Introduction
Lung cancer is one of the most common malignant
tumors worldwide and non-small cell lung cancer accounts for 80% of
all lung cancer cases. Thermotherapy is an approach taking
advantage of biological effects of heat to cure tumors.
Thermotherapy undergoes a systematic research with the medicine
development and equipment advancement (1). Now, thermotherapy is regarded as an
effective treatment such as surgery, radiotherapy, chemotherapy and
biological treatment (2). In recent
years, tumor thermotherapy has been widely used to treat cancer
(3), including lung cancer
(4). Tumor thermotherapy increased
tumor cell sensitivity to drugs in the effective temperature range,
reversed tumor cell drug-resistance and induce apoptosis (5).
Intracellular ROS could be induced and affected by
many factors such as heat, cell cytokines or some chemicals. Its
function is to signal to activate cellular apoptosis inducing cell
apoptosis pathway (6–9). However, UV radiation was found to be
able to lead to increased production of ROS and activation of
mitogen activated protein kinase (MAPK) pathway (10). As a member of MAPK family, protein
kinase B (PKB)/Akt plays key roles on regulation of cell growth and
apoptosis. Phosphorylated Akt (p-Akt) takes part in many
physiological and pathological processes directly or indirectly.
Activation of Akt pathway showed an obvious promotion on cell
growth and was necessary to normal cell growth (11,12).
In addition, activated Akt (13),
but inactivated cysteinyl aspartate-specific proteinases (caspase)
simultaneously were observed in most tumor cells, which might be
responsible for tumor occurrence and development (14). Caspase is the core regulatory
element in the formation of cell apoptosis regulating network and
could be activated by self-catalysis or caspase cascade catalysis.
Oxidative stress was able to induce elevated caspase-3 activation
and increased cell apoptosis (15)
through degradating more target proteins (16).
We observed that thermotherapy in combination with
chemotherapy showed stronger inhibitory effect on lung tumor cell
growth than thermotherapy or chemotherapy alone (17), the molecular mechanisms in the
process remained unclear.
Materials and methods
Cell culture and cell treatments
Human lung adenocarcinoma A549 cell line was
purchased from Shanghai Cell Biology Institute Cell Bank, (China)
and was cultured in F-12k cell supernatant (Sigma, St. Louis, MO,
USA) supplemented with 10% fetal calf serum (Sigma) at 37°C in a 5%
CO2 saturated humidity atmosphere. Logarithmic phase
cells were digested with 5% trypsin solution and the digestion was
inactivated in cell supernatant, and then was separated into single
cells. The cell density was diluted into 2.5×107 cells/l
and 6 ml of the cell suspension was added into each well of culture
plates. Then cells were cultured at 37°C in a 5% CO2
humidified incubator (Heraeus, Germany). After culturing for 24 h,
the cells were subjected to different group schemes as follows:
cells treated only by 50 μg/l paclitaxel (Taiji Group Sichuan Taiji
Pharmacy Co., Ltd., China; the approval number: GY H19994040) were
the chemotherapy group while cells treated with 50 μg/l paclitaxel
and 43°C heat were the thermo-chemotherapy group. Cells in
wortmannin (Alexis, USA) group were treated by thermo-chemotherapy
combined with 1 μmol/l wortmannin and cells in NAC (Sigma) group
were treated by thermo-chemotherapy combined with 30 μmol/l NAC.
The cells without any treatment were regarded as controls.
Hyperthermic treatments were performed by immersing either the
culture flasks or parafilm-wrapped 96-well plates in a waterbath at
43°C for 90 min.
MTT assay
Single cell suspension was prepared with the cells
in logarithmic phase and cell density was adjusted to
2.5×107 cells/l. Then this cell suspension was seeded
into 96-well plates, 200 μl per well, and cultured at 37°C in a 5%
CO2 humidified incubator. After 24 h, cells were treated
according to different schemes described above (6-wells were
assayed for each group and repeated three times). The treated cells
were incubated for 4 h and 20 μl of the solution containing 5 g/l
MTT (Sigma) was added to each well and incubation for another 4 h
was performed. MTT was discarded and 150 μl dimethylsulfoxide
(DMSO) (Takara, China) was added to each well, and then vortexed
for 10 min to dissolve the crystal. DMSO was used as negative
control. Absorbance (A) value was measured at 492 nm with the
microplate reader (Sunrise China). Cell proliferation rates were
calculated according to the following formula: cell proliferation
rate (%) = (average A value of experimental group/average A value
of control group) × 100%.
Intracellular ROS detection
After the treatment was performed, about
1×106 cells in the logarithmic growth phase were
collected, washed by PBS and centrifuged at 1200 r/min for 5 min.
Then reagent was added and cultured at 37°C for 20 min.
Intracellular ROS was detected using Reactive Oxygen Species
Testing kit (Shanghai Jiemei Gene Medicine Technology Co., Ltd.,
China). Preservative medium was added and detected at 490 nm
(excitation) and 530 nm (emission) qualitatively by fluorescence
microscope (Olympus, Japan) and quantitatively by F-4500
fluorospectrophotometer (Hitachi, Japan).
Western blotting
Cells in logarithmic phase (1×106) (in
vitro) or 50 mg tissue (in vivo) from each group were
harvested and suspended in cell lysis buffer containing 30 mmol/l
NaHCO3, 0.1 mmol/l EDTA, 0.1 mmol/l DTT, 0.5 mmol/l PMSF
and 2 mg/l aprotinin. Cells were lysed on ice and centrifuged at
10,000 r/min and 4°C for 10 min. The supernatant and loading buffer
was mixed. Equal amount of total protein dissolved in the mixture
of each sample was loaded and seperated by sodium dodecylsulfate
polyacrylamide gel electrophoresis (SDS-PAGE) (Bio-Rad
Laboratories, Hercules, CA, USA) and transferred onto
nitrocellulose membrane (Beijing Applygen Technology Ltd., Beijing,
China) for antibody blotting. The membrane was incubated with a
rabbit monoclonal antibody against Akt (Cell Signaling Technology,
Inc., Danvers, MA), a rabbit monoclonal antibody against p-Akt
(Ser473) (587F11) (Cell Signaling Technology, Inc.), a rabbit
monoclonal antibody against caspase-3 (Cell Signaling Technology,
Inc.) or a rabbit monoclonal antibody against β-actin (Cell
Signaling Technology, Inc.) overnight at 4°C, respectively. After
being incubated with goat anti-rabbit IgG antibodies (1:300
dilution) (Beijing Zhongshan Jinqiao Biology Technology Co., Ltd.,
Beijing, China) for 2 h. The membrane was subjected to immunoblot
analysis and the enhanced chemiluminescence method (ECL kit, China)
and HRP-DAB kit (Tiangen, Biotech Co., Ltd., Beijing, China) were
used to visualize the proteins. β-actin was used as an internal
control. Gel imaging system (Gene Genius, USA) was used to capture
images. Semi-quantitative values of p-Akt and caspase-3 were
measured by the ratio of the bands of t-Akt and β-actin.
Cell apoptosis detection
After cell concentration was adjusted, cells were
seeded into 50-ml dishes to culture, and then they were treated
according to schemes mentioned in cell culture and cell treatments.
Cells in each group were suspended as single cells. A total of
1×106 cells were collected to tubes, and fixed in
ice-cold 70% ethanol for 12 h at 4°C. The ethanol was discarded and
cells were digested by 1 ml 0.5% typsin (pH 1.5–2.0) for 10 min and
washed with PBS, labeled with propidum iodide (PI) and filtered by
500-mesh screen to obtain single cell suspension and analyzed
directly by FACS 420 flow cytometry (B&D Systems, USA).
Graft inhibition rates in nude mice
Thirty BALB/c-nu/nu nude mice (purchased from
Shanghai Slack Experimental Animals Co., Ltd., China) were treated
according to the following schemes: chemotherapy group (50 μg/kg
paclitaxel intraperitoneal injection), thermo-chemotherapy group
(50 μg/kg paclitaxel intraperitoneal injection with 43°C
waterbath), wortmannin group (thermo-chemotherapy combined with
wortmannin intraperitoneal injection), NAC group
(thermo-chemotherapy combined with NAC intraperitoneal injection)
and controls (equal saline intraperitoneal injection without any
other treatments). There were 6 nude mice in each group and every
nude mouse was treated once a week for 3 weeks. Nude mice were
sacrificed 2 days after the last treatment. Changes in behavior,
eating, defecation and weight were tracked. When grafts were
removed, graft inhibition rates were calculated according to the
following formula: graft inhibition rates (%) = (graft quality in
control group - graft quality in experimental group)/graft quality
in control group × 100%. The institutional committee for animal
research approved these experiments.
Statistical analysis
The software of SPSS 13.0 was employed for data
analysis and results are shown as mean ± SD. Comparisons among
groups were analyzed with a one-way ANOVA and LSD method was
applied for multiple comparisons. P<0.05 was regarded as
significant.
Results
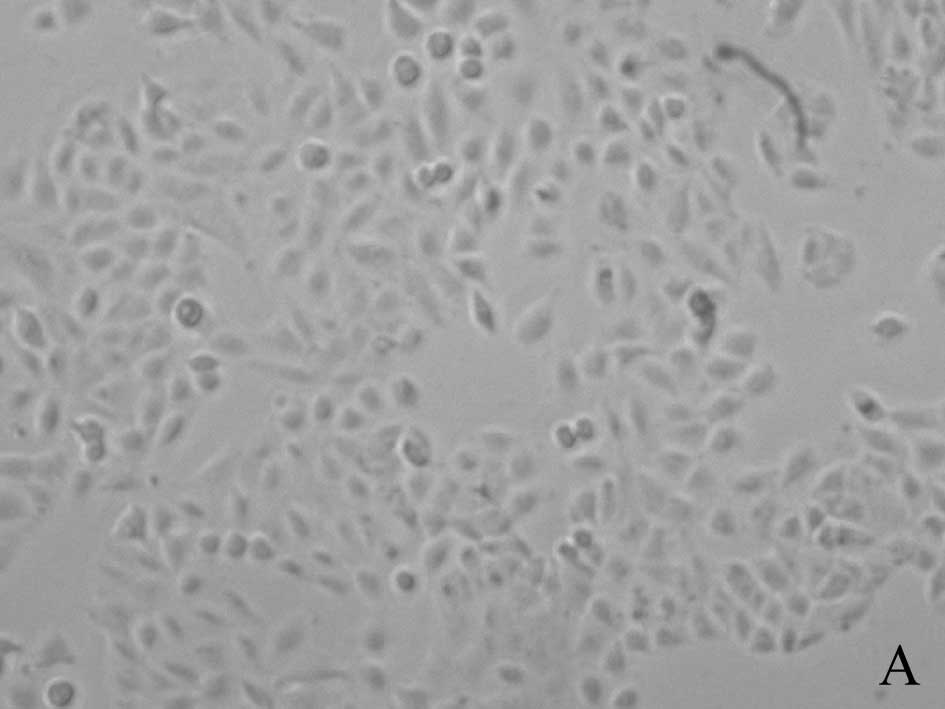
Cell proliferation rates
A549 cells in control group grew normally in
fusiform or polygonal shape. In chemotherapy group, the number of
A549 cells became less and many cells were detached from the flask
wall. Many cells were in division phase and cell bodies became
round and small. In thermo-chemotherapy group, cell morphology was
in the shape described as in chemotherapy group, but more cells
died. After wortmannin was added, the number of cells became less
but most cells were alive after NAC was added (Fig. 1).
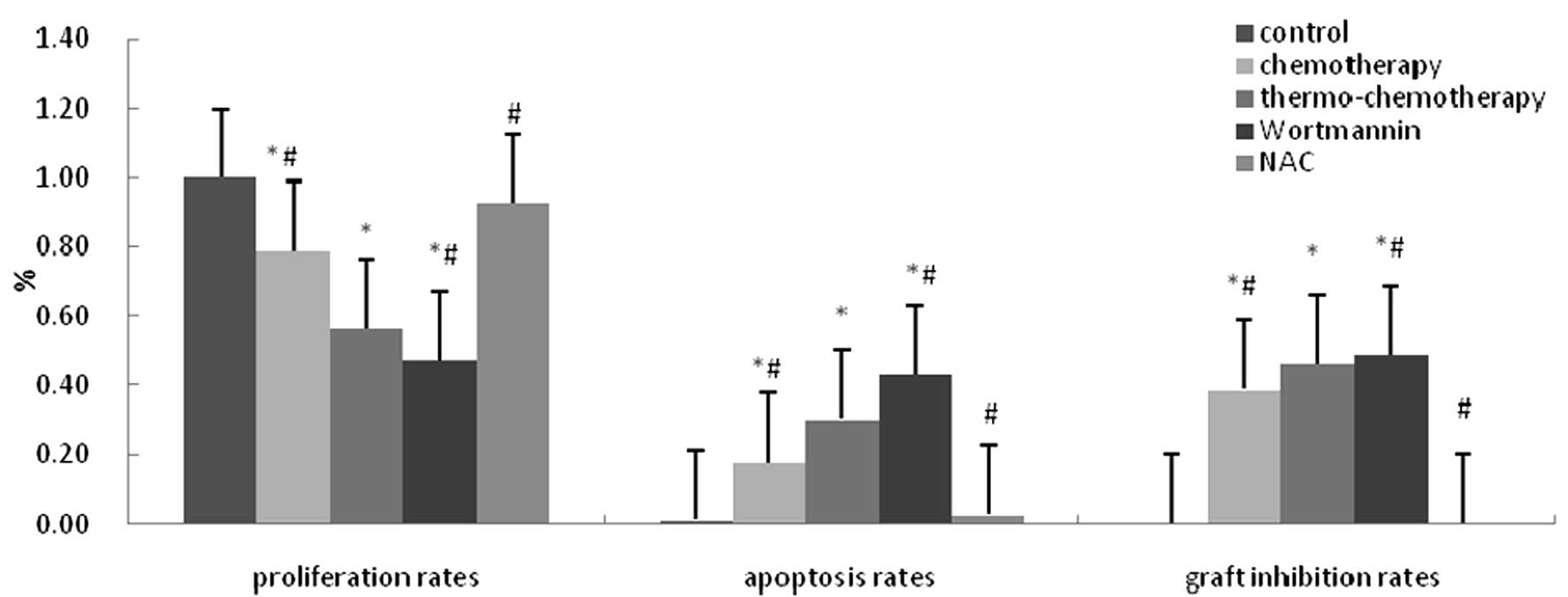
Compared with control group (100.00±0.00%),
significantly decreased proliferation rate was detected in the
groups treated with chemotherapy (78.77±2.38%) and
thermo-chemotherapy (56.34±4.30%) (P<0.05) and proliferation
rate in thermo-chemotherapy group was significantly lower than in
other groups (P<0.05). Proliferation rate in wortmannin group
(47.40±0.01%) was significantly lower than that in
thermo-chemotherapy group (P<0.05) and there was no significant
difference between NAC group (92.65±0.09%) and control group
(P>0.05), which indicated that thermotherapy in combination with
chemotherapy could inhibit A549 cells growth distinctly and ROS and
Akt pathways played a role inhibited by their own inhibitors
(Fig. 2).
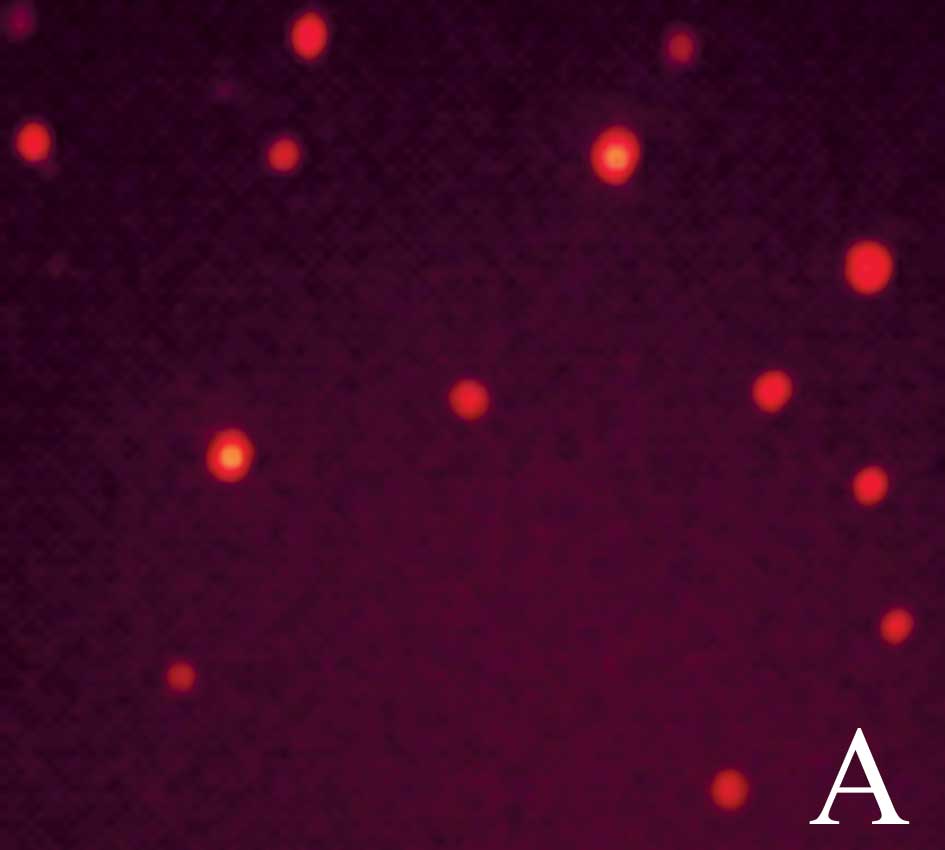
Changes of intracellular ROS
Cells in control group were red, which showed there
was no ROS in them (Fig. 3A); cells
in chemotherapy group were green, which indicated ROS in them
(Fig. 3B); cells in
thermo-chemotherapy group were bright green showing increased ROS
(Fig. 3C). Wortmannin had no effect
on ROS produced by thermo-chemotherapy (Fig. 3D), but after NAC was added, cells
became red (Fig. 3E).
Compared with thermo-chemotherapy group
(139.93±37.59%), significantly decreased ROS production was
detected in control group (58.07±2%), chemotherapy group
(74.40±3.01%) and NAC group (52.11±1.05%) (P<0.05) and
wortmannin had no effect on ROS (126.48±8.76%) (P>0.05),
indicating that thermo-chemotherapy induced production of ROS and
ROS was upstream of the Akt pathway (Fig. 3F).
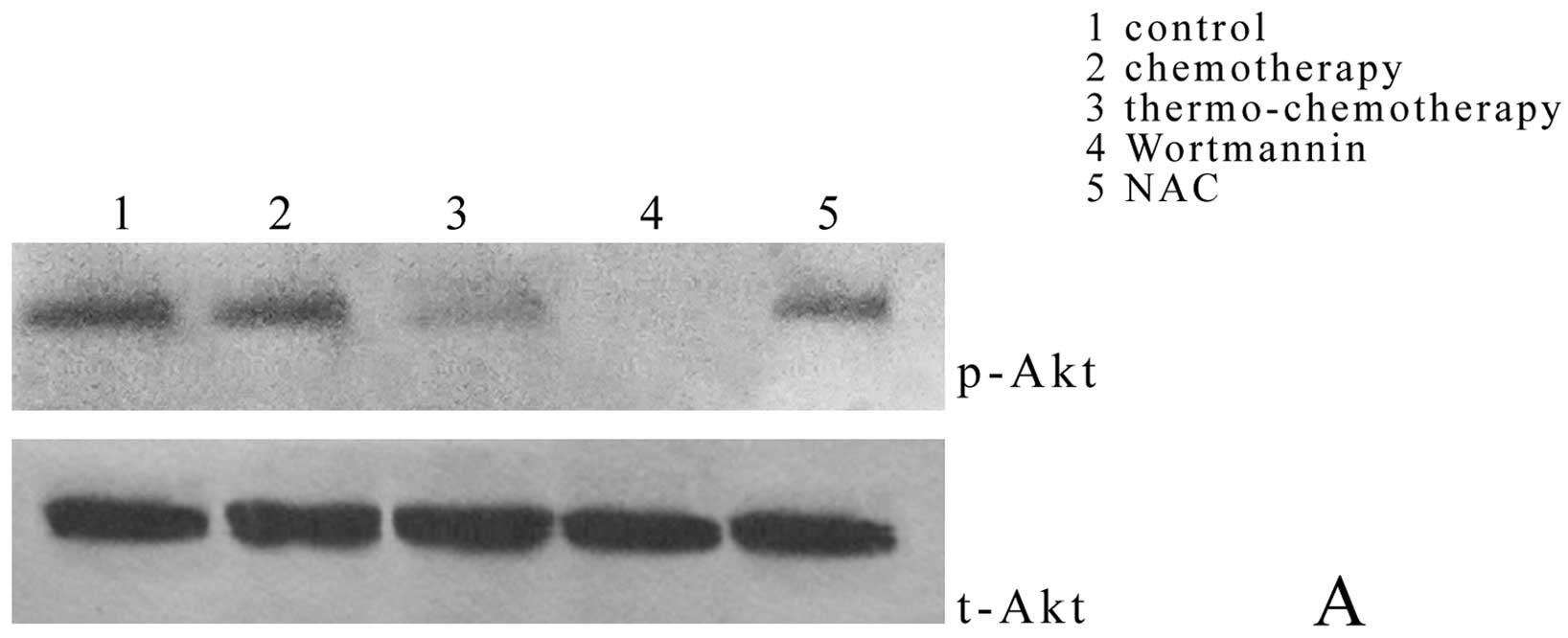
Effect of thermo-chemotherapy on
phosphorylation of Akt and caspase-3 expression
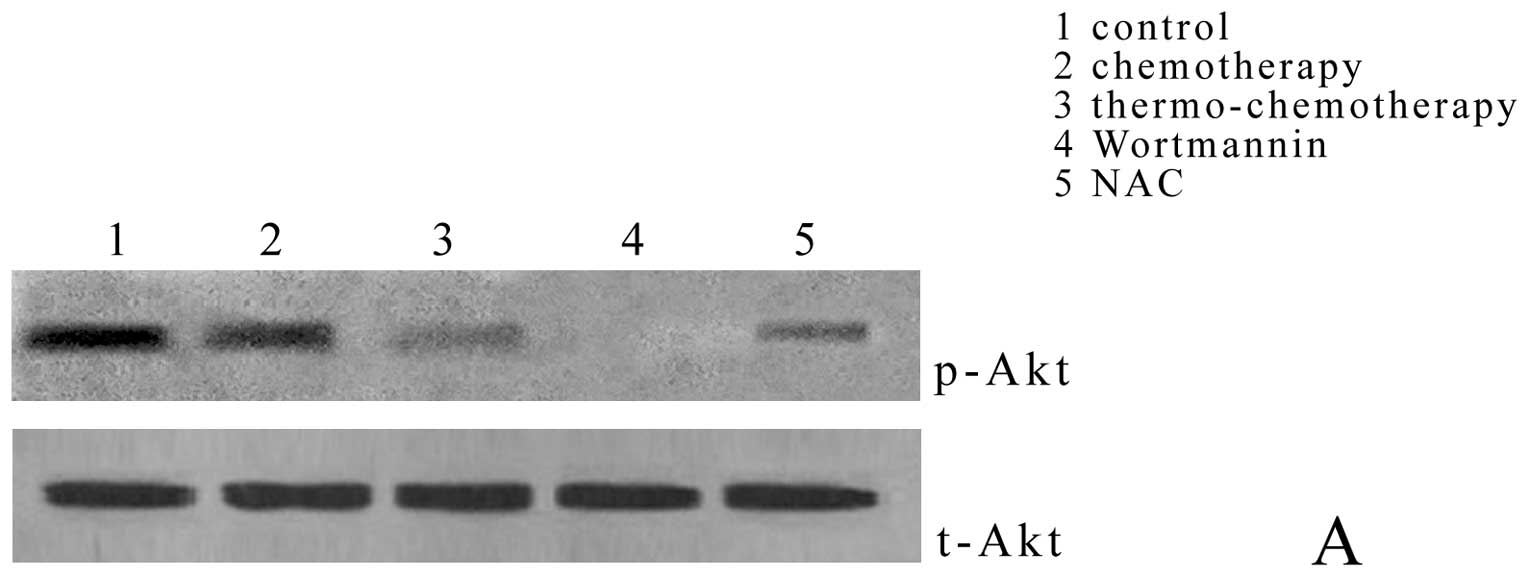
Compared with thermo-chemotherapy group
(0.63±0.00%), Akt phosphorylation in control group (1.62±0.03%) and
chemotherapy group (1.01±0.41%) decreased significantly (P<0.05)
and wortmanin completely inhibited Akt phosphorylation (0.00±0.00%)
(P<0.05); but when NAC, the specific inhibitor of ROS, was
added, Akt phosphorylation (1.08±0.24%) was significantly higher
than that in thermo-chemotherapy group (P<0.05) (Fig. 4A and C).
Caspase-3 expression in thermo-chemotherapy group
(1.02±0.02%) was significantly higher than that in control group
(0.48±0.11%) and chemotherapy group (0.67±0.02%) (P<0.05); after
Akt specific inhibitor wortmannin pretreatment, caspase-3
expression (1.05±0.09%) was significantly higher than that in
thermo-chemotherapy group (P<0.05); NAC could inhibited
caspase-3 expression completely (0.00±0.00%) (P<0.05) (Fig. 4B and C) showing that ROS could
inhibit activation of Akt pathway and activate the caspase pathway.
The Akt pathway was upstream of caspase pathway in
thermo-chemotherapy.
Cell apoptosis
Cell apoptosis rate in control group was
(1.22±0.18%) and significantly increased cell apoptosis rates were
observed in chemotherapy group (17.70±3.23%) and
thermo-chemotherapy group (30.39±2.89%) (P<0.05). Cell apoptosis
rate in thermo-chemotherapy group was significantly higher than
that in chemotherapy group (P<0.05). Compared with
thermo-chemotherapy group, cell apoptosis rate in wortmannin group
(43.23±9.26%) increased significantly and decreased significantly
in NAC group (2.80±0.99%) (P<0.05) (Fig. 2). Thermotherapy in combination with
chemotherapy inhibited A549 cell growth by leading to apoptosis
through induction of ROS production and subsequent inhibition of
Akt phosphorylation and activation of caspase.
Results of tests in vivo
A549 cells were injected subcutaneously into nude
mice and grafts occurred about 1 week later, with round or oval
shape. Compared with thermo-chemotherapy group (46.34±1.03%), graft
inhibition rate in wortmannin group (48.97±0.11%) increased
significantly, and decreased significantly in the control group
(0.00±0.00%), chemotherapy group (38.76±1.83%) and NAC group
(0.05±0.01%) (P<0.05), but there was no significant difference
between control group and NAC groups (P>0.05) (Fig. 2).
Akt phosphorylation in thermo-chemotherapy group
(0.66±0.06%) decreased significantly when compared with control
group (1.66±0.06%) (P<0.05) and increased significantly in
chemotherapy group (1.06±0.04%) and NAC group (1.07±0.02%) when
compared with thermo-chemotherapy group (P<0.05), but was
inhibited completely in wortmannin group (0.00±0.00%) (P<0.05)
(Fig. 5A and C).
Caspase-3 expression in thermo-chemotherapy group
(1.01±0.01%) was significantly higher than that in control group
(0.49±0.10%) and in chemotherapy group (0.66±0.03%) (P<0.05);
Caspase-3 expression in wortmannin group (1.06±0.07%) increased
significantly (P<0.05) and caspase-3 expression in NAC group
(0.00±0.00%) was inhibited completely (P<0.05) (Fig. 5B and C). All these results in cells
were authenticated in vivo.
Discussion
Thermotherapy is a new treatment of tumors, which
plays an important role in comprehensive treatment of tumors.
Thermotherapy in combination with chemotherapy is able to improve
tumor treatment efficiency (18).
Initially thermotherapy was regarded as ‘green treatment’ by the
clinicians (19). In 1990s, some
investigators paid attention to the phenomenon of cell apoptosis
induced by heat. This study shows that thermotherapy in combination
with chemotherapy could increase injury of drugs on tumor cells by
increasing ROS production, which leads to Akt activation, and
apoptosis of A549 cells through caspase cascade, which was
consistent with observation in vivo.
Normally, production and clearance of ROS is in a
dynamic balance (20). Cell growth
needs some ROS and ROS causes tissue cell apoptosis or necrosis
(21). When ROS balance was broken,
more ROS would be produced and oxidation stress occurred, which
would induce internal Ca2+ flow, unregulating the
expression of bax and activating caspase, then leading to cell
apoptosis (22,23). ROS was induced in the process of
tumor thermotherapy (24). The
results of this study show that thermotherapy in combination with
drugs would change induction and effect of ROS on tumor cells,
which provides a new approach to treat tumors. This study finds
that thermotherapy could increase production of intracellular ROS,
which is an important factor of apoptosis. It is also found that
this induction of ROS caused by thermotherapy could be inhibited by
the ROS inhibitor NAC, but not by the PI3K inhibitor, wortmannin
(Fig. 3). Therefore, ROS is likely
to be upstream of inhibition on Akt pathyway activation.
PI3K (phosphoinositide 3 kinase) is a phosphatidyl
kinase and induced tumor cell hyperplasia by activating
apoptosis-related Akt pathway (25). Its anti-apoptosis function may have
related to caspase activation. Phosphorylation of Akt could resist
apoptosis in chemotherapy and radiotherapy (26). Previous studies (27) have showed that phosphorylation of
Akt was increased from normal cells to atypia then to malignant
transformation with molecular markers related to loss of apoptosis
indicating that Akt extended survival of tumor cells and inhibited
apoptosis. Noske et al (28)
found that there was overexpression of Akt in 58% primary ovarian
cancer and RNA interference inhibited phosphorylation of Akt to
inhibit ovarian cancer cell proliferation. In this study, p-Akt is
highly-expressed in A549 cells and heat can decrease levels of Akt
phosphorylation. Also, p-Akt could be inhibited by NAC and
wortmannin (Figs. 4 and 5). Results of MTT and FCM show that cell
proliferation rates decreased and apoptosis rates increased in
thermo-chemotherapy group. Therefore, inhibition of thermotherapy
on lung tumor cell growth was associated with Akt pathway
activation.
In practice, wortmannin could inhibit
phosphorylation of Akt both in vivo and in vitro
(29). In our study, levels of
p-Akt in thermo-chemotherapy group were significantly lower than
those in control group and chemotherapy group. Wortmannin
completely inhibited phosphorylation of Akt (Figs. 4 and 5). Combined with results of MTT and FCM,
cell apoptosis rates in wortmannin group are higher than those in
thermo-chemotherapy group, which shows that thermotherapy inhibited
Akt pathway activation and NAC activated the Akt pathway.
Wortmannin can increase antitumor effects of thermotherapy at the
same time.
Caspases are a group of cysteine proteases, which
have similar amino acid sequences, and play an important role in
cell apoptosis. Caspase family enzymes regulate cell apoptosis.
Many apoptosis factors induce apoptosis by caspase-3 mediated
signal pathway (30). In this
study, heat could induce caspase-3 activation in A549 cells and NAC
could inhibit caspase-3 expression in heated A549 cells (Figs. 4 and 5), which suggests that production of ROS
was upstream of caspase-3 activation in A549 cell apoptosis induced
by heat. When heat inhibited phosphorylation of Akt, the expression
of caspase-3 increased, which shows that inhibition of Akt pathway
activation was upstream of caspase-3 activation.
In summary, thermotherapy could block the cell cycle
and improved induction of cell apoptosis (31). Cell apoptosis is one characteristic
of cell life and an active death form conducted by genes.
Inhibition of PI3K/Akt pathway activation could increase tumor cell
apoptosis induced by drugs (32).
Cell apoptosis rate in wortmannin group increased. But level of
p-Akt increased while cell apoptosis rate decreases after NAC
addition, which suggests that ROS inhibited activation of Akt
pathway and induced cell apoptosis. Inhibition of caspase-3
activity or antagonism of its function may inhibit cell apoptosis,
indicating that caspase-3 is essential to cell apoptosis (33). Thus, thermotherapy in combination
with chemotherapy showed a stronger inhibitory effect than
chemotherapy alone on A549 cell growth, probably through induction
of ROS production and subsequent inhibition of Akt phosphorylation
to activate caspase cascade, which would lead to lung cancer cell
apoptosis. All results were confirmed in vivo. These results
provide a theoretical basis for thermotherapy clinical
practice.
Acknowledgements
This study is supported by the National Natural
Science Foundation of China (nos. 30571552 and 30972457).
References
|
1
|
Yin WB, Yu ZH, Xu GZ and Hu YM: Radiation
Oncology. Fourth Edition. Peking Union Medical College Press; pp.
307–321. 2008
|
|
2
|
Lu H and Chen LB: Hyperthermia combined
with other therapies for cancer therapy. Postgrad Med J.
17:458–460. 2004.
|
|
3
|
Verwaal VJ, van Tinteren H, Ruth SV and
Zoetmulder FA: Toxicity of cytoreductive surgery and hyperthermic
intra-peritoneal chemotherapy. J Surg Oncol. 85:61–67. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bakhshandeh A, Bruns I, Traynor A, et al:
Ifosfamide, carboplatin and etoposide combined with 41.8 degrees C
whole body hyperthermia for malignant pleural mesothelioma. Lung
Cancer. 39:339–345. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang JH, Yao MZ, Zhang ZL, Zhang YH, Wang
YG and Liu XY: HSF1 blockade-induced tumor thermotolerance
abolishment is mediated by JNK-dependent Caspase-3 activation.
Biochem Biophys Res Commun. 321:736–745. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pletjushkina OY and Fetisova EK: Hydrogen
peroxide produced inside mitochondria takes part in cell-to-cell
transmission of apoptotic signal. Biochemistry. 71:60–67.
2006.PubMed/NCBI
|
|
7
|
Cheng Y, Chang LW and Tsou TC:
Mitogen-activated protein kinases mediate arsenic-induced
down-regulation of survivin in human lung adenocarcinoma cells.
Arch Toxicol. 80:310–318. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hamdi M, Kool J, Cornelissen-Steijger P,
et al: DNA damage in transcribed genes induces apoptosis via the
JNK pathway and the JNK-phosphatase MKP-1. Oncogene. 24:7135–7144.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vizio B, Poli G, Chiarpotto E and Biasi F:
4-hydroxynonenal and TGF-beta1 concur in inducing antiproliferative
effects on the CaCo-2 human colon adenocarcinoma cell line.
Biofactors. 24:237–246. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wen KC, Shih IC, Hu JC, Liao ST, Su TW and
Chiang HM: Inhibitory effects of terminalia catappa on UVB-induced
photodamage in fibroblast cell line. Evid Based Complement Alternat
Med. 2011:9045322011.PubMed/NCBI
|
|
11
|
Shi XY, Cai XJ, Lei JX, Cao FJ, Pan DF and
Chen P: Reversal effect of PI-3K/Akt pathway inhibitor LY294002 on
multidrug resistance of ovarian cancer cell line A2780/Taxol. Ai
Zheng. 27:343–347. 2008.(In Chinese).
|
|
12
|
Manning BD and Cantley LC: AKT/PKB
signaling: navigating downstream. Cell. 129:1261–1274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang J, Miao LJ, Wu YM, Wu YJ and Wang XC:
Expression of AKT2, cyclin D1, and MMP-9 and their correlations to
clinicopathologic features of non-small cell Lung cancer. Ai Zheng.
25:69–72. 2006.(In Chinese).
|
|
14
|
Zhuang QY, Chen XG, Dong ZQ, Liu JH and Ye
ZQ: Effects of rapamycin on prostate cancer PC-3 cells. Ai Zheng.
28:851–855. 2009.(In Chinese).
|
|
15
|
Lee MJ, Chen HM, Tzang BS, Lin CW, Wang
CJ, Liu JY and Kao SH: Ocimum gratissimum aqueous extract protects
H9c2 myocardiac cells from H(2)O(2)-induced cell apoptosis through
Akt signalling. Evid Based Complement Alternat Med.
2011:5780602011.PubMed/NCBI
|
|
16
|
Johnson GL and Lapadat R:
Mitogen-activated protein kinase pathways mediated by ERK, JNK and
p38 protein kinases. Science. 298:1911–1912. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gao S, Wang L, Wu WD and Zhou F:
Synergistic effect of thermotherapy in combination with
chemotherapy on lung tumor A549 cells growth through activation of
c-Jun N-terminal kinase and inhibition of heat shock protein70
expression. Wei Sheng Yan Jiu. 37:529–532. 2008.(In Chinese).
|
|
18
|
Patel S, Sanborn RE and Thomas CR Jr:
Definitive chemoradiotherapy for non-small cell lung cancer with N2
disease. Thorac Surg Clin. 18:393–401. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hiraoka M, Masunaga S, Nishimura Y, et al:
Regional hyperthermia combined with radiotherapy in the treatment
of lung cancers. Int J Radiat Oncol Biol Phys. 22:1009–1014. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan W, Arai A, Aoki M, Ichijo H and Miura
O: ASK1 is activated by arsenic trioxide in leukemic cells through
accumulation of reactive oxygen species and may play a negative
role in induction of apoptosis. Biochem Biophys Res Commun.
355:1038–1044. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Alexandre J, Batteux F, Nicco C, et al:
Accumulation of hydrogen peroxide is an early and crucial step for
Paclitaxel-induced tumor cell death both in vitro and in vivo. Int
J Cancer. 1:41–48. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jo PG, Choi YK and Choi CY: Cloning and
mRNA expression of antioxidant enzymes in the pacific oyster,
crassostrea gigas in response to cadmium exposure. Comp Biochem
Physiol C Toxicol Pharmacol. 147:460–469. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cho SD, Li G, Hu H, et al: Involvement of
c-Jun N-terminal kinase in G2/M arrest and caspase-mediated
apoptosis induced by sulforaphane in DU145 prostate tumor cells.
Nutr Cancer. 52:213–224. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hirano H, Tabuchi Y, Kondo T, et al:
Analysis of gene expression in apoptosis of human lymphoma U937
cells induced by heat shock and the effects of alpha-phenyl
N-tert-butylnitrone (PBN) and its derivatives. Apoptosis.
10:331–340. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Foster K, Wang Y, Zhou D and Wright C:
Dependence on PI3K/Akt signaling for malignant rhabdoid tumor cell
survival. Cancer Chemother Pharmacol. 63:783–791. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chin YR and Toker A: Function of Akt/PKB
signaling to cell motility, invasion and the tumor stroma in
cancer. Cell Signal. 21:470–476. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kobayashi I, Semba S, Mat suda Y, Kuroda Y
and Yokozaki H: Significance of Akt phosphorylation on tumor growth
and vascular endothelial growth factor expression in human gastric
carcinoma. Pathobiology. 73:8–17. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Noske A, Kaszubiak A, Weichert W, et al:
Specific inhibition of AKT2 by RNA interference results in
reduction of ovarian cancer cell proliferation: increased
expression of AKT in advanced ovarian cancer. Cancer Lett.
246:190–200. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tsurutani J, West KA, Sayyah J, Gills JJ
and Dennis PA: Inhibition of the phosphatidylinositol
3-kinase/Akt/mammalian target of rapamycin pathway but not the
MEK/ERK pathway attenuates laminin-mediated small cell lung cancer
cellular survival and resistance to imatinib mesylate or
chemotherapy. Cancer Res. 65:8423–8432. 2005. View Article : Google Scholar
|
|
30
|
Okun I, Balakin KV, Tkachenko SE and
Ivachtchenko AV: Caspase activity modulators as anticancer agents.
Anticancer Agents Med Chem. 8:322–341. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vertrees RA, Das GC, Popov VL, et al:
Synergistic interaction of hyperthermia and gemcitabine in lung
cancer. Cancer Biol Ther. 4:1144–1153. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dogra C, Changotra H, Wergedal JE and
Kumar A: Regulation of phosphatidylinositol 3 kinase (PI3K)/Akt and
nuclear factor kappa B signaling pathways in dystrophin-deficient
skeletal muscle in response to mechanical stretch. J Cell Physiol.
208:575–585. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zheng XW, Li Y, Tang FA, Ma J, Zheng PY
and Lu GF: In vivo antitumor effect of canstatin gene on human
esophageal carcinoma xenografts in nude mice. Ai Zheng. 28:350–355.
2009.(In Chinese).
|