Introduction
Osteosarcoma (OS) is a malignant bone tumor that
typically occurs in children, adolescents and young adults.
Incorporation of chemotherapy into initial treatment significantly
increases the cure rate. However, ~40% of patients still die from
lung metastases (1,2). So, it is very important to develop
biomarkers that can inform therapy and provide prognostic insight,
especially into identifying poor prognosis patients who should be
offered more aggressive therapy at an early time-point in the
clinical continuum (3,4). Cancer is also a genetic disease
developing from a multi-step process. Single or multiple mutations
in genes related to growth control, invasion and metastasis form
the molecular genetic basis of malignant transformation and tumor
progression (5). Therefore,
identification of key genes and targets related to tumorigenesis is
crucial for OS prevention and treatment.
Cyclooxygenase-2 (COX-2) is an enzyme catalyzing the
conversion of arachidonic acid and O2 to prostaglandin
H2, the committed step in prostanoid biosynthesis. The
major final end product is prostaglandin E2
(PGE2). COX-2 expression is induced by various stimuli,
and the overexpression is closely related to the pathogenesis of
some degenerative diseases including cancer (6). COX-2 expression is found increased in
metastatic rhabdomyosarcoma, leiomyosarcoma and OS, and can be
considered as a prognostic value and a target for adjuvant therapy
(7–9). COX-2 is highly expressed in high grade
OS and application of COX-2 inhibitors may improve the tumor
response to chemotherapy and the outcome of OS patients (10). Moreover, COX-2 is directly
associated with the proliferation, migration and invasion in human
OS cells, and the therapeutic value of COX-2 inhibitors should be
evaluated continuously (11). COX-2
expression correlates inversely with disease-specific survival in
patients with OS lung metastases, indicating that COX-2 expression
in metastatic OS may have prognostic significance (12).
Intriguingly, researchers hold different views
towards the prognosis of COX-2 in OS. COX-2 overexpression in the
primary tumor correlates with the occurrence of distant metastasis
in patients with OS, predicts post-metastatic survival and can be
taken into consideration in the treatment of patients with OS. COX
2 is a valuable diagnostic marker for OS (13–15).
However, it has been proven that there is no significant
relationship between COX-2 expression and clinical outcome
(16). COX-2 expression does not
correlate with outcome of OS or rhabdomyosarcoma (17).
Therefore, it is indispensible to further elucidate
the function and molecular regulatory mechanisms of COX-2 in OS. In
the present study, the expression and clinical significance of
COX-2 and SUV were assessed using immunohistochemical (IHC) assay
in biopsy samples. Human MG-63 OS cells were treated with different
concentrations of NS-398, used to investigate its effects on cell
proliferation and apoptosis. Recombinant small hairpin RNA
adenovirus vector rAd5-SUV was constructed, and the effects and
molecular mechanisms of knockdown of SUV on proliferation and
apoptosis were evaluated in MG-63 cells, attempting to find the
potential therapeutic target for the treatment of OS.
Materials and methods
Materials
MG-63 cell line used in the experiment was from the
Laboratory of Second Affiliated Hospital of Xi’an Jiaotong
University; 6-week-old female immune-deficient nude mice
(BALB/c-nu) were purchased from Shanghai SLAC Laboratory Animal
Co., Ltd. (Shanghai Laboratory Animal Center of Chinese Academy
Sciences). Adenovirus-mediated SUV small hairpin RNA vector,
negative control vector and virion-packaging elements were from
Genechem (Shanghai, China); the primers of COX-2, SUV, PCNA and
CAS-3 were synthesized by ABI Co., Ltd. (USA). All antibodies were
from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Drugs and reagents
NS-398 was purchased from Cayman Co., Ltd. (USA);
3-(4,5)-dimethylthiahiazo(-z-yl)-3,5-di-phenytetrazoliumbromide
(MTT) was from Dingguo Biology (Shanghai, China); Dulbecco’s
modified Eagle’s medium (DMEM) and fetal bovine serum (FBS) were
from Thermo Fisher Scientific Inc.(Waltham, MA, USA); TRIzol
reagent and Lipofectamine 2000 were from Invitrogen (Carlsbad, CA,
USA); M-MLV Reverse Transcriptase was from Promega (Madison, WI,
USA); SYBR Green Master Mixture was from Takara (Otsu, Japan); Cell
Cycle Analysis kit and apoptosis kit [propidium iodide (PI), RNase
A, Annexin V-FITC] were from KeyGEN Biology (Nanjing, China).
ECL-PLUS/kit was from GE Healthcare (Piscataway, NJ, USA).
Tissue samples
Fifty freshly resected OS and OC samples were
collected at the Department of Orthopedics of Second Affiliated
Hospital of Xi’an Jiaotong University during 2010 and were
classified according to American Joint Committee on Cancer (AJCC)
TNM staging system. Tissues and clinical information were obtained
as part of an approved study at Xi’an Jiaotong University. There
were 30 cases of OS tissues and 20 cases of OC tissues. A portion
of each tissue sample was stored in liquid nitrogen for
histopathological and IHC examination. All tumors and normal
tissues were diagnosed by two independent pathologists.
IHC staining
Formalin-fixed tissue samples were prepared as
paraffin-embedded sections and stained with hematoxylin and eosin.
Unstained sections were deparaffinized and incubated overnight at
4°C with primary antibodies against COX-2 and SUV and with
biotinylated secondary antibody at room temperature for 1 h,
followed by incubation with ABC peroxidase and
3,3′-diaminobenzidine (DAB; 30 mg dissolved in 100 ml Tris-buffer
containing 0.03% H2O2). Sections were
counterstained with hematoxylin. Expression of COX-2 and SUV in
each specimen was scored according to the percentage of
positive-stained cells counted in five randomly selected high
magnification fields: 0, no expression; 1, positive cell ratio
<25%; 2, positive cell ratio 26–50%; and 3, positive cell ratio
>50%. The intensity score represents the staining intensity
(score 0, no staining signal; score 1, weak positive signal; score
2, moderate positive signal; score 3, strong positive signal).
Finally, a total expression score was given ranging from 0 to 12.
According to the product of these two indicators scoring the
results, they were divided into four levels: score 0–2 is
considered as (−), score 3–4 as (+), score 5–7 as (++) and score
8–9 is considered as (+++).
Cell culture and adenovirus
transfection
MG-63 cells were cultured in DMEM medium
supplemented with 10% heat-inactivated FBS, 100 U/ml of penicillin
and 100 μg/ml of streptomycin. They were all placed in a humidified
atmosphere containing 5% CO2 at 37°C. Recombinant
adenovirus vector rAd5-SUV and negative control rAd5-GFP were
transfected into MG-63 cells. Cells were subcultured at a 1:5
dilution in 300 μg/ml G418-containing medium. Positive stable
transfectants were selected and expanded for further study. The
clone in which the rAd5-SUV virus vectors transfected was named as
rAd5-SUV group, the negative control vectors transfected was named
as GFP group and MG-63 cells as CON group.
RT-PCR
To quantitatively determine the mRNA expression
level of COX-2, SUV, PCNA and CAS-3 in MG-63 cells, RT-PCR was
used. Total RNA of each clone was extracted with TRIzol according
to the manufacturer’s instructions. Reverse-transcription was
carried out using M-MLV and cDNA amplification was carried out
using SYBR Green Master Mix kit according to the manufacturer’s
instructions. The genes were amplified using specific
oligonucleotide primer and human glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) gene was used as an endogenous control. The
PCR primer sequences were as follows: COX-2, 5′-GAAGTACCAAGCTGT
GCTTGAATAA-3′ and 5′-GGCTTGATTCCAATGCAC CTA-3′; SUV,
5′-ACCAGGTGAGAAGTGAGGGA-3′ and 5′-AACAGTAGAGGAGCCAGGGA-3′; PCNA,
5′-CCATCCT CAAGAAGGTGTTGG-3′ and 5′-GTGTCCCATATCCGCAA TTTTAT-3′;
CAS-3, 5′-AGAGGGGATCGTTGTAGAAG-3′ and 5′-GTTGCCACCTTTCGGTTAAC-3′;
GAPDH, 5′-CAAC GAATTTGGCTACAGCA-3′ and 5′-AGGGGTCTACATGGC AACTG-3′.
Data were analyzed using the comparative Ct method
(2−ΔΔCt). Three separate experiments were performed for
each clone.
Western blot assay
MG-63 cells were harvested and extracted using lysis
buffer (Tris-HCl, SDS, mercaptoethanol, glycerol). Cell extracts
were boiled for 5 min in loading buffer and then equal amount of
cell extracts was separated on 15% SDS-PAGE gels. Separated protein
bands were transferred into polyvinylidene fluoride (PVDF)
membranes and the membranes were blocked in 5% skim milk powder.
The primary antibodies against COX-2, SUV, PCNA and CAS-3 were
diluted according to the instructions of antibodies and incubated
overnight at 4°C. Then, horseradish peroxidase-linked secondary
antibodies were added at a dilution ratio of 1:1000, and incubated
at room temperature for 2 h. The membranes were washed with PBS
three times and the immunoreactive bands were visualized using
ECL-PLUS/Kit according to the kit’s instruction. The relative
protein level in different cell lines was normalized to GAPDH
concentration. Three separate experiments were performed for each
clone.
Cell proliferation assay
Cell proliferation was analyzed with the MTT assay.
Briefly, cells infected with rAd5-SUV were incubated in
96-well-plates at a density of 1×105 cells per well with
DEME medium supplemented with 10% FBS. Cells were treated with 20
μl MTT dye at 0, 24, 48 and 72 h and then incubated with 150 μl of
DMSO for 5 min. The color reaction was measured at 570 nm with
enzyme immunoassay analyzer (Bio-Rad, USA). The proliferation
activity was calculated for each clone.
Cell apoptosis analysis
To detect cell apoptosis, cells were trypsinized,
washed with cold PBS and resuspended in binding buffer according to
the instruction of the apoptosis kit. FITC-Annexin V and PI were
added to the fixed cells for 20 min in darkness at room
temperature. Then, Annexin V binding buffer was added to the
mixture before the fluorescence was measured on FACsort flow
cytometer. The cell apoptosis was analyzed using the CellQuest
software (Becton-Dickinson, USA). Three separate experiments were
performed for each clone.
In vivo tumor xenograft studies
Four mice were injected subcutaneously with
1×108 MG-63 cells in 50 μl of PBS pre-mixed with an
equal volume of matrigel matrix (Becton-Dickinson). Mice were
monitored daily, and three out of four mice developed a
subcutaneous tumor. When the tumor size reached approximately 5 mm
in length, they were surgically removed, cut into 1–2
mm3 pieces, and re-seeded individually into 18 other
mice. When tumor size reached ~5 mm in length, the mice were
randomly assigned to MG-63, rAd5-GFP and rAd5-SUV groups. In
rAd5-GFP and rAd5-SUV groups, 15 μl of adenovirus was injected into
subcutaneous tumors using a multi-site injection format. Mice in
the MG-63 group received 15 μl of PBS only. Injections were
repeated on the third day after initial treatment. The tumor volume
every three days was measured with a caliper, using the formula
volume = (length × width)2/2.
Statistical analysis
The results of each experiment are shown as mean ±
SD when applicable. Statistically significant difference in each
assay was determined by SPSS version 11.5. Difference in each group
was tested for significance using χ2 test and ANOVA
analysis of variance. P<0.05 was considered significant.
Results
The expression of COX-2 and SUV in human
OS
The expression of COX-2 and SUV in OS and OC was
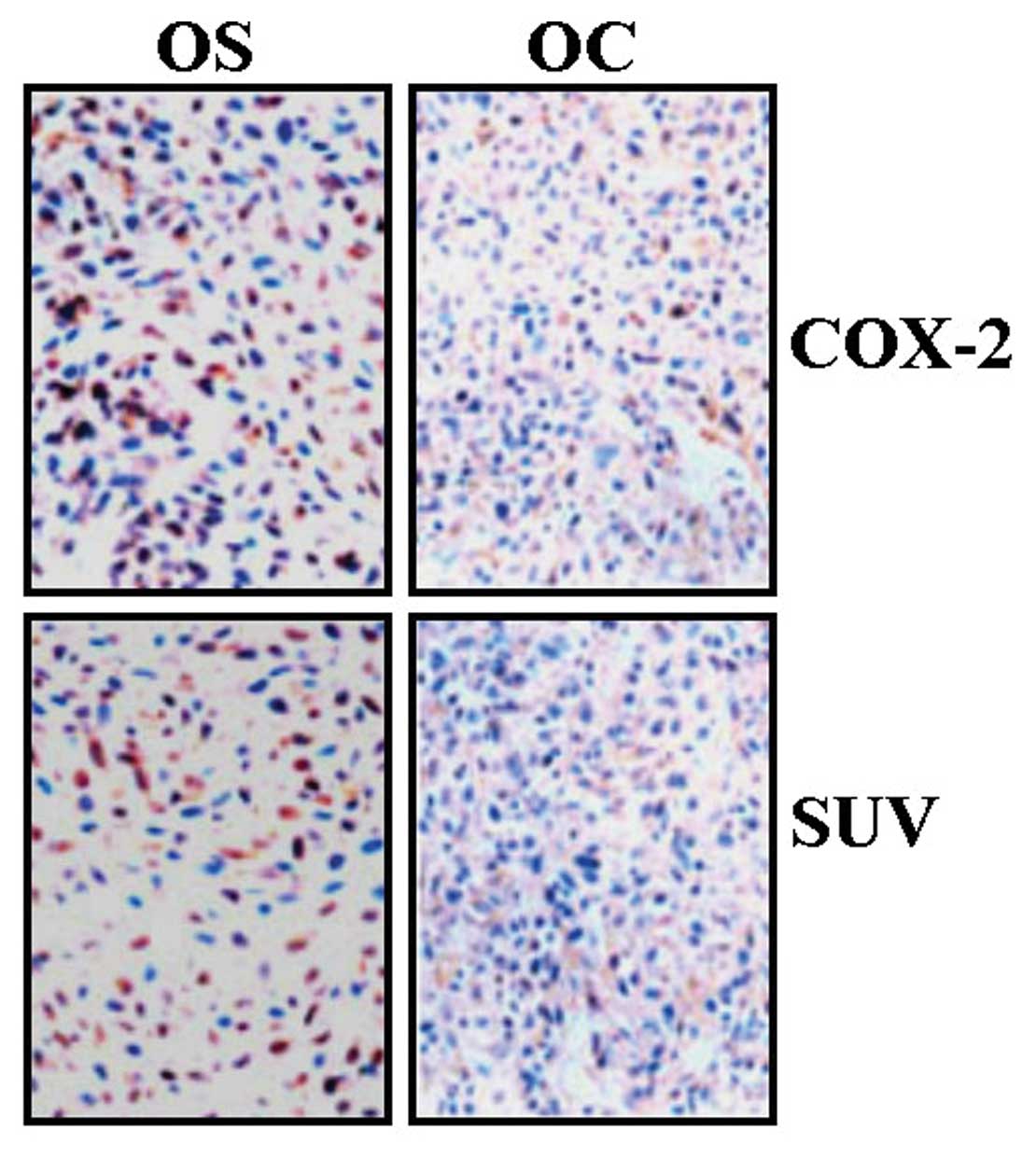
evaluated using IHC assays. As shown in Fig. 1 and Table I, the expression of COX-2 and SUV
was mainly localized in the cytoplasm. The expression of COX-2 and
SUV was respectively observed in 73.3 and 63.3% OS tissues and in
25.0 and 30.0% OC tissues, indicating their higher expression in OS
compared with OC. Spearman rank correlation analysis showed a
positive correlation between COX-2 and SUV expression in OS
(r=0.975, P=0.025).
 | Table IThe expression of COX-2 and SUV in OS
and OC tissues. |
Table I
The expression of COX-2 and SUV in OS
and OC tissues.
| | n | | | | |
|---|
| |
| | | | |
|---|
| Target | Sample | − | + | Total | Positive rate
(%) | χ2 | P-value |
|---|
| COX-2 | OS | 8 | 22 | 30 | 73.3 | 11.06 | 0.001 |
| OC | 15 | 5 | 20 | 25.0 | | |
| SUV | OS | 11 | 19 | 30 | 63.3 | 5.23 | 0.02 |
| OC | 14 | 6 | 20 | 30.0 | | |
The relationship of COX-2 and SUV
expression with the clinicopathologic features of OS
The relationship between the expression of COX-2 and
SUV and clinicopathologic features was analyzed. As shown in
Table II, no significant
correlation was found between the expression of COX-2 and SUV with
age and pathological grade and types of OS.
| Table IIThe relationship of COX-2 and SUV
expression with clinicopathological characteristics of OS. |
Table II
The relationship of COX-2 and SUV
expression with clinicopathological characteristics of OS.
| COX-2 | | | SUV | | |
|---|
|
| | |
| | |
|---|
| Clinicopathologic
factors | − | + | χ2 | P-value | − | + | χ2 | P-value |
|---|
| Age |
| >60 | 5 | 7 | 2.24 | 0.14 | 6 | 7 | 0.86 | 0.35 |
| <60 | 3 | 15 | | | 5 | 12 | | |
| Pathological
grade |
| I | 2 | 5 | | | 3 | 5 | | |
| II | 4 | 10 | 0.13 | 0.94 | 5 | 8 | 0.06 | 0.97 |
| III | 2 | 7 | | | 3 | 6 | | |
| Pathological
type |
| Osteoblastic | 4 | 9 | | | 4 | 8 | | |
|
Chondroblastic | 2 | 8 | 0.34 | 0.84 | 4 | 6 | 0.10 | 0.95 |
| Others | 2 | 5 | | | 3 | 5 | | |
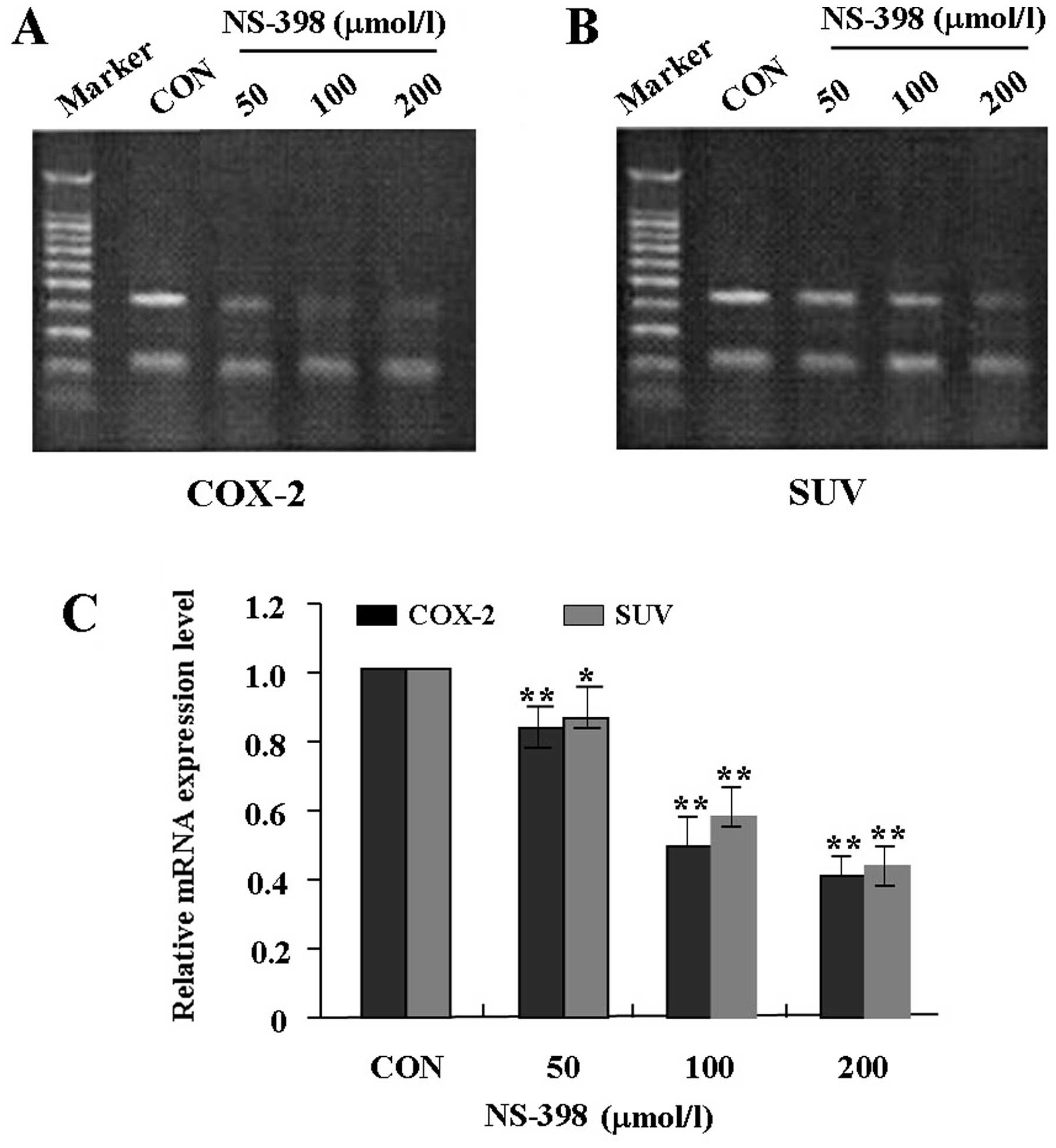
Effects of NS-398 on mRNA expression of
COX-2 and SUV in MG-63 cells
To examine the effects of NS-398 on expression of
COX-2 and SUV in MG-63 cells, MG-63 cells were treated with
different concentrations of NS-398 (0, 50, 100 and 200 μmol/l).
RT-PCR was performed at 48-h recovery to measure their mRNA
expression levels. As shown in Fig.
2, the mRNA expression levels of COX-2 and SUV were
significantly lower in NS-398 treated groups in a dose-dependent
manner than the control group, suggesting that NS-398 inhibited the
mRNA expression of COX-2 and SUV in MG-63 cells.
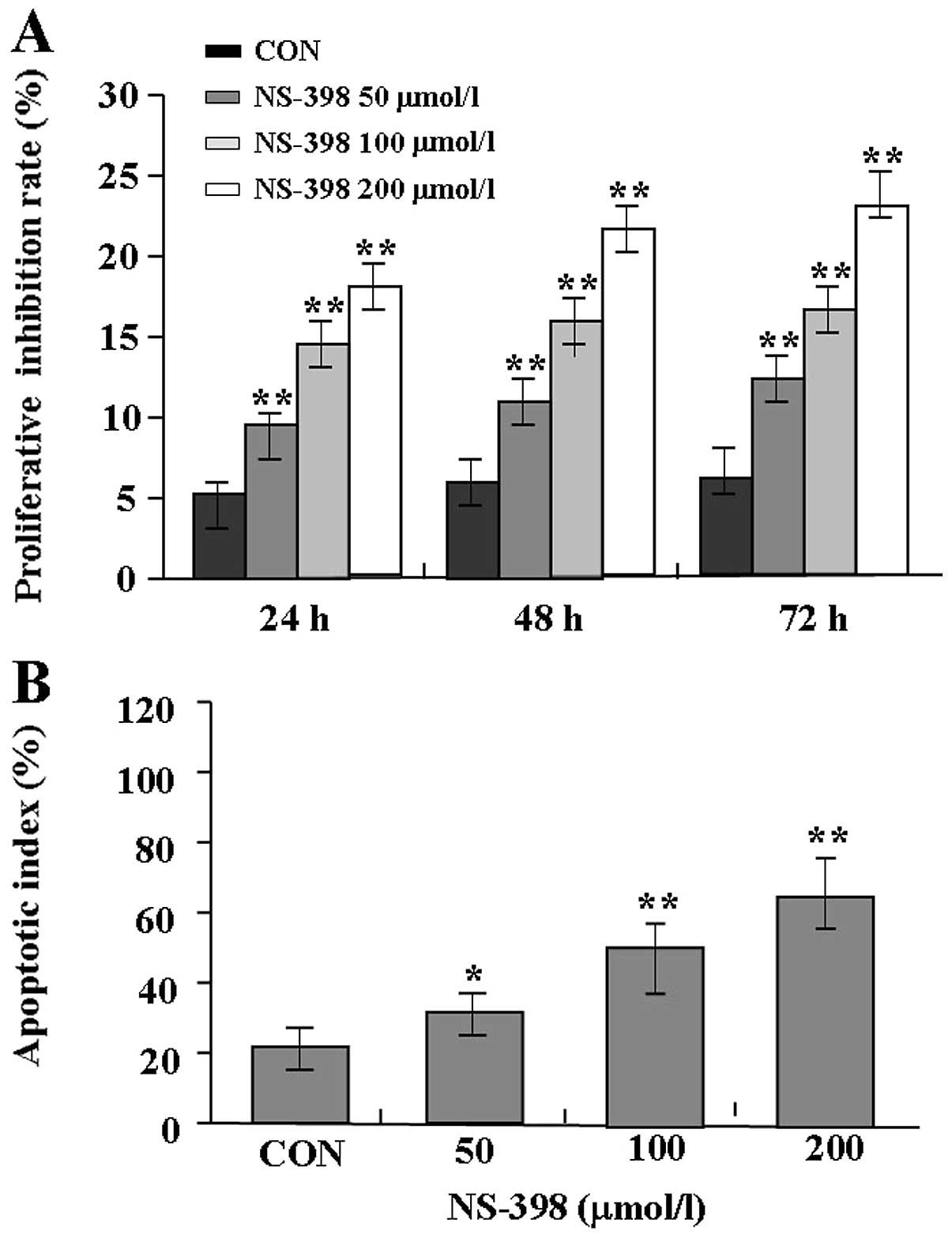
Effects of NS-398 on proliferation and
apoptosis in MG-63 cells
The proliferative activities of MG-63 cells treated
with NS-398 were examined by MTT assay, and it was found that
NS-398 could significantly reduce the proliferative activities of
MG-63 cells in a dose- and time-dependent manner in comparison with
the control group (Fig. 3A). Also,
the apoptotic index of MG-63 cells treated with NS-398 was examined
by flow cytometric analysis. The results showed that the apoptosis
index of MG-63 cells in NS-398 treated groups was markedly higher
than the control group (Fig. 3B).
Therefore, NS-398 inhibited the proliferation and induced apoptosis
in MG-63 cells.
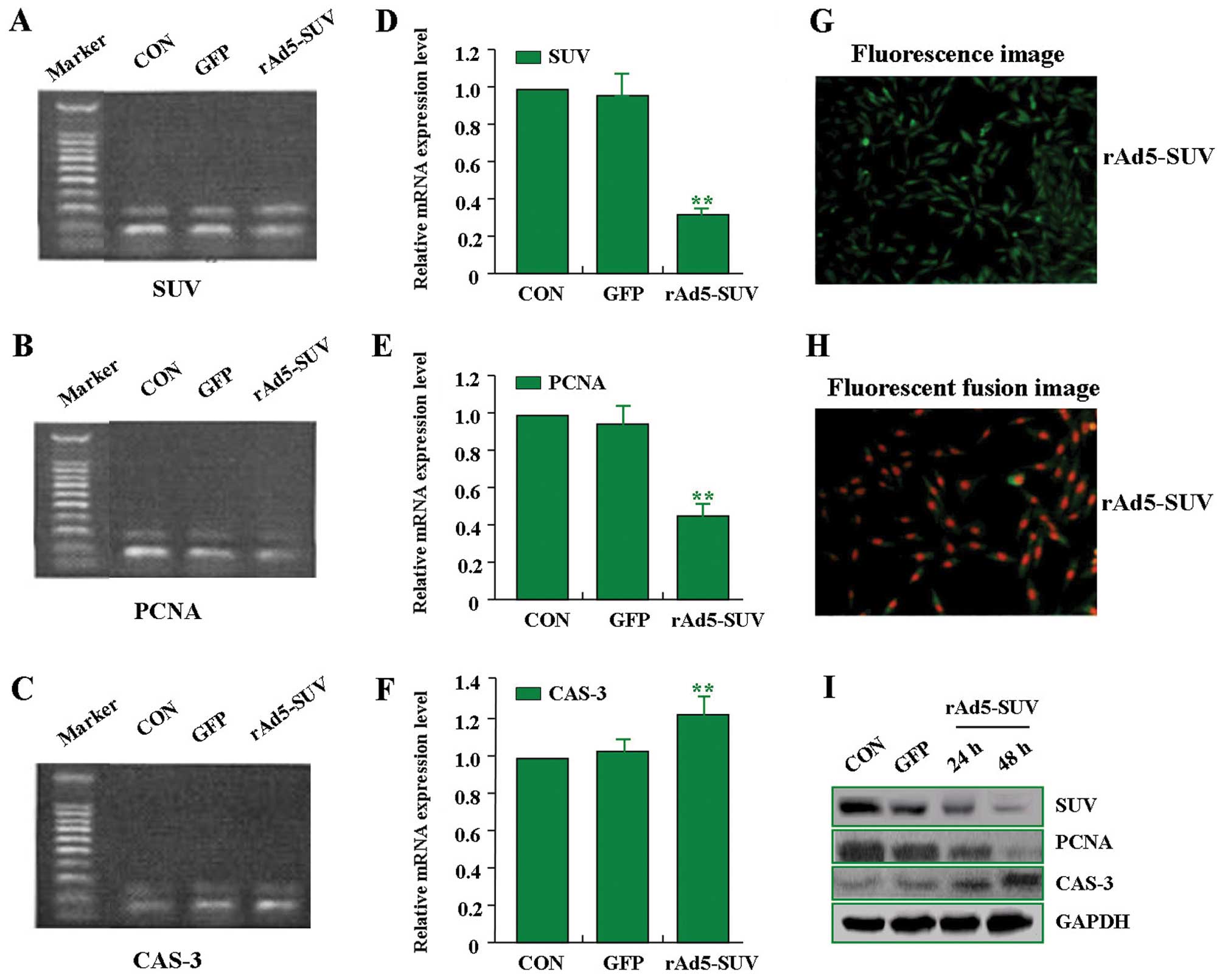
Effects of rAd5-SUV on expression of SUV,
PCNA, and CAS-3 in MG-63 cells
In order to efficiently knockdown the expression of
SUV in MG-63 OS cells, an adenovirus-mediated small hairpin RNA
approach was used to construct the rAd5-SUV vector. In pilot
studies, the transfection efficiency of rAd5-SUV (MOI=100) in MG-63
cells was >95.0% (Fig. 4G and
H). After rAd5-SUV was transfected into MG-63 cells, RT-PCR and
western blot assays were performed to measure the expression of
SUV, PCNA, and CAS-3. As shown in Fig.
4A–F and I, the expression of SUV and PCNA was decreased, while
CAS-3 expression was increased in rAd5-SUV group compared with the
GFP group and CON group. Therefore, knockdown of SUV inhibited the
expression of PCNA and enhanced the expression of CAS-3 in MG-63
cells.
Effects of rAd5-SUV on proliferation and
apoptosis of MG-63 cells
Deregulated cell proliferation is a hallmark of
cancer (18). In order to test the
effects of rAd5-SUV on OS cell proliferation and apoptosis, we
investigated the proliferative activities and apoptotic index of
MG-63 cells by MTT and flow cytometry analysis. As a result, it was
indicated that knockdown of SUV could significantly reduce the
proliferative activities of MG-63 cells compared with GFP group and
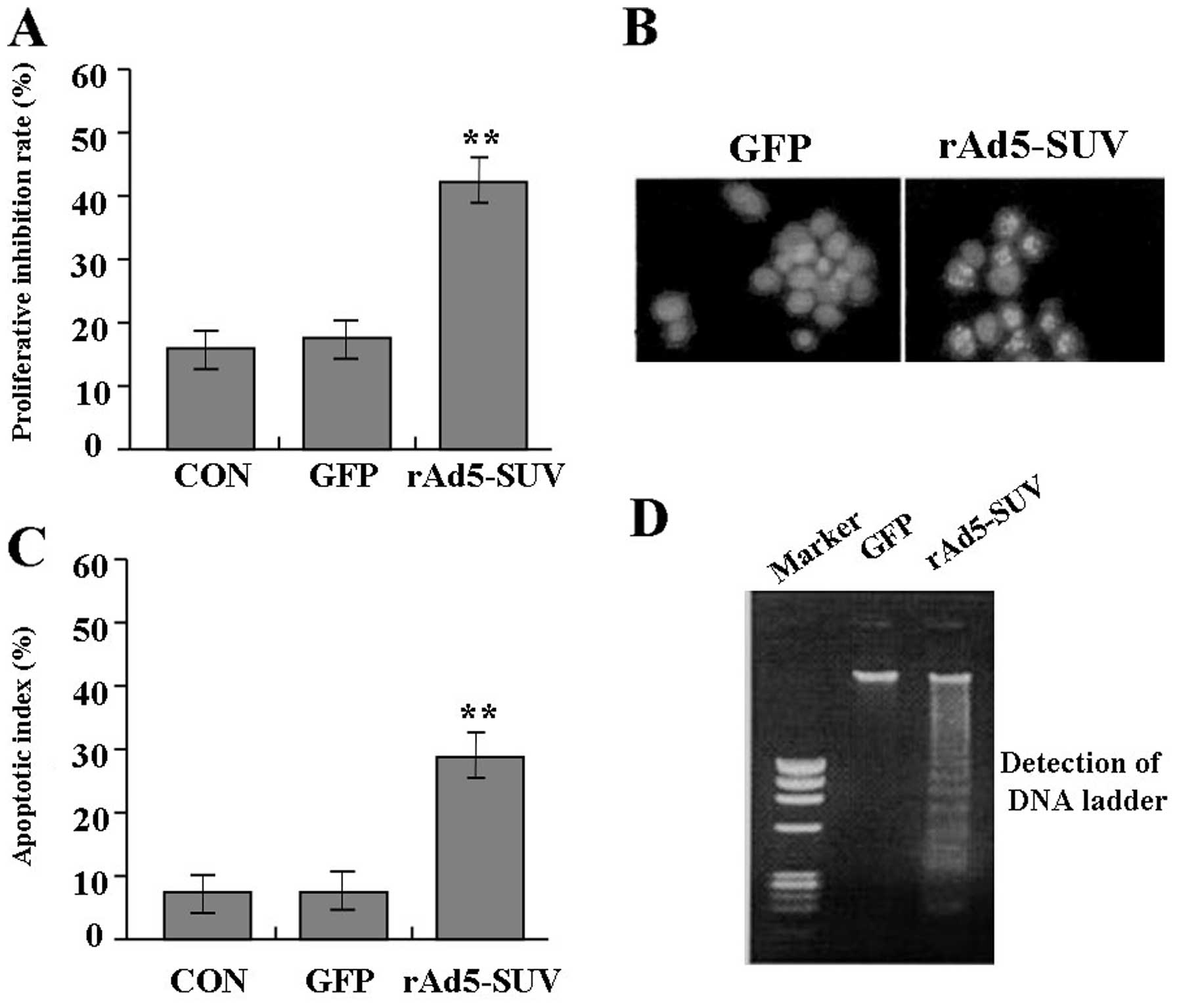
CON group (Fig. 5A). Also, cell
nuclear fragmentation, apoptotic bodies and DNA ladder turned up in
group rAd5-SUV compared with the group GFP, demonstrating the DNA
fragmentation and increase of cell apoptosis induced by knockdown
of SUV (Fig. 5B and D). Moreover,
knockdown of SUV markedly increased the apoptotic index of MG-63
cells compared with GFP group and CON group indicated by flow
cytometry (Fig. 5C). Therefore,
knockdown of SUV inhibited cell proliferation and induce apoptosis
in MG-63 cells.
Effects of rAd5-SUV on xenograft tumor
growth
Our in vitro experiments demonstrated the
inhibitory effects of knockdown of SUV on OS MG-63 cell
proliferation. Therefore, it is necessary to further investigate
the effect of knockdown of SUV on xenograft tumor growth in
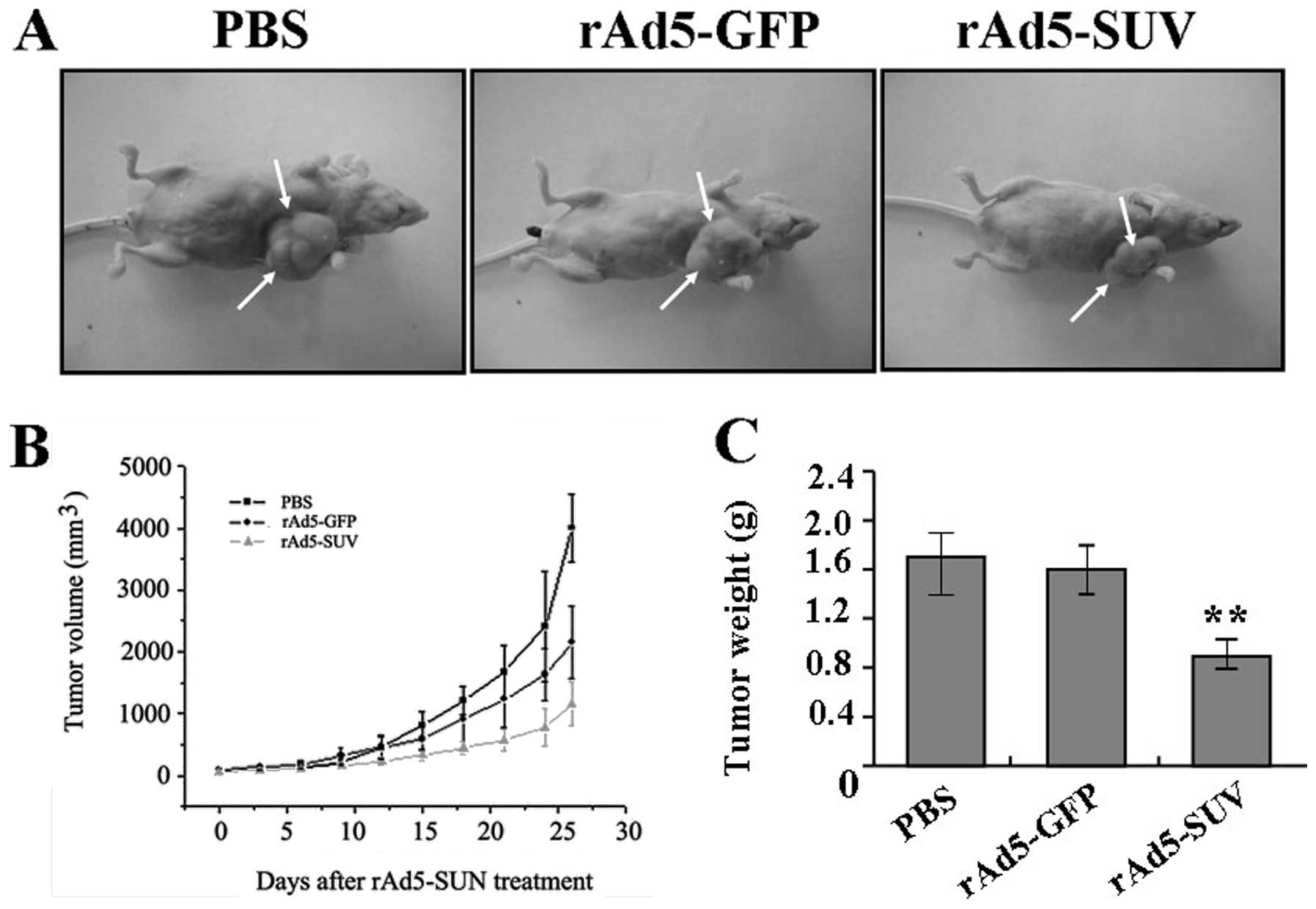
vivo. The mean volume of tumors in all experimental mice before
treatment was 101.05±36.27 mm3. During the whole tumor
growth period (Fig. 6A and B), the
tumor growth activity was measured. Tumors treated with rAd5-SUV
grew substantially slowly compared with the PBS and rAd5-GFP group.
When the tumors were harvested, the average weight of tumors in
group rAd5-SUV was significantly lower than PBS and rAd5-SUV group
(Fig. 6C). This result in
vivo indicated that knockdown of SUV could also inhibit OS cell
growth.
Discussion
OS is the most frequent malignant bone tumor with a
peak incidence in the second and third decade of life. SUV as a
member of the inhibitor of apoptosis protein family is expressed
both during normal fetal development and in human cancer.
Importantly, it is a useful prognostic marker in OS and patients
with OS exhibiting nuclear SUV expression could potentially benefit
from stratification of neoadjuvant chemotherapy (19). Elevated SUV expression in OS
correlates with histologic grade and mitotic index and a decreased
disease-free interval. SUV attenuation in canine OS cells inhibits
cell cycle progression, increased apoptosis, mitotic arrest and
chemosensitivity, and cooperates with chemotherapy to significantly
improve tumor control (20). Thus,
SUV can be considered as an independent predictor of survival for
OS patients (21,22). Coupled with the report that COX-2
expression does not correlate with outcome of OS (16), the relationship of COX-2 and SUV
with OS need to be further evaluated. In our study, the expression
of COX-2 and SUV was, respectively, observed in 73.3 and 63.3% OS
tissues and in 25.0 and 30.0% OC tissues, indicating their higher
expression in OS than in OC. Spearman rank correlation analysis
showed their positive correlation. However, consistent with a
previous study (17), our results
showed no significant correlation between the expression of COX-2
and SUV with age and pathological grade and type of OS.
In addition, COX-2 inhibitors such as NS-398 and
celecoxib have been shown to inhibit COX-2 expression, and produce
an anti-proliferative and pro-apoptotic effect on different types
of tumor cells (23,24). Meloxicam, the preferential COX-2
inhibitor, inhibits OS growth, invasiveness and metastasis by
COX-2-dependent and -independent routes (25,26).
RNAI-mediated knockdown of COX-2 inhibits the growth, invasion and
migration of OS, and COX-2 signaling pathway may provide a novel
therapeutic target for the treatment of human OS (27). Some data indicate that selective
inhibition of COX-2 exerts an effect on primary tumor growth in
Ewing sarcoma (28). Furthermore,
we investigated the effect of NS-398 on OS cell proliferation and
apoptosis, and found that NS-398 significantly inhibited the
proliferation and induce apoptosis in OS cells, enriching the
anti-tumor evidence of COX-2 inhibitors. Also, the regulatory
mechanisms of COX-2 inhibitors on OS are worth exploring. Celecoxib
induces apoptosis in human OS cells via downregulation of PI3K/Akt,
activating GSK-3β and inhibiting β-catenin-dependent signaling
pathways (29,30). Differently, our study showed that
NS-398 downregulated the mRNA expression of SUV in a dose-dependent
manner in MG-63 cells, suggesting that MG-63 might inhibit the
proliferation and induce apoptosis of MG-63 cells through
downregulation of the SUV pathway.
SUV is very important in the development of OS and
blockade of SUV markedly inhibits the proliferation and invasion of
OS cells, partially reversing their malignant phenotype. Targeting
SUV might be a promising option in the treatment of OS and
downregulation of SUV is an effective strategy to improve the
therapeutic effect of OS (31).
Similarly, our study indicated that knockdown of SUV by
adenovirus-mediated RNAI inhibited the proliferation, induced
apoptosis, and slowed the growth of xenograft tumors in MG-63
cells, providing a strategy for the treatment of OS. PCNA is
essential for the replication of deoxyribonucleic acid DNA and has
been proved to be an important marker for tumor proliferation. SUV
expression has been verified to correlate with PCNA and CAS-3 in OS
(32,33). Moreover, we found that knockdown of
SUV decreased the expression of PCNA and increased the expression
of CAS-3 in MG-63 cells, suggesting that SUV might be involved in
OS proliferation and apoptosis via regulation of PCNA and CAS-3
expression.
In conclusion, the expression of COX-2 and SUV is
closely correlated with human OS, and NS-398 inhibition of COX-2 or
knockdown of SUV by RNAI suppresses tumor proliferation and induces
apoptosis in MG-63 cells, suggesting that COX-2 may be involved in
OS cell growth and apoptosis through SUV-mediated regulation of
PCNA and CAS-3 expression, and provide a potential therapeutic
strategy for the treatment of cancer.
References
|
1
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Raymond AK, Ayala AG and Knuutila S:
Conventional osteosarcoma. World Health Organization Classification
of Tumours: Pathology and Genetics of Tumours of Soft Tissue and
Bone. Fletcher CDM, Unni KK and Mertens F: IARC Press; Lyon: pp.
264–270. 2002
|
|
3
|
Bielack SS, Kempf-Bielack B, Delling G, et
al: Prognostic factors in high-grade osteosarcoma of the
extremities or trunk: an analysis of 1,702 patients treated on
neoadjuvant cooperative osteosarcoma study group protocols. J Clin
Oncol. 20:776–790. 2002. View Article : Google Scholar
|
|
4
|
Kong C and Hansen MF: Biomarkers in
osteosarcoma. Expert Opin Med Diagn. 3:13–23. 2009. View Article : Google Scholar
|
|
5
|
Tajima Y, Yamazaki K, Makino R, et al:
Gastric and intestinal phenotypic marker expression in early
differentiated-type tumors of the stomach: clinicopathologic
significance and genetic background. Clin Cancer Res. 12:6469–6479.
2006. View Article : Google Scholar
|
|
6
|
Rodrigues S, Bruyneel E, Rodrigue CM, et
al: Cyclooxygenase-2 and carcinogenesis. Bull Cancer. 91:S61–S76.
2004.
|
|
7
|
Dickens DS, Kozielski R, Khan J, et al:
Cyclooxygenase-2 expression in pediatric sarcomas. Pediatr Dev
Pathol. 5:356–364. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mullins MN, Lana SE, Dernell WS, et al:
Cyclooxygenase-2 expression in canine appendicular osteosarcomas.
Vet Intern Med. 18:859–865. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Raspollini MR, Amunni G, Villanucci A, et
al: Cyclooxygenase-2 expression in uterine leiomyosarcomas. J
Chemother. 16:577–581. 2004. View Article : Google Scholar
|
|
10
|
Masi L, Recenti R, Silvestri S, et al:
Expression of cyclooxygenase-2 in osteosarcoma of bone. Appl
Immunohistochem Mol Morphol. 15:70–76. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee EJ, Choi EM, Kim SR, et al:
Cyclooxygenase-2 promotes cell proliferation, migration and
invasion in U2OS human osteosarcoma cells. Exp Mol Med. 39:469–476.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rodriguez NI, Hoots WK, Koshkina NV, et
al: COX-2 expression correlates with survival in patients with
osteosarcoma lung metastases. J Pediatr Hematol Oncol. 30:507–512.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Urakawa H, Nishida Y, Naruse T, et al:
Cyclooxygenase-2 overexpression predicts poor survival in patients
with high-grade extremity osteosarcoma: a pilot study. Clin Orthop
Relat Res. 467:2932–2938. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Boulytcheva IV, Soloviev YN, Kushlinskii
NE and Mahson AN: Expression of molecular markers in the tumor and
survival prognosis in osteosarcoma. Bull Exp Biol Med. 150:237–242.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hosono A, Yamaguchi U, Makimoto A, et al:
Utility of immunohistochemical analysis for cyclo-oxygenase 2 in
the differential diagnosis of osteoblastoma and osteosarcoma. J
Clin Pathol. 60:410–414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Carmody Soni EE, Miller BJ, Scarborough
MT, et al: Cyclooxygenase-2 expression is not associated with
clinical outcome in synovial sarcoma. Oncol Rep. 26:1513–1517.
2011.
|
|
17
|
Dickens DS, Kozielski R, Leavey PJ, et al:
Cyclooxygenase-2 expression does not correlate with outcome in
osteosarcoma or rhabdomyosarcoma. J Pediatr Hematol Oncol.
25:282–285. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hanahan D and Weinberg RA: The hallmarks
of cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Trieb K, Lehner R, Stulnig T, et al:
Survivin expression in human osteosarcoma is a marker for survival.
Eur J Surg Oncol. 29:379–382. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shoeneman JK, Ehrhart EJ III, Eickhoff JC,
et al: Expression and function of survivin in canine osteosarcoma.
Cancer Res. 72:249–259. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Osaka E, Suzuki T, Osaka S, et al:
Survivin expression levels as independent predictors of survival
for osteosarcoma patients. J Orthop Res. 25:116–121. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Osaka E, Suzuki T, Osaka S, et al:
Survivin as a prognostic factor for osteosarcoma patients. Acta
Histochem Cytochem. 39:95–100. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wiontzek M, Matziolis G, Schuchmann S, et
al: Effects of dexamethasone and celecoxib on calcium homeostasis
and expression of cyclooxygenase-2 mRNA in MG-63 human osteosarcoma
cells. Clin Exp Rheumatol. 24:366–372. 2006.PubMed/NCBI
|
|
24
|
Moalic S, Liagre B, Le Bail JC and
Beneytout JL: Dose-dependent modulation of apoptosis and
cyclooxygenase-2 expression in human 1547 osteosarcoma cells by
NS-398, a selective cyclooxygenase-2 inhibitor. Int J Oncol.
18:533–540. 2001.PubMed/NCBI
|
|
25
|
Naruse T, Nishida Y, Hosono K and Ishiguro
N: Meloxicam inhibits osteosarcoma growth, invasiveness and
metastasis by COX-2-dependent and independent routes.
Carcinogenesis. 27:584–592. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wolfesberger B, Hoelzl C, Walter I, et al:
In vitro effects of meloxicam with or without doxorubicin on canine
osteosarcoma cells. Vet Pharmacol Ther. 29:15–23. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao Q, Wang C, Zhu J, et al:
RNAi-mediated knockdown of cyclooxygenase2 inhibits the growth,
invasion and migration of SaOS2 human osteosarcoma cells: a case
control study. J Exp Clin Cancer Res. 30:262011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gendy AS, Lipskar A, Glick RD, et al:
Selective inhibition of cyclooxygenase-2 suppresses metastatic
disease without affecting primary tumor growth in a murine model of
Ewing sarcoma. J Pediatr Surg. 46:108–114. 2011. View Article : Google Scholar
|
|
29
|
Liu B, Shi ZL, Feng J and Tao HM:
Celecoxib, a cyclooxygenase-2 inhibitor, induces apoptosis in human
osteosarcoma cell line MG-63 via down-regulation of PI3K/Akt. Cell
Biol Int. 32:494–501. 2008. View Article : Google Scholar
|
|
30
|
Xia JJ, Pei LB, Zhuang JP, et al:
Celecoxib inhibits β-catenin-dependent survival of the human
osteosarcoma MG-63 cell line. J Int Med Res. 38:1294–1304.
2010.
|
|
31
|
Liang X, Da M, Zhuang Z, et al: Effects of
Survivin on cell proliferation and apoptosis in MG-63 cells in
vitro. Cell Biol Int. 33:119–124. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang W, Luo H and Wang A: Expression of
survivin and correlation with PCNA in osteosarcoma. J Surg Oncol.
93:578–584. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zou J, Gan M, Mao N, et al: Sensitization
of osteosarcoma cell line SaOS-2 to chemotherapy by downregulating
survivin. Arch Med Res. 41:162–169. 2010. View Article : Google Scholar : PubMed/NCBI
|