Introduction
Pancreatic cancer is the fourth leading cause of
cancer death in both men and women in the United States. It has
amongst the worst prognoses of all human malignancies. A majority
of patients are symptomatic at the time of diagnosis and have
developed advanced metastatic disease (1). Most patients die within a year of
diagnosis and the 5-year survival rate is <5% (2). Despite many multi-modal treatment
strategies, the survival rates have not improved significantly and
mortality remains high (3).
Gemcitabine (GEM) has remained the mainstay primary
chemotherapeutic agent for systemic treatment in advanced
pancreatic adenocarcinoma. Multiple studies have shown survival
benefits of GEM chemotherapy, both alone as well as in combination
with other drugs (4–6). However, despite significant advances
and improvement in therapy, the 5-year survival remains quite low.
As a result, there is now a shift towards target specific agents,
in addition to conventional cytotoxic drugs, with an aim towards
increasing overall survival (7–11).
ErbB1 and ErbB2 are members of the epidermal growth
factor (EGF) receptor family and are known to play an essential
role in regulation of cell proliferation and differentiation. The
formation of homodimers and heterodimers between different EGF
receptors is believed to regulate complex signal transduction
pathways via activation of intrinsic protein tyrosine kinase
activity. This leads to recruitment and phosphorylation of several
intracellular substrates leading to various cellular activities
including mitogenic signaling and cell growth (12,13).
Some studies have also suggested that overexpression
of ErbB1 and ErbB2 in certain cancers is associated with increased
tumor aggression and poor prognosis (12,14,15).
Targeted downregulation of these receptors has been shown to cause
apoptosis and cell death in cancers that overexpress these
receptors (12). Since a
significant percentage of pancreatic tumors overexpress both ErbB1
and ErbB2 (14–16), targeting these receptors may be a
viable strategy in patients diagnosed with such tumors.
Many studies have looked at the response of
pancreatic neoplasms to ErbB1 or ErbB2 targeted therapy.
Transtuzumab (ErbB2) (11),
erlotinib (ErbB1) (10), cetuximab
(ErbB1) (8) and gefitinib (ErbB1)
(9) have been studied in the last
few years; however, none of these drugs has shown any significant
improvement in mortality when compared to treatment with
conventional cytotoxic drugs. It was observed that most patients
eventually developed drug resistance. This negated any benefits
that these agents potentially offered. Since, all these studies
targeted either ErbB1 or ErbB2 separately, we hypothesized that
simultaneous inhibition of both ErbB1 and ErbB2 receptors might
suppress tumor growth better than targeting them individually, and
may help overcome the development of drug resistance. We selected
lapatinib (Tykerb), a dual ErbB1 and ErbB2 tyrosine kinase enzyme
inhibitor, for our study since lapatinib has been shown in multiple
studies to benefit patients who have developed drug resistance to
prior targeted therapies (17–19).
Materials and methods
Pancreatic cell lines and culture
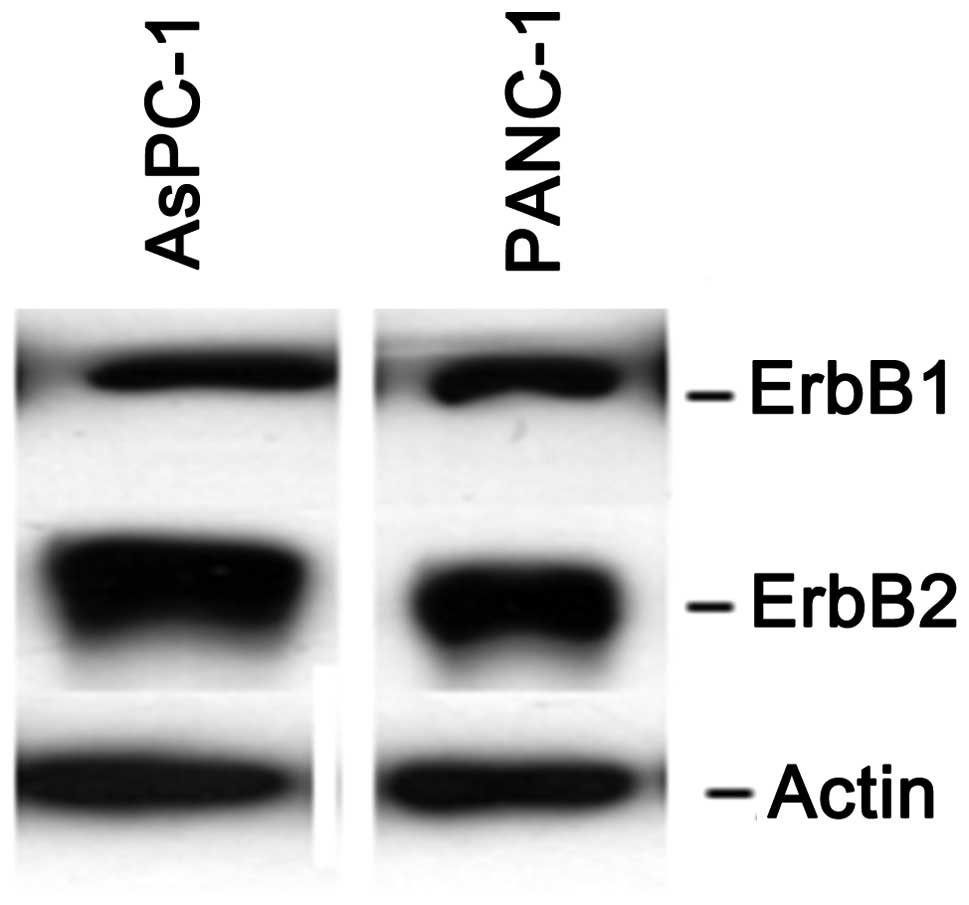
Human pancreatic cell lines AsPC-1 and PANC-1 were
selected for our experiments due to their expression of ErbB1 and
ErbB2 receptors (Fig. 1). All cell
lines were purchased from ATCC (Manassas, VA) and maintained in
recommended media supplemented with 10% fetal bovine serum
(Mediatech, Manassas, VA). All cells were cultured at 37°C in 5%
CO2 with 100% humidity.
Chemicals
Lapatinib (Tykerb, Genentech Inc, South San
Francisco, CA) was kindly provided by Genentech Inc. (South San
Francisco, CA) and gemcitabine and 5-fluorouracil (5-FU) were
obtained from Sigma/Aldrich, St. Louis, MO.
In vitro proliferation assay (MTT
assay)
Cell growth was assessed using a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
(MTT assay). Briefly, 5,000 viable cells were seeded into
flat-bottomed 96-well plates in triplicate and allowed to adhere
overnight. Cells were treated with the addition of the intended
doses of lapatinib (0–16 μM), gemcitabine (0–400 nM) and/or 5-FU
(0–16 μM). After 3–4 days of incubation,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide was
added to a final concentration of 0.5 mg/ml and cells were
incubated an additional 4 h. Cells were lysed with the addition of
isopropanol. Absorbance was measured at 595 and 655 nm in a 96-well
plate reader (Bio-Rad Model 680, Bio-Rad Laboratories, Hercules,
CA) and growth curves calculated.
In vitro apoptosis assay
Apoptosis activity was assessed using the ApoAlert
Caspase-3 Colorimetric Assay kit (Clontech, Mountain View, CA).
Briefly, 106 viable cells were seeded in 100-mm tissue
culture plates and allowed to adhere overnight. Cells were treated
with the addition of the intended doses of lapatinib (0–16 μM),
gemcitabine (0–400 nM) and/or 5-FU (0–16 μM). Cells were harvested
from plates using cell scrapers 48 h after the addition of drugs
and lysed for analysis of caspase-3 activity according to the
manufacturer’s instructions. Readings were taken at 405 nM in
disposable cuvettes in a spectrophotometer (Thermogenesis 6,
Hopkinton, MA) and caspase-3 activity was calculated.
In vitro immunoblot analyses (western
blot analysis)
Cells were plated and treated in the same manner as
described for the apoptosis assay. After incubating for 48 h, cells
were harvested using cell scrapers and washed with DPBS. Cells were
lysed on ice in HEPES lysis buffer containing 50 mM HEPES, 100 mM
NaCl, 1% Triton X-100, 10% Glycerol, 1 mM
Na3VO4, 5 mM NaFl, 1:50 dilution of Complete
Protease Inhibitor Cocktail tablets (Roche, Indianapolis, IN).
Protein concentrations were determined via Coomassie Protein Assay
Reagent (Thermo Scientific, Rockford, IL) and samples were
standardized to 10 μg in SDS-PAGE loading buffer. Samples were
loaded on a 7.5% SDS-PAGE acrylamide gel and run under reducing
conditions for 2 h at 100 V. The proteins were transferred to
Immobilon-P membranes (Millipore Co., Billerica, MA) using Towbins
transfer buffer (25 mM Tris, 192 mM glycine, 10% methanol).
Membranes were blocked in TBS (20 mM Tris, 150 mM NaCl) containing
0.1% Tween-20 and 5% non-fat dry milk or BSA and incubated with
antibodies against target proteins. Antibodies include ErbB1
(Chemicon, Temecula, CA), ErbB2 (AbCam, Cambridge, MA), Phos-ERBB1
(AbCam), Phos-ERBB2 (Cell Signaling, Beverly, MA), α-tubulin (Santa
Cruz Biotechnology, Santa Cruz, CA).
Results
The effects of lapatinib on cell
proliferation
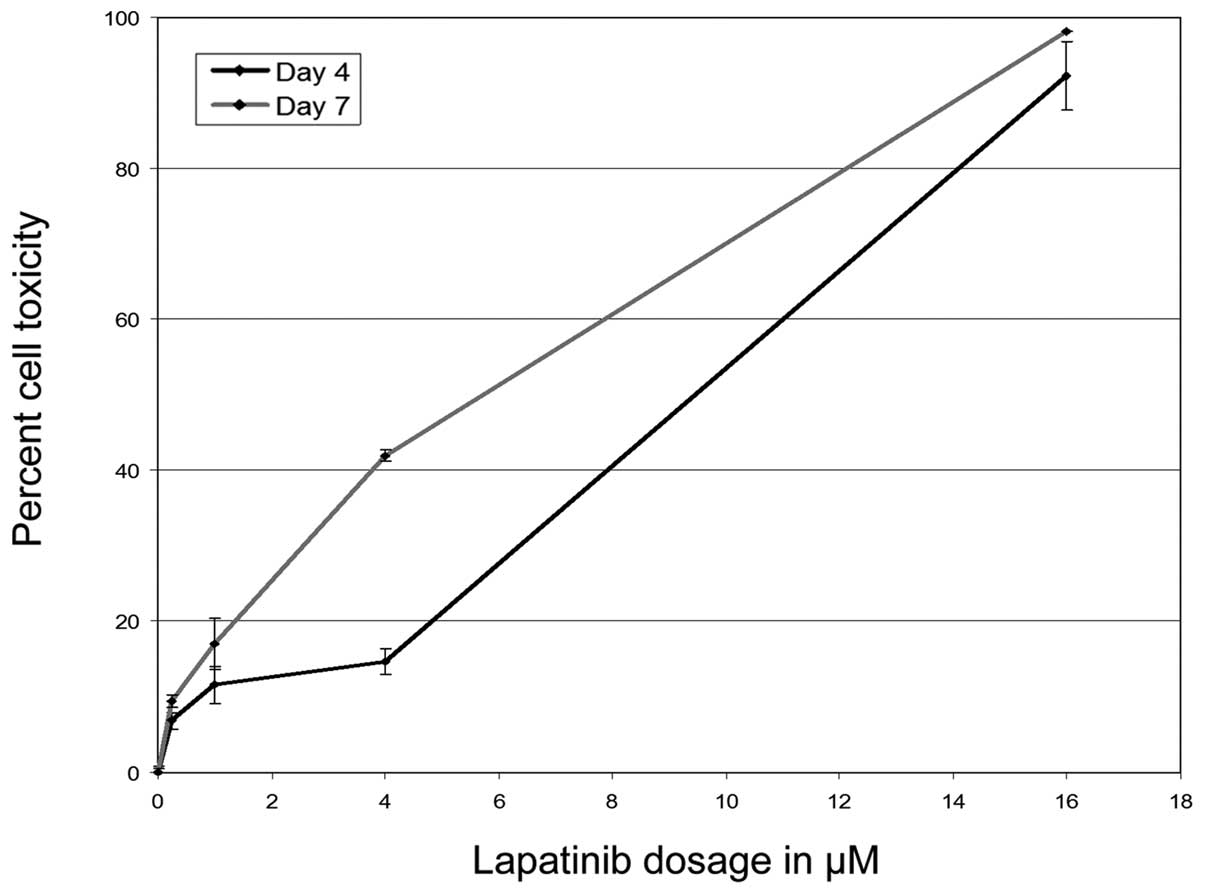
Initial experiments studied the effects of
lapatinib, both alone and in combination with 5-FU or gemcitabine,
on proliferation of AsPC and PANC-1 cells in vitro.
Lapatinib inhibited cell proliferation in a dose-dependent manner.
At lower doses (0.25–1 μM) lapatinib had no effect, but at higher
doses (4–16 μM), inhibition was significant. There was a 50%
inhibition in cell proliferation at a dose of 12 μM, but the
effects plateaued at higher doses (Fig.
2).
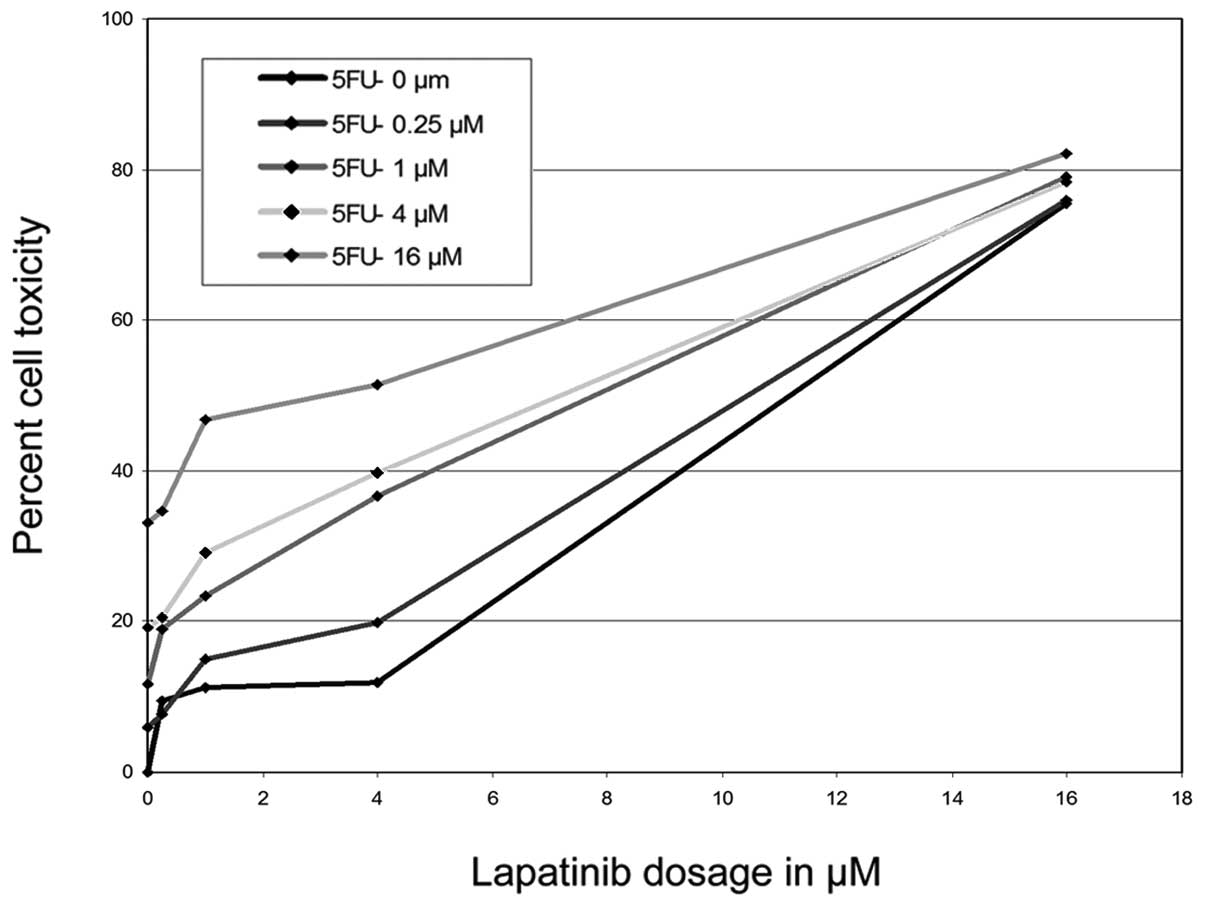
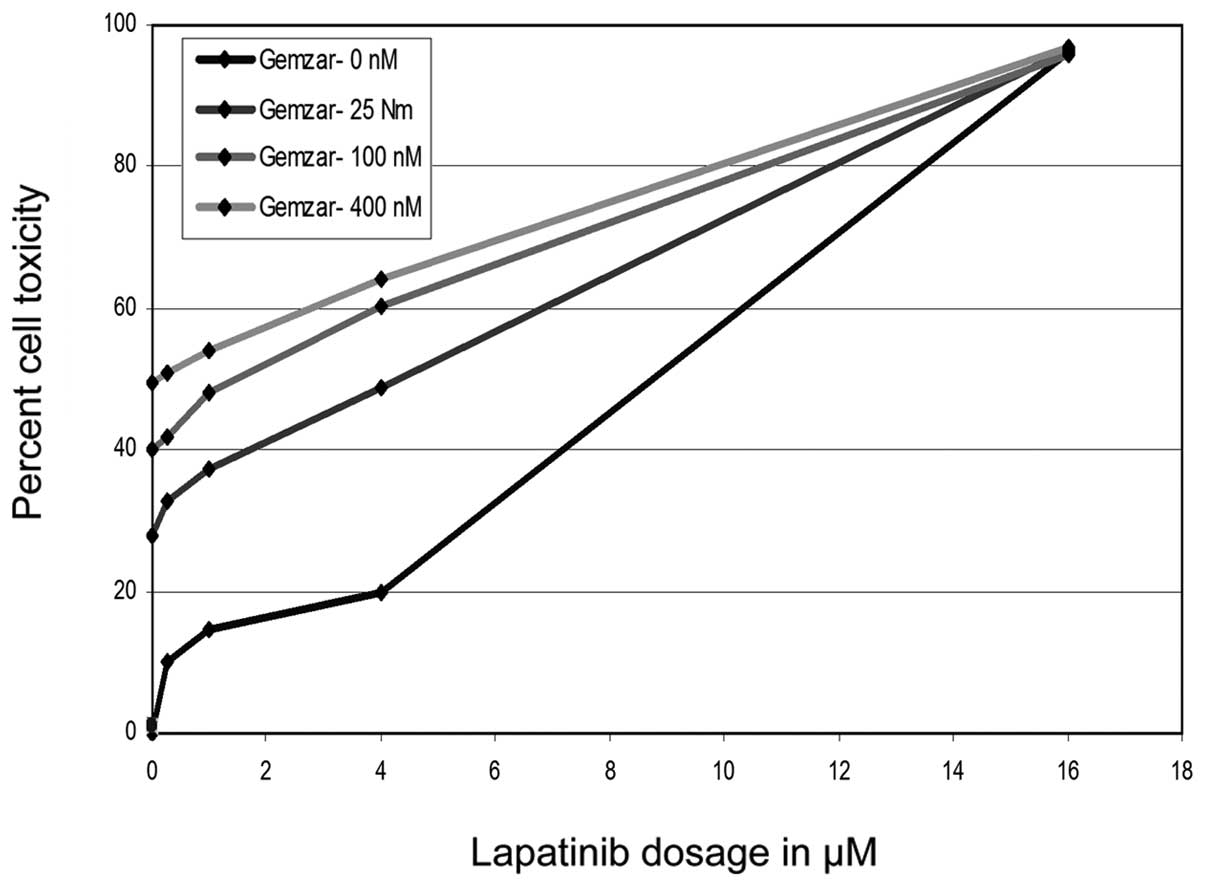
The inhibitory effects of lapatinib were potentiated
in the presence of 5-FU or GEM, with a dose-dependent shift in the
proliferation curve (Figs. 3 and
4). This suggests that lapatinib
may work synergistically with these other drugs, increasing their
effect in vitro. The combination of drugs allowed similar
inhibition at much lower doses. This may allow for reducing the
dosage of each drug in vivo and thus the incidence of
adverse side effects often noticed in current recommended dosages
(20,21).
The effect of lapatinib on ErbB1 and
ErbB2 protein levels
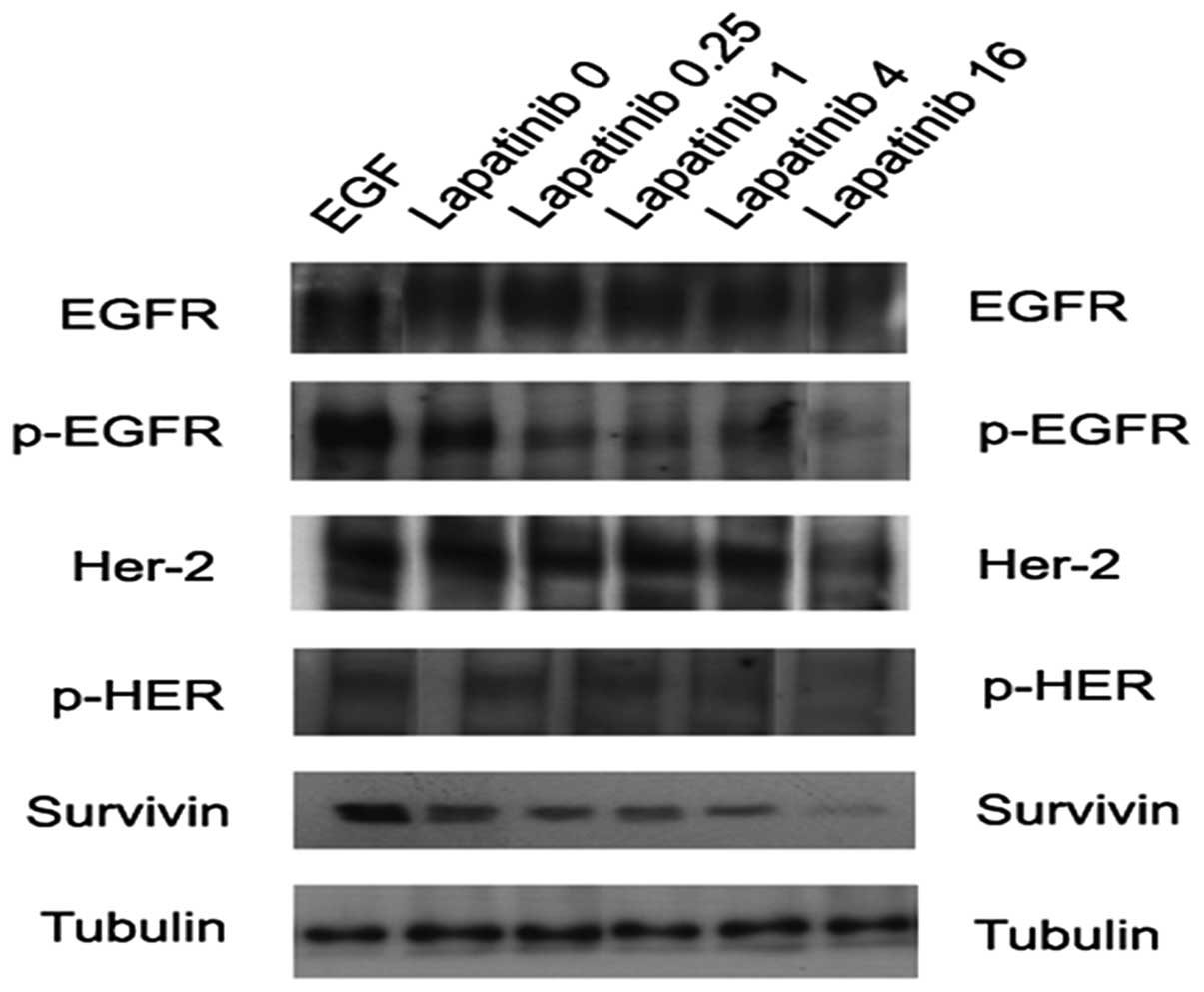
AsPC and PANC-1 cells were treated with lapatinib at
dosages similar to those used in the cell proliferation assays.
Following treatment with lapatinib, samples were analyzed for
expression of ErbB1, ErbB2 and their respective phosphorylated
proteins using immunoblotting.
While the levels of total ErbB1 and ErbB2 proteins
remained unchanged, both phosphorylated ErbB1 and ErbB2
(pErbB1-Tyr1173 and ErbB2-Tyr1248) protein levels decreased with
lapatinib treatment. Again, lower doses showed no effects; however,
at higher doses (12–16 μM) decrease in phosphorylated protein was
detectable. There was an estimated 50% reduction in phosphorylated
protein at 12 μM, which corresponds to the dose needed to inhibit
proliferation by the same amount (Fig.
5). These sites (Tyr1173 and Tyr1248) have been found to be
involved in regulation of tyrosine kinase activity and
phosphorylation of these sites couples ErbB1 and ErbB2 to
downstream kinase signal transduction pathways (22–24).
Therefore, lapatinib downregulation of phosphorylated ErbB1 and
ErbB2 may decrease tyrosine kinase activity and subsequently
decrease proliferation through these downstream pathways.
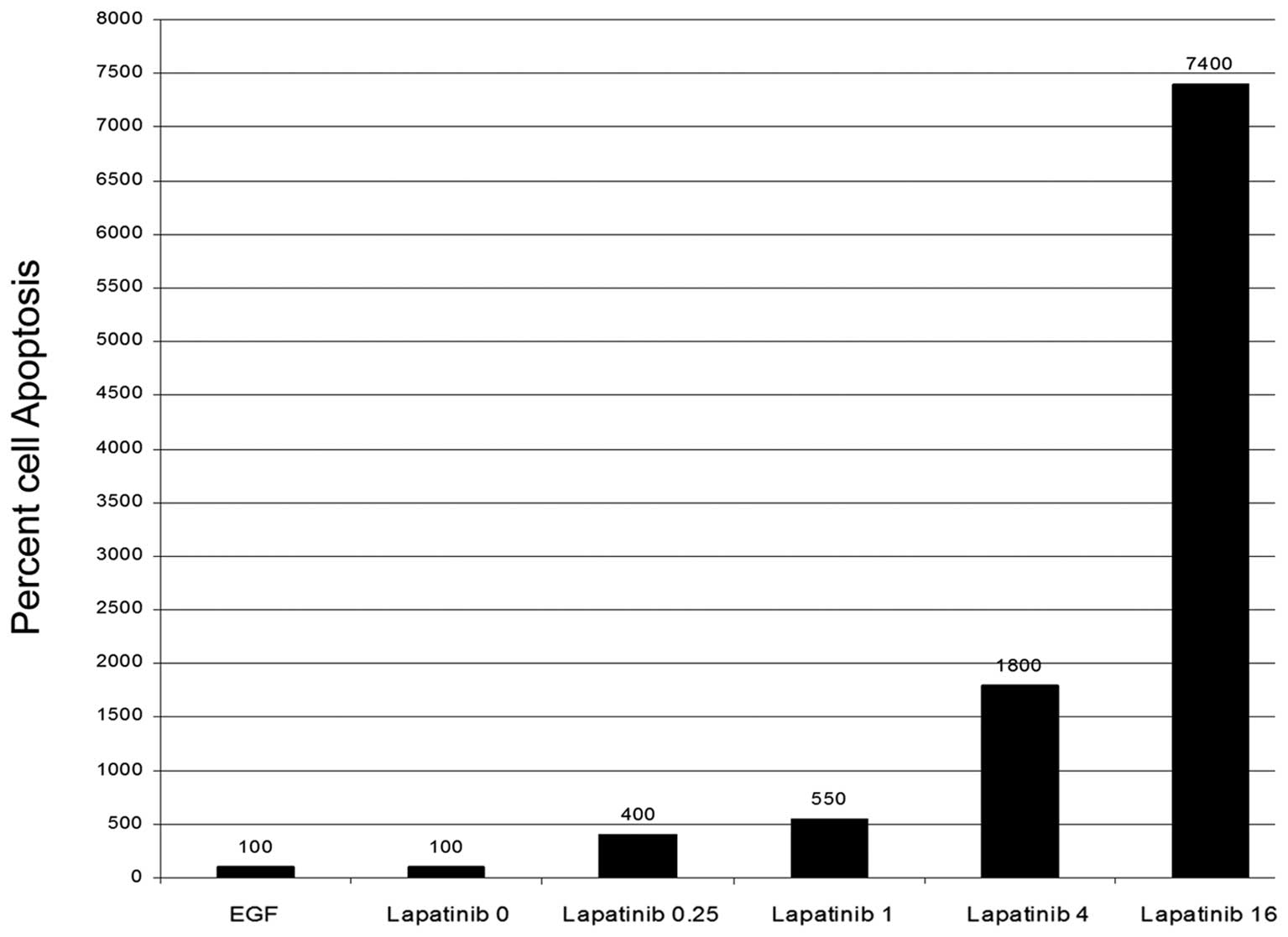
The effect of lapatinib on cell
apoptosis
AsPC and PANC-1 cells were treated in the same
manner as the proliferation assays. Forty-eight hours after
treatment, apoptosis activity was evaluated using a caspase-3 and
-8 colorimetric assay kit. Both cell lines exhibited an increase in
cleaved caspase-3 activity indicating apoptosis. This activity
corresponded with doses showing decreased proliferation with the
maximal activity in the range of 4–16 μM (Fig. 6). This would suggest that lapatinib
is causing apoptosis in addition to decreasing proliferation.
The effect of lapatinib on survivin
protein level
Based on the apoptosis results, we decided to test
survivin levels under our treatment conditions. Survivin has been
shown to inhibit apoptosis by binding to caspase-3 (25) and has also been shown to play a role
in pancreatic cancer (26–28). In order to determine if lapatinib
affects the survivin pathway, the expression of survivin protein
was assessed in both AsPC and PANC-1 cells using immunoblotting.
Again, there was a dose-dependent decrease in expression of
survivin protein levels with lapatinib treatment in both cell
lines. AsPC cells showed a complete reduction of expression at 12
μM, while PANC-1 cells showed only a 50% reduction at this same
dose (Fig. 5). Assuming survivin is
inhibiting apoptosis in these cells; lapatinib could play a key
role restoring apoptosis in pancreatic cancer cells.
Discussion
Pancreatic tumors have so far shown substantial
resistance to multiple chemotherapeutic drugs and regimens. It has
become a major challenge for clinicians to offer substantive
treatment options to patients with advanced pancreatic carcinomas.
However, with recent advancements in treatment strategies directed
against molecular targets, there has been a renewed interest. It
has been proposed that novel molecular agents targeting ErbB1 and
ErbB2 in pancreatic tumors will help overcome substantial tumor
resistance against conventional cytotoxic drugs and also offer
patient specific therapy depending upon the receptor and molecular
subtype.
As discussed earlier, some pancreatic tumors have
high levels of ErbB1 and ErbB2 cell surface receptors (14–16).
These tumors, therefore, are ideal for targeted therapeutic
strategy since both ErbB1 and ErbB2 receptors along with their
downstream proteins have been shown to promote cell growth and
survival, and mediate resistance to chemotherapy. Many studies have
been published describing the effects of newer molecular agents
directed against ErbB1 and ErbB2 receptors in pancreatic neoplasms
(8–11), however, despite theoretical
advantages; these agents have failed to yield significantly
beneficial results, either alone or in combination with
conventional cytotoxic drugs. Although, the exact mechanisms
responsible for failure to respond are a matter of intense
scrutiny, it seems that a significant number of patients develop
drug resistance. Numerous studies have been published describing
the potential mechanisms of resistance against these agents.
Engelman et al (29) showed
that amplification of MET caused gefitinib resistance by driving
HER3-dependent activation of PI3K while Scaltriti et al
(17) found that expression of
truncated HER2 altered the ErbB2 receptor such that it impaired the
access of trastuzumab to the attachment site. Furthermore, Nagy
et al (30) described the
development of resistance in some patients due to overexpression of
MUC4.
While it remains to be seen what exactly causes
development of resistance against these agents, there is ample
evidence to suggest that ErbB subunits work in tandem as a unit and
therefore, it may be possible that broad inhibition of these
subunits may be necessary to overcome the development of
resistance. Working on this hypothesis, we selected lapatinib,
based on its mechanism of dual inhibition of ErbB1 and ErbB2
tyrosine kinase and its favorable response in patients who have
developed resistance against single-receptor based molecular agents
(17,31).
We found that lapatinib markedly reduced tyrosine
phosphorylation of ErbB1 and ErbB2, which inhibits activation of
Erk1/2 and AKT, the downstream effectors of cell proliferation and
survival, respectively (32). We
also found that lapatinib caused a decrease in caspase-3 activity
and the expression of survivin, a protein inhibitor of the
apoptosis. This is significant because the exact mechanism by which
lapatinib exerts its apoptotic effect is unknown and therefore, it
may be possible that the inhibition of survivin could be one of
mechanisms by which lapatnib exerts its effects. Many studies have
shown that survivin protects tumors from programmed cell death
(33–35), however, none of the prior studies
with either ErbB1 or ErbB2 inhibitors could modulate survivin.
Investigators have added lapatinib to
chemotherapeutic agents such as paclitaxel (36), docetaxel (37), capecitabine (38), letrozole (39), and to combination regimens such as
FOLFOX (40) and FOLFOX (41), with promising results. Although
clinical trials in different tumor types are ongoing, the most
mature data thus far have been in the treatment of breast cancer,
particularly in trastuzumab-resistant patients (37). We also studied the effects of
lapatinib in combination with 5-FU and gemcitabine. In combination,
lapatinib revealed enhanced toxicity with pronounced cell death and
decreased cell survival. These effects were consistent in both cell
lines suggesting that its effects may be broad based in cells
expressing these target proteins.
In conclusion, simultaneous dual ErbB1 and ErbB2
receptor tyrosine kinase inhibition with lapatinib inhibits
pancreatic cancer cell growth and proliferation and induces
apoptotic cell death. These effects occur at clinically achievable
concentrations and are synergistic with the effects of conventional
chemotherapy agents including 5-FU or gemcitabine. These findings
support the potential role of lapatinib in the treatment of
pancreatic cancer either alone or as an adjunct to conventional
chemotherapy agents.
References
|
1
|
Nelson NJ: Pancreatic cancer research
matures. J Natl Cancer Inst. 99:1432–1434. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar
|
|
3
|
Li J and Saif MW: Any progress in the
management of advanced pancreatic cancer? JOP. 10:361–365.
2009.
|
|
4
|
Zhu CP, Shi J, Chen YX, Xie WF and Lin Y:
Gemcitabine in the chemoradiotherapy for locally advanced
pancreatic cancer: a meta-analysis. Radiother Oncol. 99:108–113.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Heinemann V, Boeck S, Hinke A, Labianca R
and Louvet C: Meta-analysis of randomized trials: evaluation of
benefit from gemcitabine-based combination chemotherapy applied in
advanced pancreatic cancer. BMC Cancer. 8:822008. View Article : Google Scholar
|
|
6
|
Yip D, Karapetis C, Strickland A, Steer CB
and Goldstein D: Chemotherapy and radiotherapy for inoperable
advanced pancreatic cancer. Cochrane Database Syst Rev.
3:CD0020932006.
|
|
7
|
Rivera F, Lopez-Tarruella S, Vega-Villegas
ME and Salcedo M: Treatment of advanced pancreatic cancer: from
gemcitabine single agent to combinations and targeted therapy.
Cancer Treat Rev. 35:335–339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Philip PA, Benedetti J, Corless CL, et al:
Phase III study comparing gemcitabine plus cetuximab versus
gemcitabine in patients with advanced pancreatic adenocarcinoma:
Southwest Oncology Group-directed intergroup trial S0205. J Clin
Oncol. 28:3605–3610. 2010. View Article : Google Scholar
|
|
9
|
Fountzilas G, Bobos M, Kalogera-Fountzila
A, et al: Gemcitabine combined with gefitinib in patients with
inoperable or metastatic pancreatic cancer: a phase II Study of the
Hellenic Cooperative Oncology Group with biomarker evaluation.
Cancer Invest. 26:784–793. 2008. View Article : Google Scholar
|
|
10
|
Moore MJ, Goldstein D, Hamm J, et al:
National Cancer Institute of Canada Clinical Trials Group.
Erlotinib plus gemcitabine compared with gemcitabine alone in
patients with advanced pancreatic cancer: a phase III trial of the
National Cancer Institute of Canada Clinical Trials Group. J Clin
Oncol. 25:1960–1966. 2007. View Article : Google Scholar
|
|
11
|
Safran H, Iannitti D, Ramanathan R, et al:
Herceptin and gemcitabine for metastatic pancreatic cancers that
overexpress ERBB2/neu. Cancer Invest. 22:706–712. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mendelsohn J and Baselga J: The EGF
receptor family as targets for cancer therapy. Oncogene.
19:6550–6565. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Riese DJ II and Stern DF: Specificity
within the EGF family/ErbB receptor family signaling network.
Bioessays. 20:41–48. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ueda S, Ogata S, Tsuda H, et al: The
correlation between cytoplasmic overexpression of epidermal growth
factor receptor and tumor aggressiveness: poor prognosis in
patients with pancreatic ductal adenocarcinoma. Pancreas. 29:e1–e8.
2004. View Article : Google Scholar
|
|
15
|
Dancer J, Takei H, Ro JY and
Lowery-Nordberg M: Coexpression of ERBB1 and ERBB2 in pancreatic
ductal adenocarcinoma: a comparative study using
immunohistochemistry correlated with gene amplification by
fluorescencent in situ hybridization. Oncol Rep. 18:151–155.
2007.
|
|
16
|
Tobita K, Kijima H, Dowaki S, et al:
Epidermal growth factor receptor expression in human pancreatic
cancer: significance for liver metastasis. Int J Mol Med.
11:305–309. 2003.PubMed/NCBI
|
|
17
|
Scaltriti M, Rojo F, Ocana A, et al:
Expression of p95HER2, a truncated form of the HER2 receptor, and
response to anti-HER2 therapies in breast cancer. J Natl Cancer
Inst. 99:628–638. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Konecny GE, Pegram MD, Venkatesan N, et
al: Activity of the dual kinase inhibitor lapatinib (GW572016)
against HER-2-overexpressing and trastuzumab-treated breast cancer
cells. Cancer Res. 66:1630–1639. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
O’Brien NA, Browne BC, Chow L, et al:
Activated phosphoinositide 3-kinase/AKT signaling confers
resistance to trastuzumab but not lapatinib. Mol Cancer Ther.
9:1489–1502. 2010.PubMed/NCBI
|
|
20
|
Conroy T, Desseigne F, Ychou M, et al:
FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N
Engl J Med. 364:1817–1825. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mehta SP: Palliative chemotherapy for
pancreatic malignancies. Surg Clin North Am. 90:365–375. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Muthuswamy SK, Gilman M and Brugge JS:
Controlled dimerization of ErbB receptors provides evidence for
differential signaling by homo- and heterodimers. Mol Cell Biol.
19:6845–6857. 1999.PubMed/NCBI
|
|
23
|
Zwick E, Hackel PO, Prenzel N and Ullrich
A: The EGF receptor as central transducer of heterologous
signalling systems. Trends Pharmacol Sci. 20:408–412. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sorkin A, Helin K, Waters CM, Carpenter G
and Beguinot L: Multiple autophosphorylation sites of the epidermal
growth factor receptor are essential for receptor kinase activity
and internalization. Contrasting significance of tyrosine 992 in
the native and truncated receptors. J Biol Chem. 267:8672–8678.
1992.
|
|
25
|
Li F, Ambrosini G, Chu EY, Plescia J,
Tognin S, Marchisio PC and Altieri DC: Control of apoptosis and
mitotic spindle checkpoint by survivin. Nature. 396:580–584. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu BB and Wang WH: Survivin and
pancreatic cancer. World J Clin Oncol. 2:164–168. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Theodoropoulos GE, Michalopoulos NV,
Panoussopoulos SG, Taka S and Gazouli M: Effects of caspase-9 and
survivin gene polymorphisms in pancreatic cancer risk and tumor
characteristics. Pancreas. 39:976–980. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shen YM, Yang XC, Song ML, Qin CH, Yang C
and Sun YH: Growth inhibition induced by short hairpin RNA to
silence survivin gene in human pancreatic cancer cells.
Hepatobiliary Pancreat Dis Int. 9:69–77. 2010.PubMed/NCBI
|
|
29
|
Engelman JA, Zejnullahu K, Mitsudomi T, et
al: MET amplification leads to gefitinib resistance in lung cancer
by activating ERBB3 signaling. Science. 316:1039–1043. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nagy P, Friedlander E, Tanner M, Kapanen
AI, Carraway KL, Isola J and Jovin TM: Decreased accessibility and
lack of activation of ErbB2 in JIMT-1, a herceptin-resistant,
MUC4-expressing breast cancer cell line. Cancer Res. 65:473–482.
2005.PubMed/NCBI
|
|
31
|
Xia W, Liu LH, Ho P and Spector NL:
Truncated ErbB2 receptor (p95ErbB2) is regulated by heregulin
through heterodimer formation with ErbB3 yet remains sensitive to
the dual EGFR/ErbB2 kinase inhibitor GW572016. Oncogene.
23:646–653. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xia W, Mullin RJ, Keith BR, et al:
Anti-tumor activity of GW572016: a dual tyrosine kinase inhibitor
blocks EGF activation of EGFR/erbB2 and downstream Erk1/2 and AKT
pathways. Oncogene. 21:6255–6263. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Song Z, Yao X and Wu M: Direct interaction
between survivin and Smac/DIABLO is essential for the
anti-apoptotic activity of survivin during taxol-induced apoptosis.
J Biol Chem. 278:23130–23140. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Z, Fukuda S and Pelus LM: Survivin
regulates the p53 tumor suppressor gene family. Oncogene.
23:8146–8153. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Desplanques G, Hulin P, Juin P and
Barille-Nion S: Survivin Protects Breast Cancer Cells from DNA
Damage during Mitosis (abstract). In: Thirty-Second Annual
CTRC-AACR San Antonio Breast Cancer Symposium; Dec 10–13; San
Antonio, TX. pp. 2009Cancer Res. 69(Suppl 3)2009, View Article : Google Scholar
|
|
36
|
Boussen H, Cristofanilli M, Zaks T,
DeSilvio M, Salazar V and Spector N: Phase II study to evaluate the
efficacy and safety of neoadjuvant lapatinib plus paclitaxel in
patients with inflammatory breast cancer. J Clin Oncol.
28:3248–3255. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
LoRusso PM, Jones SF, Koch KM, et al:
Phase I and pharmacokinetic study of lapatinib and docetaxel in
patients with advanced cancer. J Clin Oncol. 26:3051–3056. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cameron D, Casey M, Press M, et al: A
phase III randomized comparison of lapatinib plus capecitabine
versus capecitabine alone in women with advanced breast cancer that
has progressed on trastuzumab: updated efficacy and biomarker
analyses. Breast Cancer Res Treat. 112:533–543. 2008. View Article : Google Scholar
|
|
39
|
Johnston S, Pippen J Jr, Pivot X, et al:
Lapatinib combined with letrozole versus letrozole and placebo as
first-line therapy for postmenopausal hormone receptor-positive
metastatic breast cancer. J Clin Oncol. 27:5538–5546. 2009.
View Article : Google Scholar
|
|
40
|
Siegel-Lakhai WS, Beijnen JH, Vervenne WL,
et al: Phase I pharmacokinetic study of the safety and tolerability
of lapatinib (GW572016) in combination with
oxaliplatin/fluorouracil/leucovorin (FOLFOX4) in patients with
solid tumors. Clin Cancer Res. 13:4495–4502. 2007. View Article : Google Scholar
|
|
41
|
Midgley RS, Kerr DJ, Flaherty KT, et al: A
phase I and pharmacokinetic study of lapatinib in combination with
infusional 5-fluorouracil, leucovorin and irinotecan. Ann Oncol.
18:2025–2029. 2007. View Article : Google Scholar : PubMed/NCBI
|