis of
hepatocellular carcinoma through suppressing snail degradation.
Biochem Biophys Res Commun. 504:820–826. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chandhanayingyong C, Kim Y, Staples JR,
Hahn C and Lee FY: MAPK/ERK signaling in osteosarcomas, Ewing
sarcomas and chondrosarcomas: Therapeutic implications and future
directions. Sarcoma. 2012:4048102012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao Z, Zhao GS, Lv Y, Peng D, Tang X, Song
H and Guo QN: Anoikis-resistant human osteosarcoma cells display
significant angiogenesis by activating the Src kinase-mediated MAPK
pathway. Oncol Rep. 41:235–245. 2019.PubMed/NCBI
|
|
16
|
Liu W, Wang B, Duan A, Shen K, Zhang Q,
Tang X, Wei Y, Tang J and Zhang S: Exosomal transfer of miR-769-5p
promotes osteosarcoma proliferation and metastasis by targeting
DUSP16. Cancer Cell Int. 21:5412021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Noh K, Kim KO, Patel NR, Staples JR,
Minematsu H, Nair K and Lee FY: Targeting inflammatory kinase as an
adjuvant treatment for osteosarcomas. J Bone Joint Surg Am.
93:723–732. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meltzer PS and Helman LJ: New horizons in
the treatment of osteosarcoma. N Engl J Med. 385:2066–2076. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tian H, Cao J, Li B, Nice EC, Mao H, Zhang
Y and Huang C: Managing the immune microenvironment of
osteosarcoma: The outlook for osteosarcoma treatment. Bone Res.
11:112023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou Y, Yang D, Yang Q, Lv X, Huang W,
Zhou Z, Wang Y, Zhang Z, Yuan T, Ding X, et al: Single-cell RNA
landscape of intratumoral heterogeneity and immunosuppressive
microenvironment in advanced osteosarcoma. Nat Commun. 11:63222020.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu Z, Wen S, Huo Z, Wang Q, Zhao J, Wang
Z, Chen Y, Zhang L, Zhou F, Guo Z, et al: Current status and
prospects of targeted therapy for osteosarcoma. Cells. 11:35072022.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu Y, Zhang J, Chen Y, Kang Y, Liao Z, He
Y and Zhang C: Novel immunotherapies for osteosarcoma. Front Oncol.
12:8305462022. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Harris MA and Hawkins CJ: Recent and
ongoing research into metastatic osteosarcoma treatments. Int J Mol
Sci. 23:38172022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Duffaud F, Mir O, Boudou-Rouquette P,
Piperno-Neumann S, Penel N, Bompas E, Delcambre C, Kalbacher E,
Italiano A, Collard O, et al: Efficacy and safety of regorafenib in
adult patients with metastatic osteosarcoma: A non-comparative,
randomised, double-blind, placebo-controlled, phase 2 study. Lancet
Oncol. 20:120–133. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mayank AK, Pandey V, Vashisht AA, Barshop
WD, Rayatpisheh S, Sharma T, Haque T, Powers DN and Wohlschlegel
JA: An Oxygen-dependent interaction between FBXL5 and the
CIA-targeting complex regulates iron homeostasis. Mol Cell.
75:382–393.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Viñas-Castells R, Frías Á, Robles-Lanuza
E, Zhang K, Longmore GD, García de Herreros A and Díaz VM: Nuclear
ubiquitination by FBXL5 modulates Snail1 DNA binding and stability.
Nucleic Acids Res. 42:1079–1094. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gong J, Cao J, Liu G and Huo JR: Function
and mechanism of F-box proteins in gastric cancer (review). Int J
Oncol. 47:43–50. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang M, Dai W, Ke Z and Li Y: Functional
roles of E3 ubiquitin ligases in gastric cancer. Oncol Lett.
20:222020.PubMed/NCBI
|
|
30
|
Wu W, Ding H, Cao J and Zhang W: FBXL5
inhibits metastasis of gastric cancer through suppressing Snail1.
Cell Physiol Biochem. 35:1764–1772. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hinokuma H, Kanamori Y, Ikeda K, Hao L,
Maruno M, Yamane T, Maeda A, Nita A, Shimoda M, Niimura M, et al:
Distinct functions between ferrous and ferric iron in lung cancer
cell growth. Cancer Sci. 114:4355–4364. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cho YA, Kim SE, Park CK, Koh HH, Park CK
and Ha SY: Loss of F-Box and leucine rich repeat protein 5 (FBXL5)
expression is associated with poor survival in patients with
hepatocellular carcinoma after curative resection: A Two-institute
study. Cancer Genomics Proteomics. 20:298–307. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Park CK, Heo J, Ham WS, Choi YD, Shin SJ
and Cho NH: Ferroportin and FBXL5 as prognostic markers in advanced
stage clear cell renal cell carcinoma. Cancer Res Treat.
53:1174–1183. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang H, Lu Y, Wang M, Wu Y, Wang X and Li
Y: Roles of E3 ubiquitin ligases in gastric cancer carcinogenesis
and their effects on cisplatin resistance. J Mol Med (Berl).
99:193–212. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu C, Liu Y, Yu Y, Zhao Y and Yu A:
Comprehensive analysis of ferroptosis-related genes and prognosis
of cutaneous melanoma. BMC Med Genomics. 15:392022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jiao Q, Du X, Wei J, Li Y and Jiang H:
Oxidative stress regulated iron regulatory protein IRP2 through
FBXL5-Mediated Ubiquitination-proteasome way in SH-SY5Y cells.
Front Neurosci. 13:202019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sasaki K, Hitora T, Nakamura O, Kono R and
Yamamoto T: The role of MAPK pathway in bone and soft tissue
tumors. Anticancer Res. 31:549–553. 2011.PubMed/NCBI
|
|
38
|
Huang X, Zeng J, Ruan S, Lei Z, Zhang J
and Cao H: The use of matrine to inhibit osteosarcoma cell
proliferation via the regulation of the MAPK/ERK signaling pathway.
Front Oncol. 14:13388112024. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ren J, Lv L, Tao X, Zhai X, Chen X, Yu H,
Zhao X, Kong X, Yu Z, Dong D and Liu J: The role of CBL family
ubiquitin ligases in cancer progression and therapeutic strategies.
Front Pharmacol. 15:14325452024. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Terzi EM, Sviderskiy VO, Alvarez SW,
Whiten GC and Possemato R: Iron-sulfur cluster deficiency can be
sensed by IRP2 and regulates iron homeostasis and sensitivity to
ferroptosis independent of IRP1 and FBXL5. Sci Adv. 7:eabg43022021.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ruiz JC and Bruick RK: F-box and
leucine-rich repeat protein 5 (FBXL5): Sensing intracellular iron
and oxygen. J Inorg Biochem. 133:73–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Qiu C, Liu T, Luo D, Luan D, Cheng L and
Wang S: Novel therapeutic savior for osteosarcoma: The endorsement
of ferroptosis. Front Oncol. 12:7460302022. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang M, Jike Y, Liu K, Gan F, Zhang K,
Xie M, Zhang J, Chen C, Zou X, Jiang X, et al: Exosome-mediated
miR-144-3p promotes ferroptosis to inhibit osteosarcoma
proliferation, migration, and invasion through regulating ZEB1. Mol
Cancer. 22:1132023. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ma Y, Cong L, Shen W, Yang C and Ye K:
Ferroptosis defense mechanisms: The future and hope for treating
osteosarcoma. Cell Biochem Funct. 42:e40802024. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nantajit D, Lin D and Li JJ: The network
of epithelial-mesenchymal transition: Potential new targets for
tumor resistance. J Cancer Res Clin Oncol. 141:1697–1713. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cheng ZX, Wang DW, Liu T, Liu WX, Xia WB,
Xu J, Zhang YH, Qu YK, Guo LQ, Ding L, et al: Effects of the HIF-1α
and NF-κB loop on epithelial-mesenchymal transition and
chemoresistance induced by hypoxia in pancreatic cancer cells.
Oncol Rep. 31:1891–1898. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Meyers PA: Muramyl Tripeptide-Phosphatidyl
ethanolamine encapsulated in liposomes (L-MTP-PE) in the treatment
of osteosarcoma. Adv Exp Med Biol. 1257:133–139. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Traweek RS, Cope BM, Roland CL, Keung EZ,
Nassif EF and Erstad DJ: Targeting the MDM2-p53 pathway in
dedifferentiated liposarcoma. Front Oncol. 12:10069592022.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Marvalim C, Datta A and Lee SC: Role of
p53 in breast cancer progression: An insight into p53 targeted
therapy. Theranostics. 13:1421–1442. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Huang R, Yang L, Zhang Z, Liu X, Fei Y,
Tong WM, Niu Y and Liang Z: RNA m6A Demethylase ALKBH5 protects
against pancreatic ductal adenocarcinoma via targeting regulators
of iron metabolism. Front Cell Dev Biol. 9:7242822021. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xu M, Tao J, Yang Y, Tan S, Liu H, Jiang
J, Zheng F and Wu B: Ferroptosis involves in intestinal epithelial
cell death in ulcerative colitis. Cell Death Dis. 11:862020.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Geneen LJ, Dorée C and Estcourt LJ:
Interventions for improving adherence to iron chelation therapy in
people with sickle cell disease or thalassaemia. Cochrane Database
Syst Rev. 3:CD0123492023.PubMed/NCBI
|
|
53
|
Sandoval-Acuña C, Torrealba N, Tomkova V,
Jadhav SB, Blazkova K, Merta L, Lettlova S, Adamcová MK, Rosel D,
Brábek J, et al: Targeting mitochondrial iron metabolism suppresses
tumor growth and metastasis by inducing mitochondrial dysfunction
and mitophagy. Cancer Res. 81:2289–2303. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu Y, Yu C, Luo M, Cen C, Qiu J, Zhang S
and Hu K: Ferroptosis in cancer treatment: Another way to rome.
Front Oncol. 10:5711272020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhang B and Zhang Y, Li R, Li J, Lu X and
Zhang Y: The efficacy and safety comparison of first-line
chemotherapeutic agents (high-dose methotrexate, doxorubicin,
cisplatin, and ifosfamide) for osteosarcoma: A network
meta-analysis. J Orthop Surg Res. 15:512020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wu Y, Song Y, Wang R and Wang T: Molecular
mechanisms of tumor resistance to radiotherapy. Mol Cancer.
22:962023. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhou J, Lan F, Liu M, Wang F, Ning X, Yang
H and Sun H: Hypoxia inducible factor-1α as a potential therapeutic
target for osteosarcoma metastasis. Front Pharmacol.
15:13501872024. View Article : Google Scholar : PubMed/NCBI
|