Introduction
Multiple myeloma (MM) is a malignant hematologic
tumor that originates in the bone marrow. Annually, ~588,161
individuals worldwide are diagnosed with MM (1). In 2020, among 1,278,362 cases of
leukemia, lymphoma and MM, 176,404 cases of MM were reported,
accounting for 14% of the total (2). Despite the unprecedented response and
survival rates in MM treatment, the disease remains considered
incurable due to its complex pathogenesis, recurrence and drug
resistance (3). Furthermore, MM
manifests with various clinical symptoms, leading to misdiagnosis
and underdiagnosis, which can delay optimal treatment and pose a
significant threat to patients' lives (4). Therefore, it is crucial to explore the
pathogenesis of MM and identify reliable laboratory diagnostic
biomarkers.
Aging is a natural and inevitable process that
occurs over time in organisms (5).
Pathologically, aging results from a cumulative accumulation of
stress, injury, infection, immune response and metabolic disorders
(6). Aging is one of the most
prominent risk factors for various malignancies (7). There is substantial evidence of a
bidirectional relationship between aging and malignant diseases,
both of which share numerous common characteristics (8). Notably, cellular aging processes
contribute to the pathogenesis of MM (9). Additionally, aging-related dysfunction
of T cells and the immune system can exacerbate MM onset and
progression (10–12). Thus, investigating aging at the
molecular level may offer novel insights for the clinical
translation of MM therapies.
By integrating bulk profiles with machine learning
algorithms [least absolute shrinkage and selection operator (LASSO)
regression and support vector machine recursive feature elimination
(SVM-RFE)], this study identified TXN as an aging-associated
diagnostic biomarker for MM. TXN functions as a pro-inflammatory
factor by activating the NF-κB signaling pathway and generating
reactive oxygen species (ROS) (13,14).
In vitro assays further confirmed its upregulated expression
in MM cell lines, highlighting its pathogenic potential.
Materials and methods
ARGs and sequencing dataset
In this study, 307 genes were selected from the
Human Ageing Genomic Resources (http://hagr.ageing-map.org/). The mRNA expression
profile datasets GSE6477 and GSE16558 were downloaded from the Gene
Expression Omnibus database (https://www.ncbi.nlm.nih.gov/geo/). The GSE6477
dataset was derived from the GPL96 (HG-U133A) Affymetrix Human
Genome U133A Array, while the GSE16558 dataset was derived from the
GPL6244 (HuGene-1_0-st) Affymetrix Human Gene 1.0 ST Array. GSE6477
contains bone marrow CD138+ samples from 125 patients
with MM and 15 normal bone marrow CD138+ controls, while
GSE16558 includes bone marrow CD138+ samples from 60
patients with MM and 5 normal bone marrow CD138+
controls. The bulk data were preprocessed using R software (version
4.2.3), with batch effects removed using the ‘SVA’ package and
normalization performed using the ‘normalize’ package for
subsequent analysis. All datasets included the corresponding
clinical information for both patients with MM and normal
individuals.
Screening of differentially expressed
genes (DEGs)
DEGs between MM and normal bone marrow were
identified in the integrated GSE6477 and GSE16558 dataset using a
corrected P-value <0.05 and |log2fold change
(FC)|>1 as screening criteria. The ‘limma’ package was used for
DEG analysis and the intersection of aging-related genes (ARGs) was
further identified to obtain aging-related DEGs (ARDEGs). The
Wilcoxon rank-sum test was employed to analyze the expression of
ARDEGs between MM and normal samples. Cluster analysis was
performed on the selected DEGs, and visualizations including
heatmaps, volcano plots and box plots were generated using the
‘pheatmap’ and ‘ggplot2’ packages.
Protein-protein interactions (PPI) and
correlation analysis of ARDEGs
To examine the interactions between ARDEGs, the
Search Tool for the Retrieval of Interacting Genes and proteins
(STRING) database (https://string-db.org/) was utilized. The interaction
network was constructed with a confidence score threshold of
>0.7. The PPI network was visualized and further analyzed using
Cytoscape software (version 3.8.1; http://cytoscape.org/). The Pearson correlation was
calculated using the ‘correlation graph’ function in R software to
identify the relationships between ARDEGs.
Gene Ontology (GO) and pathway
enrichment analysis of ARDEGs
GO, Kyoto Encyclopedia of Genes and Genomes (KEGG)
and Disease Ontology (DO) enrichment analyses were performed using
the ‘GOplot’, ‘KEGGplot’ and ‘DOplot’ packages in R software. GO
analysis was performed for the categories biological process,
cellular component and molecular function, while KEGG analysis
examined the signaling pathways of ARDEGs. DO analysis was used to
explore disease enrichment. A P<0.05 was considered indicative
of significantly differential enrichment of DEGs.
Machine learning screening for
ARGs
LASSO regression analysis of ARDEGs was conducted
using the ‘glmnet’ package on the integrated GSE6477 and GSE16558
datasets. The SVM-RFE algorithm was applied using the ‘e1071’
package and the ‘caret’ package was used to intersect the results
of the two algorithms to identify ARGs in MM.
Evaluating the potential value of
candidate genes
To validate the potential diagnostic value of the
candidate genes, receiver operating characteristic (ROC) curves
were plotted and the area under the ROC curve (AUC) was calculated
using the ‘pROC’ package.
Validation of ARGs in other
datasets
The GSE39754 and GSE5900 datasets were used to
validate the expression patterns of the candidate genes. GSE39754
was derived from the GPL5175 (HuEx-1_0-st) Affymetrix Human Exon
1.0 ST Array, while GSE5900 was derived from the GPL570
(HG-U133_Plus_2) Affymetrix Human Genome U133 Plus 2.0 Array. Both
datasets included bone marrow CD138+ samples from 170
patients with MM and 28 normal bone marrow CD138+
controls, providing a basis for evaluating the diagnostic efficacy
of the candidate genes. Clinical information for both patients with
MM and normal individuals was included in all datasets.
Cell lines and culture
The cell lines GM12878 (human normal B lymphocyte
cell line), U266 (human MM lymphocyte-like cell line) and RPMI8226
(human MM peripheral blood B lymphocyte cell line) were obtained
from the Shanghai Academy of Biological Sciences. U266 and GM12878
were designated as the MM group and GM12878 was designated as the
control group. All cell lines underwent short tandem repeat
authentication and mycoplasma testing, conducted by the Shanghai
Academy of Biological Sciences. These cell lines were cultured in
complete RPMI 1640 medium supplemented with 1%
penicillin-streptomycin (Gibco; Thermo Fisher Scientific, Inc.) and
10% fetal bovine serum (FBS Gibco; Thermo Fisher Scientific, Inc.).
Cell incubations were performed under strictly regulated conditions
of 37°C and 5% CO2 in a humidified incubator.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol reagent (Takara
Bio, Inc.), and its concentration, purity and integrity were
evaluated with a NanoDrop™2000 spectrophotometer (Thermo Fisher
Scientific, Inc.). According to the manufacturer's instructions, RT
was performed using 1 µg of total RNA, processed with HiScript II Q
RT SuperMix for qPCR (+gDNA wiper) (Vazyme Biotech Co., Ltd.) along
with a gDNA eraser (Vazyme Biotech Co., Ltd.). The concentration,
purity and integrity of the resulting cDNA were subsequently
assessed using the same NanoDrop spectrophotometer. Real-time qPCR
was carried out using SYBR Green MasterMix (cat. no. 11203ES50;
Yeasen Biotechnology Co., Ltd.) and StepOne Software v.2.3 (Applied
Biosystems; Thermo Fisher Scientific, Inc.).
The reaction conditions were as follows:
Pre-denaturation at 95°C for 5 min, followed by 40 cycles of
denaturation at 95°C for 30 sec, annealing at 55°C for 30 sec and
extension at 72°C for 30 sec, with a final extension at 72°C for 10
min, with three biological replicates for each sample. Data
analysis was conducted using the 2−∆∆Cq method (15), normalizing against the expression
levels of the reference gene GAPDH. The primer sequences used in
the RT-qPCR assays are provided in Table I.
 | Table I.PCR primers used in this study. |
Table I.
PCR primers used in this study.
| TXN forward
primer |
5′-CTGGATTATGCAGAGTACGTTCG-3′ |
|---|
| TXN reverse
primer |
5′-CGACTTGCTGCTTGCTCAATTT-3′ |
| GADPH forward
primer |
5′-TGACTTCAACAGCGACACCCA-3′ |
| GADPH reverse
primer |
5′-CACCCTGTTGCTGTAGCCAAA-3′ |
Statistical analysis
All statistical analyses in this bioinformatics
study were performed using R software (version 4.2.2). Pearson
correlation analysis was conducted to explore associations between
different variables. A P-value or false discovery rate threshold of
<0.05 was set as the criterion for statistical significance. For
the experimental component, statistical analyses were conducted
using GraphPad Prism (version 8.0.2; Dotmatics), with each
experiment including at least three biological replicates. Values
are expressed as the mean ± SD. Differences between the two
datasets were evaluated using either two-way ANOVA (no post-hoc
test was performed) or Student's t-test, with P<0.05 considered
statistically significant.
Results
Identification of ARDEGs based on
MM-retrospective analysis
After preprocessing the integrated GSE6477 and
GSE16558 dataset, differential gene analysis was conducted on bone
marrow CD138+ samples from 185 patients with MM and 20
normal CD138+ samples using the merged data (from
GSE6477 and GSE16558). A correction with criteria of P<0.05 and
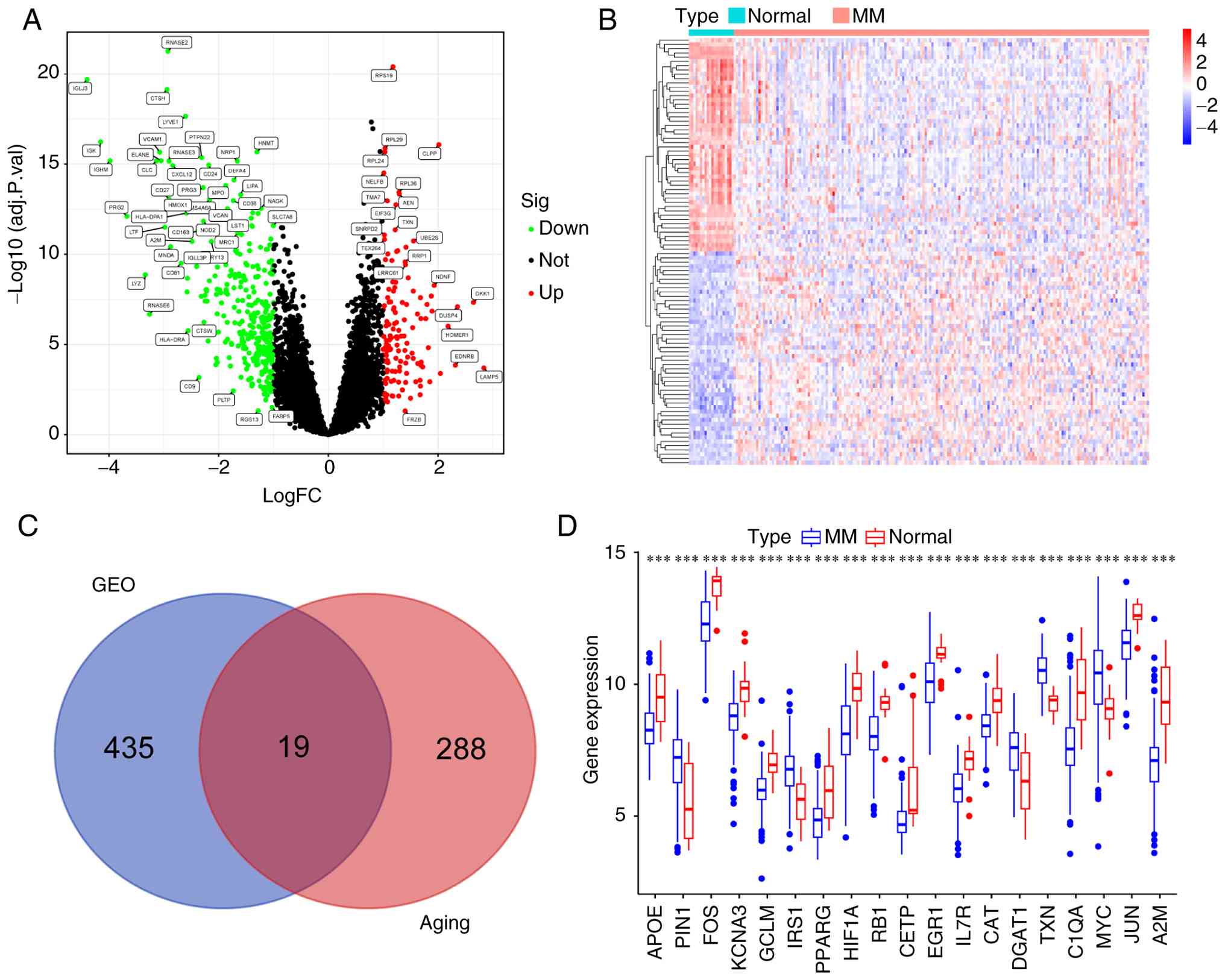
|log2FC|>1 yielded 454 DEGs, which are depicted in
the volcano plot and heatmap (Fig. 1A
and B). Additionally, principal component analysis (PCA) was
performed to assess the overall variance and clustering of samples,
as shown in Fig. S1. By
intersecting these DEGs with the Aging-Associated Gene Library, 19
ARGs linked to MM were identified, including A2M, APOE, PIN1, FOS,
KCNA3, GCLM, IRS1, PPARG, HIF1A, RB1, CETP, EGR1, IL7R, CAT, DGAT1,
TXN, C1QA, MYC and JUN (Fig. 1C).
Among these, 5 genes were upregulated, while 14 genes were
downregulated in patients with MM compared to normal controls. The
box plot illustrates significant expression differences of these 19
MM-related aging genes between MM and normal samples (Fig. 1D). PIN1, TXN and MYC were the top
three upregulated genes, whereas A2M, C1QA and HIF1A were the top
three downregulated genes (Table
II).
| Table II.Comparison of the 19 ARDEGs in
multiple myeloma samples with those in normal samples. |
Table II.
Comparison of the 19 ARDEGs in
multiple myeloma samples with those in normal samples.
| Gene symbol |
Log2FC | Direction of
differential expression | P-value | Adjusted
P-value | Chromosome |
|---|
| PIN1 | 1.413596257 | Up |
9.50×10−6 |
1.08×10−4 | 19p13 |
| TXN | 1.221137514 | Up |
2.28×10−14 |
4.40×10−12 | 9q31.3 |
| MYC | 1.177462797 | Up |
1.59×10−3 |
7.38×10−3 | 8q24.21 |
| IRS1 | 1.100398111 | Up |
7.24×10−7 |
1.23×10−5 | 2q36.3 |
| DGAT1 | 1.079197683 | Up |
8.10×10−6 |
9.50×10−5 | 8q24.3 |
| A2M | −2.48525999 | Down |
1.20×10−13 |
1.94×10−11 | 12p13.31 |
| C1QA | −2.027119477 | Down |
1.47×10−10 |
8.60×10−9 | 1p36.12 |
| HIF1A | −1.61783154 | Down |
3.65×10−8 |
7.35×10−7 | 14q23.2 |
| FOS | −1.473078631 | Down |
4.39×10−9 |
1.54×10−7 | 14q24.3 |
| CETP | −1.353050943 | Down |
5.50×10−9 |
1.90×10−7 | 16q13 |
| RB1 | −1.284638924 | Down |
4.99×10−9 |
7.73×10−7 | 13q14.2 |
| PPARG | −1.22131096 | Down |
1.04×10−8 |
3.30×10−7 | 3p25.2 |
| APOE | −1.198617718 | Down |
5.17×10−8 |
1.29×10−6 | 19q13.32 |
| KCNA3 | −1.196436349 | Down |
1.88×10−7 |
6.92×10−6 | 1p13.3 |
| JUN | −1.146427429 | Down |
1.08×10−8 |
3.40×10−7 | 1p32.1 |
| EGR1 | −1.033395792 | Down |
3.45×10−5 |
3.23×10−4 | 5q31.2 |
| GCLM | −1.006391329 | Down |
4.29×10−9 |
1.52×10−7 | 1p22.1 |
| IL7R | −1.001299766 | Down |
2.55×10−6 |
3.60×10−5 | 5p13.2 |
| CAT | −1.000447561 | Down |
1.12×10−9 |
4.82×10−8 | 11p13 |
PPI network and correlation analysis
of ARDEGs
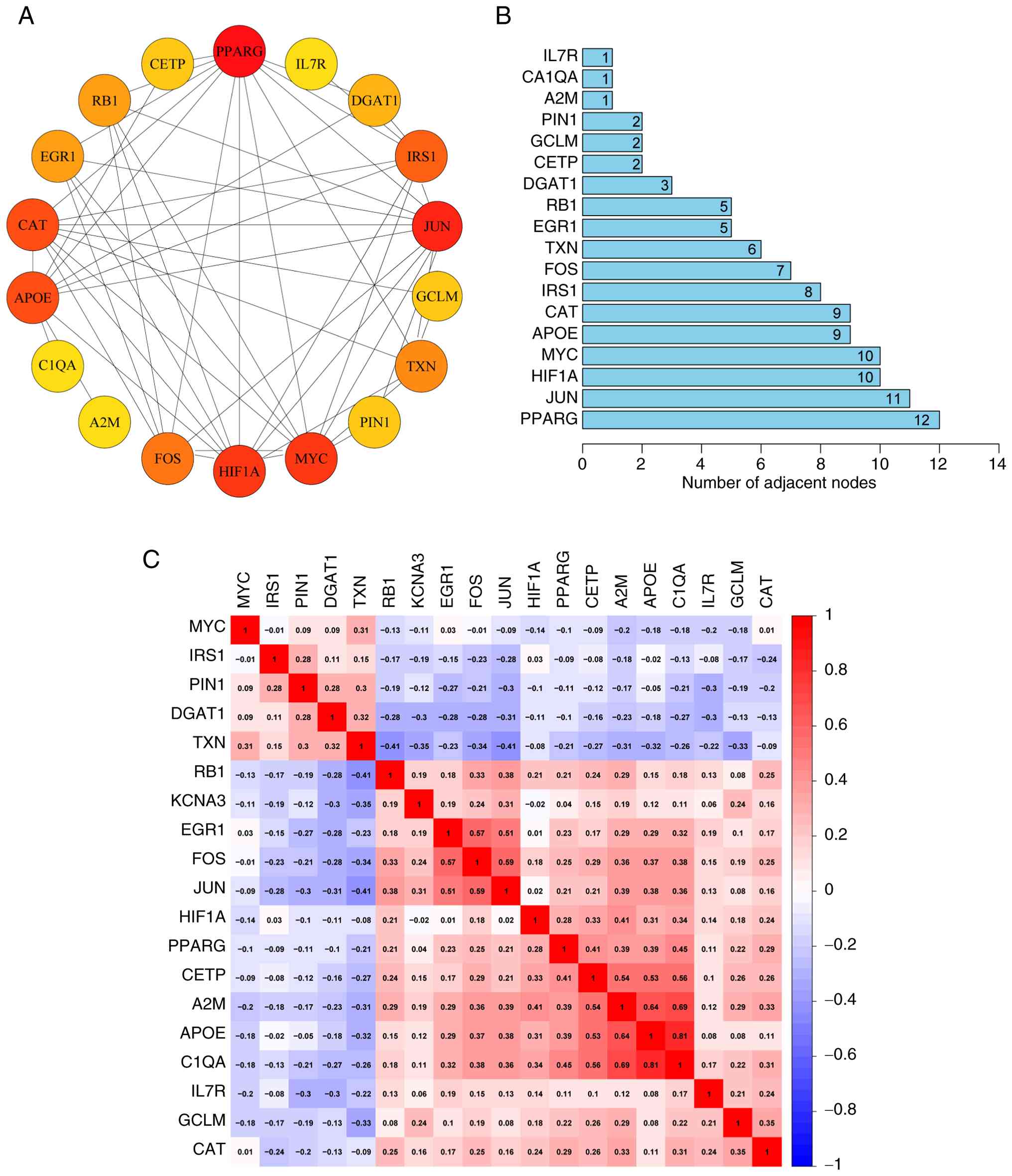
PPI analysis was performed to examine the
interactions among the 19 ARDEGs (Fig.
2A), and the number of genes interacting with each other was
quantified (Fig. 2B). The analysis
revealed that 18 genes interacted with at least one other gene.
PPARG interacted with 12 genes, JUN with 11, and both MYC and HIF1A
interacted with 10 genes. Correlation analysis of the expression
levels of the 18 interacting genes showed significant associations
between them. Notably, the expression levels of C1QA, APOE and A2M
were significantly positively correlated (r>0.6). The top three
genes associated with C1QA were APOE (r=0.81), A2M (r=0.69) and
CETP (r=0.56). For TXN, the top three associated genes were JUN
(r=−0.41), RB1 (r=−0.41) and KCNA3 (r=−0.35) (Fig. 2C).
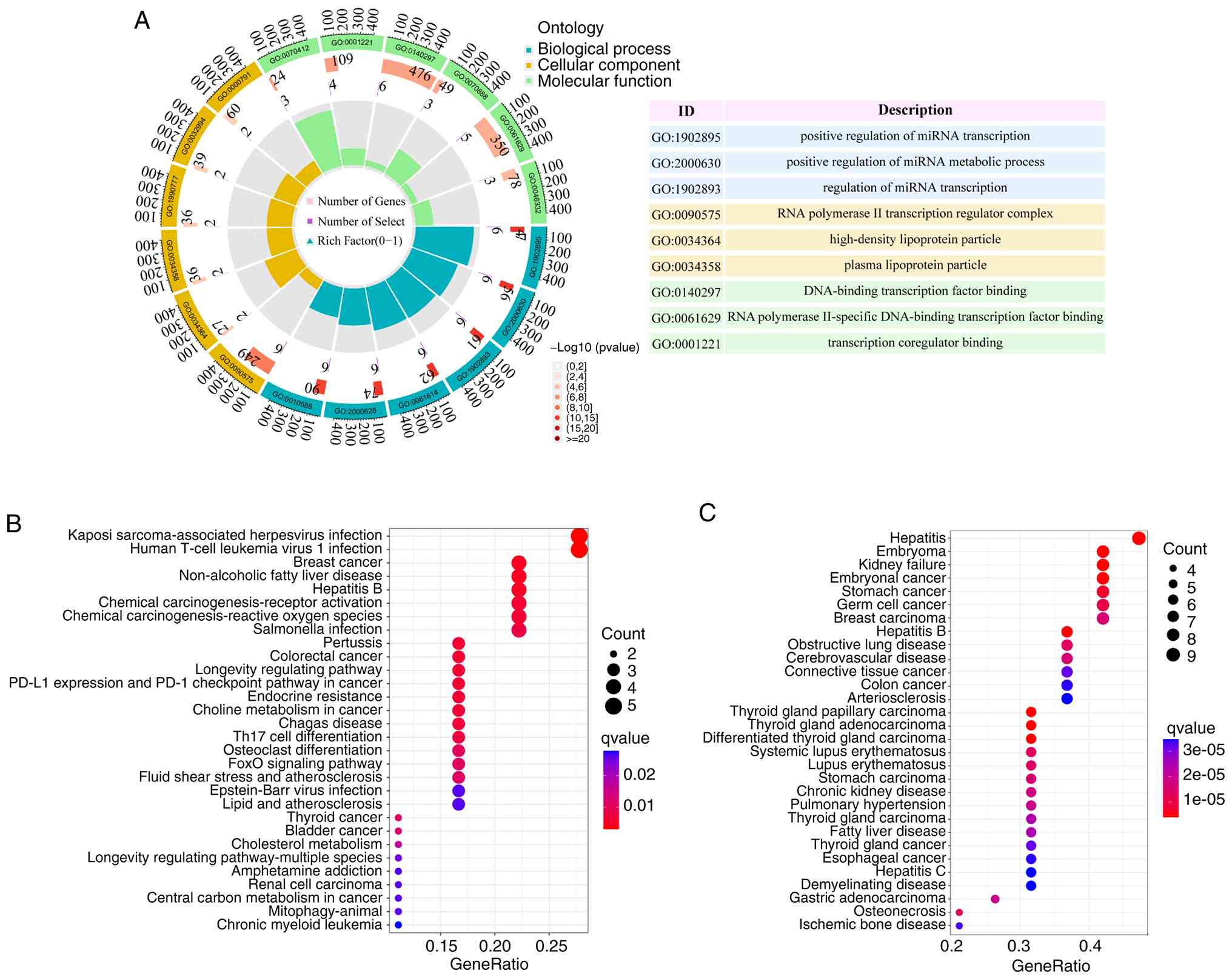
Enrichment analysis of ARDEGs
GO, KEGG and DO enrichment analyses were conducted
using R software to determine the potential biological functions of
the ARDEGs. The most significant GO enrichment terms included the
‘positive regulation of miRNA transcription’, ‘regulation of miRNA
transcription’ (biological process), ‘RNA polymerase II
transcription regulator complex’, ‘high-density lipoprotein
particle’, ‘plasma lipoprotein particle’ (cellular component),
‘DNA-binding transcription factor binding’, ‘RNA polymerase
II-specific DNA-binding transcription factor binding’ and
‘transcription co-regulator binding’ (molecular function) (Fig. 3A). KEGG enrichment analysis revealed
that the ARDEGs were primarily involved in signaling pathways
related to Kaposi sarcoma-associated herpesvirus infection and
human T-cell leukemia virus 1 infection, suggesting their role in
the dynamic regulation of the immune system (Fig. 3B). DO enrichment analysis showed
that ARDEGs are crucial in the occurrence and development of
hepatitis (Fig. 3C).
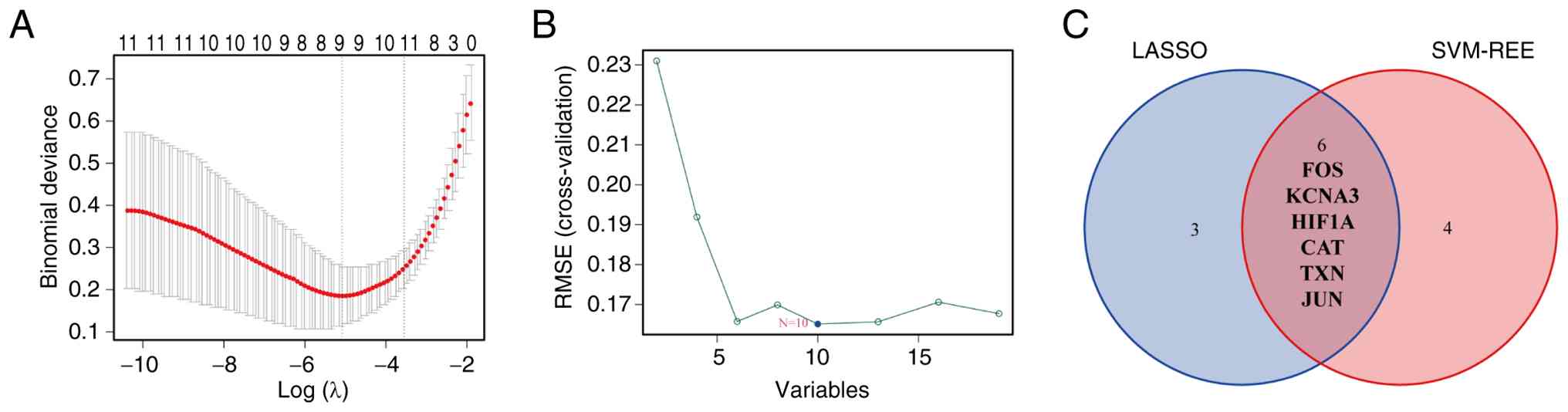
Screened candidate genes of MM
The LASSO regression algorithm identified 9 genes
among the 19 MM ARDEGs: APOE, FOS, KCNA3, IRS1, PPARG, HIF1A, CAT,
TXN and JUN (Fig. 4A). In
comparison, the SVM-RFE algorithm identified 10 genes: TXN, RB1,
A2M, FOS, JUN, GCLM, CAT, KCNA3, HIF1A and IL7R (Fig. 4B). A total of six candidate genes,
overlapping between the two algorithms, were selected: FOS, KCNA3,
HIF1A, CAT, TXN and JUN (Fig. 4C).
Notably, all these 6 molecules are closely associated with
aging.
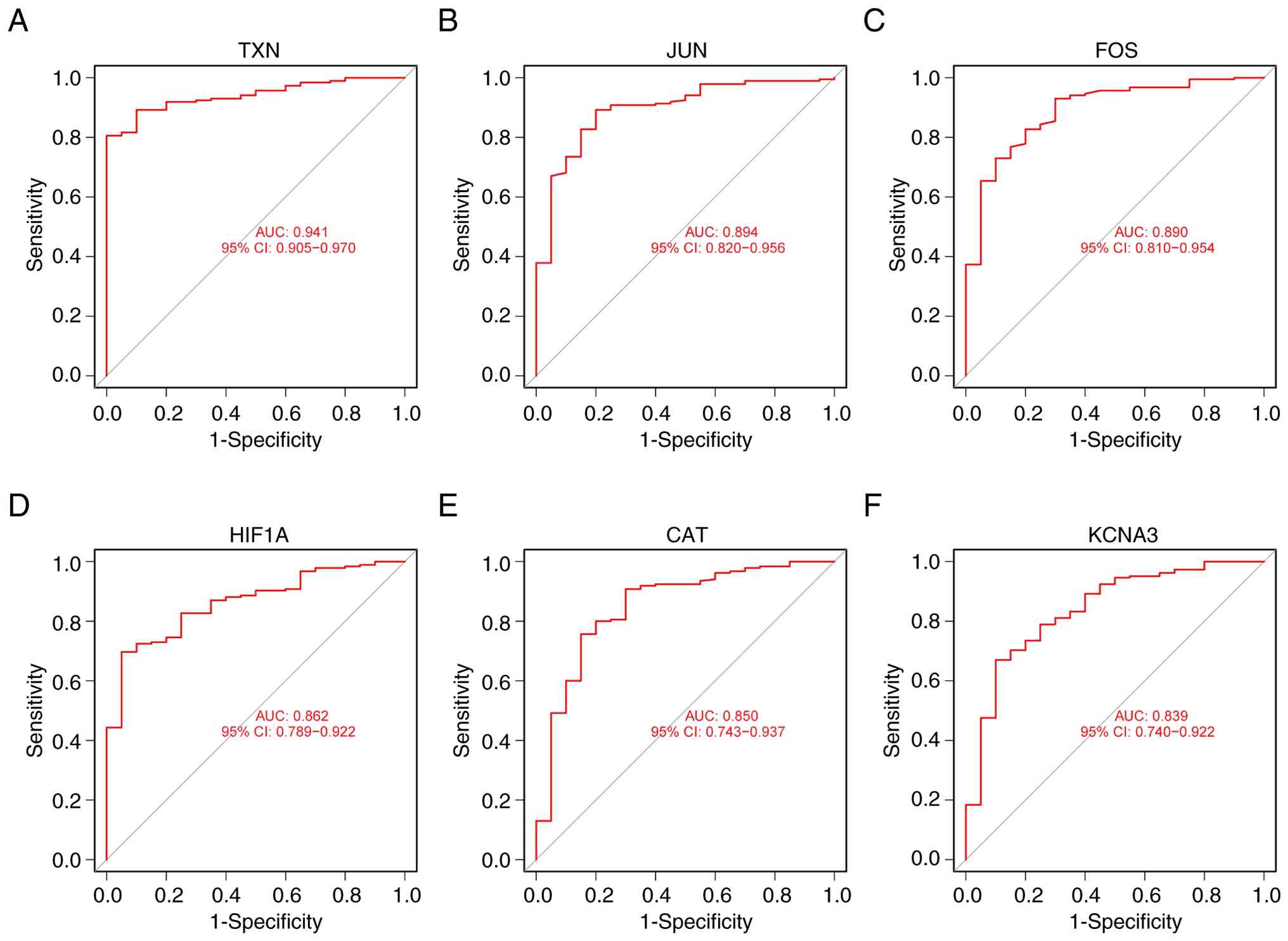
ROC curves of six candidate genes in
MM and normal bone marrow
The ‘pROC’ package was used to analyze the
expression of six candidate genes in 185 MM and 20 normal bone
marrow samples from the GSE6477 and GSE16558 datasets, generating
ROC curves. The AUC, which combines sensitivity and specificity,
was used to assess the diagnostic effectiveness of these genes. The
six candidate genes demonstrated high diagnostic value for MM.
Among them, TXN exhibited the highest diagnostic value in MM
samples (AUC=0.941). The diagnostic values for the other genes were
as follows: JUN (AUC=0.894), FOS (AUC=0.890), HIF1A (AUC=0.862),
CAT (AUC=0.850) and KCNA3 (AUC=0.839) (Fig. 5A-F).
Validation of candidate genes in other
datasets
The GSE39754 and GSE5900 datasets were used as a
validation cohort to assess the accuracy of the analysis results
and the expression levels of the six candidate genes. As shown in
Fig. 6A-F, a significant difference
in TXN and HIF1A expression was observed between MM and normal
samples. TXN was overexpressed and HIF1A expression was reduced in
MM samples. Notably, HIF1A expression was more variable in the MM
samples. Additionally, the ROC curve analysis in the validation
cohort (Fig. 6G-L) showed that TXN
(AUC=0.707), JUN (AUC=0.527), FOS (AUC=0.503), HIF1A (AUC=0.728),
CAT (AUC=0.569) and KCNA3 (AUC=0.589) exhibited diagnostic value.
Among these, TXN had the highest diagnostic value and can serve as
a key diagnostic marker for MM.
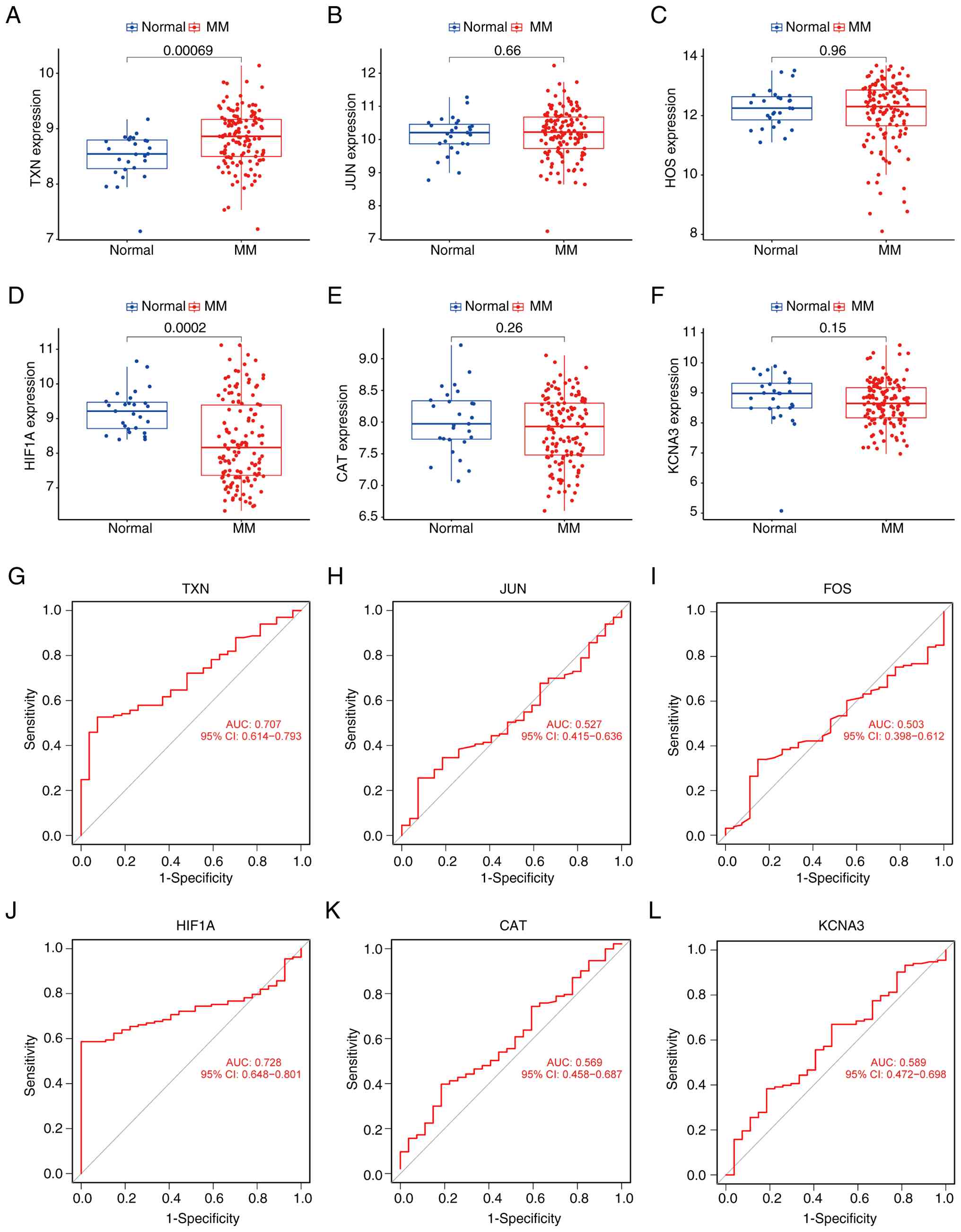 | Figure 6.Validation of hub genes in other
datasets. (A-F) Expression levels of the candidate diagnostic
markers (A) TXN, (B) JUN, (C) FOS, (D) HIF1A, (E) CAT and (F) KCNA3
in the MM and normal samples in the validation cohort. (G-L) ROC
curve of the six candidate genes in MM and normal samples
(validation dataset). (G) TXN, (H) JUN, (I) FOS, (J) HIF1A, (K) CAT
and (L) KCNA3. AUC, area under the ROC curve; ROC, receiver
operating characteristic; MM, multiple myeloma. |
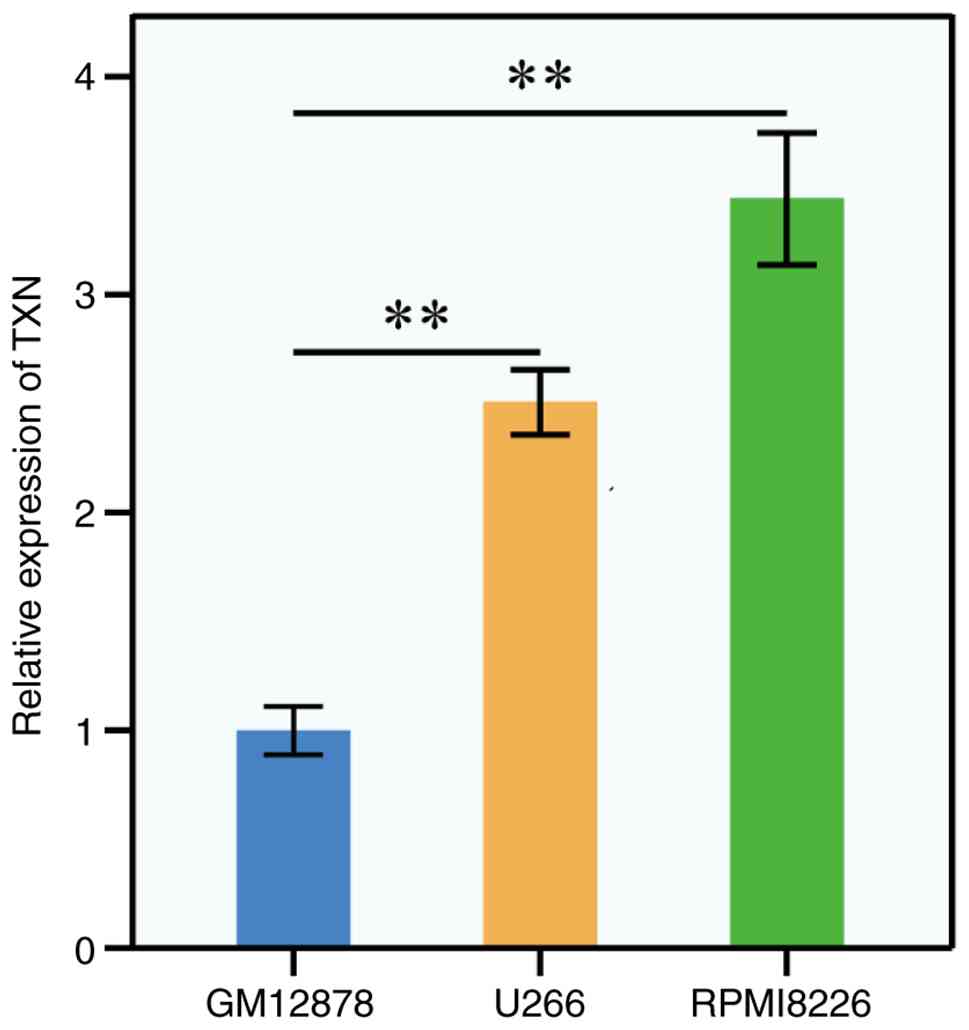
Verification of the key genes in the
human MM cell line
To validate the bioinformatics results, RT-qPCR was
performed on the human MM cell lines U266 and RPMI8226 to further
analyze TXN expression. The results from the U266 and RPMI8226 cell
validation tests were consistent with the bioinformatics analysis,
showing significantly higher expression levels of TXN compared to
normal cells. Specifically, the expression of TXN was significantly
greater in U266 (P<0.01) and RPMI8226 (P<0.01) cells than in
normal cells (Fig. 7).
Discussion
In the present study, bioinformatics analysis and
machine learning techniques were utilized to identify aging-related
diagnostic and druggable targets for patients with MM. The results
demonstrated that TXN could serve as a robust diagnostic biomarker
for detecting MM pathogenesis. Additionally, the potential
biological and molecular functions of TXN in MM were explored,
offering novel insights into the disease's underlying
mechanisms.
Aging is the primary risk factor for most
malignancies (5). For instance, MM
is an aging-related disease that poses significant health
challenges for the elderly (16).
Epidemiological investigation pointed out the global burden of MM
is increasing in numerous countries due to an aging population
(17). Studies have highlighted
aging-associated epigenetic and genetic alterations that
characterize both MM development and normal aging processes
(9). For example, aging can destroy
the immune systems, which leads to the impairment of immune
surveillance and pathogenesis of MM (18). Besides, enhancer of zeste homolog 2
inhibition induces senescence via the ERK1/2 signaling pathway in
MM (19). Additionally,
inositol-requiring enzyme 1α inhibition enhances mitochondrial
quality in CD8+ T cells, thereby improving their
anti-tumor efficacy (20).
Furthermore, genetic instability in patients with MM contributes to
an anti-senescence effect, promoting disease progression (21). Age-related mesenchymal stromal cell
senescence has also been linked to the progression from monoclonal
gammopathy of undetermined significance to MM (22). In the present study, 6 molecules
were identified (FOS, KCNA3, HIF1A, CAT, TXN and JUN) that are
closely associated with aging. The m6A mRNA demethylase FTO in
granulosa cells can slow FOS-dependent ovarian aging (23). KCNA3 is implicated in cognitive
aging (24). P53/HIF-1α regulates
neuronal aging and autophagy in spinal cord ischemia/reperfusion
injury (25). CAT contributes to
autophagy-induced vascular smooth muscle cell senescence (26). Additionally, Astragalus
polysaccharide alleviates oxidative stress and senescence in
osteoarthritis chondrocytes via the GCN2/ATF4/TXN axis
(27). Activation of JUN has long
been considered a hallmark of cellular senescence (28). The present study confirmed that TXN
can serve as a promising diagnostic biomarker for MM, a malignant
blood cancer characterized by uncontrolled plasma cell
proliferation. TXN plays a pivotal role in the pathophysiology of
MM, affecting both the cellular aging process and the maintenance
of cellular redox balance. This study further explored the novel
mechanisms through which TXN contributes to MM, providing valuable
insights into the disease's pathophysiology. Additionally, the
TXN-ALDH1L2 axis has been shown to promote the progression of
colorectal cancer and radioresistance by activating the NF-κB
signaling pathway (14), which is
also closely associated with MM pathogenesis (29). Thus, TXN may regulate NF-κB
signaling to modulate MM progression.
Despite these insights, there are still gaps in our
understanding. Specifically, further studies are needed to
elucidate the precise mechanisms by which TXN acts in MM, as well
as to develop targeted therapeutic strategies based on TXN's
regulatory pathways, particularly regarding its role in apoptosis
regulation. Furthermore, additional research is required to
identify therapeutic agents targeting TXN for MM treatment, which
could enhance the clinical applicability of the present findings.
The in vitro results obtained from the GM12878
EBV-transformed lymphoblastoid cell line should also be validated
in patient samples to ensure authenticity. Besides, ARGs identified
in the present study that are also involved in the pathogenesis of
other disease or biological functions showed a weaker association
with MM pathogenesis, indicating a need for further pre-clinical
validation of these DEG mechanisms in MM. Furthermore, differences
in the diagnostic performance of TXN between the training and
validation sets of patients with MM suggest predictive
heterogeneity. Therefore, future studies should assess TXN's
diagnostic performance in multi-center cohorts to strengthen its
diagnostic robustness. Lastly, the impact of TXN dysregulation on
the overall survival of patients with MM should be explored in
clinical studies to provide further evidence for the clinical
relevance of TXN.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by the Hubei Key Laboratory of Central
Nervous System Tumor and Intervention (grant no. ZZYKF202210).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
HL and SS conducted the research. YWu and YWa
performed the data analysis. HL and SS wrote the manuscript. YuL
and YiL contributed to the conception and design of the study,
provided supervision and critically revised the manuscript. YuL and
YiL checked and confirm the authenticity and accuracy of all the
raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kumar SK, Rajkumar V, Kyle RA, van Duin M,
Sonneveld P, Mateos MV, Gay F and Anderson KC: Multiple myeloma.
Nat Rev Dis Primers. 3:170462017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Palumbo A and Anderson K: Multiple
myeloma. N Engl J Med. 364:1046–1060. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rajkumar SV: Multiple myeloma: Every year
a new standard? Hematol Oncol. 37 (Suppl 1):62–65. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Raab MS, Podar K, Breitkreutz I,
Richardson PG and Anderson KC: Multiple myeloma. Lancet.
374:324–339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
De Winter G: Aging as disease. Med Health
Care Philos. 18:237–243. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hou Y, Dan X, Babbar M, Wei Y, Hasselbalch
SG, Croteau DL and Bohr VA: Ageing as a risk factor for
neurodegenerative disease. Nat Rev Neurol. 15:565–581. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fulop T, Larbi A, Douziech N, Levesque I,
Varin A and Herbein G: Cytokine receptor signalling and aging. Mech
Ageing Dev. 127:526–537. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kanagal-Shamanna R, Beck DB and Calvo KR:
Clonal hematopoiesis, inflammation, and hematologic malignancy.
Annu Rev Pathol. 19:479–506. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Urban VS, Cegledi A and Mikala G: Multiple
myeloma, a quintessential malignant disease of aging: A geroscience
perspective on pathogenesis and treatment. Geroscience. 45:727–746.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ullrich F, Bröckelmann PJ, Turki AT, Khan
AM, Chiru ED, Vetter M, Tresckow BV, Wirth R, Cordoba R,
Ortiz-Maldonado V, et al: Impact of immunological aging on T
cell-mediated therapies in older adults with multiple myeloma and
lymphoma. J Immunother Cancer. 12:e0094622024. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Visram A and Kourelis TV: Aging-associated
immune system changes in multiple myeloma: The dark side of the
moon. Cancer Treat Res Commun. 29:1004942021.PubMed/NCBI
|
|
12
|
Zweegman S, Palumbo A, Bringhen S and
Sonneveld P: Age and aging in blood disorders: Multiple myeloma.
Haematologica. 99:1133–1137. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang B and Zhao S: Polydatin regulates
proliferation, apoptosis and autophagy in multiple myeloma cells
through mTOR/p70s6k pathway. Onco Targets Ther. 10:935–944. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu L, Guo Q, Luo Z, Wang Y, Weng J, Chen
Y, Liang W, Li Y, Zhang Y, Chen K, et al: TXN inhibitor impedes
radioresistance of colorectal cancer cells with decreased ALDH1L2
expression via TXN/NF-κB signaling pathway. Br J Cancer.
127:637–648. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fotiou D, Ntanasis-Stathopoulos I,
Gavriatopoulou M and Dimopoulos MA: Multiple myeloma: Current and
future management in the aging population. Maturitas. 138:8–13.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mafra A, Laversanne M, Marcos-Gragera R,
Chaves HVS, Mcshane C, Bray F and Znaor A: The global multiple
myeloma incidence and mortality burden in 2022 and predictions for
2045. J Natl Cancer Inst. 117:907–914. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Casey M and Nakamura K: The
cancer-immunity cycle in multiple myeloma. Immunotargets Ther.
10:247–260. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guo S, Tang Q, Gao X, Hu L, Hu K, Zhang H,
Zhang Q, Lai Y, Liu Y, Wang Z, et al: EZH2 inhibition induces
senescence via ERK1/2 signaling pathway in multiple myeloma. Acta
Biochim Biophys Sin (Shanghai). 56:1055–1064. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wan Y, Wang J, Chen M, Wang J, Nan F,
Huang H, Liu Z and Hou J: Dual roles of IRE1α inhibition in
reversing mitochondrial ROS-induced CD8(+) T-cell senescence and
exerting direct antitumor effects in multiple myeloma. J Immunother
Cancer. 13:e0110442025. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Alagpulinsa DA, Szalat RE, Poznansky MC
and Shmookler Reis RJ: Genomic instability in multiple myeloma.
Trends Cancer. 6:858–873. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Plakhova N, Panagopoulos V, Cantley MD,
Trainor LJ, Hewett DR, Clark KC, Gardiner J, Yong A, Lee C, Horvath
N, et al: Age-related mesenchymal stromal cell senescence is
associated with progression from MGUS to multiple myeloma.
Leukemia. 39:1464–1475. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang ZX, Wang YN, Li ZY, Dai ZH, He Y,
Chu K, Gu JY, Ji YX, Sun NX, Yang F and Li W: The m6A mRNA
demethylase FTO in granulosa cells retards FOS-dependent ovarian
aging. Cell Death Dis. 12:7442021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Abdul Sani NF, Amir Hamzah AIZ, Abu Bakar
ZH, Mohd Yusof YA, Makpol S, Wan Ngah WZ and Damanhuri HA: Gene
expression profile in different age groups and its association with
cognitive function in healthy malay adults in Malaysia. Cells.
10:16112021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu X, Wang J, Shen K and Jin W:
p53/HIF-1α regulates neuronal aging and autophagy in spinal cord
ischemia/reperfusion injury. Mech Ageing Dev. 222:1120002024.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Salazar G, Cullen A, Huang J, Zhao Y,
Serino A, Hilenski L, Patrushev N, Forouzandeh F and Hwang HS:
SQSTM1/p62 and PPARGC1A/PGC-1alpha at the interface of autophagy
and vascular senescence. Autophagy. 16:1092–1110. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen K, Qin C, Ji C, Yu Y, Wu Y, Xu L,
Jiang Y and Zou G: Astragalus polysaccharide alleviates oxidative
stress and senescence in chondrocytes in osteoarthritis via
GCN2/ATF4/TXN axis. Int J Biol Macromol. 310((Pt 3)): 1432852025.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hernandez-Segura A, Nehme J and Demaria M:
Hallmarks of Cellular Senescence. Trends Cell Biol. 2018.28.
436–453. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Matthews GM, de Matos Simoes R, Dhimolea
E, Sheffer M, Gandolfi S, Dashevsky O, Sorrell JD and Mitsiades CS:
NF-κB dysregulation in multiple myeloma. Semin Cancer Biol.
39:68–76. 2016. View Article : Google Scholar : PubMed/NCBI
|