Introduction
Ovarian cancer is a common aggressive disease and is
the fifth leading cause of cancer-related death in women (1). The life-time risk of ovarian cancer is
approximately 1.4% (2).
Approximately 200,000 women are diagnosed with this highly lethal
disease each year (3). Due to the
lack of obvious symptoms, substantial delays in diagnosis are quite
common, and the overall survival rate for ovarian cancer has only
slightly increased over the last 30 years in spite of advances in
surgery and chemotherapy. Therefore, understanding of the tumor
molecular biology is an essential step for the selection of the
most effective treatment strategy for ovarian cancer.
Tumorigenesis is a result of the accumulation of
genetic changes. Genomic instability leads to the transformation of
normal cells to cancer cells. This includes aberrant reduction or
loss of tumor-suppressor genes and activation of proto-oncogenes
(4).
P53 binding-proteιn 1 (53BP1) was initially
identified in a yeast two-hybrid system by Iwabuchi et al in
1994, and has been mapped to chromosomes 15q15–21 and predicts a
protein of 1972 amino acids (217 kb) (5). Similar to other mediators of
checkpoint responses (e.g., BRCA1, MDC1), 53BP1 also contains a
highly conserved tandem BRCA1 C-terminal (BRCT) domain, which is
essential for tumor-suppressor function via interaction with
ezrin-radixin-moesin (ERM) at the plasma membrane (6). Owing to its rapid accumulation to
sites of DNA double-strand breaks (DSB), 53BP1 has been considered
a cytologic marker for endogenous DSB (7). The functions of 53BP1 range from
participation in DNA-damage repair to negotiating cell cycle
checkpoints (7,8). By triggering a cell cycle checkpoint,
cells suffering DNA damage undergo cell cycle arrest and apoptosis
avoiding genomic instability which can generate cancer (9). Defects in 53BP1 were found to
contribute to the pathogenesis of various types of human cancer
(10,11), but our knowledge of the role of
53BP1 in ovarian cancer is still rudimentary. Here, we presented a
preliminary analysis of 53BP1 in the prevention of ovarian cancer
development.
Materials and methods
E. coli DH5α competent cells, Lipofectamine
2000 and RPMI-1640 medium were purchased from Invitrogen (Carlsbad,
CA, USA). N-Myc-53BP1 WT pLPC-Puro, pSUPER-neo-GFP, and
pSUPER-neo-GFP-53BP1 were purchased from Addgene, OligoEngine and
Sangon Biotech (Shanghai, China), respectively. The N-Myc-WT
pLPC-Puro was a kind gift from Professor Titia de Lange (Laboratory
of Cell Biology and Genetics of Rockefeller University). ECL for
western blotting and the gel electrophoresis device were from GE
Healthcare (USA). The antibodies for Akt, p-Akt,
P21waf1/Cip1, Bcl-2, caspase-3 and β-actin were from
Cell Signaling Technologies (Boston, MA).
Cell culture and transfection
Human ovarian cancer cell line A2780 was originally
obtained from the American Type Culture Collection, and HO-8910PM,
a highly metastatic ovarian cancer cell line (12), was obtained from the Chinese Academy
of Sciences (Shanghai, China). The cells were grown in
Rossman-Park-Memorial-Institute (RPMI)-1640 media supplemented with
10% fetal bovine serum (FBS; Haoyang Biological Manufacture Co.
Ltd., Tianjin, China) at 37°C in a humidified atmosphere containing
5% CO2. Cells in a logarithmic growth phase were
harvested for the experiments. Endogenous 53BP1 was knocked down
and overexpressed using the pSUPER-neo-GFP-53BP1 and N-Myc-WT
pLPC-Puro plasmids, respectively. The cells were transfected with
plasmids using Lipofectamine 2000 according to the manufacturer’s
protocol. Cells were selected in complete medium containing 375
μg/ml of G418 or puromycin (0.75 μg/ml) (Invitrogen). Western
blotting was used to detect the expression level of 53BP1 in all of
the cells described above.
MTT assay for cell growth
The proliferative ability of cells modified by the
variable gene was measured using
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT)
assay according to a standard protocol. The cells were seeded into
96-well plates (1.5×103 cells/well) for 1–7 days in
triplicate. At specified time points, a total of 20 μl of MTT
(Amresco, Solon, OH, USA) solution (0.5 mg/ml) was added to each
well, and the wells were incubated for an additional 4 h at 37°C.
The purple-blue MTT formazan precipitate was dissolved in 200 μl of
DMSO (Shenggong, Shanghai, China). The activity of the
mitochondria, reflecting cellular growth and viability was
evaluated at 550 nm with a microplate reader (Bio-Rad, Hercules,
CA, USA).
Colony forming assay
Cells (300) modified by the variable gene were
plated on 60-mm culture dishes in triplicate. After cells were
cultured for 10 days, colonies were fixed with methanol for 15 min
and stained with 0.1% crystal violet. Colonies of at least 50 cells
were counted and recorded.
Flow cytometric analysis of the cell
cycle
The transfected cells were seeded at a density of
106 cells/60-mm dish (BD Biosciences, San Jose, CA, USA)
in culture media contained corresponding selective antibiotics for
48 h. The cells were then washed with ice-PBS, harvested and
resuspended in 1 ml staining solution (50 μg/ml PI, 20 μg/ml RNase
A). After incubation of 30 min at room temperature, samples were
analyzed using a FACSCalibur flow cytometer (BD Biosciences).
Flow cytometric analysis of
apoptosis
Annexin V and propidium iodide (PI) staining was
performed using an Annexin V-fluorescein isothiocyanate (FITC)
apoptosis detection kit (BD Biosciences) to measure apoptosis.
Cells (1×105) cultured in 60-mm dishes were washed twice
with ice-PBS, collected, and re-suspended in 100 μl 1X binding
buffer. Annexin V-FITC (5 μl) conjugate and 5 μl of PI buffer were
added, and the cells were incubated at room temperature for 15 min
in the dark. After addition of 400 μl of 1X binding buffer, the
cells were analyzed using a flow cytometer (PE and 7AAD were used
for 53BP1-knockdown HO-8910PM cells).
Western blot analysis
Whole-cell lysates of the cells were prepared by
incubating cells in RIPA buffer (Shennengbocai, Shanghai, China)
(1% NP-40, 0.1% SDS, 5 mM EDTA, 0.5% sodium deoxycholate, 1 mM
sodium vandate) containing protease inhibitors (1 mM PMSF, 1 mM
sodium fluoride). Cell lysates were centrifuged at 12,000 rpm for
15 min at 4°C. The supernatant was collected, and the protein
concentration was measured using the BCA protein assay kit (Merck,
Darmstadt, Germany). Proteins in equal amounts were separated by
appropriate concentration using SDS-polyacrylamide gel
electrophoresis and transferred onto PVDF membranes (Millipore,
Billeriaca, MA, USA). The membranes were blocked in TBST for 1 h at
room temperature and then incubated with primary antibodies
overnight at 4°C. The membranes were then washed three times with
TBST, followed by incubation with HRP-labeled secondary antibodies
(KPL, Gaithersburg, MD, USA) (1:5000). Bound antibody was
visualized using ECL detection reagent (Merck).
Statistical analysis
The SPSS version 14.0 software was used for
statistical analysis. Results were reported as means ± standard
deviation (SD). The Q-test was used to analyze statistical
differences between groups. The alpha (α) level was set at
0.05.
Results
Evaluation of 53BP1 protein expression in
ovarian cancer cells after transfection with the 53BP1-overpressing
and -knockdown plasmids
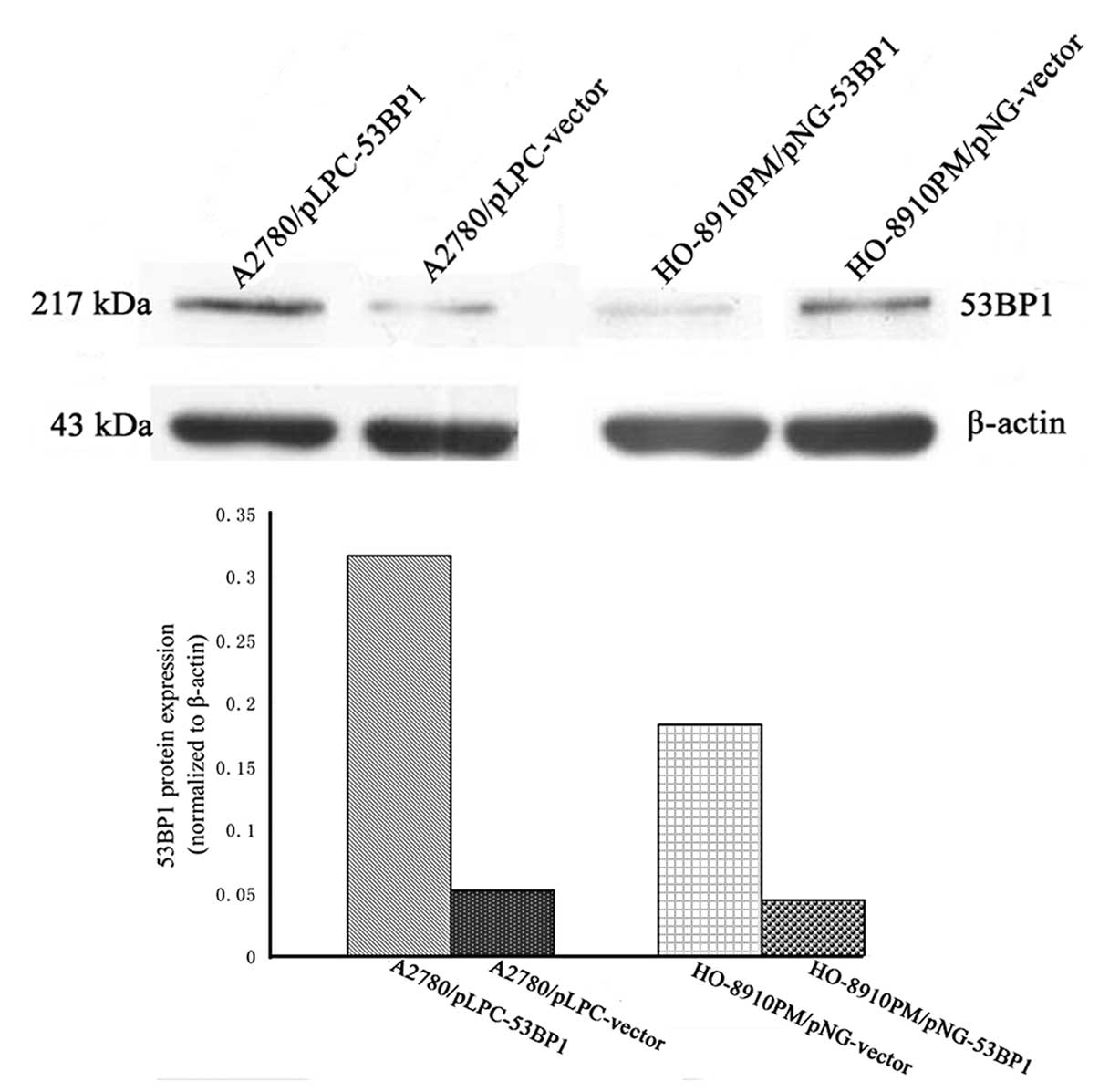
Stable transfected cell lines were generated by
expanding the resistant (G418 and puromycin, respectively)
colonies, including A2780/pLPC-vector, A2780/pLPC-53BP1 and
HO-8910PM/pNG-vector-transfected cells lines, and the expression of
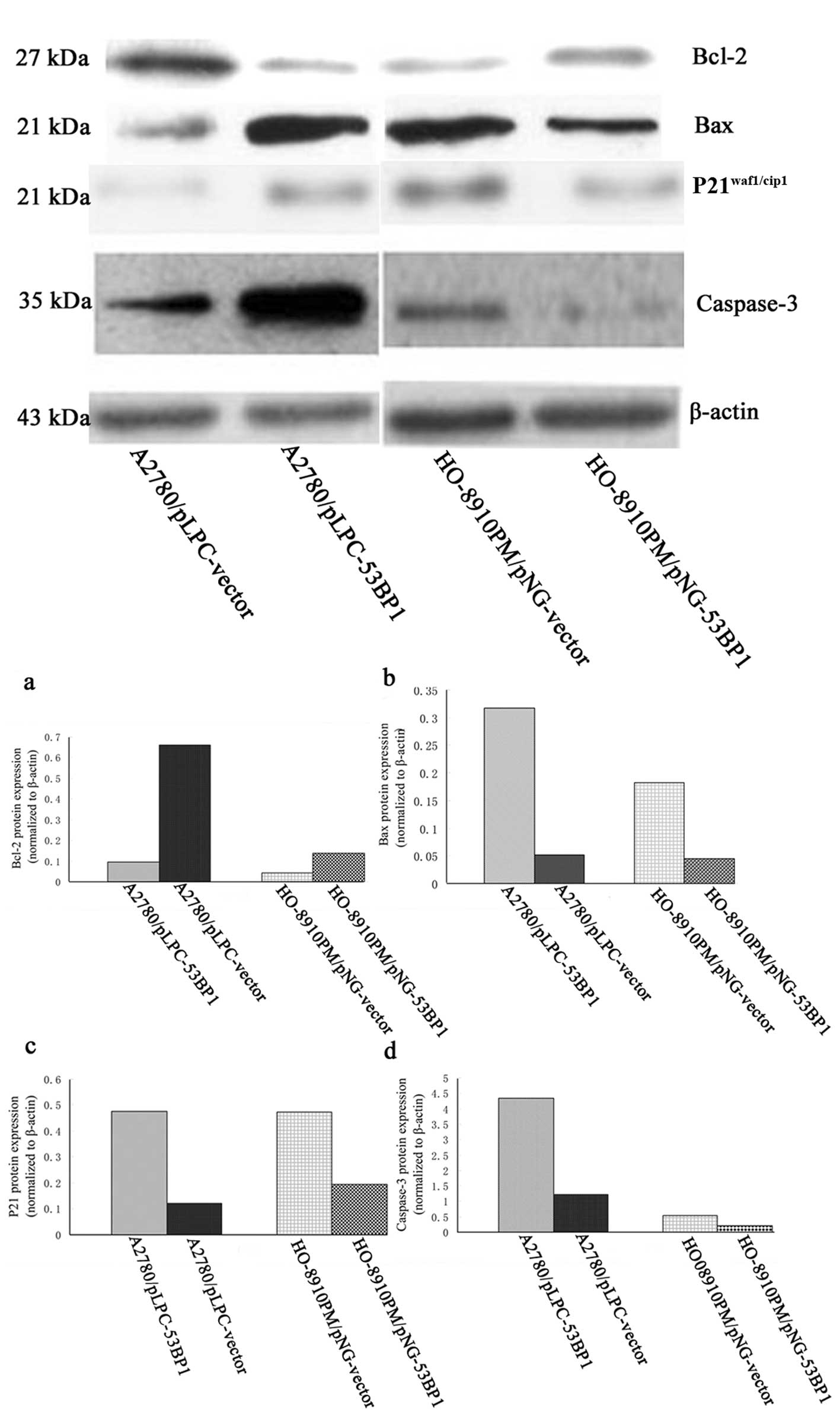
53BP1 in these cell lines was examined by western blot analysis. As
shown in Fig. 1, the level of 53BP1
was significantly increased and decreased compared with the
corresponding control group.
53BP1 ectopic expression markedly alters
cell proliferation in the derived cell lines of A2780 and
HO-8910PM
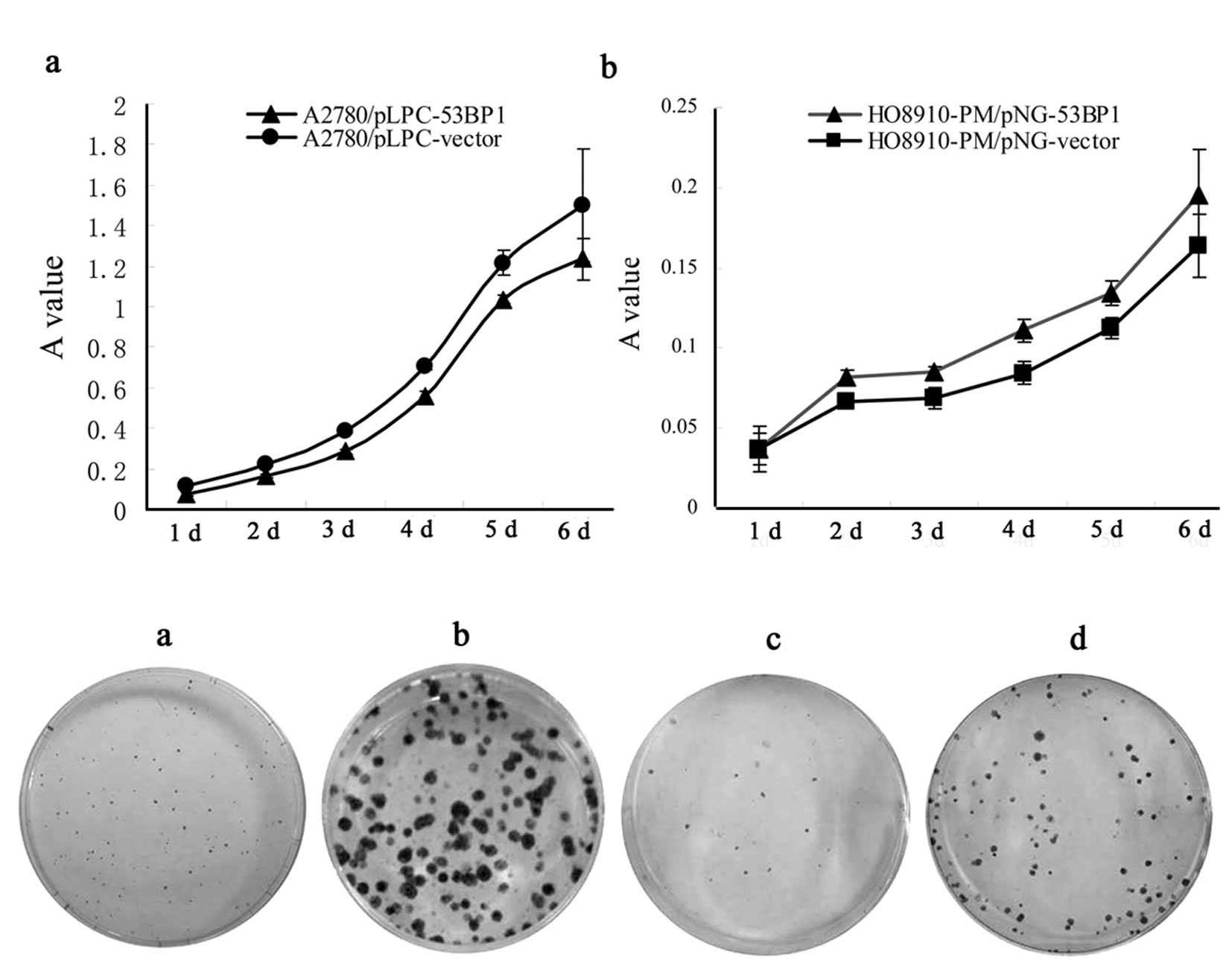
Using MTT, we investigated the effect of 53BP1 on
the proliferation of the A2780/pLPC-53BP1 cells, compared to the
A2780/pLPC-vector cells, and in the HO-8910PM/pNG-53BP1 cells
compared to the HO-8910PM/pNG-vector cells. As shown in Fig. 2A, A2780/pLPC-53BP1 and
HO-8910PM/pNG-vector cells consistently exhibited a significant
decrease in celluar proliferation compared to the A2780/pLPC-vector
and HO-8910PM/pNG-53BP1 cells, respectively (P<0.05).
Colony formation assay was performed on A2780 and
HO-8910PM cells, respectively. As far as congenetic cells, the
colony formation significantly increased with expression of 53BP1
and vice versa (Fig. 2B).
53BP1-mediated Akt activation contributes
to induce G2/M cell cycle arrest and apoptosis
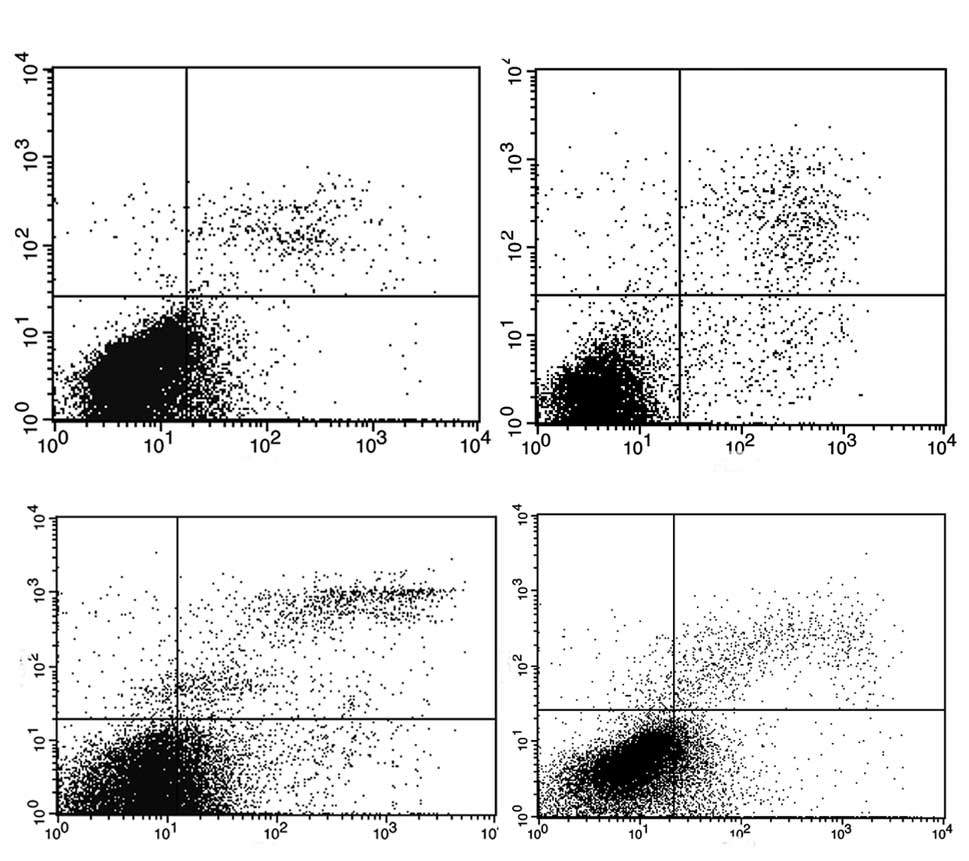
As compared to the empty vector, 53BP1 transfection
caused a highly dramatical cell cycle arrest in the G2/M
phase in A2780/pLPC-53BP1 cells (P<0.05). Similar results were
found in the HO-8910PM/pNG-vector and HO-8910PM/pNG-53BP1
(P<0.05) (Fig. 3).
Fig. 4 shows the
results of Annexin V/PI staining, which was used to quantify
apoptosis. The proportion of apoptotic cells in A2780/pLPC-53BP1
and A2780/pLPC-vector was ~20.01±1.10% and 7.89±0.23% (P<0.05),
respectively. Downregulation of 53BP1 in the HO-8910PM/pNG-53BP1
cells decreased the proportion of apoptosis (4.24±0.25%) compared
to that of the HO-8910PM/pNG-vector cells (10.15±1.1%,
P<0.05)
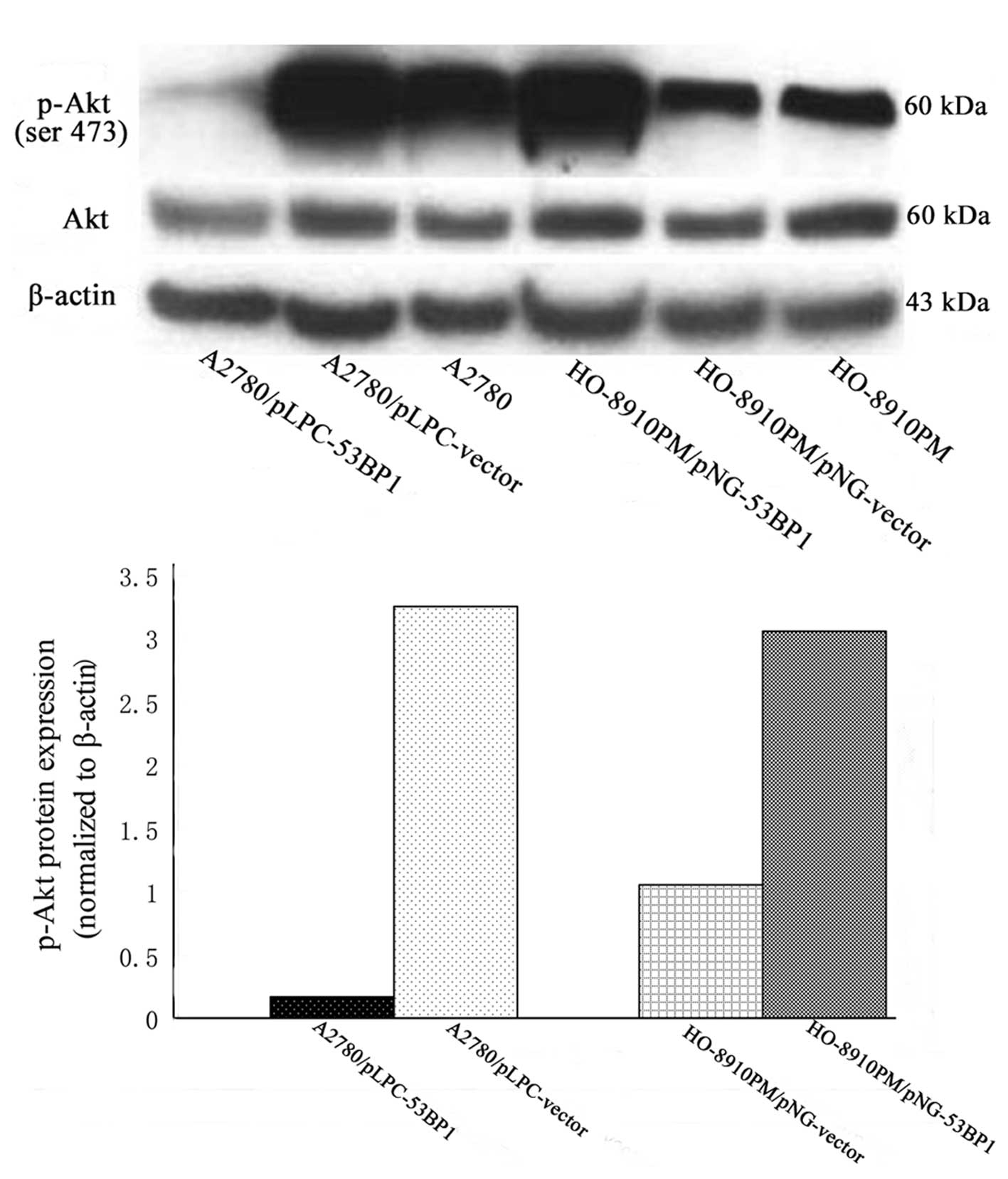
When the expression of 53BP1 was relatively high,
genes involved in G2/M phase transition and inhibition
of apoptosis such as P21waf1/Cip1, caspase-3 and Bax
were significantly upregulated (P<0.05), while, genes promoting
the ability of proliferation and apoptosis, such as p-Akt, Bcl-2
were downregulated (P<0.05) (Figs.
5 and 6).
Discussion
A healthy DNA-damage repair system is essential for
maintaining a harmonious stable relationship among apoptosis, cell
cycle checkpoints and cell proliferation. In contrast, a
dysfunctional DNA-damage repair system results in serious
consequence to cells: unlimited growth, resistance to apoptosis,
and tumor formation. The crucial roles of 53BP1 in DNA-damage
repair have been intensely debated. Studies have demonstrated that
mutation or loss of 53BP1 is linked to a variety of common human
cancers, including breast, lymphoma, and prostate cancer (11,13–15).
53BP1−/ − or 53BP1+/− greatly promotes
lymphoma-genesis in a p53-null background (16). This present study provides a new
paradigm for understanding the multiple signaling pathways involved
in 53BP1 as a candidate tumor-suppressor gene which regulates
ovarian cancer cell survival.
To determine whether 53BP1 plays a role in the
reduction of the proliferation rate of cells, we analyzed the
effect of the alteration of 53BP1 expression on the induction of
antiproliferation of ovarian cancer cells, A2780 and HO-8910PM. We
showed that the expression level of 53BP1 was inversely correlated
to the proliferation rate of ovarian cancer cells. This effect was
mostly based on the 53BP1-mediated downregulation of p-Akt
(phosphorylation-Akt), while, Akt protein levels exhibited no
significant change pre-and post-alteration of 53BP1. Akt is a
serine/threonine protein kinase and is predominantly localized to
the endoplasm (17). PIP3
(phosphatidylinositol 3,4,5-trisphosphate) synthesis is a result of
activation of PI3K (PI3-kinase) by growth factor stimulation of
cells. Subsequently, PIP3 triggers PDK1
(3-phosphoinositide-dependent kinase 1) to phosphorylate Akt
(18). Several lines of evidence
suggest that activated p-Akt serves as a multifunctional regulator
of cellular processes, including cell proliferation, cell cycle and
apoptosis (19–21). Research has shown that modulation of
the genetic background of cancer cells or therapeutic interventions
may induce G2/M arrest accompanied by the downregulation
of p-Akt (22–24).
There is a subtle relationship among growth,
apoptosis and cell cycle arrest of cancer cells. Cell cycle arrest
and apoptosis can induce the inhibition of proliferation in cancer
cells. Previous reports have revealed the influence of Akt on
proliferation, apoptosis and cell cycle. Herein, we investigated
the cell cycle profile, apoptosis and expression of related
signaling proteins by modulating the expression of 53BP1 in ovarian
cancer cells. We found that overexpression of 53BP1 significantly
promoted apoptosis, induced G2/M arrest and vice versa.
Furthermore, the protein levels of Bax, caspase-3 and
P21waf1/Cip1 were significantly increased, acompanied by
a decrease in Bcl-2.
Recent studies have shown that the PI3K/Akt pathway
is involved in the regulation of Bcl-2 family proteins (25,26).
The Bcl-2 family is an important apoptosis controller; Bcl-2 and
Bax are members of the Bcl-2 family, and they belong to
anti-apoptotic protein and pro-apoptotic protein, respectively. The
ratio of Bax/Bcl-2 has a great influence on determining whether
cells undergo apoptosis (27). In
our research, we found that 53BP1 induced apoptosis of ovarian
cancer cells by increasing pro-apoptotic Bax expression and
decreasing anti-apoptotic Bcl-2 expression, leading to
up-regulation of the ratio of Bax/Bcl-2. Although Akt plays an
important role in the mechanism by which 53BP1 exerts its
antiapoptosis effect, Akt is clearly not the only molecule
involved. Other potential targets, such as caspase-3, were found to
stimulate apoptosis. Our results confirmed the upregulation of
caspase-3 expression in A2780/pLPC-53BP1 and HO-8910PM/pNG-vector
cells compared to their control groups, A2780/pLPC-vector and
HO-8910PM/pNG-53BP1, respectively. Bax/Bcl-2 may have an effect on
caspase-3 by enhancing apoptosis of cancer cells by cytochrome c,
APAF-1, caspase-9 and other death substrates (28,29).
Yet, further studies are needed to confirm that other proteins are
involved in the anti-apoptosis caspase pathway in relation to the
effect of the alteration of 53BP1 in ovarian cancer cells.
Coincident with our results, recent studies have shown that the
downregulation of Akt has an impact on inducing G2/M
arrest (22–24). P21waf1/Cip1 is an
inhibitor of CDK (cyclin-dependent kinase) in mammalian cells and
the transcriptional target of P53 (30). The tumor suppressor protein
P21waf1/Cip1 serves to inhibit kinase activity and block
progression through G1/S (31), while P21waf1/Cip1 is
associated with the induction and maintenance of G2/M
cell cycle arrest (32). Akt was
found to lose the ability to play a negative role on the cell cycle
by directly phosphorylating Thr145 and Ser146 of
P21waf1/Cip1, and then the phosphorylated
P21waf1/Cip1 is unable to enter the nucleus from the
cytoplasm (33).
Overall, our results confirmed that 53BP1 inhibits
cell proliferation and induces G2/M arrest and
apoptosis. The antitumor effect of 53BP1 on ovarian cancer cells
possibly involves the down-regulation of the Akt signaling pathway.
These findings suggest a new target for tumors refractory to
current treatments.
Acknowledgements
The work was supported by the National Natural
Science Foundations of China to Beihua Kong (No. 30872738), and by
the Natural Science Foundation of Shandong Province of China to
Shuhui Hong (No. ZR2009CL015).
References
|
1
|
Wright JW, Pejovic T, Jurevic L, Bishop
CV, Hobbs T and Stouffer RL: Ovarian surface epitheliectomy in the
non-human primate: continued cyclic ovarian function and limited
epithelial replacement. Hum Reprod. 26:1422–1430. 2011. View Article : Google Scholar
|
|
2
|
Ovarian cancer: screening, treatment, and
followup. NIH Consens Statement. 12:1–30. 1994.
|
|
3
|
Fong MY, McDunn J and Kakar SS:
Identification of metabolites in the normal ovary and their
transformation in primary and metastatic ovarian cancer. PLoS One.
6:e199632011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Altomare DA, Menges CW, Xu J, et al:
Losses of both products of the cdkn2a/arf locus contribute to
asbestos-induced mesothelioma development and cooperate to
accelerate tumorigenesis. PLoS One. 6:e188282011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Iwabuchi K, Bartel PL, Li B, Marraccino R
and Fields S: Two cellular proteins that bind to wild-type but not
mutant p53. Proc Natl Acad Sci USA. 91:6098–6102. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lowndes NF: The interplay between BRCA1
and 53BP1 influences death, aging, senescence and cancer. DNA
Repair (Amst). 9:1112–1116. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mochan TA, Venere M, DiTullio RA and
Halazonetis TD: 53BP1 and NFBD1/MDC1-Nbs1 function in parallel
interacting pathways activating ataxia-telangiectasia mutated (ATM)
in response to DNA damage. Cancer Res. 63:8586–8591.
2003.PubMed/NCBI
|
|
8
|
Cescutti R, Negrini S, Kohzaki M and
Halazonetis TD: TopBP1 functions with 53BP1 in the G1 DNA damage
checkpoint. EMBO J. 29:3723–3732. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu PP, Chung HW, Liu KC, et al: Diallyl
sulfide induces cell cycle arrest and apoptosis in HeLa human
cervical cancer cells through the p53, caspase- and
mitochondria-dependent pathways. Int J Oncol. 38:1605–1613.
2011.PubMed/NCBI
|
|
10
|
Iwabuchi K, Matsui T, Hashimoto M,
Matsumoto Y, Kurihara T and Date T: Characterization of a cancer
cell line that expresses a splicing variant form of 53BP1:
separation of checkpoint and repair functions in 53BP1. Biochem
Biophys Res Commun. 376:509–513. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bartkova J, Horejsi Z, Sehested M, et al:
DNA damage response mediators MDC1 and 53BP1: constitutive
activation and aberrant loss in breast and lung cancer, but not in
testicular germ cell tumours. Oncogene. 26:7414–7422. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shenhua X, Lijuan Q, Hanzhou N, et al:
Establishment of a highly metastatic human ovarian cancer cell line
(HO-8910PM) and its characterization. J Exp Clin Cancer Res.
18:233–239. 1999.PubMed/NCBI
|
|
13
|
Takeyama K, Monti S, Manis JP, et al:
Integrative analysis reveals 53BP1 copy loss and decreased
expression in a subset of human diffuse large B-cell lymphomas.
Oncogene. 27:318–322. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nuciforo PG, Luise C, Capra M, Pelosi G
and d’Adda di Fagagna F: Complex engagement of DNA damage response
pathways in human cancer and in lung tumor progression.
Carcinogenesis. 28:2082–2088. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Du RZH, Chen J and Jiang ZM: Relationship
between loss of 53BP1 expression and clinical pathology in prostate
adenocarcinoma. Chin J Clin Exp Pathol. 1:44–47. 2011.
|
|
16
|
Ward IM, Difilippantonio S, Minn K, et al:
53BP1 cooperates with p53 and functions as a haploinsufficient
tumor suppressor in mice. Mol Cell Biol. 25:10079–10086. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dai RY, Chen SK, Yan DM, et al: PI3K/Akt
promotes GRP78 accumulation and inhibits endoplasmic reticulum
stress-induced apoptosis in HEK293 cells. Folia Biol (Praha).
56:37–46. 2010.PubMed/NCBI
|
|
18
|
Miao B, Skidan I, Yang J, et al: Small
molecule inhibition of phosphatidylinositol-3,4,5-triphosphate
(PIP3) binding to pleckstrin homology domains. Proc Natl Acad Sci
USA. 107:20126–20131. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Franke TF, Yang SI, Chan TO, et al: The
protein kinase encoded by the Akt proto-oncogene is a target of the
PDGF-activated phosphatidylinositol 3-kinase. Cell. 81:727–736.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hemmings BA: PH domains - a universal
membrane adapter. Science. 275:18991997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hemmings BA: PtdIns(3,4,5)P3 gets its
message across. Science. 277:5341997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu X, Zhang Y, Qu D, Jiang T and Li S:
Osthole induces G2/M arrest and apoptosis in lung cancer A549 cells
by modulating PI3K/Akt pathway. J Exp Clin Cancer Res. 30:332011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee DH, Thoennissen NH, Goff C, et al:
Synergistic effect of low-dose cucurbitacin B and low-dose
methotrexate for treatment of human osteosarcoma. Cancer Lett.
306:161–170. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yingchun L, Xiujuan Q, Jinglei Q, et al:
E3 ubiquitin ligase Cbl-b potentiates the apoptotic action of
arsenic trioxide by inhibiting the PI3K/Akt pathway. Braz J Med
Biol Res. 44:105–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kucharzewska P, Welch JE, Svensson KJ and
Belting M: The polyamines regulate endothelial cell survival during
hypoxic stress through PI3K/AKT and MCL-1. Biochem Biophys Res
Commun. 380:413–418. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang F, Zheng W, Pi R, et al:
Cryptotanshinone protects primary rat cortical neurons from
glutamate-induced neurotoxicity via the activation of the
phosphatidylinositol 3-kinase/Akt signaling pathway. Exp Brain Res.
193:109–118. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rondelet B, Dewachter C and Kerbaul F: et
al Prolonged overcirculation-induced pulmonary arterial
hypertension as a cause of right ventricular failure. Eur Heart J.
May 23–2011.(E-pub ahead of print).
|
|
28
|
Davies MA, Koul D, Dhesi H, et al:
Regulation of Akt/PKB activity, cellular growth, and apoptosis in
prostate carcinoma cells by MMAC/PTEN. Cancer Res. 59:2551–2556.
1999.PubMed/NCBI
|
|
29
|
Cardone MH, Roy N, Stennicke HR, et al:
Regulation of cell death protease caspase-9 by phosphorylation.
Science. 282:1318–1321. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu W, Konduri SD, Bansal S, et al:
Estrogen receptor-alpha binds p53 tumor suppressor protein directly
and represses its function. J Biol Chem. 281:9837–9840. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pestell RG, Albanese C, Reutens AT, Segall
JE, Lee RJ and Arnold A: The cyclins and cyclin-dependent kinase
inhibitors in hormonal regulation of proliferation and
differentiation. Endocr Rev. 20:501–534. 1999.PubMed/NCBI
|
|
32
|
Shin SY, Kim CG, Kim SH, Kim YS, Lim Y and
Lee YH: Chlorpromazine activates p21Waf1/Cip1 gene
transcription via early growth response-1 (Egr-1) in C6 glioma
cells. Exp Mol Med. 42:395–405. 2010.PubMed/NCBI
|
|
33
|
Zhou BP, Liao Y, Xia W, Spohn B, Lee MH
and Hung MC: Cytoplasmic localization of p21Cip1/WAF1 by
Akt-induced phosphorylation in HER-2/neu-overexpressing cells. Nat
Cell Biol. 3:245–252. 2001. View
Article : Google Scholar : PubMed/NCBI
|