Introduction
The Hedgehog (Hh) signaling pathway plays an
important role in embryogenesis and histogenesis (1). There are three mammalian Hh genes:
Sonic hedgehog (Shh), Desert hedgehog (Dhh) and
Indian hedgehog (Ihh). Ihh plays an important role in
chondrogenesis (2), whereas Dhh is
necessary for the development of peripheral nerves and
spermatogenesis (3,4). In mammals, the role of Shh is well
established in limb development, tissue polarity and patterning of
the central nervous system (1). Shh
is also involved in the morphogenesis of the axial skeleton, lungs,
skin, hair and teeth (1,5). Hh proteins activate the signaling
cascade by binding to the membrane-bound receptor Patched that
leads to the activation of transmembrane protein Smoothened and the
sequential activation of intracellular signal transduction
(1). As a result, the expression of
Hh target genes is initialized through the activation of Gli
transcription factors, which in mammals contains Gli1, Gli2 and
Gli3 homologues (5,6).
The role of Shh is not restricted to embryonic
development. Previous studies proved that Shh is also required for
adult stem cell (SC) proliferation and differentiation (7). Thus, anti-Shh antibodies block the
proliferation of human haematopoietic SC; the addition of purified
Shh accordingly induces cell proliferation and differentiation
in vitro (8). However, an
inappropriate activation of Hh signaling can initiate tumorigenesis
in different tissues (9–11). It has been shown that human tumors
such as basal cell carcinoma, medulloblastoma, rhabdomyosarcoma,
fibroma and meningioma are associated with abnormalities in the Shh
signaling pathway (12).
Recent data have shown that the Shh pathway is
involved in prostate development (13). These data have also implicated the
Shh pathway in the development of prostate cancer, the second most
prevalent cause of male cancer-related death in the world. Thus,
in vitro blocking of the Shh pathway inhibits the
proliferation of human prostate cancer cell lines and cells in
primary prostate cancer cultures (10,14,15).
Moreover, the activation of the Shh pathway has been detected in
human metastatic and high-grade prostate tumors (15). Human prostate cancer is associated
with mutations and loss of heterozygosity in the Shh pathway
negative regulator suppressor of fused (15). In contrast to accumulated human
data, recent in vivo data from the LADY prostate cancer mice
model have shown that the expression level of Shh and other
components of the Hh signaling pathway (Ptc1 and Gli1) are not
increased during prostate tumor development (16).
The present study aimed to evaluate the activity of
the Shh pathway during prostate cancer progression. To assess a
possible correlation between activation of the Shh pathway and
prostate cancer development, we examined the expression of the
different Shh pathway proteins (Shh, Gli1, Gli2 and Gli3) and the
proliferation markers FoxA1 and Notch1 in prostate biopsies in
C57Bl/6 (WT) and transgenic adenocarcinoma of the mouse prostate
(TRAMP) mice at 12, 17 and 21 weeks of age.
Materials and methods
Animals and tissue preparation
Heterozygous male TRAMP and WT mice (Jackson
Laboratory, USA) were maintained under conventional housing
conditions. Animal procedures were approved by the Ethics Committee
of the University of Tartu. Animals were euthanized using cervical
dislocation. After excision, a part of each prostate was
immediately frozen for RNA isolation. For immunohistochemistry,
tissues were embedded in Tissue-Tek O.C.T (Sakura Finetek, The
Netherlands) and frozen. Samples were stored at −70°C until further
analysis.
Immunohistochemistry
Prostate blocks were cut into 10-μm sections (Microm
HM500OM; Carl Zeiss, Germany). After fixation, peroxidase was
blocked in 0.75% hydrogen peroxide in methanol. After incubation
with 3% normal goat serum (Sigma-Aldrich, USA), sections were
incubated overnight at 4°C with either rabbit anti-Gli1 IgG
polyclonal antibody (pAb) (1:200; Santa Cruz Biotechnology, USA)
(17,18), rabbit anti-Gli2 IgG, pAb (1:200;
Santa Cruz Biotechnology) (19),
mouse anti-Gli3 IgM, monoclonal antibody (mAb) (1:100, clone 5E1;
reviewed in ref. 20), mouse
anti-Shh IgG, mAb (1:50, clone 5E1; Developmental Studies Hybridoma
Bank, USA) (21,22), mouse anti-FoxA1 IgG pAb (1:50;
Cemines, USA) (23) or rat
anti-Notch1 IgG pAb (1:10; Developmental Studies Hybridoma Bank)
(24,25). Sections were then incubated with
secondary antibodies for 1 h at room temperature. HRP-conjugated
goat anti-rabbit IgG and goat anti-rat IgG antibodies were used for
rabbit and rat pAb, respectively (1:200; Jackson ImmunoResearch
Laboratories, USA). HRP-conjugated goat anti-mouse IgG -positive
IgM antibody (1:200; Jackson ImmunoResearch Laboratories) was used
for mouse pAb. Mouse on mouse staining protocol (Jackson
ImmunoResearch Laboratories) was used if the pAB had mouse origin.
As a negative control pAB was replaced with vehicle. Positive
staining was visualized using diaminobenzidine (Vector
Laboratories, USA). Counterstaining was performed with Harris
hematoxylin, followed by tissue dehydration and mounting with
Assistant Histokitt (Chemi-Teknik, Norway). Enumeration of the
cells was determined using a Zeiss light microscope. Cells were
counted in two randomly selected high-power fields (HPF;
magnification, ×400) pro animal.
RNA isolation and RT-PCR
Total prostate RNA was isolated using TRI Reagent
(Ambion, USA). mRNA expression analysis of Shh, Gli1, Gli2 and Gli3
was carried out by RT-PCR. RNA was reversibly transcribed using
oligo-dT primers and Stratascript III Reverse Transcriptase
(Invitrogen, USA). The specific primer pairs used were: Gli1
5′-GTCCACCAAC CAACTATG-3′ forward, 5′-TCCAGGTCAAGAGAGTCC-3′ reverse
(product is 481 bp); Gli2 5′-GAACGAAGAGA CAGCTCCAC-3′ forward,
5′-CTGTGGAAACGTTGCACT-3′ reverse (337 bp); Gli3
5′-CTGGAAAGGAGCGGGAAAG-3′ forward, 5′-CTGAGGCTGCAGTGGGATTAC-3′
reverse (256 bp); Shh 5′-TACAAGCAGTTTATTCCCAACGT-3′ forward,
5′-GACCCTCATAGTGTAGAGACT-3′ reverse (243 bp); actin
5′-TACCACAGGCATTGTGATGGA-3′ forward, 5′-CAACGTCACACTTCATGATGG-3′
reverse (272 bp). Reaction was initially performed at 95°C for 2
min, followed by 35 cycles of denaturation (at 94°C for 30 sec),
annealing (for Gli1 at 52°C, for Gli2 at 54°C, for Gli3 at 60°C,
for Shh at 58°C and for actin at 57°C, for 30 sec) and extension
(at 72°C for 1 min). The PCR products were identified by
electrophoresis in 1.5% agarose gels. DNA bands were visualized
using ethidium bromide.
Statistical analysis
Data are expressed as the mean number of positive
cells pro HPF ± SEM. Statistical analysis was carried out using
non-parametric analysis of variance (Kruskal-Wallis test) to
evaluate variance among the groups studied. If a significant
variance was found, an unpaired two-group test (Mann-Whitney U
test) was used to determine significant differences between
individual groups. The Spearman-rank correlation test was used to
detect the relationship between variables. P<0.05 was considered
to be a statistically significant difference.
Results
Shh pathway components in prostate
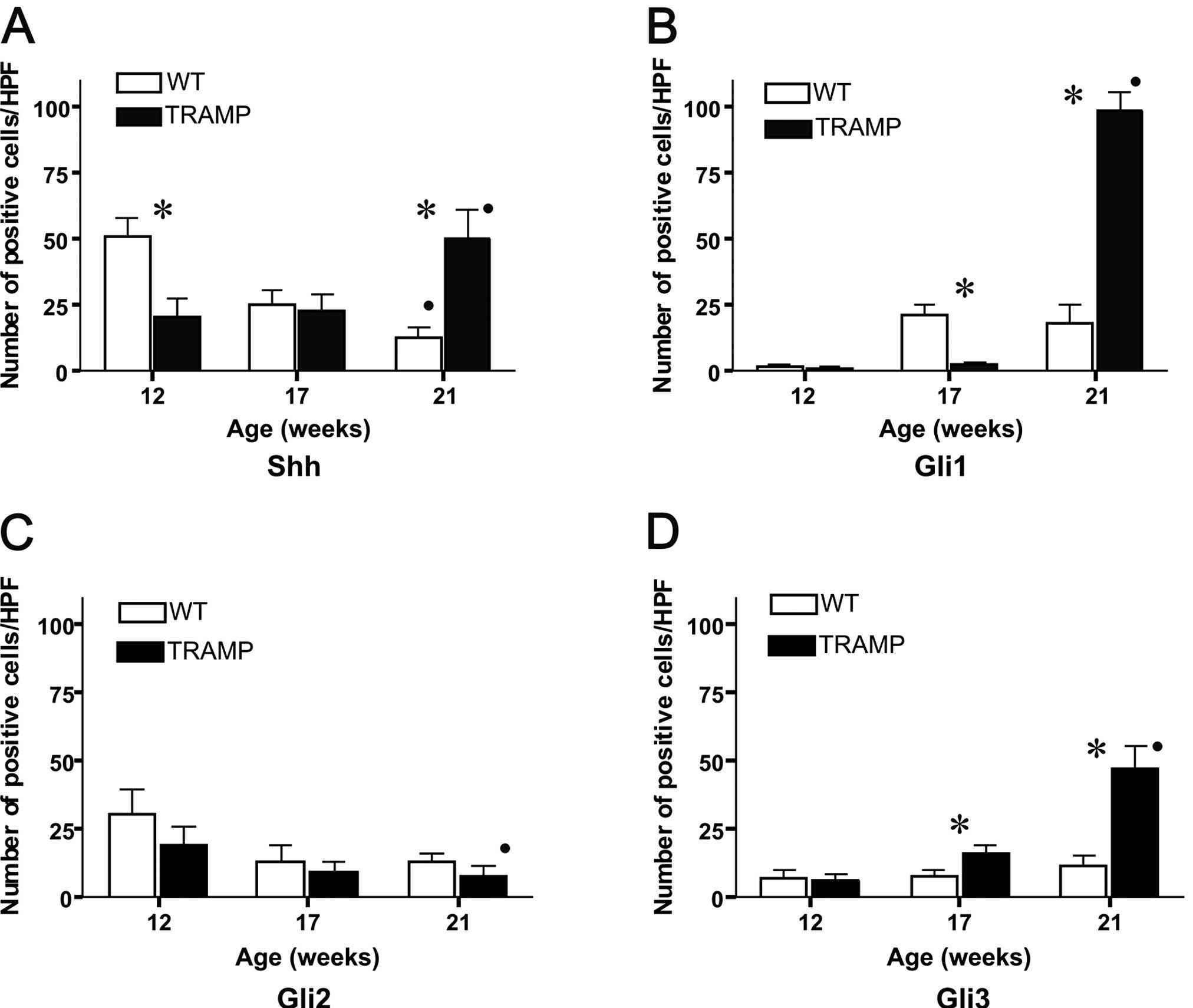
In the WT mice the number of Shh-positive cells
decreased significantly in an age-dependent manner, and were 50±6,
25±5 and 13±4 cells/HPF at 12, 17 and 21 weeks of age, respectively
(P=0.002, Rs=−0.95). In the TRAMP mice, the number of Shh-positive
cells increased significantly during the period studied (20±7, 23±6
and 50±10 cells/HPF at 12, 17 and 21 weeks of age, respectively;
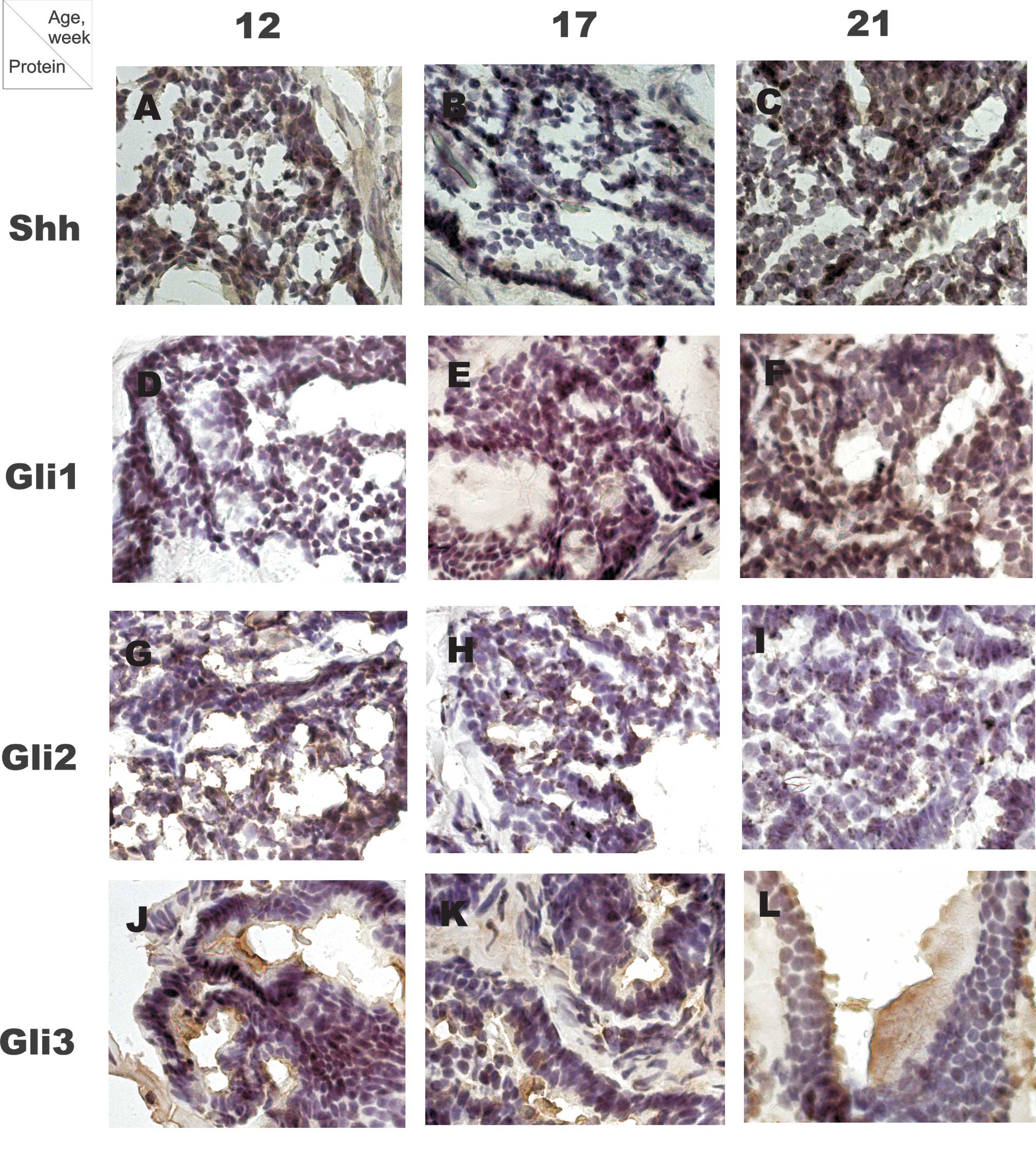
P=0.01, Rs=0.73) (Fig. 1A-C,
Fig. 2A). The number of
Shh-positive cells was higher in WT compared to TRAMP mice at 12
weeks of age, whereas in older mice, at 21 weeks of age, the number
of Shh-positive cells was higher in TRAMP mice (P=0.02 for both
comparisons).
The number of Gli1-positive cells in WT was 2±0.9
cells/HPF at 12 weeks, 21±4 at 17 weeks and 18±7 cells/HPF at 21
weeks of age. In the TRAMP mice, the number of Gli1-positive cells
increased from 2±0.5 cells/HPF at 12 weeks to 4±0.8 cells at 17
weeks and 100±7 cells/HPF at 21 weeks of age (P=0.002, Rs=0.93)
(Fig. 1D-F, Fig. 2B). At 12 weeks of age no difference
was noted in the number of Gli1-positive cells between WT and TRAMP
mice. At 17 weeks the number of Gli1-positive cells was
significantly lower in TRAMP compared to WT mice, but in older
mice, at 21 weeks of age, the number of Gli1-positive cells was
higher in TRAMP compared to WT mice (P=0.02 for both
comparisons).
In the WT mice, the number of Gli2-positive cells
was 30±9 cells/HPF at 12 weeks, 13±6 at 17 weeks and 13±3 cells/HPF
at 21 weeks of age. In the TRAMP mice, the number of Gli2-positive
cells decreased in an age-dependent manner and were 19±7 cells/HPF
at 12 weeks, 10±3 at 17 weeks and 8±3 cells/HPF at 21 weeks of age
(P=0.03, Rs=−0.7) (Fig. 1G-I,
Fig. 2C). No significant
differences were noted in the number of Gli2-positive cells between
WT and TRAMP mice at all time points studied.
In the WT mice, the number of Gli3-positive cells at
12 weeks of age was 7±3 cells/HPF, and at 17 and 21 weeks 8±2 and
11±3 cells/HPF, respectively. The number of Gli3-positive cells in
the TRAMP mice increased during the study period, and were 6±2
cells/HPF at 12 weeks, 16±3 at 17 weeks and 48±8 cells/HPF at 21
weeks of age (P=0.002, Rs=0.095) (Fig.
1J-L, Fig. 2D). No significant
difference was noted in the number of Gli3-positive cells between
WT and TRAMP mice at 12 weeks of age. The number of Gli3-positive
cells was higher in TRAMP compared to WT mice at 17 and 21 weeks of
age (P=0.02 for both comparisons).
Messenger RNA expression of the Shh
signaling pathway components
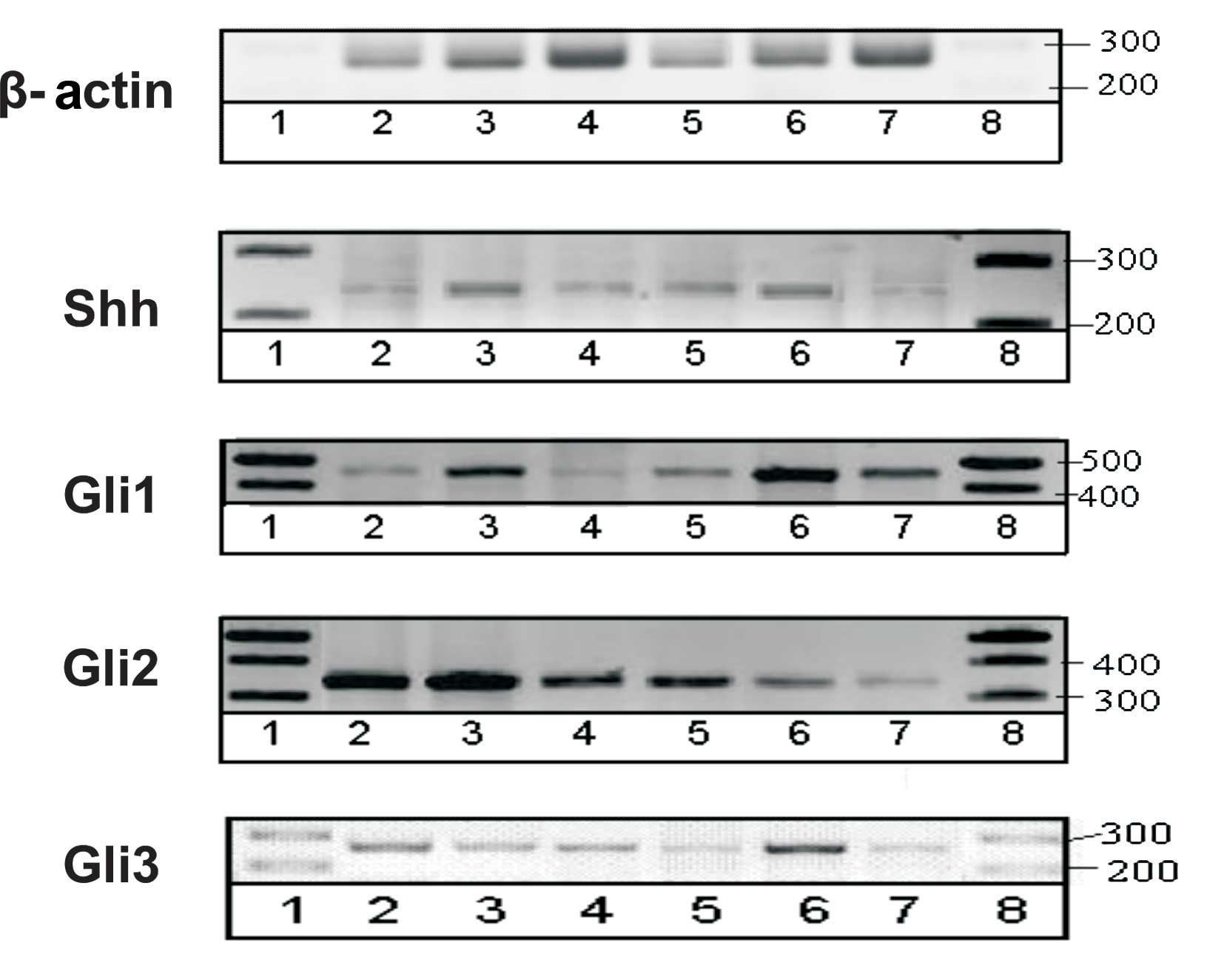
RT-PCR yields semi-quantitative results and,
therefore, was performed to verify the presence of respective genes
in mouse prostate. The scan of representative gels is shown in
Fig. 4.
FoxA1 protein in prostate
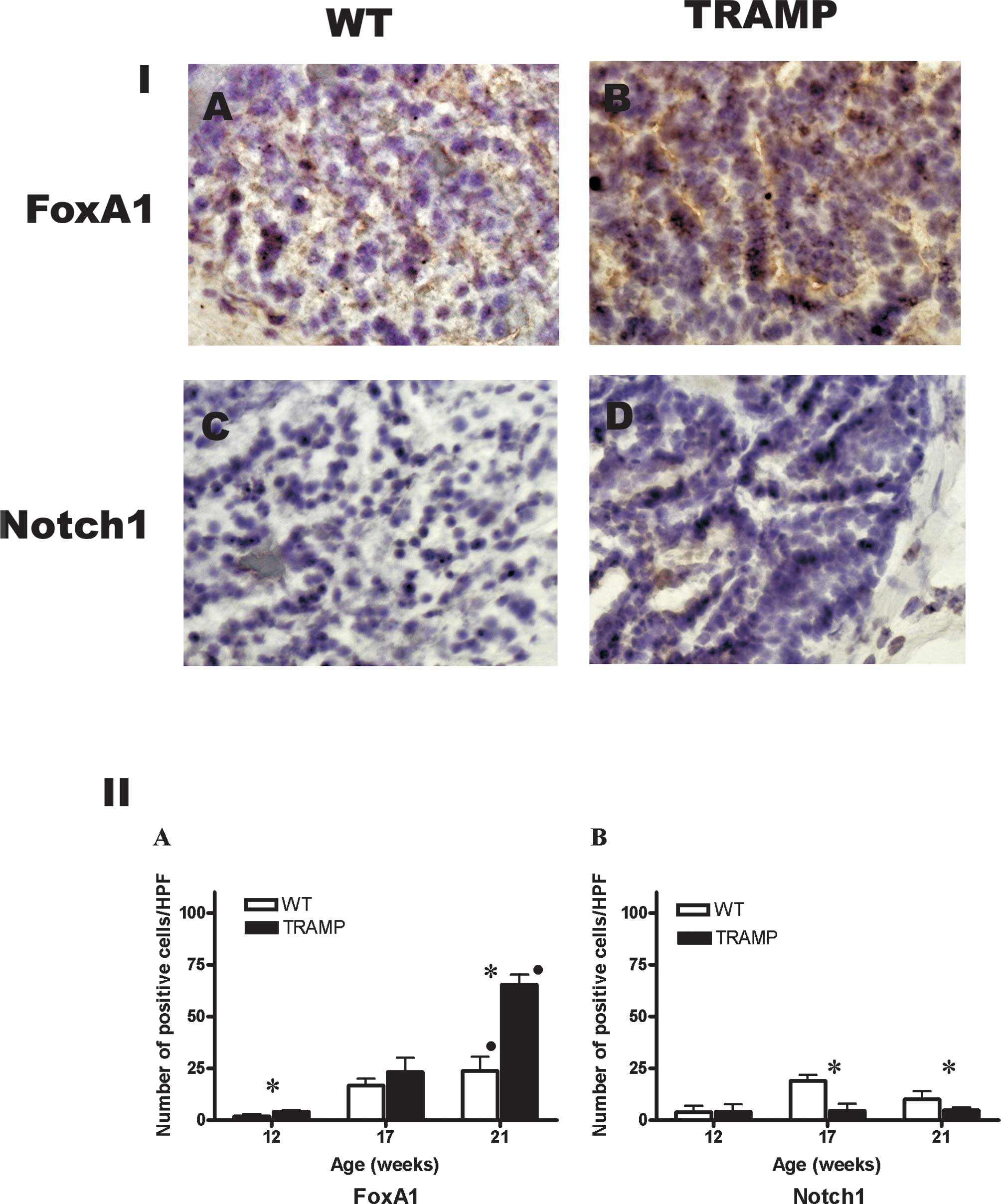
In the WT mice, the number of FoxA1-positive cells
increased significantly at 2±0.9, 16±3 and 24±6 cells/HPF at 12, 17
and 21 weeks of age, respectively (P=0.004, Rs=0.9). In TRAMP mice,
the number of FoxA1-positive cells increased significantly at
4±0.8, 24±6 and 65±5 cells/HPF at 12, 17 and 21 weeks of age,
respectively (P=0.002, Rs=0.95) (Fig.
3, Panel IA-B and Panel IIA). The number of FoxA1-positive
cells was higher in WT compared to TRAMP mice at 12 and 21 weeks of
age (P=0.03 and 0.02, respectively).
Notch1 protein in prostate
In WT mice, the number of Notch1-positive cells was
4±2.5, 18±3 and 10±4 cells/HPF at 12, 17 and 21 weeks of age,
respectively. In TRAMP mice, there were 4±3, 4.5 ±3 and 5±1
Notch1-positive cells/HPF at 12, 17 and 21 weeks of age,
respectively (Fig. 3, Panel IC-D
and Panel IIB). The number of Notch1-positive cells was higher in
WT compared to TRAMP mice at 17 and 21 weeks of age (P=0.02 and
0.03, respectively).
Discussion
Our study proves that with aging the number of
Shh-positive cells increased in TRAMP and decreased in wild-type
mice. In the TRAMP mice, the increase in the number of Shh-positive
cells was paralleled by the age-dependent increase in Gli1-and
Gli3-positive cells that in turn reflects activation of the whole
Shh pathway.
Prostate cancer is the most common type of cancer in
males and is the second cause of male cancer-related death. In
Estonia, frequency of prostate cancer is 107/100,000 individuals,
making it the first cause of cancer-related death in males
(www.cancer.ee). The risk for developing prostate
cancer rises significantly with age, and 60% of newly diagnosed
cases occur in men over the age of 70.
Mouse experimental models are the most frequently
used in vivo research tools in the study of human diseases.
However, due to species-specific differences there are no suitable
wild-type mice strains to study human prostate cancer, mainly
because prostate cancer does not develop in rodents spontaneously
(26). Possible reasons for this
include the short life span of the rodent and that in contrast to
human prostate, mouse prostate atrophies with aging (26). Additionally, prostate anatomy
considerably differs between human and rodents (26,27).
Currently, well-validated prostate cancer transgenic mouse strains
TRAMP (28,29) and LADY (30) are available. In these mice strains
rat probasin gene promoter is used to direct the expression of
T/t-antigens of the SV40 virus in the prostate epithelium. Besides
the different size of the promoter, another interesting difference
between the two models is that the TRAMP mice epithelial cells may
express both large and small T/t-antigens of the SV40 virus, while
LADY is designed to express only the large T-antigen. Notably,
TRAMP mice develop high-grade prostatic intraepithelial neoplasia
and adenocarcinoma, which later metastasize primarily to the lungs
and lymph nodes. On the other hand, LADY mice develop low-grade
prostatic intraepithelial neoplasia and invasive carcinoma, which
generally fail to metastasize (31). Therefore, taking into account the
fact that human prostate adenocarcinoma is an early metastasizing
cancer, we utilized a more relevant TRAMP model.
Involvement of the Shh pathway in prostate cancer
has previously been demonstrated. Currently, the activation of
Ptch1 and Gli1, but not Shh, is considered to be a feature of the
Shh pathway activation in a signal-receiving cell (5,6). Thus,
despite the constant presence of Shh transcripts in human
metastatic prostate tumors, the Shh pathway is activated in only
25% of tumors (10). Nevertheless,
human prostate carcinomas as well as human metastatic and
high-grade prostate tumors are associated with a high expression
level of Shh mRNA (14,15).
The present study demonstrated that the number of
prostate Shh-positive cells increased in an age-dependent manner in
the TRAMP mouse model for prostate cancer. A significant difference
in the number of Shh-positive cells as well as the Shh mRNA
expression level between transgenic and wild-type mice was detected
only in older mice, at the 21st week of age. Based on the TRAMP
mice breeder description, prostate adenocarcinoma develops by the
24th week and metastasizes by the 30th week of age (www.jax.org). Therefore, the increase in the number of
Shh-positive cells preceded the estimated appearance of
adenocarcinoma per se or its metastasis in TRAMP mice. The
presence of Shh mRNA in normal adult murine and human prostate
tissue has been demonstrated (14,16).
This observation was extended and it was shown that Shh is present
in normal prostate on the protein level. Furthermore, we detected a
significant age-dependent decrease in the number of cells in
wild-type mice. Thus, our data showed that the Shh protein
expression decreases with age in healthy conditions, whereas in
prostate cancer development the Shh protein expression increases
with age and precedes cancer manifestation.
Shh signal transduction leads to the activation of
the Gli transcription factors, Gli1, Gli2 and Gli3. Gli1 serves as
a direct readout of Hh signaling and as a transcriptional activator
of the Shh pathway (5). Increased
expression level of Gli1 in human prostate cancer cell lines and
human prostate carcinomas was previously demonstrated (10,14).
We showed an age-dependent increase in the number of Gli1-positive
cells in transgenic mice. Similar to the number of the Shh-positive
cells, the number of Gli1-positive cells and its mRNA expression
was higher only in older TRAMP mice. It was shown that while Gli1
acts solely as an activator, Gli2 and Gli3 are bi-potential
transcriptional regulators (32).
Gli2 and Gli3 are activators or repressors, depending on the
context. Although we detected an age-dependent decrease in the
number of Gli2-positive cells and the level of its mRNA expression
in transgenic mice, no significant differences were noted in the
number of cells between transgenic and wild-type mice. On the other
hand, the number of Gli3-positive cells increased in an
age-dependent manner in transgenic mice, with the number of cells
beng significantly higher compared to wild-type mice in the 17th
and 21st weeks of age. This is consistent with our mRNA data and
earlier observations (33). Our
data showed that tumor development in prostate cancer-prone TRAMP
mice is associated with an age-dependent increase in Shh, Gli1 and
Gli3 and decrease in Gli2 mRNA and protein levels in prostate
tissue. Our data support the hypothesis that the following order of
events may occur during prostate carcinogenesis (34): i) a small amount of Shh activates
Gli3 and Gli1 proteins; ii) Gli3 binds to the Gli1 promoter and
induces the additional expression of Gli1 and iii) Shh pathway
proteins activate relevant target genes that lead to tumorigenesis.
Whether this hypothesis is correct remains to be tested in further
studies.
Tumor development in TRAMP mice is associated with
an increased rate of epithelial proliferation (28). FoxA1 is an important regulator of
cell proliferation and differentiation, and is continuously
expressed in human and mice prostate from its development till
aging (35,36). FoxA1 protein-enhanced expression was
repeatedly detected in human prostate carcinomas (35). We detected an age-dependent increase
in the number of prostate FoxA1-positive cells in both transgenic
and wild-type mice. Transgenic mice had an increased number of
prostate FoxA1-positive cells compared to wild-type mice. Thus, in
the current model prostate tumor progression is associated with an
increase in the number of FoxA1-positive cells, which is in
agreement with human data (35).
Taking into account that FoxA1 is constantly expressed in the human
prostate and the fact that its activation is time-dependently
associated with prostate carcinoma development, the possible role
of this marker in prostate tumor development as well as its
possible use as a diagnostic marker warrants further
investigation.
Notch1 is crucial for prostate development and the
differentiation of prostate cells (37). An interesting aspect of Notch1 is
its apparently opposite functions in tumor development, where it
can act either as an oncogene or as a tumor suppressor. For
example, a high expression of Notch1 was shown in 14 human prostate
cancer cell lines in vitro and in TRAMP prostate tissue
in vivo (38,39). On the other hand, Notch1 prostate
tumor suppressor features were demonstrated in human prostate
adenocarcinomas in vivo and Notch1 knock-out mice (37). Nevertheless, although the role of
Notch1 in the prostate cancer remains controversial, this
proliferation marker certainly warrants further investigation in
prostate tumors (reviewed in ref. 40). In this study, the number of
Notch1-positive cells was decreased at the 17th and 21st week of
age in transgenic compared to wild-type mice, suggesting that
Notch1 functions as a tumor-suppressor protein in the current
prostate cancer model.
Our study showed age-dependent activation of the Shh
pathway in the TRAMP transgenic mice. As this particular mouse
strain is likely to develop metastasizing prostate cancer, our data
actually implicate Shh pathway activation not only in the
development of primary prostate carcinomas but also in its
metastatic spread. Therefore, further use of this particular strain
as a valid tool for prostate cancer research is significant.
Acknowledgements
The authors thank Dr Tõnis Timmusk for providing the
FoxA1 and Notch1 antibodies. We are also grateful to Dr Illar Pata
for the help with the mouse work and to Drs Torben Østerlund, Piret
Michelson and Robert Tsanev for critical review of the manuscript.
The study was supported by the Estonian Science Foundation (grants
ETF7242 to S.S. and ETF8116 to P.K.), and the Estonian Ministry of
Education (Directional financing to P.K.). P.K. was supported by a
Wellcome Trust International Senior Research Fellowship for part of
these studies.
References
|
1
|
Osterlund T and Kogerman P: Hedgehog
signalling: how to get from Smo to Ci and Gli. Trends Cell Biol.
16:176–180. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chung UI, Schipani E, McMahon AP and
Kronenberg HM: Indian hedgehog couples chondrogenesis to
osteogenesis in endochondral bone development. J Clin Invest.
107:295–304. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sharghi-Namini S, Turmaine M, Meier C, et
al: The structural and functional integrity of peripheral nerves
depends on the glial-derived signal desert hedgehog. J Neurosci.
26:6364–6376. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bitgood MJ, Shen L and McMahon AP: Sertoli
cell signaling by Desert hedgehog regulates the male germline. Curr
Biol. 6:298–304. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kinzler KW, Ruppert JM, Bigner SH and
Vogelstein B: The GLI gene is a member of the Kruppel family of
zinc finger proteins. Nature. 332:371–374. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ruppert JM, Kinzler KW, Wong AJ, et al:
The GLI-Kruppel family of human genes. Mol Cell Biol. 8:3104–3113.
1988.PubMed/NCBI
|
|
7
|
Rowitch DH, BSJ, Lee SM, Flax JD, Snyder
EY and McMahon AP: Sonic hedgehog regulates proliferation and
inhibits differentiation of CNS precursor cells. J Neurosci.
19:8954–8965. 1999.PubMed/NCBI
|
|
8
|
Bhardwaj G, Murdoch B, Wu D, et al: Sonic
hedgehog induces the proliferation of primitive human hematopoietic
cells via BMP regulation. Nat Immunol. 2:172–180. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stecca B, Mas C and Ruiz i Altaba A:
Interference with HH-GLI signaling inhibits prostate cancer. Trends
Mol Med. 11:199–203. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Karhadkar SS, Bova GS, Abdallah N, et al:
Hedgehog signalling in prostate regeneration, neoplasia and
metastasis. Nature. 431:707–712. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ruiz i Altaba A: Therapeutic inhibition of
Hedgehog-GLI signaling in cancer: epithelial, stromal, or stem cell
targets? Cancer Cell. 14:281–283. 2008.PubMed/NCBI
|
|
12
|
Oldak M, Grzela T, Lazarczyk M, Malejczyk
J and Skopinski P: Clinical aspects of disrupted Hedgehog signaling
(Review). Int J Mol Med. 8:445–452. 2001.PubMed/NCBI
|
|
13
|
Datta S and Datta MW: Sonic Hedgehog
signaling in advanced prostate cancer. Cell Mol Life Sci.
63:435–448. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sanchez P, Hernandez AM, Stecca B, et al:
Inhibition of prostate cancer proliferation by interference with
Sonic Hedgehog-Gli1 signaling. Proc Natl Acad Sci USA.
101:12561–12566. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sheng T, Li C, Zhang X, et al: Activation
of the hedgehog pathway in advanced prostate cancer. Mol Cancer.
3:292004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gipp J, Gu G, Crylen C, Kasper S and
Bushman W: Hedgehog pathway activity in the LADY prostate tumor
model. Mol Cancer. 6:192007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mukherjee S, Frolova N, Sadlonova A, et
al: Hedgehog signaling and response to cyclopamine differ in
epithelial and stromal cells in benign breast and breast cancer.
Cancer Biol Ther. 5:674–683. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao J, Chen G, Cao D, et al: Expression
of Gli1 correlates with the transition of breast cancer cells to
estrogen-independent growth. Breast Cancer Res Treat. 119:39–51.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nielsen SK, Mollgard K, Clement CA, et al:
Characterization of primary cilia and Hedgehog signaling during
development of the human pancreas and in human pancreatic duct
cancer cell lines. Dev Dyn. 237:2039–2052. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hunt R, Bragina O, Drews M, et al:
Generation and characterization of mouse monoclonal antibody 5E1
against human transcription factor GLI3. Hybridoma (Larchmt).
26:231–240. 2007. View Article : Google Scholar
|
|
21
|
Unger S, Copland I, Tibboel D and Post M:
Down-regulation of sonic hedgehog expression in pulmonary
hypoplasia is associated with congenital diaphragmatic hernia. Am J
Pathol. 162:547–555. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang BE, Shou J, Ross S, Koeppen H, De
Sauvage FJ and Gao WQ: Inhibition of epithelial ductal branching in
the prostate by sonic hedgehog is indirectly mediated by stromal
cells. J Biol Chem. 278:18506–18513. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Minoo P, Hu L, Zhu N, et al: SMAD3
prevents binding of NKX2.1 and FOXA1 to the SpB promoter through
its MH1 and MH2 domains. Nucleic Acids Res. 36:179–188. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fox V, Gokhale PJ, Walsh JR, Matin M,
Jones M and Andrews PW: Cell-cell signaling through NOTCH regulates
human embryonic stem cell proliferation. Stem Cells. 26:715–723.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Djalilian AR, Namavari A, Ito A, et al:
Down-regulation of Notch signaling during corneal epithelial
proliferation. Mol Vis. 14:1041–1049. 2008.PubMed/NCBI
|
|
26
|
Maini A, Archer C, Wang CY and Haas GP:
Comparative pathology of benign prostatic hyperplasia and prostate
cancer. In Vivo. 11:293–299. 1997.PubMed/NCBI
|
|
27
|
Cunha GR, Donjacour AA, Cooke PS, et al:
The endocrinology and developmental biology of the prostate. Endocr
Rev. 8:338–362. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gingrich JR, Barrios RJ, Morton RA, et al:
Metastatic prostate cancer in a transgenic mouse. Cancer Res.
56:4096–4102. 1996.PubMed/NCBI
|
|
29
|
Gingrich JR, Barrios RJ, Kattan MW, Nahm
HS, Finegold MJ and Greenberg NM: Androgen-independent prostate
cancer progression in the TRAMP model. Cancer Res. 57:4687–4691.
1997.PubMed/NCBI
|
|
30
|
Kasper S, Sheppard PC, Yan Y, et al:
Development, progression and androgen-dependence of prostate tumors
in probasin-large T antigen transgenic mice: a model for prostate
cancer. Lab Invest. 78:319–333. 1998.
|
|
31
|
Abate-Shen C and Shen MM: Mouse models of
prostate carcinogenesis. Trends Genet. 18:S1–S5. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sasaki H, Nishizaki Y, Hui C, Nakafuku M
and Kondoh H: Regulation of Gli2 and Gli3 activities by an
amino-terminal repression domain: implication of Gli2 and Gli3 as
primary mediators of Shh signaling. Development. 126:3915–3924.
1999.PubMed/NCBI
|
|
33
|
Dai P, Akimaru H, Tanaka Y, Maekawa T,
Nakafuku M and Ishii S: Sonic Hedgehog-induced activation of the
Gli1 promoter is mediated by GLI3. J Biol Chem. 274:8143–8152.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang C, Ruther U and Wang B: The
Shh-independent activator function of the full-length Gli3 protein
and its role in vertebrate limb digit patterning. Dev Biol.
305:460–469. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mirosevich J, Gao N, Gupta A, Shappell SB,
Jove R and Matusik RJ: Expression and role of Foxa proteins in
prostate cancer. Prostate. 66:1013–1028. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gao N, Ishii K, Mirosevich J, et al:
Forkhead box A1 regulates prostate ductal morphogenesis and
promotes epithelial cell maturation. Development. 132:3431–3443.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang XD, Leow CC, Zha J, et al: Notch
signaling is required for normal prostatic epithelial cell
proliferation and differentiation. Dev Biol. 290:66–80. 2006.
View Article : Google Scholar
|
|
38
|
Leong KG and Gao WQ: The Notch pathway in
prostate development and cancer. Differentiation. 76:699–716. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shou J, Ross S, Koeppen H, de Sauvage FJ
and Gao WQ: Dynamics of notch expression during murine prostate
development and tumorigenesis. Cancer Res. 61:7291–7297.
2001.PubMed/NCBI
|
|
40
|
Bolos V, Grego-Bessa J and de la Pompa JL:
Notch signaling in development and cancer. Endocr Rev. 28:339–363.
2007. View Article : Google Scholar : PubMed/NCBI
|