Introduction
Hyperthermia therapy is an important approach to
tumor treatment. Acting as an external stimuli, high temperatures
activate the mitochondrial apoptosis pathway to promote the
apoptosis of tumor cells, which is regulated by the Bcl-2 gene
family. Interference and disruption of the hyperthermia-induced
apoptosis pathway stimulates tumor cell insensitivity to
heat-induced cell apoptosis(1).
For many years, research on the effect and
regulation of hyperthermia treatment has focused mainly on the
tumor cell (2,3). However, the regulation of the thermal
destruction of tumor cells in a stromal microenvironment is rarely
investigated. Recent studies have shown that tumor development and
treatment are not only determined by tumor cells but are also
closely related to the balance of the tumor-stroma microenvironment
(4,5).
The majority of tumor stromal cells are fibroblasts.
Compared to normal fibroblasts (NFs), tumor fibroblasts are
significantly altered. Tumor fibroblasts have certain biological
characteristics of both fibroblasts and myofibroblasts and are also
known as cancer-associated fibroblast cells (CAFs). CAFs produce a
tumor extracellular matrix, which provides a suitable
microenvironment to support the growth of tumors (6,7) and
also to promote tumor cell proliferation, invasion and metastasis
(8). Animal experiments have
demonstrated that CAFs shorten the incubation period and increase
the formation rate of tumor cells, reduce the dose of tumor cells
required for tumor formation, promote the invasion and metastasis
of tumor cells and reduce the effects of tumor treatment when tumor
cells are injected into nude mice (7,9–11).
Previous studies have shown that in in vitro models of
pancreatic cancer, the efficacy of chemotherapy was significantly
decreased and cell apoptosis was reduced when cancer cells were
co-cultured with CAFs (12,13). It was also found, based on in
vitro experiments, that the addition of CAFs in breast cancer
enhanced the tolerance of the cancer to tamoxifen (chemotherapeutic
drug) (14). In clinical studies,
it was found that patients with rectal or pancreatic cancer had a
poor prognosis if their pathological examination of surgical
specimens showed high expression levels of α-SMA, which indirectly
proved that CAFs protect and promote tumor development (15,16).
As mentioned above, considering the regulation of
the tumor stroma in cancer therapy, this study is focused on the
main component of tumor stroma, CAFs, to screen differentially
expressed secreted proteins in oral cancer CAFs and NFs using
cytokine chip technology. Additionally, we aimed to explore the
characteristics of the secreted factor of oral CAFs and the
regulation of the oral CAFs in cell apoptosis of heat-induced
Tca8113 cells through in vitro indirect co-culture of oral
cancer CAFs and Tca8113 cells as a simulation of in vivo
tumor-stroma interaction.
Materials and methods
Materials
The following materials were used in this study:
tongue squamous carcinoma cell line Tca8113 (Shanghai Bio-Cell
Bank, China); RPMI-1640 medium (Hyclone Biochemical Products,
Beijing Co., Ltd., China); mouse anti-human α-smooth muscle actin,
cytokeratin, vimentin monoclonal antibodies and EnVision
immunohistochemistry kit (Fuzhou Maixin Biotechnology Development
Co., Ltd., China); mouse anti-human Bcl-2, Bax, CXCL9/MIG, CXCR3
antibodies (USA RD Cos.); and RayBio® human cytokine
antibody chip kit (RayBiotech, Inc., USA).
Fibroblast culture
Oral NFs and oral CAFs were isolated from normal
oral mucosa and oral squamous cell carcinoma tissues confirmed by
pathological analysis, respectively, following standard tissue
culture instructions. Cells were cultured in an RPMI-1640 medium
containing 10% FBS and incubated at 37°C, in 5% CO2 in a
humidified incubator.
Tca8113 cell culture and heat
treatment
The Tca8113 cells were cultured in an RPMI-1640
medium containing 10% FBS (complete medium) and incubated at 37°C,
in 5% CO2 in a humidified incubator. Heat treatment was
performed in a 43°C water bath for 80 min.
Immunocytochemistry
Logarithmic-growing fibroblasts and Tca8113 cells
were seeded and then fixed by cold acetone. Immunocytochemistry was
performed using the EnVision immunocytochemistry kit, visualized by
DAB and restained with hematoxylin.
Cytokine antibody microarray
hybridization
The cells were cultured for 24 h and the culture
supernatants were collected. Cytokine antibody microarray
hybridization was performed using a RayBio® human
cytokine antibody chip kit according to the manufacturer’s
instructions and the differences between the 2 groups were analyzed
with ScanAlyze software after X-ray film exposure.
Preparation of conditioned medium
NFs and CAFs were cultured in an RPMI-1640 medium
containing 10% FBS (complete medium) and the supernatants were
collected. Conditioned medium was prepared by mixing the collected
supernatants with a complete medium at a different volume ratio
(Table I).
 | Table IPreparation of the conditioned
medium. |
Table I
Preparation of the conditioned
medium.
| NF-conditioned
medium | | CAF-conditioned
medium |
|---|
|
| |
|
|---|
| No. | NF supernatants | Complete medium | No. | CAF supernatants | Complete medium |
|---|
| 1 | 0/6 | 6/6 | 4 | 0/6 | 6/6 |
| 2 | 1/6 | 5/6 | 5 | 1/6 | 5/6 |
| 3 | 2/6 | 4/6 | 6 | 2/6 | 4/6 |
Antibody neutralization
Tca8113 cells were cultured with a conditioned
medium containing 20 μg/ml of an antibody against CXCL9/MIG or
CXCR3 for 24 h, and then incubated in a 43°C water bath for 80
min.
Apoptosis analysis by flow cytometry
Cells were harvested, fixed with 70% cold ethanol
and stained with propidium iodide (PI) staining solution (100 μg/ml
PI, 0.1 mg/ml RNase, 0.1% Triton X-100). The DNA content was
analyzed using Beckman Coulter Epics XL flow cytometry, and the
apoptosis rate was calculated by using the sub-diploid peak prior
to the G0/G1 phase as the apoptosis signal.
Western blot analysis
Cellular proteins were transferred to a PVDF
membrane after the SDS-PAGE electrophoresis. The membrane was
incubated with a primary antibody solution (1:1000 diluted in TBST)
at 4°C overnight, washed with 1X TBST, incubated with a secondary
antibody solution (1:5000 diluted in TBST), washed with 1X TBST for
1 h and then exposed to film.
Statistical analysis
Statistical analysis was performed with SPSS17.0
statistical software using a one-way ANOVA and a comparison of the
2 groups was performed using an LSD test; P<0.05 was considered
to indicate a statistically significant difference.
Results
Culture of fibroblasts
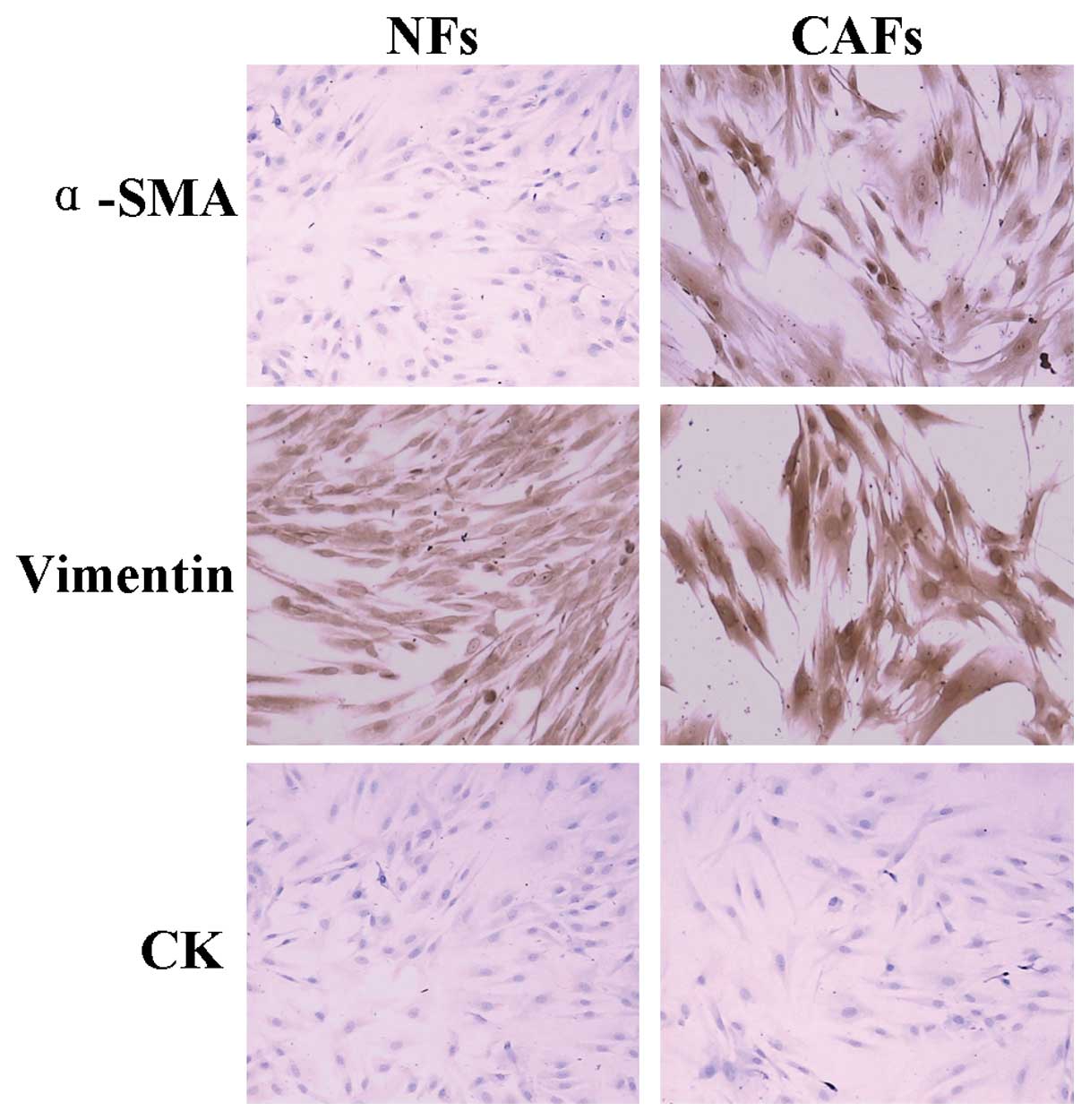
Oral mucosa NFs have an elongated spindle shape and
have no cell-overlapping growth. However, oral tongue squamous CAFs
have a short spindle shape with clear cell protuberances, they are
irregular, multinucleated and have local cell-overlapping growth
(Fig. 1). Immunocytochemical
staining shows NFs are α-SMA negative, while CAFs are α-SMA
positive (Fig. 2).
Cytokine microarray hybridization
Cytokines secreted by CAFs and NFs were measured by
the Human-L Series of protein factors (RayBio), which
simultaneously measures 507 types of protein factors, including
growth factors, interleukins, tumor necrosis factor and chemokines.
The results showed that the following CAF-secreted cytokines
significantly increased: CD40/TNFRSF5 (4.39-fold of NFs), IL-17C
(6.55-fold of NFs), MFRP (16.70-fold of NFs), MIG (19.22-fold of
NFs), NCAM-1/CD56 (4.60-fold of NFs), NeuroD1 (4.42-fold of NFs)
and Smad4 (5.95-fold of NFs). The cytokines which were
significantly reduced were CCL28/VIC (0.09-fold of NFs), GFR α-1
(0.08-fold of NFs), Granzyme A (0.03-fold of NFs), Neuregulin
(0.04-fold of NFs), GRO (0.08-fold of NFs), TSLP (0.09-fold of NFs)
and WISP-1/CCN4 (0.06-fold of NFs) (Fig. 3).
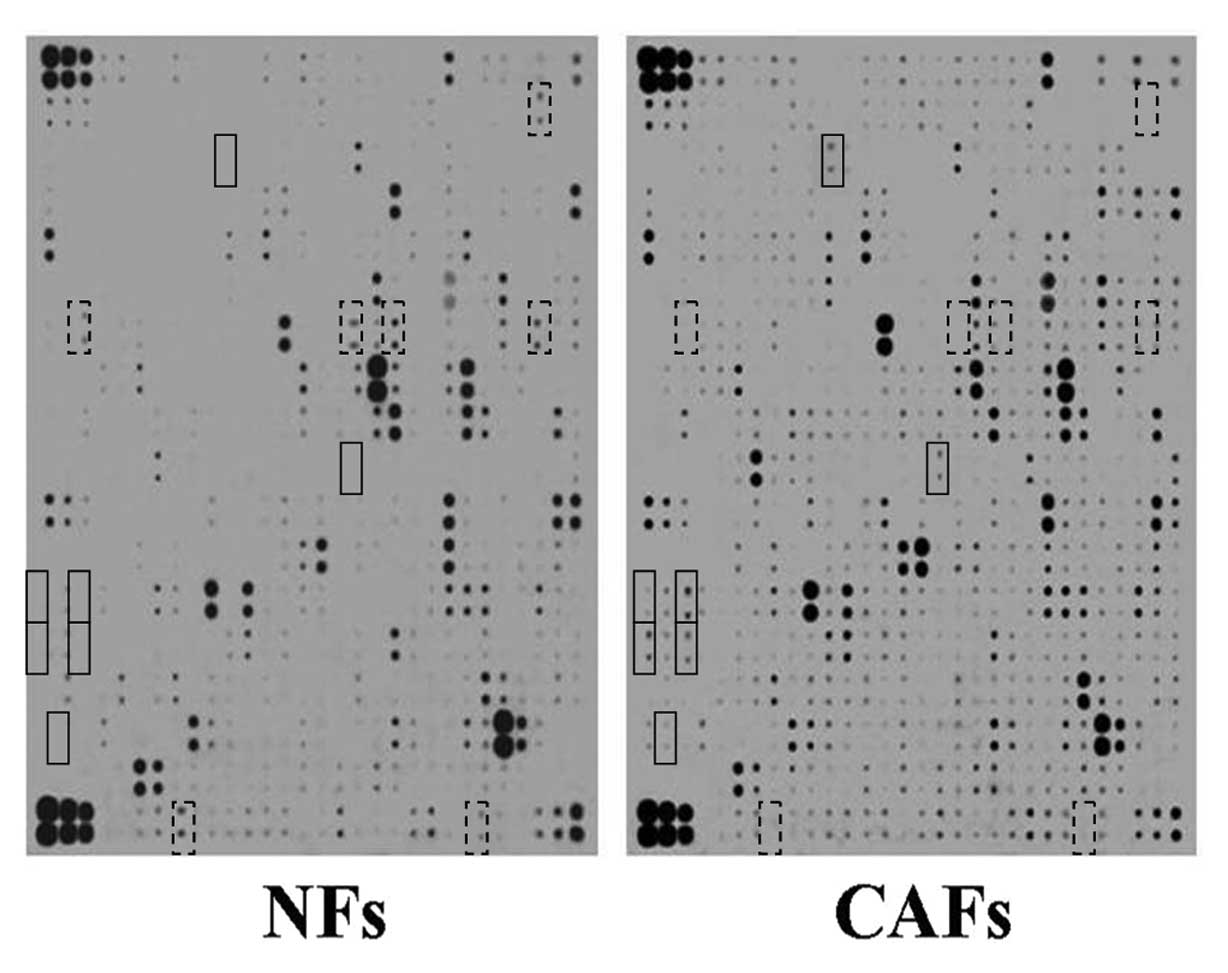 | Figure 3Chemiluminescence of the cytokine
microarray. Hybridization results of NF or CAF supernatant using
RayBio human cytokine antibody microarray. The cytokines which were
significantly increased as shown in the solid line boxes were:
CD40/TNFRSF5 (rows 5 and 6, column 11), IL-17C (rows 19 and 20,
column 17), MFRP (rows 25 and 26, column 1), MIG (rows 25 and 26,
column 3), NCAM-1/CD56 (rows 27 and 28, column 1), NeuroD1 (rows 27
and 28, column 3) and Smad4 (rows 31 and 32, column 2). The
cytokines which were significantly reduced as shown in the dotted
line boxes were: CCL28/VIC (rows 3 and 4, column 27), GFR α-1 (rows
13 and 14, column 3), Granzyme A (rows 13 and 14, column 18), GRO
(row 13 and 14, column 20), Heregulin/NDF/GGF/Neuregulin (rows 13
and 14, column 28), TSLP (rows 35 and 36, column 8) and WISP-1/CCN4
(rows 35 and 36, column 25). |
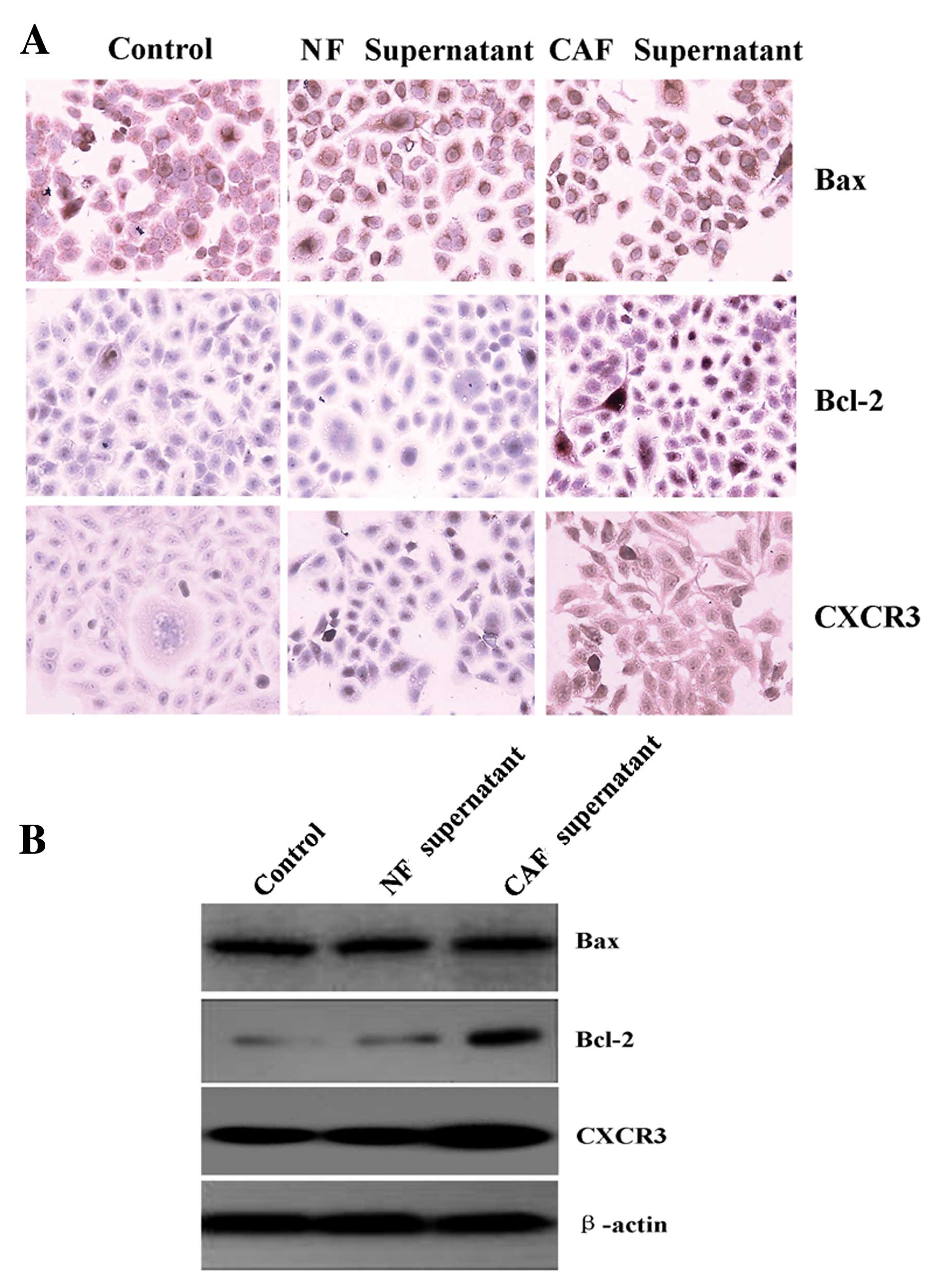
CAF supernatant promotes Tca8113 cell
expression of Bcl-2 and CXCR3
The expression level of Bax in the Tca8113 cells had
no obvious change, after treatment with a 2/6 volume NF- or a
CAF-conditioned medium supernatant. After treatment with a 2/6
volume NF-conditioned medium supernatant, expression levels of
Bcl-2 and CXCR3 in the Tca8113 cells had no obvious changes.
However, after treatment with a 2/6 volume CAF-conditioned medium
supernatant, the expression levels significantly increased
(Fig. 4).
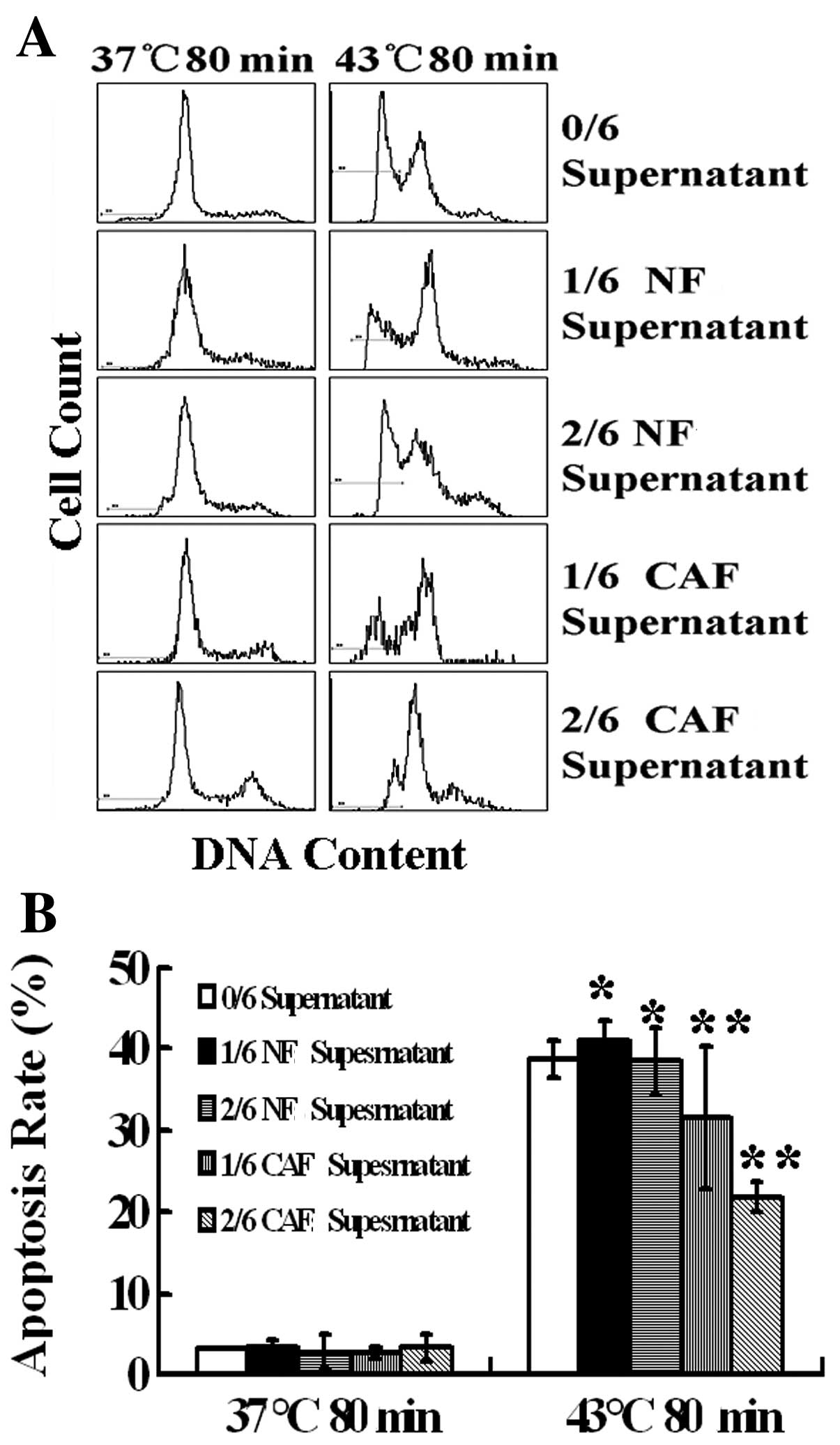
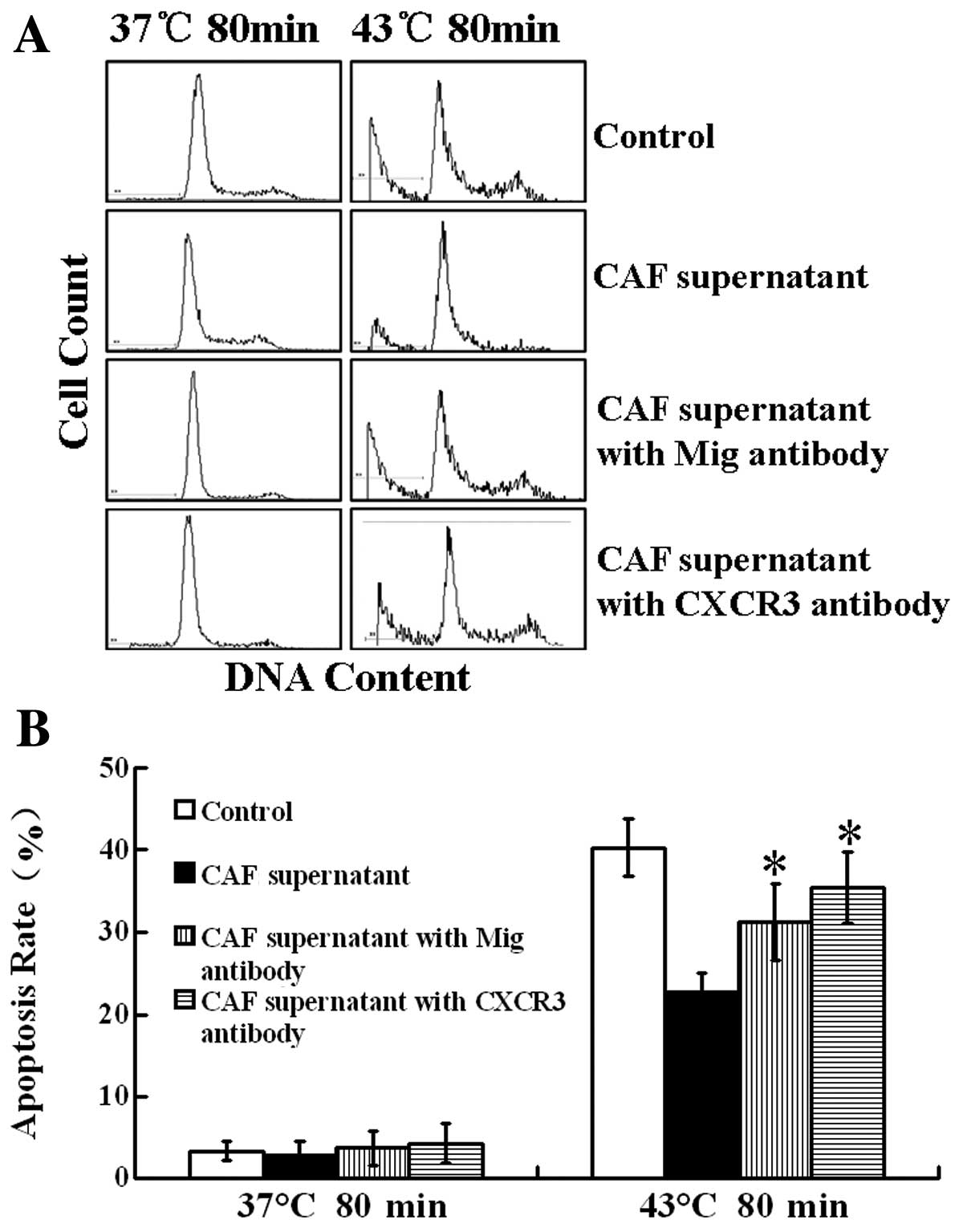
CAF supernatant inhibits heat-induced
Tca8113 cell apoptosis
Heat treatment at 43°C for 80 min induced Tca8113
cell apoptosis, and the apoptosis rate was 38.8% according to
propidium iodide staining combined with flow cytometry. After
treatment with a 1/6 and 2/6 volume of the NF supernatant, the
apoptosis rates were 41.06 and 38.43% (P>0.05), respectively;
thus, there was no significant effect on the heat-induced cell
apoptosis rate. After treatment with a 1/6 volume of CAF
supernatant, the apoptosis rate was 31.6%. After a pretreatment
with a 2/6 volume of CAF supernatant, the rate of apoptosis was
only 21.8% (P<0.05), significantly inhibiting the heat-induced
Tca8113 cell apoptosis (Fig.
5).
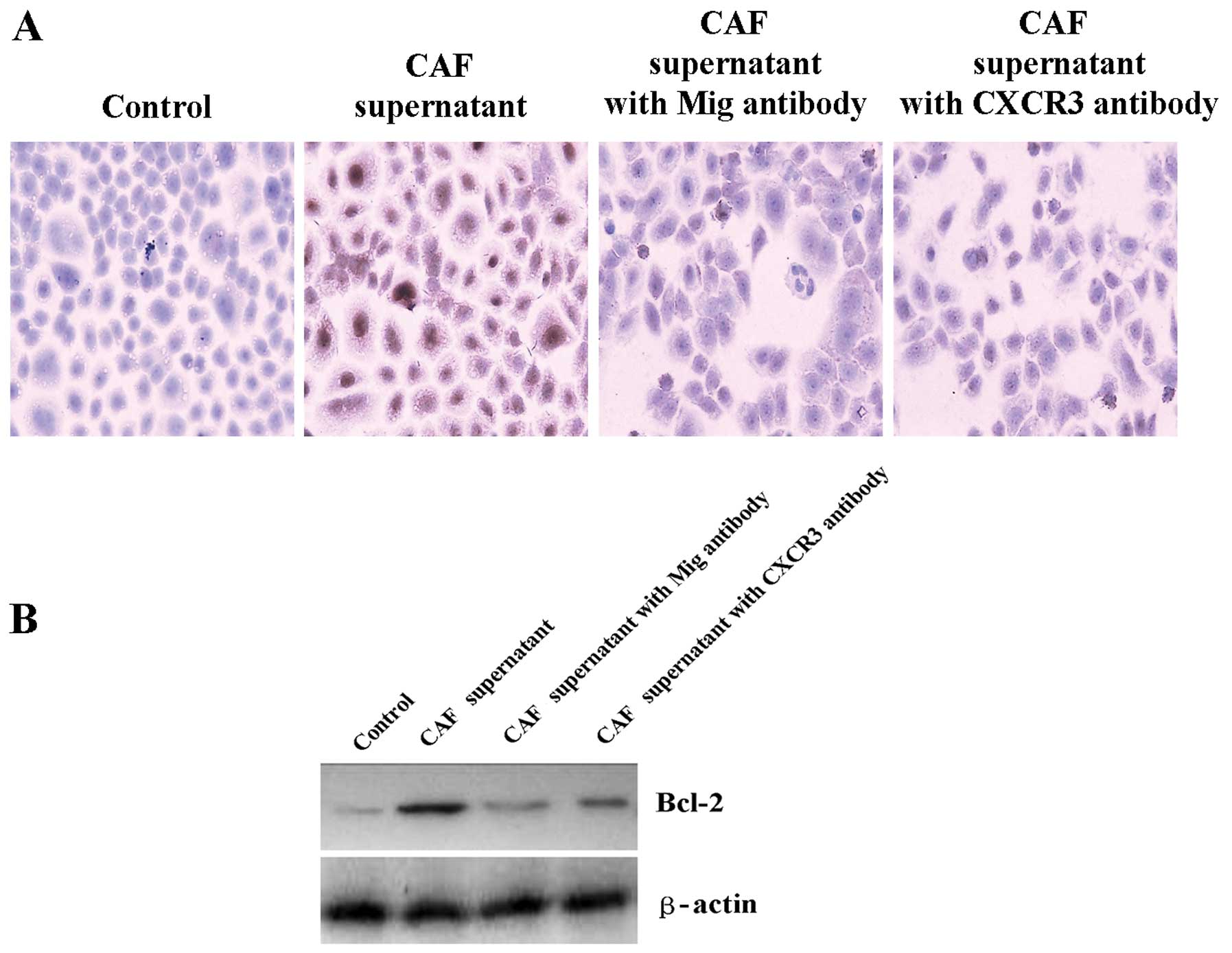
CAFs enhance Bcl-2 expression and inhibit
heat-induced Tca8113 cell apoptosis through the Mig/CXCR3 axis
As mentioned above, the protein level of the Mig
factor in the CAFs was significantly higher than that in the NFs,
and the expression level of CXCR3 in the Tca8113 cells
significantly increased after treatment with the CAF supernatant.
Therefore, we used neutralizing antibodies against the Mig factor
or its receptor CXCR3, treated the Tca8113 cells with a 2/6 volume
of CAF supernatant and tested the expression of the Bcl-2 protein
in the Tca8113 cells using western blot analysis. We tested the
rate of the heat-induced Tca8113 cell apoptosis using flow
cytometry. The results showed that after neutralizing antibodies
against the Mig factor or its receptor CXCR3, enhanced expression
of Bcl-2 protein in Tca8113 and inhibition of heat-induced Tca8113
cell apoptosis in CAF supernatant were significantly reduced. After
a heat treatment at 43°C for 80 min, the heat-induced Tca8113 cell
apoptosis rate was 40.3%. After pretreatment with a 2/6 volume CAF
supernatant and heat treatment at 43°C for 80 min, the apoptosis
rate dropped to 22.6%. After adding neutralizing antibodies against
the Mig factor, the apoptosis rate increased to 31.2%. After adding
a neutralizing antibody against CXCR3, the apoptosis rate increased
to 35.4% (Figs. 6 and 7). These results suggest that CAFs may
enhance Bcl-2 expression and inhibit heat-induced cell apoptosis
through the Mig factor and its receptor CXCR3.
Discussion
A tumor is composed of tumor cells, extracellular
matrix and a vascular system. The majority of tumor stromal cells
are fibroblasts. CAFs produce a tumor extracellular matrix that
provides a suitable microenvironment to support the growth of the
tumor (6,7). Through their interaction with tumor
cells, CAFs alter their morphological and physiological
characteristics, express various cytokines, proteases, adhesion
molecules and increase the malignant phenotype of tumor cells to
different degrees. These events play an important role in cancer
proliferation, development, invasion and metastasis (8).
In this study, we derived CAFs from oral tongue
squamous carcinoma tissue. Compared with NFs, the biological
characteristics of the CAFs were altered. The secreted factors in
the CAFs and NFs were significantly different as determined by the
cytokine microarray test. Compared with the NFs, the levels of the
CAF-secreted cytokine MIG was significantly higher.
Hyperthermia not only induces cell apoptosis but may
also have a direct lethal effect on a tumor. Commonly used
temperatures in clinical hyperthermia are between 40 and 45°C, and
the death of tumor cells is mainly caused by cell apoptosis during
this treatment (17). For many
years, research on the effect and regulation of hyperthermia
treatment has been mainly focused on the tumor cell itself; for
example, examining the relationship between the expression levels
of apoptosis-related proteins and heat shock proteins in tumor
cells and the efficacy of hyperthermia (2,3).
However, the regulation of the thermal destruction of tumor cells
in the stromal microenvironment is rarely investigated. Recent
studies have shown that the incidence of tumor development and
treatment is not simply determined by tumor cells or stroma. It is
closely related to the balance of the tumor-stroma microenvironment
formed through the interaction between both the tumor and the
stroma (4,5). CAFs are the most important host cells
in this microenvironment, and they play an important role in
regulating the balance of the system, which is dependent on direct
cell-to-cell contact, secretion of soluble factors and
extracellular matrix modification (18,19).
Studies have shown that CAFs promote tumor development and enhance
tumor capacity to fight against radiation therapy and chemotherapy
(12–14). In this study, we derived CAFs from
oral tongue squamous carcinoma, established an in vitro
co-culture model for CAFs and Tca8113 cells and found that the CAF
supernatant significantly inhibited heat-induced Tca8113 cell
apoptosis. Heat treatment at 43°C for 80 min induced Tca8113 cell
apoptosis, and the apoptosis rate was 38.8%. After treatment with a
1/6 or 2/6 volume NF supernatant, there was no significant effect
on the heat-induced cell apoptosis rate of Tca8113. However, after
treatment with a 1/6 volume CAF supernatant, the rate was impacted
and lowered to 31.6%. A pretreatment with a 2/6 volume CAF
supernatant significantly inhibited the heat-induced Tca8113 cell
apoptosis and the rate of apoptosis was only 21.8%. These results
suggest that CAFs significantly enhance the ability of cancer cells
to fight against thermal destruction. Furthermore, the tumor
microenvironment is another important factor in determining the
efficacy of hyperthermia.
A high temperature also inhibits the expression of
the anti-apoptotic genes, such as Bcl-2, activates
apoptosis-related pathways and induces cell apoptosis (1,17,20,21).
Interference and disruption of a hyperthermia-induced apoptosis
pathway may induce tumor cells which are no longer sensitive to the
stimulation of cell apoptosis. Our previous studies found that the
intracellular expression of Bax and Bcl-2 is one of the intrinsic
factors determining the efficacy of hyperthermia. In this study, we
tested the expression levels of Bax and Bcl-2 in Tca8113 cells
after treatment with CAF and NF supernatants, and we found that the
expression level of Bax was not affected. We also found that the
expression level of Bcl-2 after the NF treatment was not affected
whereas the level was significantly enhanced after the CAF
treatment. These results suggest that CAFs promote the expression
of an anti-apoptotic gene, Bcl-2, enhance tumor resistance to
heat-induced apoptosis and mediate thermal tolerance.
As mentioned above, the secreted cytokines of CAFs
are significantly different from the secreted cytokines of NFs.
CAFs can mediate the heat tolerance of cancer cells dependent on
these factors. Comparing secreted cytokines of oral cancer CAFs
with those of NFs shows that the quality of the Mig factor, one of
the chemokines, is clearly increased by 19-folds compared with that
in NFs. Recent studies have shown that the chemokine receptor,
CXCR3, is highly expressed in a variety of tumor cells.
Furthermore, specific binding of CXCR3 with its ligand chemokine
Mig (CXCL9), IP-10 (CXCL10) or I-TAC (CXCL11) regulates tumor
invasion, metastasis, survival and proliferation (22–24).
The results from tissue-specific analysis and research among
various types of tumors are varied, which may be related to
different shearing variants of CXCR3 and the culture conditions of
in vitro studies, such as the presence or absence of serum
(25). In this study, we found that
Tca8113 cells expressed CXCR3, and the expression level of CXCR3 in
the Tca8113 cells increased after treatment with the CAF
supernatant. Furthermore, we treated the Tca8113 cells with the CAF
supernatants while using neutralizing antibodies against the Mig
factor or its receptor CXCR3, and we found that this enhanced the
expression of the Bcl-2 protein in Tca8113 cells and that the
inhibition of heat-induced Tca8113 cell apoptosis in the CAF
supernatant was significantly reduced.
In summary, secreted cytokines of oral tongue
squamous cell carcinoma CAFs are significantly different from NFs.
CAFs increase the expression level of the Bcl-2 protein in Tca8113
cells, enhance resistance to heat-induced cancer cell apoptosis and
mediate thermal tolerance. Further studies should investigate
whether or not CAFs initiate cancer-related downstream signaling
pathways and regulate the expression and activation of target
proteins to regulate heat-induced tumor cell apoptosis by secreting
the Mig chemokine factor to directly bind the CXCR3 receptor of
cancer cells, the Mig/CXCR3 paracrine axis.
Acknowledgements
This research was supported in part by grants from
the National Natural Science Foundation of China (nos. 81160325,
30760272 and 30960422, to Y.W.H.) and the Project of the Department
of Science and Technology of Yunnan Province (no. 2009CC021, to
Y.W.H.).
References
|
1
|
Kajihara A, Takahashi A, Ohnishi K, Imai
Y, Yamakawa N, Yasumoto J, et al: Protein microarray analysis of
apoptosis-related protein expression following heat shock in human
tongue squamous cell carcinomas containing different p53
phenotypes. Int J Hyperthermia. 24:605–612. 2008. View Article : Google Scholar
|
|
2
|
Setroikromo R, Wierenga PK, van Waarde MA,
Brunsting JF, Vellenga E and Kampinga HH: Heat shock proteins and
Bcl-2 expression and function in relation to the differential
hyperthermic sensitivity between leukemic and normal hematopoietic
cells. Cell Stress Chaperones. 12:320–330. 2007. View Article : Google Scholar
|
|
3
|
Shelton SN, Dillard CD and Robertson JD:
Activation of caspase-9, but not caspase-2 or caspase-8, is
essential for heat-induced apoptosis in Jurkat cells. J Biol Chem.
285:40525–40533. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Karnoub AE, Dash AB, Vo AP, Sullivan A,
Brooks MW, Bell GW, et al: Mesenchymal stem cells within tumour
stroma promote breast cancer metastasis. Nature. 449:557–563. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sautes-Fridman C, Cherfils-Vicini J,
Damotte D, Fisson S, Fridman WH, Cremer I, et al: Tumor
microenvironment is multifaceted. Cancer Metastasis Rev. 30:13–25.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ostman A and Augsten M: Cancer-associated
fibroblasts and tumor growth-bystanders turning into key players.
Curr Opin Genet Dev. 19:67–73. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xing F, Saidou J and Watabe K: Cancer
associated fibroblasts (CAFs) in tumor microenvironment. Front
Biosci. 15:166–179. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Angeli F, Koumakis G, Chen MC, Kumar S and
Delinassios JG: Role of stromal fibroblasts in cancer: promoting or
impeding? Tumour Biol. 30:109–120. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin JW, Chen QM, Li SF, Song HY, Long D
and Zhou HM: The study on oral carcinoma-associated fibroblast
promoting the proliferation of lingual carcinoma cell line. Sichuan
Da Xue Xue Bao Yi Xue Ban. 39:184–187. 2008.(In Chinese).
|
|
10
|
Olsen CJ, Moreira J, Lukanidin EM and
Ambartsumian NS: Human mammary fibroblasts stimulate invasion of
breast cancer cells in a three-dimensional culture and increase
stroma development in mouse xenografts. BMC Cancer. 10:4442010.
View Article : Google Scholar
|
|
11
|
Paland N, Kamer I, Kogan-Sakin I, Madar S,
Goldfinger N and Rotter V: Differential influence of normal and
cancer-associated fibroblasts on the growth of human epithelial
cells in an in vitro cocultivation model of prostate cancer. Mol
Cancer Res. 7:1212–1223. 2009. View Article : Google Scholar
|
|
12
|
Hwang RF, Moore T, Arumugam T,
Ramachandran V, Amos KD, Rivera A, et al: Cancer-associated stromal
fibroblasts promote pancreatic tumor progression. Cancer Res.
68:918–926. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Muerkoster S, Wegehenkel K, Arlt A, Witt
M, Sipos B, Kruse ML, et al: Tumor stroma interactions induce
chemoresistance in pancreatic ductal carcinoma cells involving
increased secretion and paracrine effects of nitric oxide and
interleukin-1β. Cancer Res. 64:1331–1337. 2004.PubMed/NCBI
|
|
14
|
Shekhar MP, Santner S, Carolin KA and Tait
L: Direct involvement of breast tumor fibroblasts in the modulation
of tamoxifen sensitivity. Am J Pathol. 170:1546–1560. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cohen SJ, Alpaugh RK, Palazzo I, Meropol
NJ, Rogatko A, Xu Z, et al: Fibroblast activation protein and its
relationship to clinical outcome in pancreatic adenocarcinoma.
Pancreas. 37:154–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsujino T, Seshimo I, Yamamoto H, Ngan CY,
Ezumi K, Takemasa I, et al: Stromal myofibroblasts predict disease
recurrence for colorectal cancer. Clin Cancer Res. 13:2082–2090.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang W, Bian L, Ma LJ, Tang RZ, Xun S and
He YW: Hyperthermia-induced apoptosis in Tca8113 cells is inhibited
by heat shock protein 27 through blocking phospholipid scramblase 3
phosphorylation. Int J Hyperthermia. 26:523–537. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Grugan KD, Miller CG, Yao Y, Michaylira
CZ, Ohashi S, Klein-Szanto AJ, et al: Fibroblast-secreted
hepatocyte growth factor plays a functional role in esophageal
squamous cell carcinoma invasion. Proc Natl Acad Sci USA.
107:11026–11031. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Orimo A, Gupta PB, Sgroi DC,
Arenzana-Seisdedos F, Delaunay T, Naeem R, et al: Stromal
fibroblasts present in invasive human breast carcinomas promote
tumor growth and angiogenesis through elevated SDF-1/CXCL12
secretion. Cell. 121:335–348. 2005. View Article : Google Scholar
|
|
20
|
Alcala MA Jr, Park K, Yoo J, Lee DH, Park
BH, Lee BC, et al: Effect of hyperthermia in combination with TRAIL
on the JNK-Bim signal transduction pathway and growth of xenograft
tumors. J Cell Biochem. 110:1073–1081. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang W, Bian L, Li GQ, Ma LJ, Tang RZ and
He YW: Role of protein kinase C-delta in hyperthermia-induced
apoptosis in tongue squamous cell carcinoma Tca8113 cells. Hua Xi
Kou Qiang Yi Xue Za Zhi. 28:539–542. 5462010.(In Chinese).
|
|
22
|
Lo BK, Yu M, Zloty D, Cowan B, Shapiro J
and McElwee KJ: CXCR3/ligands are significantly involved in the
tumorigenesis of basal cell carcinomas. Am J Pathol. 176:2435–2446.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ma X, Norsworthy K, Kundu N, Rodgers WH,
Gimotty PA, Goloubeva O, et al: CXCR3 expression is associated with
poor survival in breast cancer and promotes metastasis in a murine
model. Mol Cancer Ther. 8:490–498. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pradelli E, Karimdjee-Soilihi B, Michiels
JF, Ricci JE, Millet MA, Vandenbos F, et al: Antagonism of
chemokine receptor CXCR3 inhibits osteosarcoma metastasis to lungs.
Int J Cancer. 125:2586–2594. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Datta D, Banerjee P, Gasser M,
Waaga-Gasser AM and Pal S: CXCR3-B can mediate growth-inhibitory
signals in human renal cancer cells by down-regulating the
expression of heme oxygenase-1. J Biol Chem. 285:36842–36848. 2010.
View Article : Google Scholar : PubMed/NCBI
|