Introduction
Despite the advances in combinatorial chemistry, 60%
of the agents used for cancer treatment are substances of natural
origin or derivatives (1).
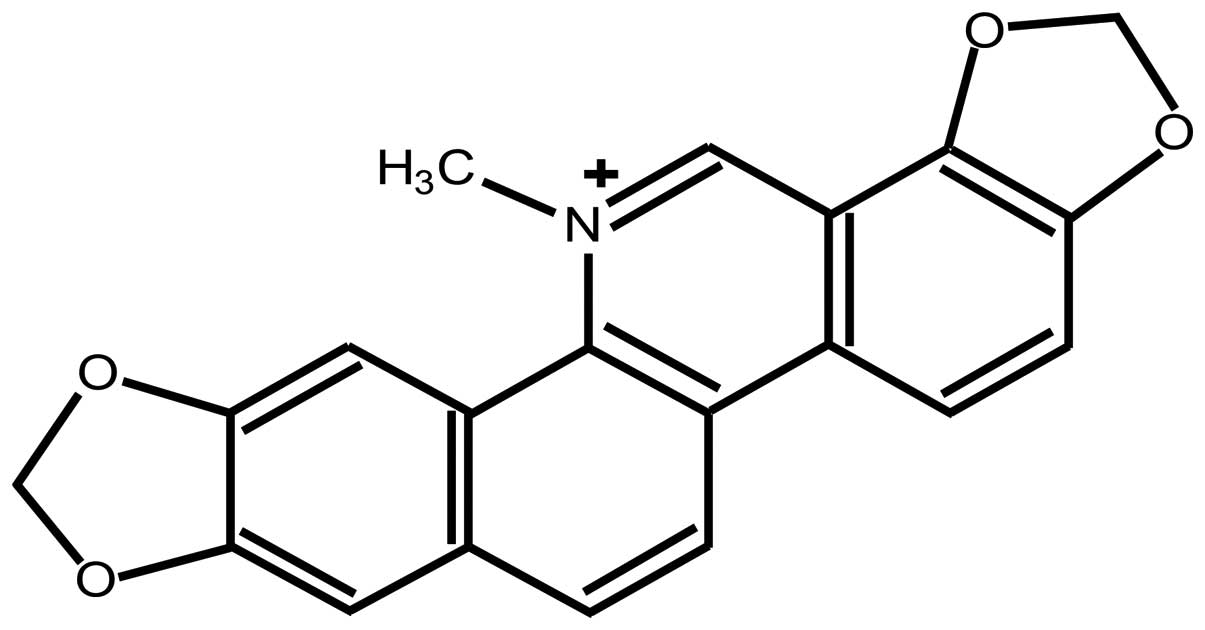
Sanguinarine [13-methyl(1,3)benzodioxolo(5,6-c)-1,3-dioxolo(4,5)
phenanthridinium](Fig. 1), derived
from the root of sanguinaria canadensis and other
poppy-fumaria species, is the most widely-used benzophenanthridine
alkaloid. It has been used in many over-the-counter products such
as toothpaste, mouthwash, cough and cold remedies due to its
antimicrobial, antioxidant and anti-inflammatory properties
(2,3). The cytotoxic and cytostatic effects of
sanguinarine on a variety of human cancer cells, including human
epidermoid carcinoma, erythroleukemia, prostate cancer, pancreatic
carcinoma, colon cancer, breast cancer, lung cancer, promyelocytic
leukemia and bone cancer (4–13),
have been reported. Sanguinarine exhibits the highest cytotoxicity
among benzophenanthridine alkaloids(14). Markedly, sanguinarine results in
antiproliferative effects on human epidermoid carcinoma cells
(A431), but not in the normal human epidermal keratinytes (4). This different anti-proliferative
response is also observed in a study where mouse lymphocytic
leukemic cells were significantly more sensitive to sanguinarine
compared with normal spleen cells (15). Thus, sanguinarine has gained
increasing attention as a potential agent in the treatment of
cancer.
Cervical cancer is the second most common cancer in
women worldwide, and its incidence is disproportionately high
(>80%) in the developing countries (16). The major cause of mortality from
this disease is metastatic cancer cells that fail to respond to
chemoradiation therapy (17). Since
sanguinarine possesses the potential ability for cancer treatment,
the availability of sanguinarine in advanced, recurrent, metastatic
cervical cancer merits further research. To the best of our
knowledge, the effect of sanguinarine on cervical carcinoma has yet
to be studied extensively, despite discovering that sanguinarine
caused apoptosis in HeLa cells (18,19)
when compared with other benzophenanthridine alkaloids. Thus, a
systemic study is required to elucidate the mechanism by which
sanguinarine inhibits cell growth in some types of cervical cancer
cells.
Sanguinarine-induced apoptosis, which is the process
of programmed cell death, is the key issue in cancer cells
mentioned above. The Bax and Bcl-2 proteins belong to the Bcl-2
family, which is the best-characterized group of
apoptosis-regulating factors. The Bcl-2 family can be divided into
two main groups according to their functional properties: the
anti-apoptotic proteins such as Bcl-2 and Bcl-XL, and the
pro-apoptotic proteins, such as Bax, Bak and Bad. Sanguinarine
decreased anti-apoptotic Bcl-2 and increased the pro-apoptotic Bax
protein in a dose-dependent pattern in K562 erythroleukemia cells
(5,6), human pancreatic carcinoma AsPC-1 and
BxPC-3 cells (8), and immortalized
human HaCaT keratinytes (20).
The transcription factor NF-κB is retained normally
in an inactive form in the cytoplasm through interaction with IκB
(inhibitor κB) family proteins and can be activated by various
inflammatory and stress stimuli (21). NF-κB activation requires nuclear
translocation. Active nuclear form of the NF-κB transcription
factor complex is composed of two DNA binding subunits, NF-κB p65
and p50. Studies have indicated that NF-κB promotes cell survival
by inhibiting apoptosis and NF-κB has been recognized as an
anti-apoptotic regulator. The decreased expression of NF-κB by
sanguinarine has been observed in K562 erythroleukemia cells
(5) and human myeloid ML-1a cells
(21).
During the last decade, the incidence of cervical
adenocarcinoma has increased, particularly in young women (22). In the present study, the
antiproliferative and apoptosis-inducing effects of sanguinarine
were investigated in cervical adenocarcinoma HeLa and cervical
squamous cell carcinoma SiHa cells.
Materials and methods
Reagents and antibodies
Sanguinarine (>99.0%
pure),3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide(MTT), dimethylsulfoxide (DMSO), protein lysis buffer, R
Nase A and proteinase K were purchased from Sigma-Aldrich (St.
Louis, MO, USA). Sanguinarine was dissolved to a stock solution of
20 mmol/l in DMSO and directly diluted in medium to appropriate
concentrations before the experiments. Primary antibodies against
Bcl-2, Bax and NF-κB and secondary antibodies were obtained from
Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell culture and treatment
HeLa and SiHa cells were purchased from the American
Type Culture Collection (ATCC, Manassas, VA, USA) and maintained in
Dulbecco’s modified Eagle’s medium (D-MEM; Invitrogen, Carlsbad,
CA, USA) supplemented with 10% FBS, 100 U/ml penicillin, 100 μg/ml
streptomycin, 4 mmol/l L-glutamine, and nonessential amino acids at
37°C in a 5% CO2 humid environment. Unless otherwise
indicated, cells were plated in 100-mm Petri dishes at a density of
2×106 cells/dish.
MTT assay
Cell viability was determined by MTT assay. Cells
were plated at 1×105 cells/well in 200 μl D-MEM
containing sanguinarine in a 96-well microtiter plate. Each
concentration was repeated in 6 wells. After 12, 24, 36, 48, 60 h
of incubation at 37°C in a humidified chamber, 20 μl MTT (5 mg/ml
in PBS) was added to each well and further incubated for 4 h. Then,
the microtiter plate containing the cells was centrifuged at 1,800
rpm for 5 min at 4°C. MTT solution was removed from the wells by
aspiration and formazan crystals were dissolved in 150 μl DMSO.
Absorbance was measured in an ELISA reader at 570 nm, with the
absorbance at 630 nm to correct for background. The effect of
sanguinarine on growth inhibition was assessed as percentage
inhibition in cell growth where untreated cells were taken as
100%.
Clonogenic assay
Cell survival was assessed using the colony
formation assay. Briefly, cells were treated with 0, 0.5, 1, 2 and
3 μmol/l sanguinarine for 12 h and collected immediately by
trypsinization. The cells were counted and re-plated in 60-mm
tissue culture dishes in 2 sets of triplicates for each
concentration with 500, 1000, 2000, 5000 cells/well. Sufficient
numbers were seeded to ensure that 50~100 macroscopic colonies
would appear in untreated cells at the end of 14 days. Media were
replaced every 3 days. At the end, cells were stained with 0.5%
crystal violet in 1:1 methanol-water. Fifty cells were used as
minimum number to define a colony and colonies were counted under a
light microscope.
Cell morphology
Alteration of cell morphology was observed by an
inverted microscope (Olympus Corporation, Tokyo, Japan). The key
morphological criterion for the detection of apoptosis was the
formation of apoptotic bodies according to other reports (4,5,18).
TUNEL assay
Terminal deoxynucleotidyl transferase-mediated
dUTP-biotin nick end labeling (TUNEL) assay was carried out using
the In Situ Cell Death Detection kit according to the
manufacturer’s instructions (Roche Diagnostics, Indianapolis, IN,
USA). Briefly, cells were fixed in 4% paraformaldehyde and
permeabilized in ice-cold 0.1% triton X-100, 0.1% sodium citrate.
TUNEL reaction mix was added at 1:5 dilution of TUNEL reaction mix
and incubated for 1 h at 37°C in the dark under humidified
conditions. TUNEL-Peroxidase (POD) converter was then added and
incubated for an additional hour. Fluorescein-based TSA
fluorescence system (PerkinElmer Life Sciences, Waltham, MA, USA)
was added for 20 min before visualizing with a Nikon microscope
using a 20×0.50 objective and photographed with a camera.
TUNEL-positive cells were counted using particle analysis and cell
counter plug-ins with Image software.
DNA fragmentation gel electrophoresis
assay
Briefly, cells were exposed to sanguinarine for 24 h
and then lysed with 200 μl lysis buffer [10 mM Tris-HCl (pH 7.6),
100 mmol/l EDTA, and 20 mmol/l NaCl] for 30 min. Following
centrifugation, the supernatant was transferred into new tubes and
incubated by adding 20 μl SDS and 200 μl R Nase A (10 mg/ml) for 2
h at 56°C, 30 μl proteinase K (50 mg/ml) was added to each tube.
The cell lysates were incubated for another 2 h at 37°C. The DNA
was finally precipitated with the addition of 10 μl of 10 M
potassium acetate and 1 ml 100% ethanol at −80°C for 30 min. The
extracted DNA samples were centrifuged and washed with 70% ethanol.
Pure DNA was finally loaded and run on a 1% agarose gel at 80 v in
running buffer (89 mmol/l Tris-acetate, 2 mmol/l Na2EDTA, and 89
mmol/l boric acid). The gels were stained with 0.1 μg/ml ethidium
bromide and photographed. A DNA molecular weight ladder
(Invitrogen) was included as a molecular size marker. Gel images
were evaluated for typical ladder patterns of low molecular weight
DNA fragments in multiples of 180–200 base pairs, a hallmark of
apoptosis. The bands were visualized under a UV transilluminator
(Model TM-36; UVP Inc., San Gabriel, CA, USA) followed by Polaroid
photography in MP-4 Photographic System (Fotodyne Inc ., Hartland,
WI, USA).
Cell cycle analysis
Cells were harvested, centrifuged, incubated with 5
μl R Nase (20 μg/ml final concentration) for 30 min and propidium
iodide (50 μg/ml final concentration) for 1 h. Cell cycle analysis
was carried out by FACScan benchtop cytometry (BD Biosciences, San
Jose, CA, USA), Cell Quest software (BD Biosciences) and ModFit LT
software (Verity Software House, Topsham, ME).
Western blot assay
Western blot assay was performed as previously
described (23). Briefly, cells
were harvested and lysed with a lysis buffer containing [50 mmol/l
HEPES (pH 7.5), 150 mmol/l NaCl, 1.5 mmol/l MgCl, 1 mmol/l EDTA,
10% glycerol and 1% Triton X-100] containing protease inhibitors
(phenylmethylsulfonyl fluoride, leupeptin, aprotinin, and trypsin
inhibitor). Protein concentration was determined by Bio-Rad protein
assay (Invitrogen). Total lysates (50 μg) were separated on
SDS-PAGE and electroblotted onto nitrocellulose membranes
(Millipore, Billerica, MA, USA). Following overnight incubation
with the appropriate primary antibodies, blots were treated with
horseradish peroxidase conjugated secondary antibodies.
Chemiluminescence detection was finally performed with a
chemiluminescence (ECL) system (Amersham Biosciences, Amersham,
UK).
Statistical analysis
Assays were performed in triplicate for each sample.
Average value was determined and expressed as the mean ± SD.
Statistical analysis was performed by means of multifactorial ANOVA
and SPSS software for Windows (11.0, Chicago, IL, USA). P<0.05
was considered to indicate statistically significant
differences.
Results
Sanguinarine inhibits viability of HeLa
and SiHa cells
We first assessed the effect of sanguinarine on the
proliferation of two human cervical cancer cell lines, HeLa and
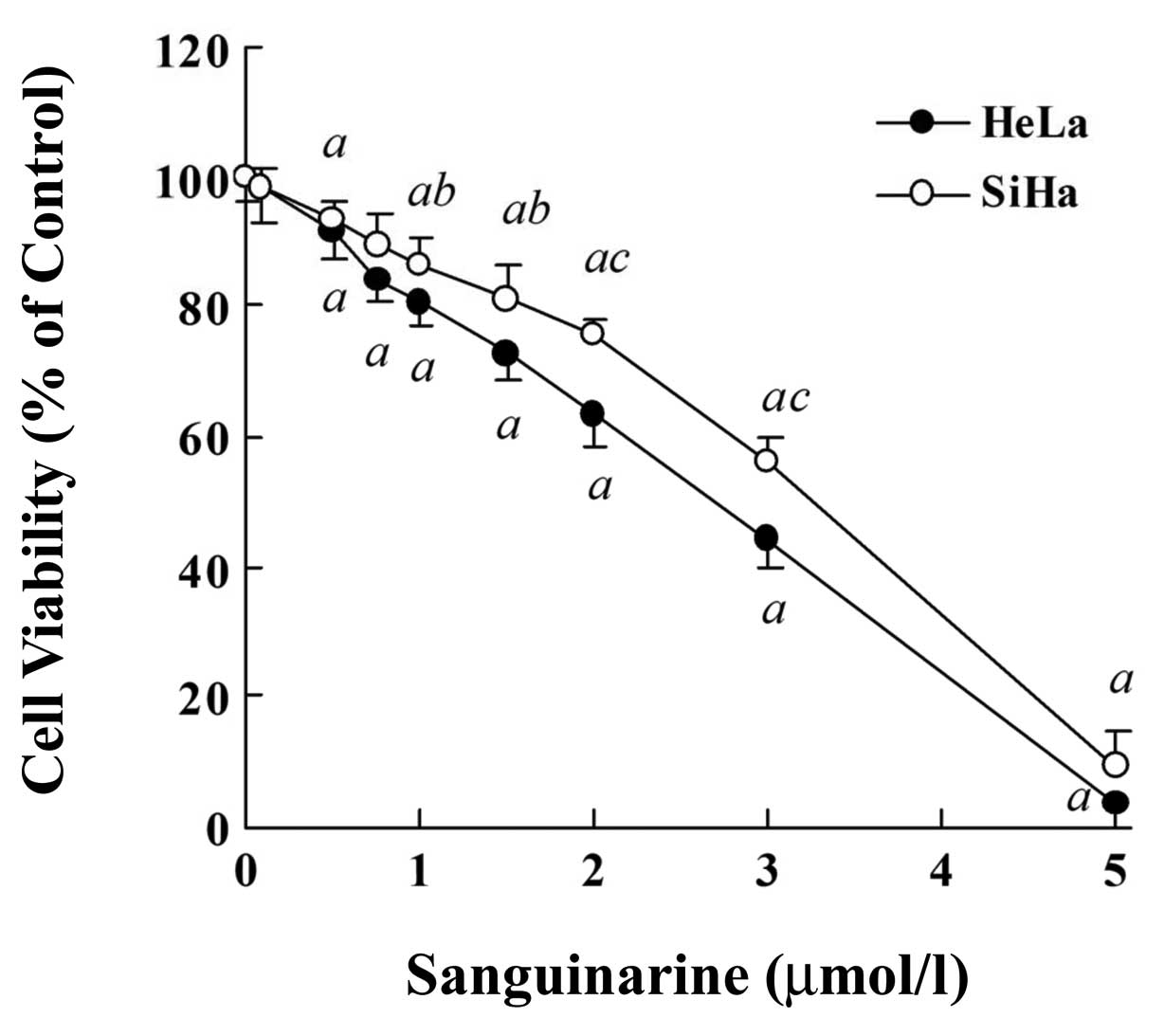
SiHa, by MTT assays. As shown in Fig.
2, a significant dose-dependent inhibition of cell
proliferation was observed in HeLa and SiHa cells exposed to
sanguinarine (24 h), >90% cells lost their viability following
24 exposure to 5 μmol/l sanguinarine in both cell lines. An
IC50 value was 2.62±0.21 μmol/l in HeLa cells and
3.07±0.23 μmol/l in SiHa cells (P<0.05), respectively,
indicating that HeLa cells were slightly more sensitive than SiHa
cells. Similar results were obtained by trypan blue dye exclusion
assay, which was used to enumerate the proportion of live and dead
cells in a population (data not shown).
Sanguinarine inhibits cell colony
formation
To assess the reproductive potential and long-term
cell survival, the effect of sanguinarine on cell ability to form
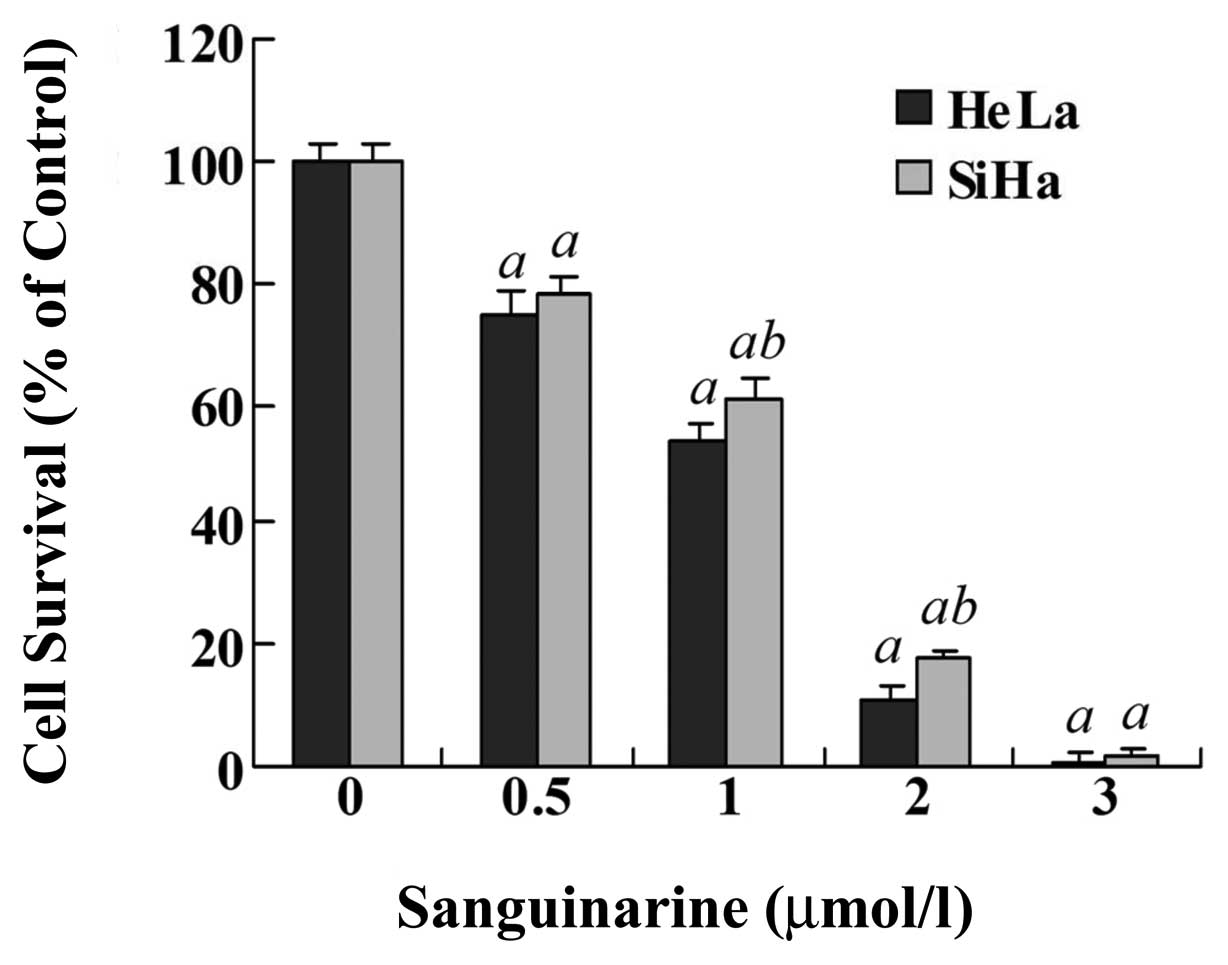
colonies was detected. As shown in Fig.
3, sanguinarine resulted in a dose-dependent inhibition of
colony formation in HeLa and SiHa cells. For example, 1.0 μmol/l
sanguinarine clearly decreased the colony formation to 53.5% in
HeLa cells and to 61.0% in SiHa cells. Similarly, sanguinarine was
more effective in HeLa cells than in SiHa cells (P<0.05).
Sanguinarine induces apoptosis in HeLa
and SiHa cells
We determined whether the decrease of cell viability
and colony formation by sanguinarine was via the occurrence of
apoptosis. For this purpose, apoptotic cell death was examined by
cell morphology, DNA fragmentation and flow cytometry assays. Using
light microscopy, apoptotic bodies were observed in HeLa and SiHa
cells treated with sanguinarine at various doses. The cytosolic
condensation and nuclear condensation, typical of apoptotic cell
death, were observed in the cells treated with sanguinarine at all
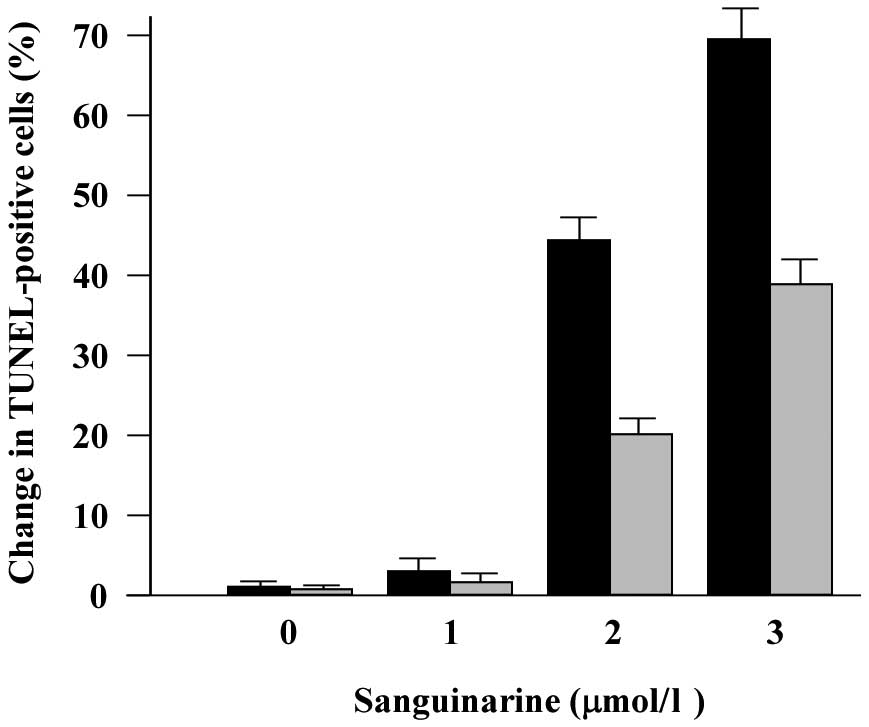
doses tested (data not shown). Moreover, sanguinarine induced a
significant increase in the number of TUNEL-labeled apoptotic cells
in a dose-dependent manner, and the change of TUNEL-positive cell
numbers in HeLa cells was evident compared with SiHa cells
following sanguinarine treatment at the same dose (Fig. 4).
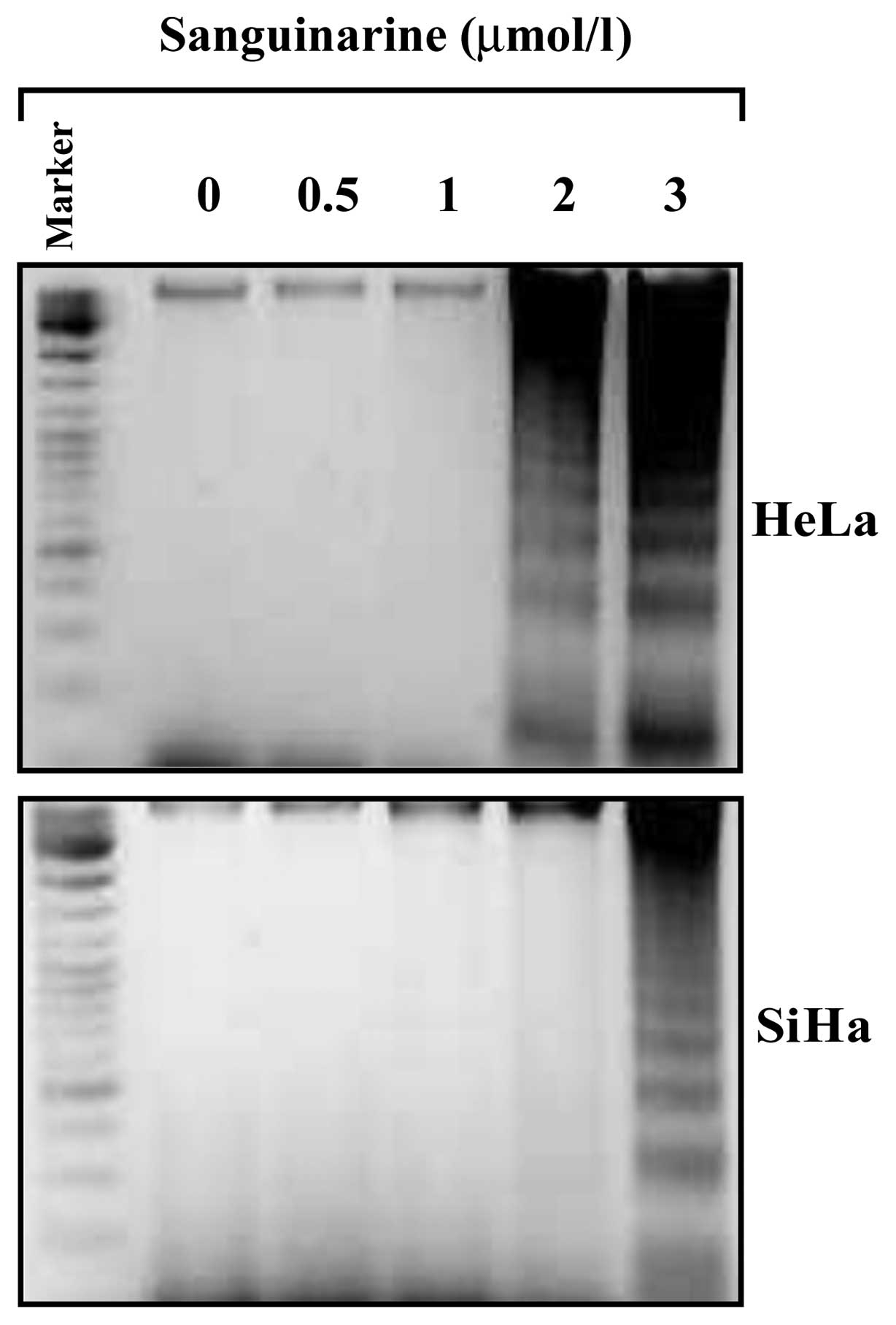
Apoptosis was also assessed by a classical DNA
fragmentation assay. As shown in Fig.
5, fragmented DNAs were observed in HeLa cells treated with 2.0
μmol/l sanguinarine and became more obvious in cells treated with
3.0 μmol/l sanguinarine. DNA fragmentation was also observed in
SiHa cells treated with sanguinarine at 3.0 μmol/l, but not at 2.0
μmol/l.
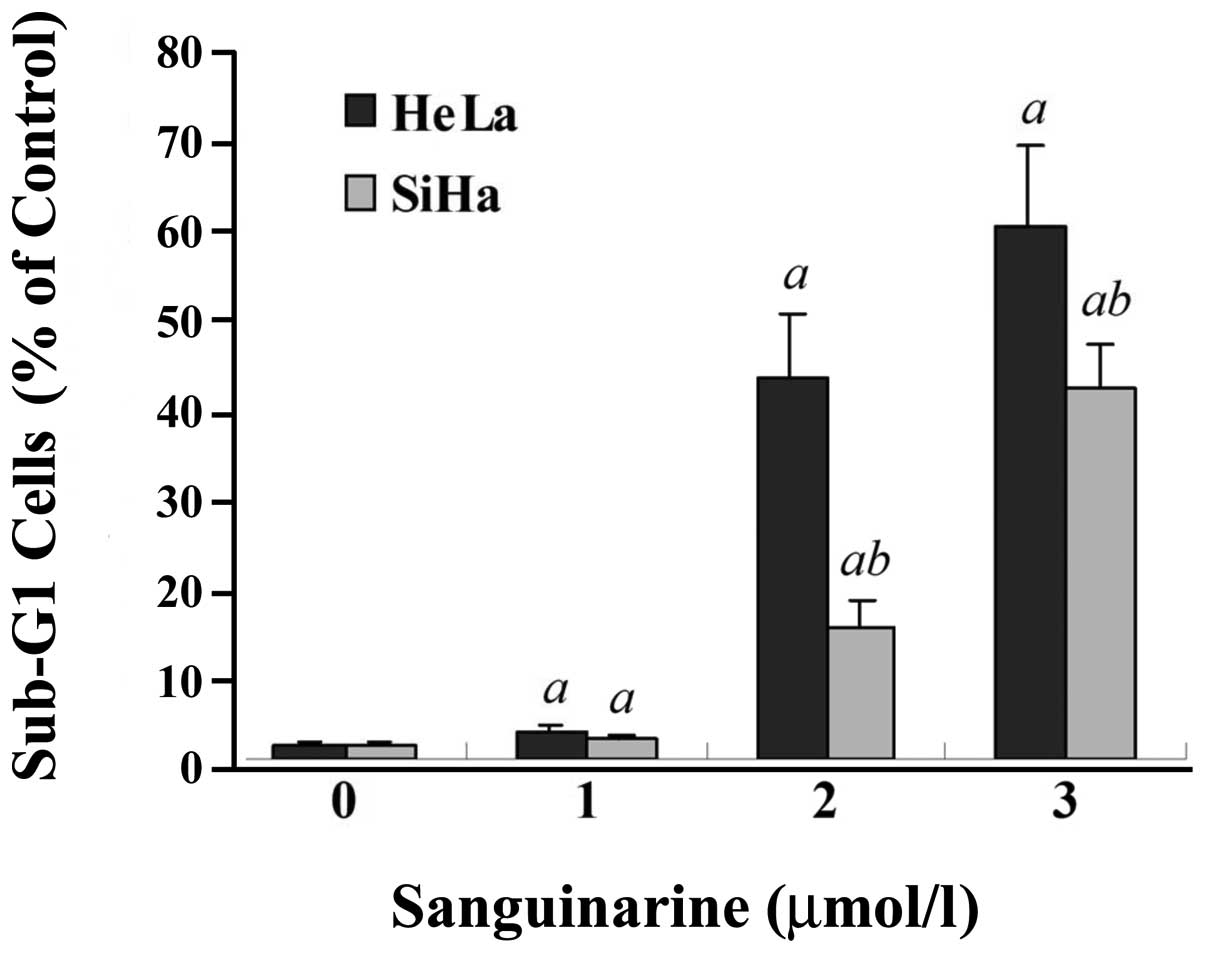
Additionally, as evidenced by flow cytometry, a
significant increase of sub-G1 fraction, a characteristic of
apoptotic cells, on a DNA histogram was also observed in both HeLa
and SiHa cell lines exposed to sanguinarine (Fig. 6). A larger Sub-G1 population was
seen in HeLa than in SiHa cells at the same concentration of
sanguinarine (P<0.01); for example, in HeLa cells treated with
2.0 and 3.0 μmol/l sanguinarine, the sub-G1 fraction increased from
1.7% (untreated control) to 42.5 and 59.7% (P<0.01),
respectively.
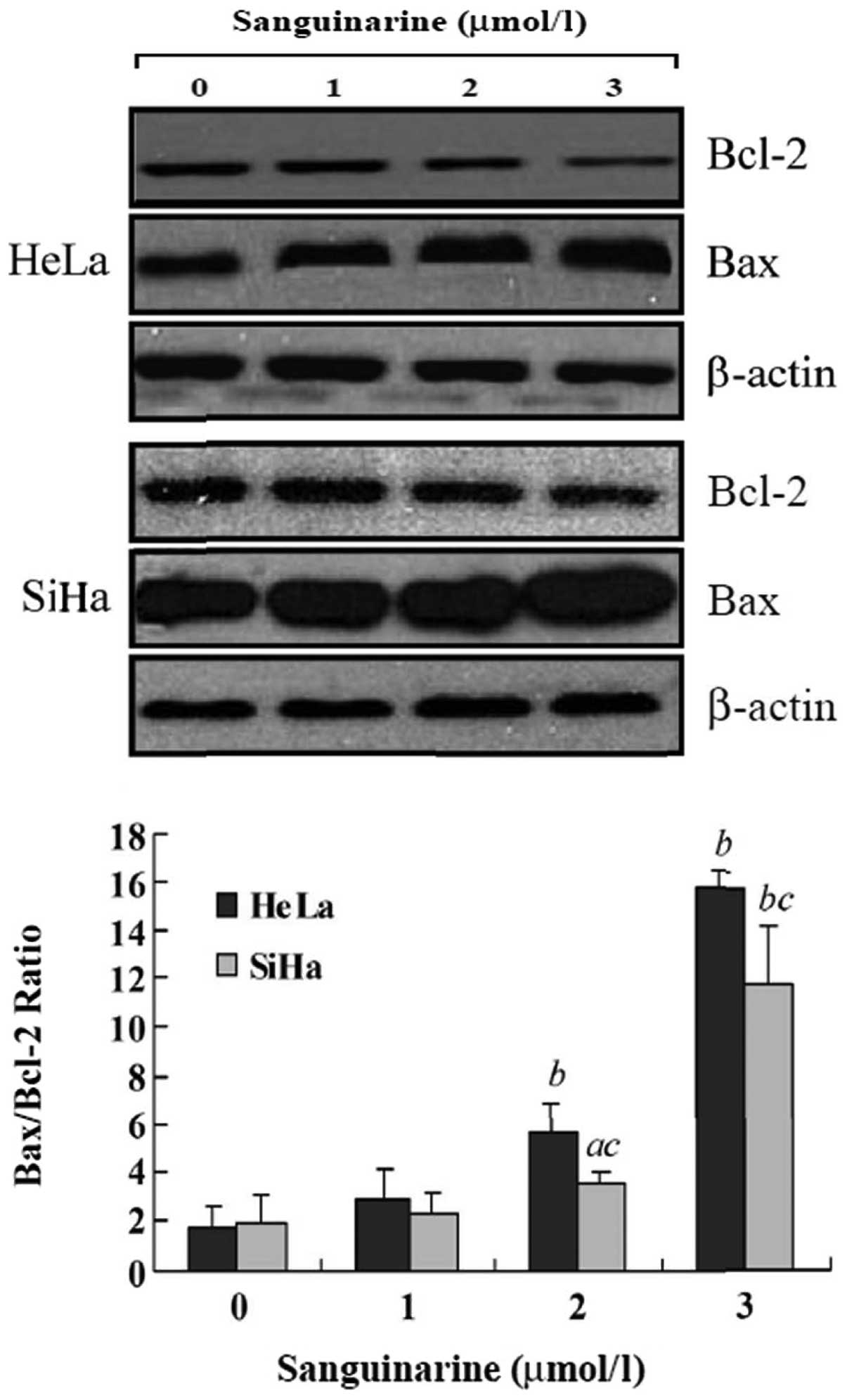
Effects of sanguinarine on
apoptosis-related proteins
To investigate the potential mechanism(s) involved
in sanguinarine-induced apoptosis, we determined the impact of
sanguinarine on Bcl-2 and Bax, two key apoptosis regulator
proteins. As shown in Fig. 7, 24 h
of exposure to sanguinarine resulted in a dose-dependent increase
of pro-apoptotic protein Bax expression and a concomitant decrease
of anti-apoptotic protein Bcl-2 levels. After treatment with 2.0
and 3.0 μmol/l sanguinarine, the Bax/Bcl-2 ratio, which favors
apoptosis, significantly increased in HeLa cells (P<0.01) and in
SiHa cells (P<0.05 for 2.0 μmol/l sanguinarine; P<0.01 for
3.0 μmol/l sanguinarine).
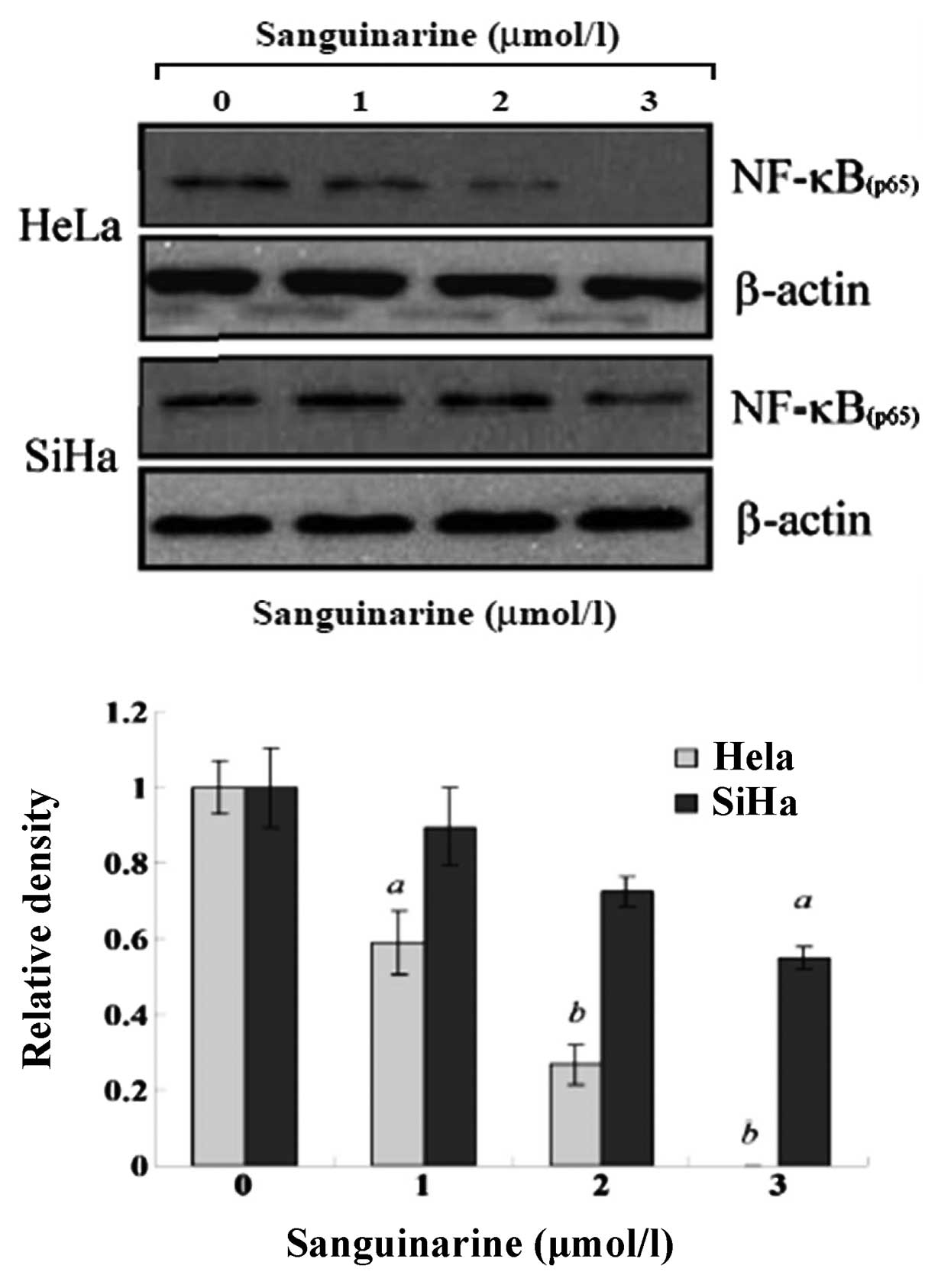
To investigate the alteration of NF-κB in
sanguinarine-induced apoptosis, the p65 subunit of NF-κB was
determined by western blot assay. As shown in Fig. 8, a dose-dependent decrease of NF-κB
protein expression was detectable in HeLa and SiHa cells. Moreover,
such reduction of NF-κB was more evident in HeLa cells compared to
SiHa cells.
Discussion
In the present study, we demonstrated that
sanguinarine significantly inhibited cell viability and colony
formation in HeLa and SiHa human cervical cancer cells in a
dose-dependent manner. The IC50 of sanguinarine was
2.62±0.21 μmol/l in HeLa cells and 3.07±0.23 μmol/l in SiHa cells,
respectively, which is consistent with the results of previous
studies(14). Other studies
reported that IC50 of sanguinarine is 0.72±0.33 μmol/l
in HL-60 human leukemia cells (12)
and 5.2 μmol/l in L1210, a mouse lymphocytic leukemic cell
line(15). Thus, the anticancer
ability of sanguinarine may be cell-type dependent. The clonal
adaptation and growth of cancer cells are crucial for tumor growth
(23), hence, suppression of cell
colony formation in the present study may suggest potential
anticancer use of sanguinarine at different stages of
carcinogenesis.
It has been reported that cervical adenocarcinoma is
chemo- and radio-resistant and patients with adenocarcinoma of
uterine cervix exhibit significantly poorer recurrence-free
survival than those with squamous cell carcinoma (24,25).The inhibitory effect of sanguinarine
on chemo-resistant cells was also observed using multidrug
resistance (MDR) cells to human papillomavirus type 16-immortalized
endocervical cells (18). Similar
sanguinarine sensitivity of HeLa (cervical adenocarcinoma) cells to
SiHa (squamous cell carcinoma) cells may suggest that sanguinarine
is a promising agent for possible development as an anticancer drug
for cervical cancer, particularly for radio-chemo-resistant
cervical adenocarcinoma.
Both normal and cancer cells within a living system
are affected by the rate of apoptosis, which is regarded as an
ideal way to eliminate damaged cells. Agents that induce apoptosis
may be used for the management and therapy of cancer by modulating
the steady-state cell population. To determine whether the decrease
of viability and colony formation in cervical cells exposed to
sanguinarine was due to apoptosis, we performed morphological
observation by light microscopy and found a significant change of
apoptotic morphology following treatment with sanguinarine. The
induction of apoptosis by sanguinarine was also evident from the
assay of DNA fragmentation using classical DNA ladder assay since
fragmentation of DNA by endonucleases is considered a hallmark of
apoptotic cell death (26).
Sanguinarine provoked the DNA fragmentation in cervical cancer
cells, which is in agreement with findings of previous studies in
human epidermoid carcinoma (A431), human breast cancer cells
(MDA-231), KB cancer cells and primary effusion lymphoma cells
(4,10,27,28).
However, Slunská et al found that compared with other
benzophenanth ridine alkaloids, sanguinarine did not induce
significant DNA ladder formation in HL-60 cells (19). Weerasinghe et al also found
that human erythroleukemia cells treated with a low level (4.1
μmol/l) of sanguinarine showed the morphology of apoptosis in ~96%
of cells, and sanguinarine at a high dose of 34.4 μmol/l resulted
in necrotic cell death morphology in over 90% of cells (5,6).
The appearance of sub-G1 peak on a DNA histogram, as
determined by flow cytometry, represents apoptotic cells with
fractional DNA content which underwent activation-induced apoptosis
(29). Our current results provide
additional evidence that sanguinarine results in dose-dependent
apoptosis induction in cervical cancer cells as the percentage of
sub-G1 gradually increased after exposure to sanguinarine, which is
consistent with other studies (4,12,19,27,30.31).
Members of the Bcl-2 protein family regulate and
execute many cell intrinsic apoptotic pathways; for example, Bcl-2
has been shown to function as an inhibitor of apoptosis. The
increased expression of Bcl-2 confers drug resistance, which is a
major obstacle to successful chemotherapy (32). By contrast, Bax, another member of
the Bcl-2 family, is found to be involved in the apoptotic cell
death process (33).Bax has been
shown to significantly increase the sensitivity to chemotherapeutic
drugs such as gemcitabine and 5-Fu in pancreatic cancer AsPC-1
cells (34). Our results also show
an upregulation of the Bax protein and a downregulation of the
Bcl-2 protein in response to sanguinarine, further suggesting a
role of apoptosis induction in controlling growth of cervical
cancer cells by sanguinarine. On the other hand, the imbalance
between pro-apoptotic and anti-apoptotic proteins is involved in
the distinctive biological features of cancer cells. An increase of
Bax/Bcl2 ratio favors apoptosis induction and enhances
chemotherapeutic effects, which has been found inhuman pancreatic
carcinoma cells, K562 erythroleukemia cells, immortalized human
keratinytes (HaCaT) (5,7,20) and
human cervical cancer cells used in our study following treatment
with sanguinarine. Bax expression increases sensitivity of cancer
cells to chemotherapeutic drugs (34) and Bcl-2 increase is related to drug
resistance (32).
NF-κB is a nuclear transcription factor that
regulates expression of a large number of genes involved in the
regulation of apoptosis, tumorigenesis, inflammation,
atherosclerosis, viral replication and several autoimmune
diseases(35). NF-κB is commonly
upregulated in cancer cells and several attempts are currently
underway to block its activation in order to render cells more
susceptible to anticancer drugs (35,36).
Chaturvedi et al first reported that the tumor necrosis
factor (TNF)-induced appearance of p65 was blocked by sanguinarine
and the inhibition of NF-κB activation by sanguinarine is not
cell-type specific (21). In our
present study, sanguinarine reduced the expression of the NF-κB
protein. This inhibitory effect may be specific to sanguinarine and
may not be observed in other structural analogues of sanguinarine
(21,37). Our results are also supported by
other studies where the decrease of NF-κB expression in K562
erythroleukemia cells was observed following treatment with 4.5
μmol/l of sanguinarine (5).
In conclusion, sanguinarine in the range of observed
concentrations, significantly inhibits proliferation and induces
apoptosis in both HeLa and SiHa human cervical cancer cell lines.
The increase of apoptotic bodies, DNA fragmentation and sub-G1
phase by sanguinarine was more obvious in HeLa cells than in SiHa
cells. Furthermore, sanguinarine decreased the expression of
anti-apoptotic proteins Bcl-2 and transcription factor NF-κB and
increased the pro-apoptotic protein Bax in both cell lines,
particularly in HeLa cells. Therefore, the higher sensitivity to
sanguinarine in HeLa cells compared to SiHa cells is contributed to
the higher susceptibility of HeLa cells to sanguinarine-induced
apoptosis. These observations further support the possible use of
sanguinarine in the treatment of cervical cancer, although further
studies are required to explore the exact mechanism(s) of the
responses evoked by sanguinarine as well as to verify their
effectiveness in an animal model system.
Acknowledgements
This study was supported by grants from the NNSFC
(81071906 and 81172127), the Program for Scientific Innovation
Research of College Graduate in Jiangsu Province (CX10B-0452); the
Outstanding Doctoral Thesis Project of Soochow University
(233209749); the Medical Scientific Research Project of Soochow
University (HZ200905) and the Program for Preliminary Scientific
Research of Soochow University (SDY2011A45).
References
|
1
|
Newman DJ and Cragg GM: Natural products
as sources of new drugs over the last 25 years. J Nat Prod.
70:461–477. 2007.PubMed/NCBI
|
|
2
|
Laster LL and Lobene RR: New perspectives
on sanguinaria clinicals: individual toothpaste and oral rinse
testing. J Can Dent Assoc. 56:19–30. 1990.
|
|
3
|
Firatli E, Unal T, Onan U, et al:
Antioxidative activities of some chemotherapeutics. A possible
mechanism in reducing gingival inflammation. J Clin Periodontol.
21:680–683. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ahmad N, Gupta S, Husain MM, et al:
Differential antiproliferative and apoptotic response of
sanguinarine for cancer cells versus normal cells. Clin Cancer Res.
6:1524–1528. 2000.PubMed/NCBI
|
|
5
|
Weerasinghe P, Hallock S and Liepins A:
Bax, Bcl-2, and NF-kappaB expression in sanguinarine induced
bimodal cell death. Exp Mol Pathol. 71:89–98. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Weerasinghe P, Hallock S, Tang SC, et al:
Role of Bcl-2 family proteins and caspase-3 in sanguinarine-induced
bimodal cell death. Cell Biol Toxicol. 17:371–381. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Adhami VM, Aziz MH, Reagan-Shaw SR, et al:
Sanguinarine causes cell cycle blockade and apoptosis of human
prostate carcinoma cells via modulation of cyclin kinase
inhibitor-cyclin-cyclin-dependent kinase machinery. Mol Cancer
Ther. 3:933–940. 2004.
|
|
8
|
Ahsan H, Reagan-Shaw S, Breur J, et al:
Sanguinarine induces apoptosis of human pancreatic carcinoma AsPC-1
and BxPC-3cells via modulations in Bcl-2 family proteins. Cancer
Lett. 249:198–208. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Matkar SS, Wrischnik LA and
Hellmann-Blumberg U: Sanguinarine causes DNA damage and
p53-independent cell death in human colon cancer cell lines. Chem
Biol Interact. 172:63–71. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim S, Lee TJ, Leem J, et al:
Sanguinarine-induced apoptosis: generation of ROS, down-regulation
of Bcl-2, c-FLIP, and synergy with TRAIL. J Cell Biochem.
104:895–907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jang BC, Park JG, Song DK, et al:
Sanguinarine induces apoptosis in A549 human lung cancer cells
primarily via cellular glutathione depletion. Toxicol In vitro.
23:281–287. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
vrba J, Dolezel P, vicar J, et al:
Cytotoxic activity of sanguinarine and dihydrosanguinarine in human
promyelocytic leukemia HL-60 cells. Toxicol In vitro. 23:580–588.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Park H, Bergeron E, Senta H, et al:
Sanguinarine induces apoptosis of human osteosarcoma cells through
the extrinsic and intrinsic pathways. Biochem Biophys Res Commun.
399:446–451. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Slaninová I, Táborská E, Bochoráková H, et
al: Interaction of benzo[c]phenanthridine and protoberberine
alkaloids with animal and yeast cells. Cell Biol Toxicol. 17:51–63.
2001.
|
|
15
|
Kaminskyy V, Lin KW, Filyak Y, et al:
Differential effect of sanguinarine, chelerythrine and chelidonine
on DNA damage and cell viability in primary mouse spleen cells and
mouse leukemic cells. Cell Biol Int. 32:271–277. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Scarinci IC, Garcia FA, Kobetz E, et al:
Cervical cancer prevention: new tools and old barriers. Cancer.
116:2531–2542. 2010.PubMed/NCBI
|
|
17
|
Long HJ: Management of metastatic cervical
cancer: review of the literature. J Clin Oncol. 25:2966–2974. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ding Z, Tang SC, Weerasinghe P, et al: The
alkaloid sanguinarine is effective against multidrug resistance in
human cervical cells via bimodal cell death. Biochem Pharmacol.
63:1415–1421. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Slunská Z, Gelnarová E, Hammerová J, et
al: Effect of quaternary benzo[c]phenanthridine alkaloids
sanguilutine and chelilutine on normal and cancer cells. Toxicol In
vitro. 24:697–706. 2010.
|
|
20
|
Adhami VM, Aziz MH, Mukhtar H, et al:
Activation of prodeath Bcl-2 family proteins and mithondrial
apoptosis pathway by sanguinarine in immortalized human HaCaT
keratinytes. ClinCancer Res. 9:3176–3182. 2003.PubMed/NCBI
|
|
21
|
Chaturvedi MM, Kumar A, Darnay BG, et al:
Sanguinarine(pseudhelerythrine) is a potent inhibitor of NF-kappaB
activation, IkappaBalpha phosphorylation, and degradation. J Biol
Chem. 272:30129–30134. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu S, Semenciw R, Probert A, et al:
Cervical cancer in Canada: changing patterns in incidence and
mortality. Int J Gynecol Cancer. 11:24–31. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Laconi E, Pani P and Farber E: The
resistance phenotype in the development and treatment of cancer.
Lancet Oncol. 1:235–241. 2000. View Article : Google Scholar
|
|
24
|
Park JY, Kim DY, Kim JH, et al: Outcomes
after radical hysterectomy in patients with early-stage
adenocarcinoma of uterine cervix. Br J Cancer. 102:1692–1698. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Quinn MA: Adenocarcinoma of the cervix -
are there arguments for a different treatment policy? Curr Opin
Obstet Gynecol. 9:21–24. 1997.PubMed/NCBI
|
|
26
|
Collins JA, Schandi CA, Young KK, et al:
Major DNA fragmentation is a late event in apoptosis. J Histochem
Cytochem. 45:923–934. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chang MC, Chan CP, Wang YJ, et al:
Induction of necrosis and apoptosis to KB cancer cells by
sanguinarine is associated with reactive oxygen species production
and mitochondrial membrane depolarization. Toxicol Appl Pharmacol.
218:143–151. 2007. View Article : Google Scholar
|
|
28
|
Hussain AR, Al-Jomah NA, Siraj AK, et al:
Sanguinarine-dependent induction of apoptosis in primary effusion
lymphoma cells. Cancer Res. 67:3888–3897. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kajstura M, Halicka HD, Pryjma J and
Darzynkiewicz Z: Discontinuous fragmentation of nuclear DNA during
apoptosis revealed by discrete ‘sub-G1’ peaks on DNA content
histograms. Cytometry A. 71:125–131. 2007.PubMed/NCBI
|
|
30
|
McDonald ER III and El-Deiry WS: Cell
cycle control as a basis for cancer drug development (Review). Int
J Oncol. 16:871–886. 2000.PubMed/NCBI
|
|
31
|
Malikova J, Zdarilova A and Hlobilkova A:
Effects of sanguinarine and chelerythrine on the cell cycle and
apoptosis. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub.
150:5–12. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Reed JC: Bcl-2: prevention of apoptosis as
a mechanism of drug resistance. Hematol Oncol Clin North Am.
9:451–473. 1995.PubMed/NCBI
|
|
33
|
Brady HJ and Gil-Gómez G: Bax. The
pro-apoptotic Bcl-2 family member, Bax. Int J Biochem Cell Biol.
30:647–650. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu ZW, Friess H, Büchler MW, et al:
Overexpression of Bax sensitizes human pancreatic cancer cells to
apoptosis induced by chemotherapeutic agents. Cancer Chemother
Pharmacol. 49:504–510. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Heafner B: NF-kappa B: arresting a major
culprit in cancer. Drug Discov Today. 7:653–663. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nakanishi C and Toi M: Nuclear
factor-kappaB inhibitors as sensitizers to anticancer drugs. Nat
Rev Cancer. 5:153–159. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mendoza J, Zamora R, Gallardo JC, et al:
NF-kappaB does not influence the induction of apoptosis by Ukrain.
Cancer Biol Ther. 5:788–793. 2006. View Article : Google Scholar : PubMed/NCBI
|