Introduction
Cytokines are a large group of soluble extracellular
proteins or glycoproteins, which are involved in many essential
cellular processes, including cell growth, death, differentiation,
angiogenesis and regulation of normal hematopoiesis (1). One of the types of glycoproteins,
well-known since 1960, is granulocyte-colony stimulating factor
(G-CSF), now also referred as CSF3 (2,3). G-CSF
is mainly produced by hematopoietic cells but also by bone marrow
stromal cells, endothelial cells, fibroblasts and macrophages
(4). G-CSF promotes several
cellular processes such as proliferation, maturation and
differentiation. In addition, CSF3 regulates the survival of mature
granulocytes and enhances chemotaxis, motility and phagocytosis of
mature neutrophils (2–6). This cytokine interacts with the
specific cell receptor G-CSFR, which belongs to the cytokine
receptor type I superfamily and is expressed on the surface of
early myeloid progenitors, mature granulocytes and
monocytes/macrophages (3). The
G-CSF gene was cloned in 1984 and allowed the production of
recombinant human G-CSF (rhG-CSF) (7). The recombinant human protein consists
of two forms: the first form is a 174-amino acid glycosylated
molecule called Lenograstim and the second one is a smaller
non-glycosylated 175-amino acid molecule called G-CSF (7,8). Both
were established as useful clinical agents that are aimed at
increasing the level of granulocytes after the chemotherapy or
radiotherapy (7,9,10).
It has been shown that the reorganization of the
cytoskeleton takes place during cell proliferation, maturation and
differentiation (11–14). Actin filaments are a major component
of the cytoskeleton, which plays an important role in the cellular
processes such as cell migration, growth, cytokinesis, endocytosis,
determination of cell shape and vesicle trafficking. In cells,
actin is found both in a monomeric form (G-actin) and in the form
of filaments (F-actin), which can be arranged in bundles or
networks (15–19). The cytoplasmic functions of actin
are well investigated. However, the role and location of actin in
the cell nucleus has been controversial for decades. The first
evidence for the presence of actin in the nucleus was presented
almost 40 years ago, but was treated with skepticism (20,21).
In recent years, it was demonstrated that actin and actin-binding
proteins play an important role in diverse nuclear activities.
Actin was discovered as a component of chromatin remodeling
complex.
It was also observed that nuclear actin is
associated with the RNA transcription machineries, long-range
chromatin organization and newly synthesized ribonucleoproteins
(RNP) (22–26). Nowadays, it is well known that the
pool of cytoplasmatic and nuclear actin undergoes various
alterations (27–29). Early studies of the effect of G-CSF
showed only the reorganization of actin cytoskeleton as a
consequence of morphological changes in human neutrophils (30). Moreover, it has been shown that this
effect is dependent on the kind of used recombinant human G-CSF
(glycosylated or non-glycosylated) (8). Xu et al demonstrated the
translocation of actin from the cytoplasm to the nucleus during
macrophage differentiation of HL-60 cells (29). In contrast to the HL-60 cell line,
the K562 cell line seems to have no detectable G-CSF receptors and
the effect of G-CSF on this line has not been fully understood so
far. Nevertheless, El-Sonbaty et al showed the influence of
G-CSF on the K562 cell line based on the plasmid-induced expression
of G-CSFR (31–33).
The aim of our studies was to show the influence of
non-glycosylated form of rhG-CSF on the F-actin cytoskeleton and
the morphology of HL-60 and K562 cell lines during their
differentiation.
Materials and methods
Cell culture
The human leukemia cell line HL-60 (ATCC CCL-240)
and erythroleukemia cell line K562 (ATCC CCL-243) were used in the
present study. Both cell lines were grown in RPMI-1640 medium
supplemented with 10% (v/v) heat-inactivated fetal bovine serum
(FBS) and 50 μg/ml gentamycin, in a fully humidified atmosphere of
5% CO2 at 37°C. Cells were incubated with 5 and 10 ng/ml
concentrations of non-glycosylated G-CSF (Amgen Europe B.V.) for 24
h. Control cells were grown under identical conditions, in the
absence of growth factors.
Isolation of nuclei
For analysis of nuclear F-actin content, the
isolation of nuclei was performed. After washing with PBS and
centrifugation, the cells were incubated with homogenizing buffer
consisting of 50 mM Tris-HCl, pH 7.5; 0.3 M sucrose; 5 mM
CaCl2; 5 mM MgCl2; 10 mM 2-mercaptoethanol;
and 0.5% (v/v) Nonidet-P40 substitute (Sigma-Aldrich).
After the homogenization on ice, the cell suspension
was centrifuged and the supernatant was decanted. The sediment was
suspended in 1 ml of homogenized buffer and placed on the top of
the buffer for purification of nuclei, which consists of 50 mM
Tris-HCl, pH 7.5; 0.3 M sucrose; 5 mM KCl; 5 mM MgCl2;
10 mM 2-mercaptoethanol (Sigma-Aldrich); and 41% glycerol (Roth).
Following centrifugation, the nuclei were washed with PBS and fixed
in 4% paraformaldehyde (Serva).
Classical fluorescence and confocal
microscopy
Control cells and cells treated with G-CSF were
fixed in 4% paraformaldehyde (Serva) for 20 min, washed three times
with PBS and centrifuged onto glass slides. Cells on slides were
permeabilized in 0.1% Triton X-100 in PBS for 5 min and washed with
PBS. Next, the cells were incubated with phalloidin conjugated to
Alexa Fluor® 488 (Invitrogen, 1:40) for 20 min, at room
temperature (RT). Nuclear staining was performed with
4′,6′-diamidino-2-phenylindole dihydrochloride (DAPI,
Sigma-Aldrich, 100 ng/ml) for 10 min. After counter-staining, the
cells were rinsed with PBS and mounted in Aqua-Poly/Mount
(Polysciences Inc.). The cells were examined using an Eclipse E800
fluorescence microscope (Nikon) and C1 confocal microscope
(Nikon).
Flow cytometry
Cytoplasmic and nuclear F-actin
staining
Cells and isolated nuclei were fixed in 4%
paraformaldehyde for 20 min and afterwards rinsed three times with
PBS. Then, the cells and nuclei were stained for F-actin with
phalloidin conjugated to Alexa Fluor 488 (Invitrogen, 1:40) for 20
min. After washing with PBS, DNA was labeled with
7-aminoactinomycin (7-AAD, Sigma-Aldrich) for 5 min in RT. All
reaction steps, during which the fluorochrome was used, were
performed in the dark. The samples were analyzed using
Becton-Dickinson FACScan flow cytometer.
Cell cycle analysis
The cells were stained with PI solution consisting
of 50 μg/ml propidium iodide (PI) and 0.03% (v/v)
nonylphenylpolyethylene glycol (Nonidet-P40 substitute;
Sigma-Aldrich) for 15 min in the dark (RT). After centrifugation
and supernatant removal, the cells were incubated with RNase A
solution for 15 min in the dark (RT). The percentage of cells in
respective phases of the cell cycle was analyzed by
Becton-Dickinson FACScan flow cytometer.
Double-staining with Annexin V and
7-AAD
In order to detect the apoptotic and necrotic cell
death, cells were stained with using the Annexin V-FITC Apoptosis
Detection kit (BD Biosciences Pharmingen) according to the
manufacturer’s instructions. After washing with PBS, the
supernatant was incubated with 195 μl of Annexin V binding buffer
and 5 μl of Annexin V-FITC. Following the 15-min incubation in the
dark (RT) and centrifugation, the cells were incubated with 190 μl
of Annexin V binding buffer and 10 μl of 7-AAD for 5 min in the
dark. Then cells were detected by Becton-Dickinson FACScan flow
cytometer.
Transmission electron microscopy
For observation of the morphology and ultrastructure
of HL-60 and K562 cells, a transmission electron microscopy was
used. The control cells and cells treated with G-CSF were washed
with PBS and fixed in 3.6% (v/v) glutaraldehyde (Merck) for 30 min
(RT). After washing with 0.1 M sodium cacodylate buffer
(Sigma-Aldrich), the cells were entrapped in fibrin clot and
postfixed in 2% (w/v) OsO4 (Serva) in 0.1 M cacodylate buffer for 1
h (RT). Then the cells were passed through a series of ethanol and
acetone solutions and embedded in Epon 812 (Roth). Ultrathin
sections were cut and double-stained with uranyl acetate and lead
citrate. The samples were examined using a JEM 100CX transmission
electron microscope (JEOL).
In order to determine the localization of F-actin at
the ultrastructural level, the cells were fixed in 4%
paraformaldehyde for 20 min (RT), washed three times with PBS and
incubated with 0.1% Triton X-100 in PBS for 5 min. Then, the cells
were rinsed with PBS and treated with endogenous biotin blocking
kit (Invitrogen) and 6% BSA in PBS (bovine serum albumin, Sigma
Aldrich) for 1 h (RT). Next, the cells were incubated for 20 min
with biotinylated phalloidin (Sigma-Aldrich) diluted 1:85 from
stock solution. This step was followed by rinsing with PBS,
postfixation in 2% (w/v) OsO4 (Serva), dehydration in series of
ethanol and embedding in LR White. After the LR White
polymerization, the samples were cut by the ultramicrotome and
collected on nickel grids (Sigma). Then, cells were treated with
0.1% Triton X-100 in PBS for 5 min and three times with 0.001%
Tween-20 in TBS. F-actin was stained for 1 h in the dark using
Qdots® 525 streptavidin conjugate (Invitrogen) diluted
1:50 in PBS. The preparations were examined using a JEM 100CX
(JEOL) transmission electron microscope.
Statistical analysis
To determine the differences between control cells
and cells treated with G-CSF, the non-parametric Mann-Whitney U
test was used. Results were considered significant at
P<0.05.
Results
Fluorescent staining of F-actin
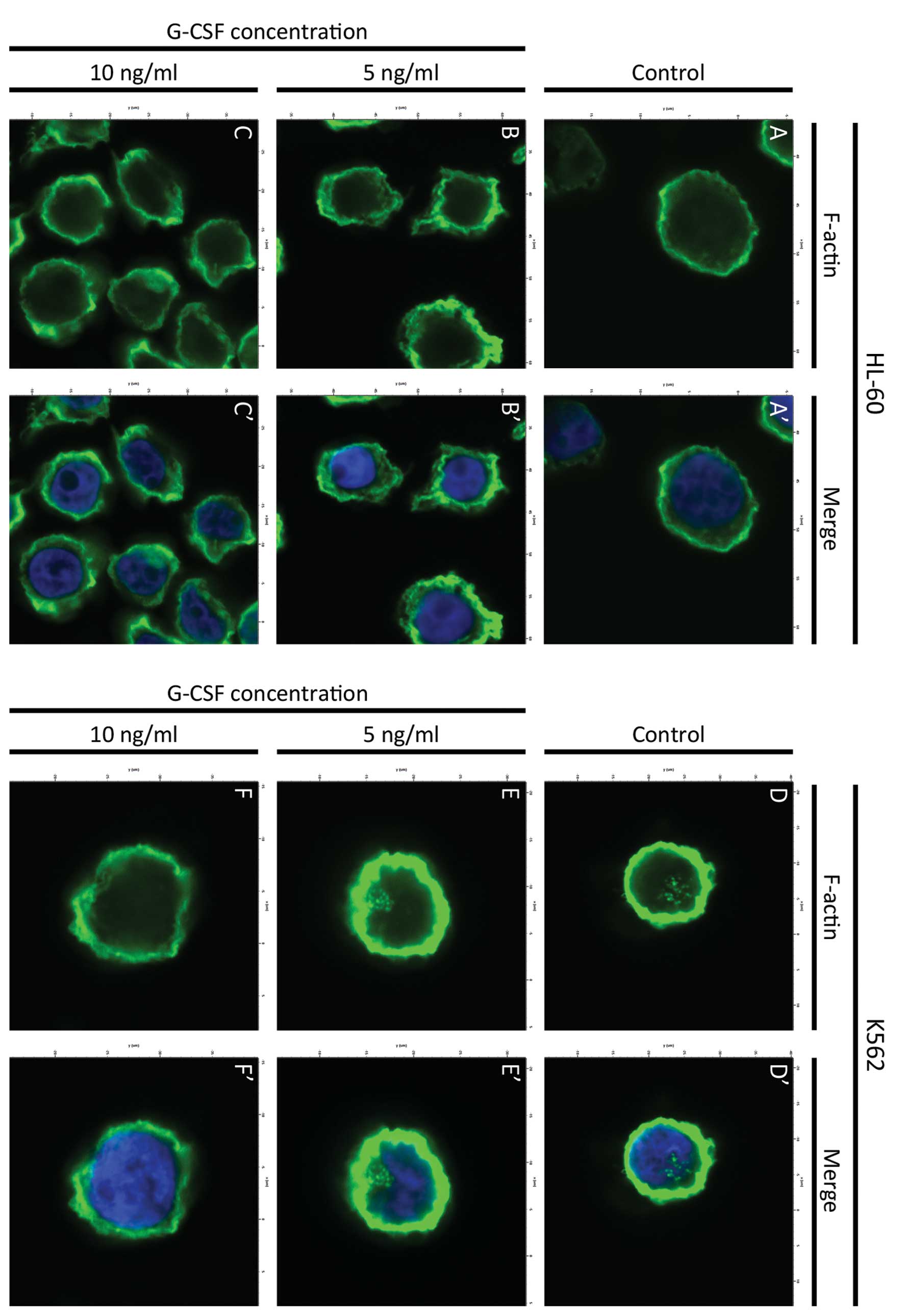
The reorganization of F-actin cytoskeleton in HL-60
and K562 cell lines after treatment with G-CSF was estimated using
phalloidin conjugated to Alexa Fluor 488. It was shown that in
HL-60 control cells, the F-actin fluorescence was increased on the
cell periphery. There were also observed low fluorescence signals
in the form of small aggregates in the cytoplasm (Figs. 1A′, A″ and 2A, A′). Visualization of actin filaments
in HL-60 cells treated with the lowest dose of G-CSF (5 ng/ml) in
both classical fluorescence and confocal microscope, revealed a
higher fluorescence intensity in whole cells, compared to the
control. In these cells small aggregates and short fibers of
F-actin were formed (Figs. 1B′, B″
and 2B, B′). At 10 ng/ml
concentration of G-CSF, the reorganization of F-actin was more
evident and much more networks and aggregates of actin were
observed, compared to the control cells and cells treated with the
lowest G-CSF dose (Figs. 1C′, C″
and 2C, C′). Moreover, at both
doses of G-CSF, only few cells with apoptotic features were
observed (Fig. 1B and C).
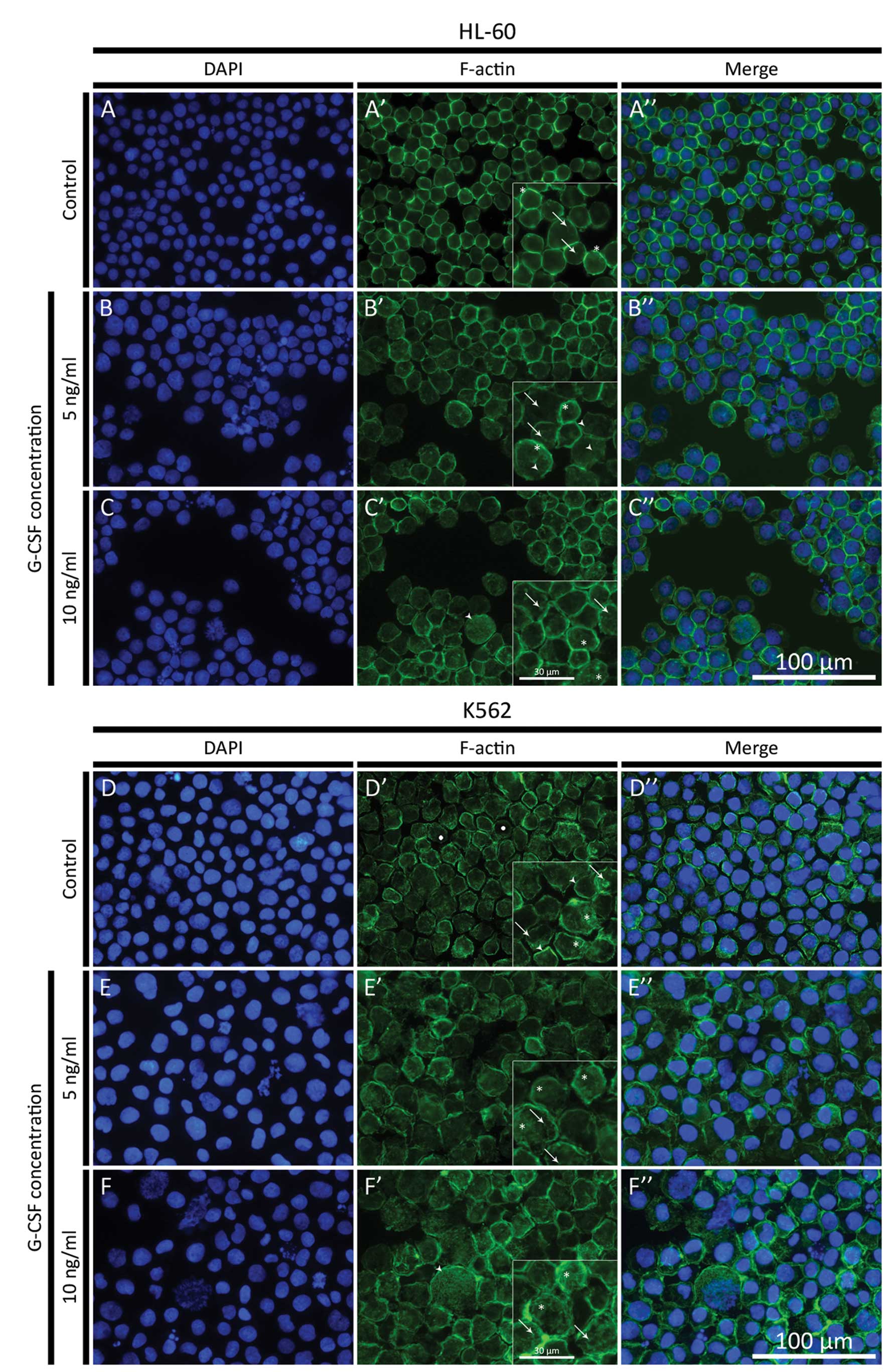 | Figure 1(A–C) Fluorescence microscopic
changes in F-actin cytoskeleton organization in HL-60 cell line.
The cells were treated with 5 and 10 ng/ml G-CSF. (A–C) DAPI;
(A′–C′) F-actin; (A″–C″) Merge. (A) HL-60 control cells (asterisks,
ring-like structures formed by F-actin on the cell periphery;
arrows, F-actin aggregates in the cytoplasm); (B) HL-60 cells
treated with 5 ng/ml G-CSF (asterisks, layer of F-actin beneath the
plasma membrane; arrows, F-actin aggregates; arrowheads, short
F-actin fibers); (C) HL-60 cells treated with 10 ng/ml G-CSF
(arrows, F-actin aggregates in the cytoplasm; arrowhead, high
F-actin fluorescence signal in giant cell; asterisks, F-actin
networks). (D–F) Fluorescence microscopic changes in F-actin
cytoskeleton organization in K562 cell line. The cells were treated
with 5 and 10 ng/ml G-CSF. (D–F) DAPI; (D′–F′) F-actin; (D″–F″)
Merge. (D) K562 control cells (arrows, F-actin aggregates in the
cytoplasm; arrowheads, ring-like structures formed by F-actin on
the cell periphery; asterisks, F-actin networks; dots, low F-actin
fluorescence signal in the area of nucleus); (E) K562 cells treated
with 5 ng/ml G-CSF (arrows, F-actin aggregates in the cytoplasm;
asterisks, F-actin networks); (F) K562 cells treated with 10 ng/ml
G-CSF (arrows, F-actin aggregates in the cytoplasm; arrowhead, high
F-actin fluorescence signal in giant cell; asterisks, F-actin
networks). |
In K562 control cells, many different forms of
F-actin were observed. Beside the high fluorescence intensity on
the cell periphery, the F-actin formed small aggregates and a
network in the cytoplasm (Fig. 1D′ and
D″). In the area of the cell nucleus, the fluorescence
intensity of F-actin was high in form of aggregates (Fig. 2D and D′). After treatment with 5
ng/ml of G-CSF a cytoplasmic F-actin labeling, in the form of
aggregates and networks, was seen (Fig.
1E′ and E″), however, nuclear staining was decreased (Fig. 2E and E′). After 10 ng/ml of G-CSF,
F-actin labeling was also decreased in both cytoplasm and cell
nucleus, as compared to control (Fig.
2F and F′). DAPI staining showed that only few nuclei of cells
treated with the highest dose of G-CSF displayed apoptotic features
(Fig. 1D–F) and these cells were
characterized by low fluorescence intensity of F-actin (Fig. 1E″ and F″). High signal of F-actin
fluorescence in giant HL-60 and K562 cells was also observed
(Fig. 1C′, C″, F′ and F″).
Transmission electron microscopy
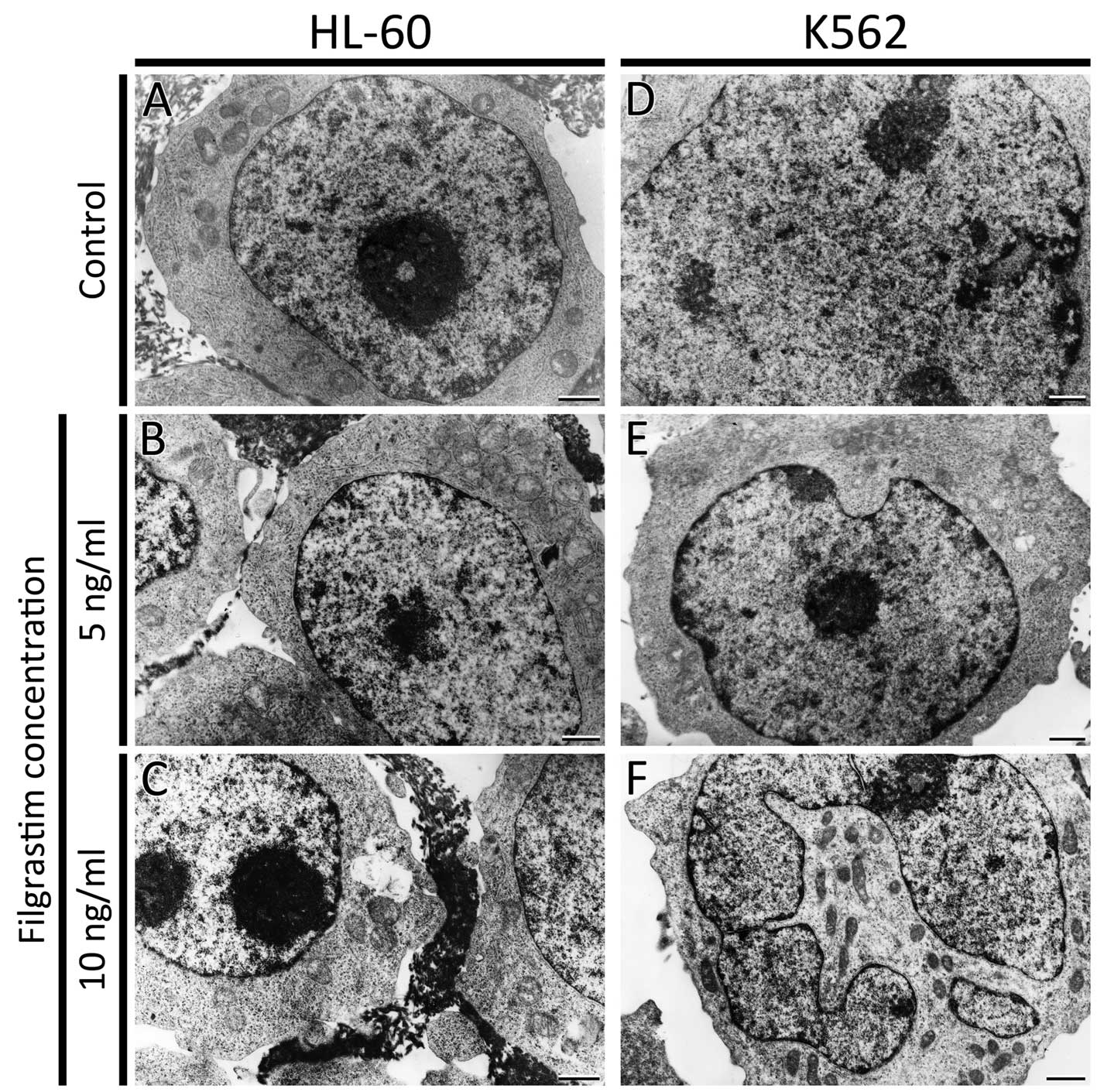
HL-60 and K562 cells were examined by transmission
electron microscopy. The ultrastructure of HL-60 cells, after
treatment with both doses of G-CSF, was unchanged. The control
cells and cells incubated with G-CSF showed normal morphology with
an oval nucleus. There were no changes observed in mitochondria,
Golgi apparatus and endoplasmic reticulum (Fig. 3A–C). K562 cells morphology after
treatment with the lowest dose of G-CSF, was comparable to the
control (Fig. 3D and E). However,
in few K562 cells treated with 10 ng/ml of G-CSF multi-lobular
nuclei were noted (Fig. 3F).
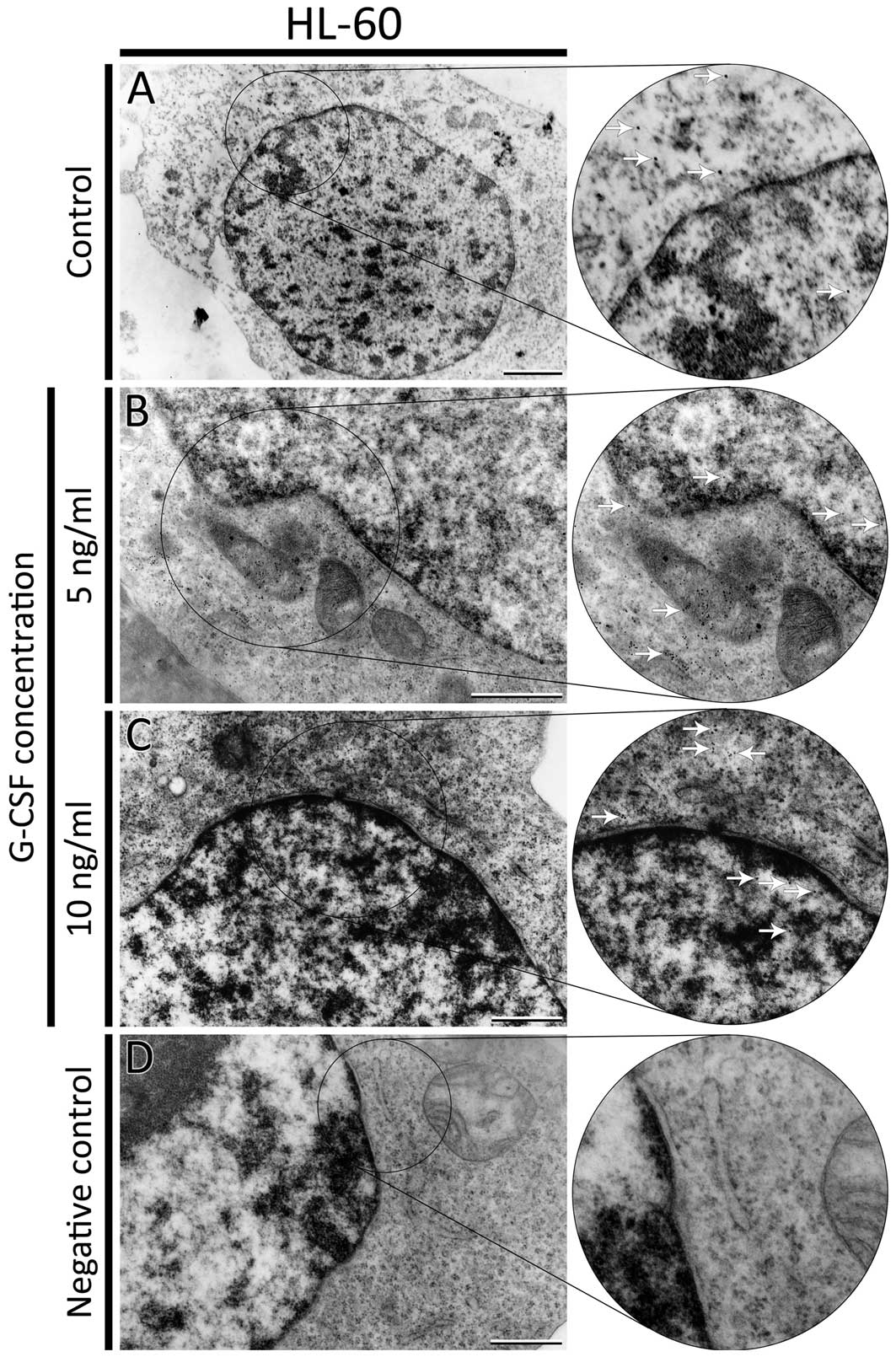
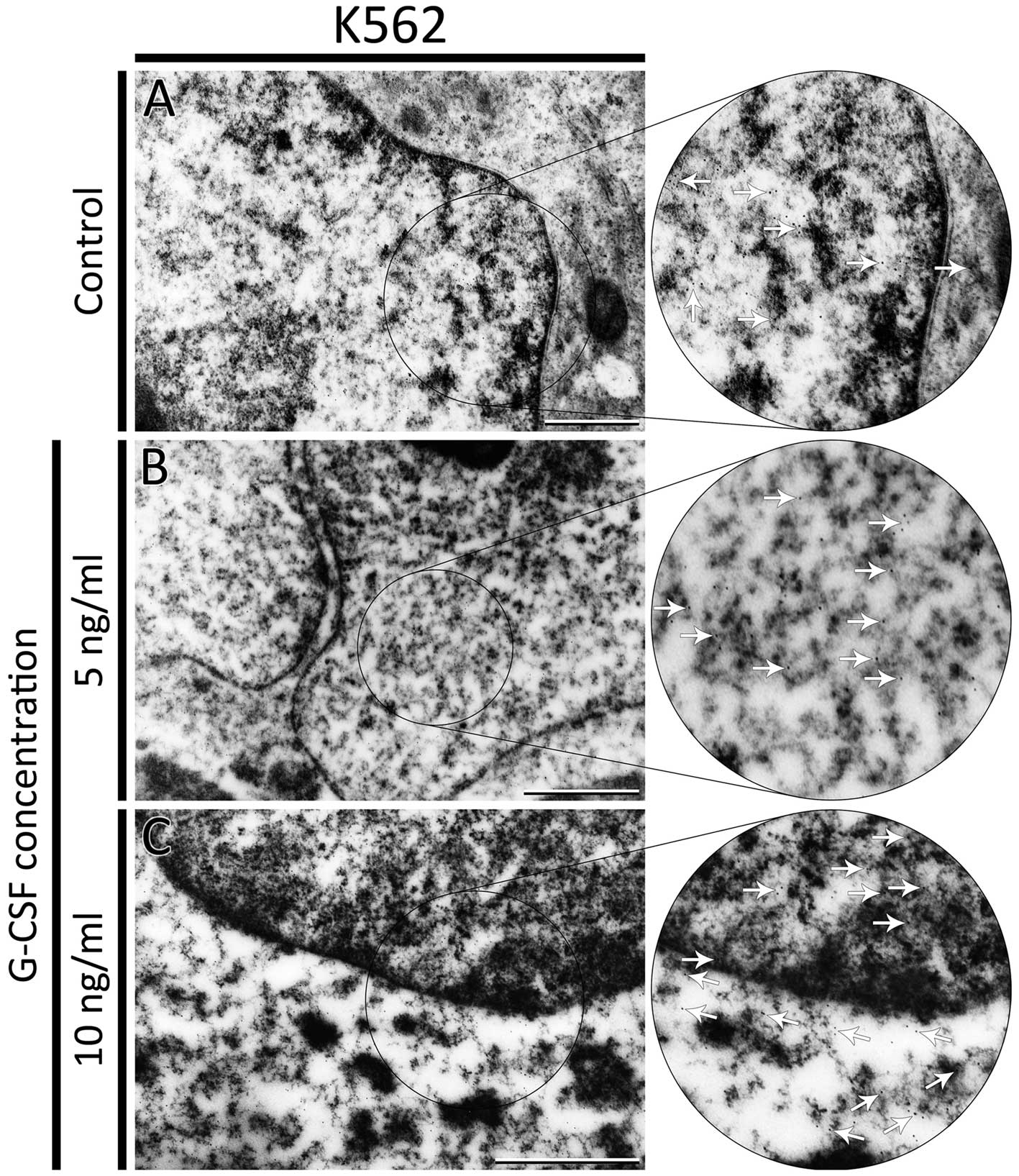
F-actin was also visualized at the ultrastructural
level using biotinylated phalloidin and Qdots semiconductor
nanocrystals conjugated with streptavidin. Quantum dots were found
in control cells and cells treated with all doses of G-CSF. In
HL-60 control cells, nanocrystals were observed in the cytoplasm
and nucleus (Fig. 4A). After
treatment with 5 ng/ml of G-CSF, a lot of Qdots were localized
mainly in the cytoplasm. In the nucleus, single aggregates of
nanocrystals were observed close to the heterochromatin and nuclear
membrane (Fig. 4B). In HL-60 cells
incubated with 10 ng/ml of G-CSF only few aggregates of Qdots were
localized in the cytoplasm and nucleus (Fig. 4C). In K562 control cells, the Qdots
were observed mainly in the nucleus close to the heterochromatin
(Fig. 5A). After treatment of K562
cells with 5 and 10 ng/ml of G-CSF nanocrystals were seen in both
cytoplasm and nucleus. However, the number of nanomolecules in the
nucleus was smaller than in the control (Fig. 5B and C). Moreover, after treatment
with 10 ng/ml of G-CSF, the number of Qdots in cytoplasm was
greater. For negative control, the cells were incubated as
previously described but without biotinyled phalloidin (Fig. 4D).
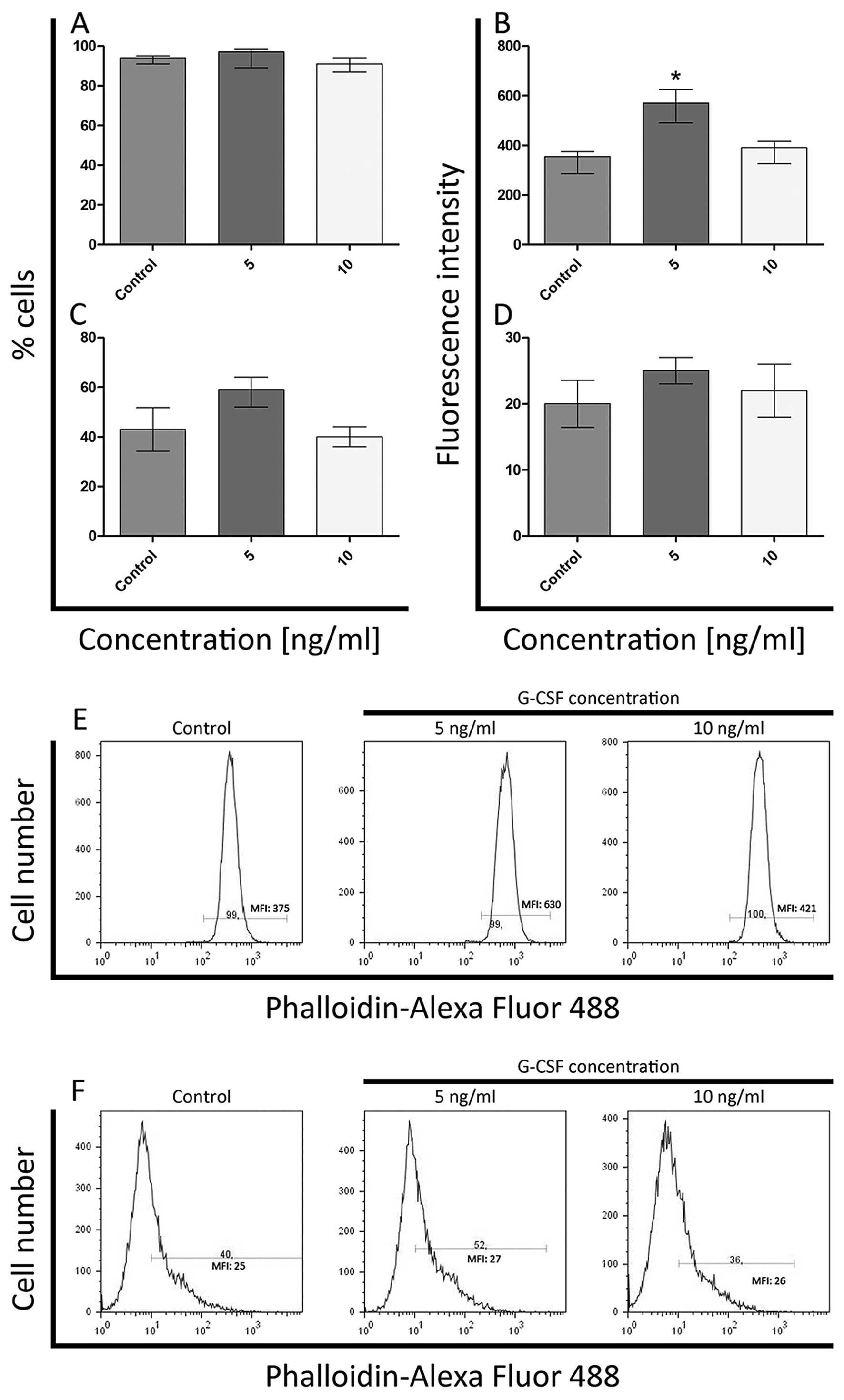
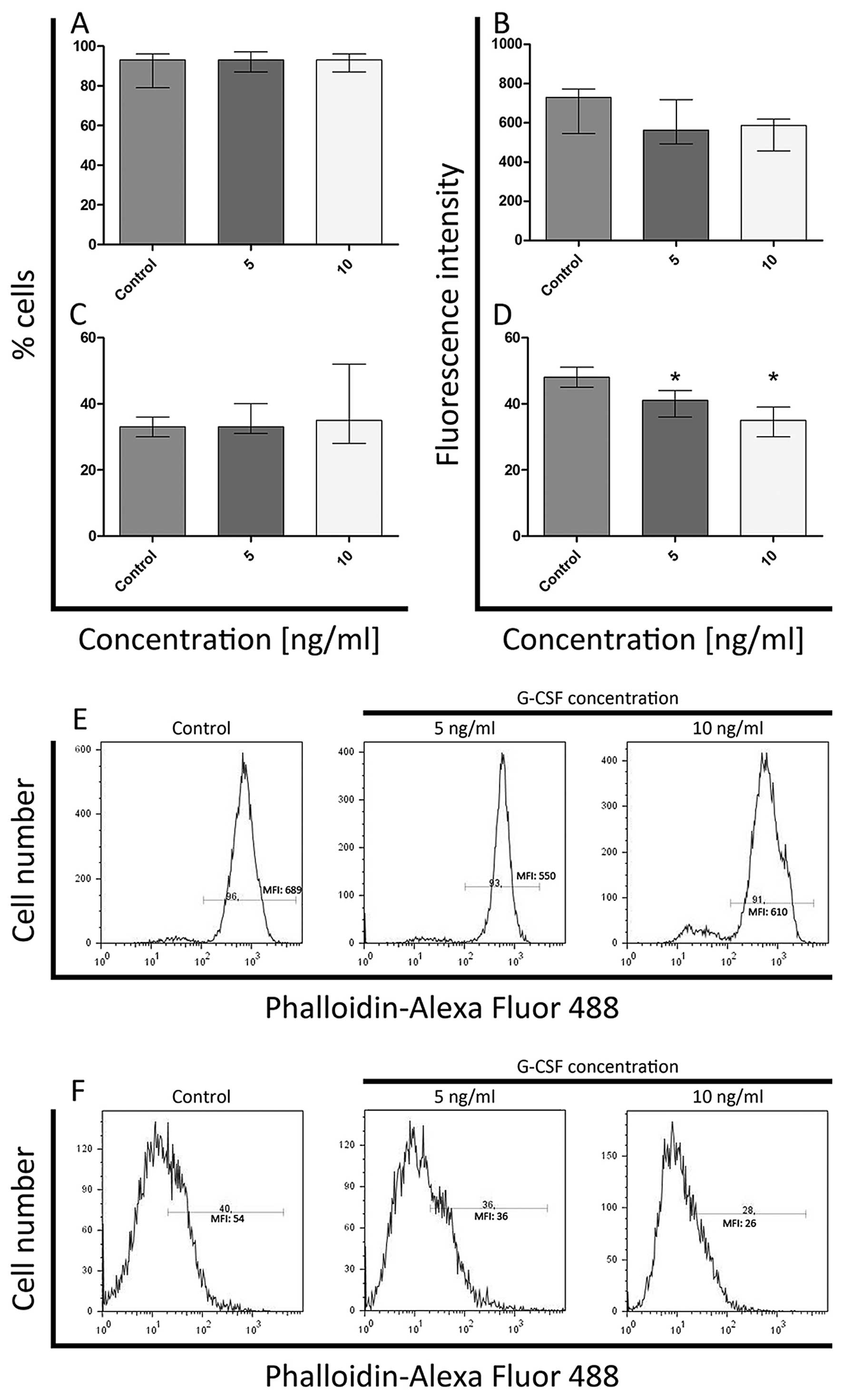
Flow cytometric analysis
F-actin staining
Flow cytometric analysis after the G-CSF treatment
demonstrated quantitative changes in F-actin fluorescence staining
both in whole cells and isolated nuclei in comparison to the
control. There were not observed statistically significant
differences in the percentage of F-actin-positive cells incubated
with G-CSF in comparison to the control cells. The levels of
F-actin ranged from 99 to 100% in HL-60 cell line and in K562 cells
from 91 to 96% (Figs. 6A and
7A). The significant increase
(P<0.05) of F-actin level in HL-60 cells was observed only after
exposure to 5ng/ml of G-CSF (Fig.
6B). At 10 ng/ml of G-CSF, the fluorescence intensity was
comparable to the control (Fig.
6B). In whole K562 cells, the statistically significant
differences in fluorescence intensity of F-actin were not observed
(Fig. 7B).
In the case of cell nuclei treated with G-CSF, there
were no statistically significant differences in the percentage of
F-actin-positive nuclei in both cell lines and in F-actin
fluorescence intensity in isolated HL-60 nuclei (Figs. 6C, D and 7C). Whereas in K562 cell nuclei, the
fluorescence intensity of F-actin decreased in a dose-dependent
manner (it was the lowest after treatment with 10 ng/ml
concentration) and statistically significant differences were
observed (Fig. 7D).
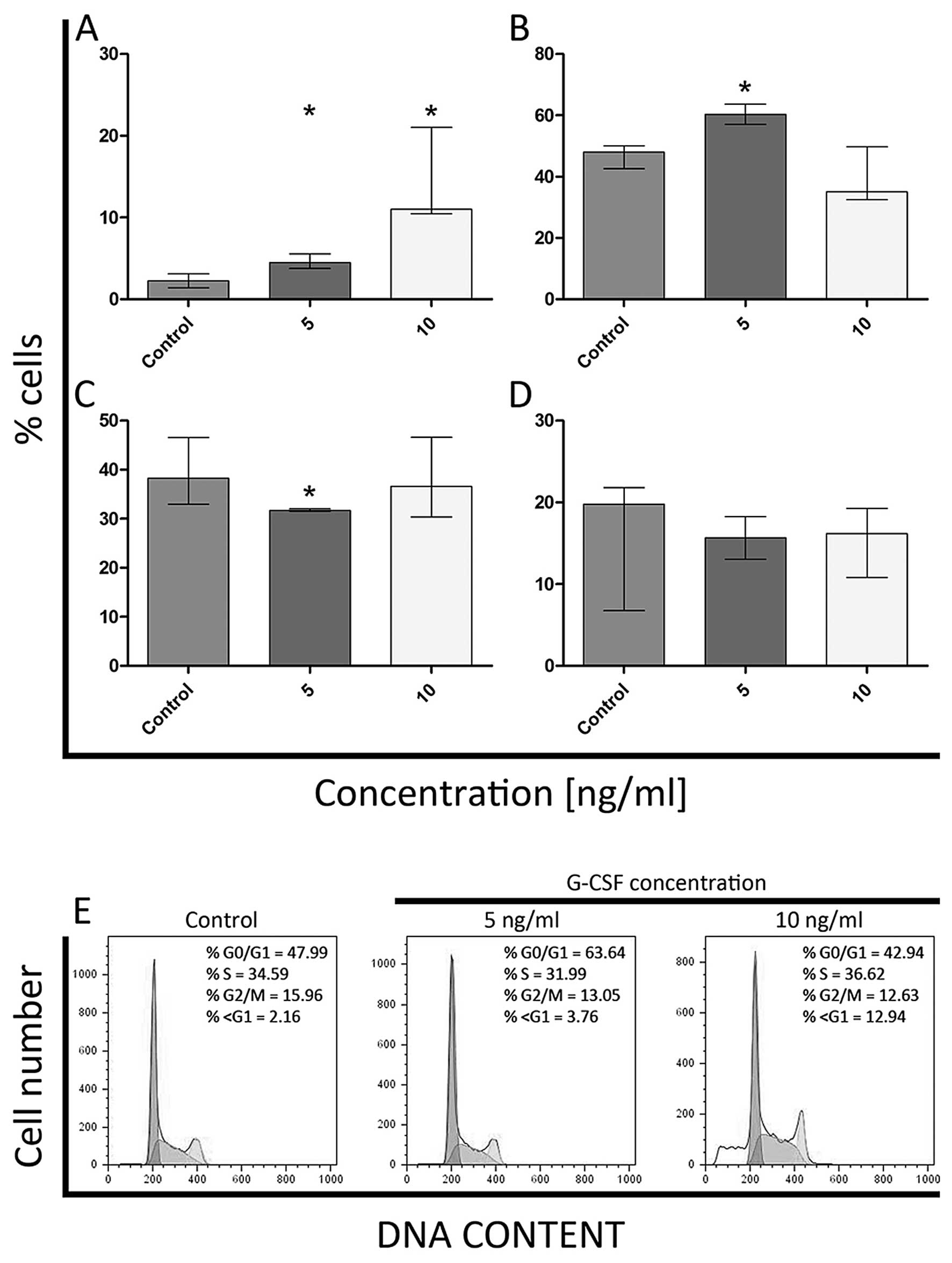
Cell cycle analysis
In order to examine the effect of granulocyte colony
stimulating factor (G-CSF) on HL-60 and K562 cell cycle
progression, standard and widely used method that employs PI/RNase
was applied. Accordingly, there was observed a statistically
significant increase in the mean percentage of cells in G0/G1 and
sub-G1 phases after treatment of HL-60 cells with 5 ng/ml of G-CSF,
as compared to the control (Fig. 8A and
B). On the other hand, after treatment of HL-60 cells with the
same dose of G-CSF, the percentage of S-phase cells was decreased
(Fig. 8C). Moreover, after
incubation with 10 ng/ml of G-CSF, a significant increase in the
percentage of cells in sub-G1 phase was noticed (Fig. 8A). There were no statistically
significant differences in the percentage of HL-60 cells in G2/M
phase for both doses of G-CSF (Fig.
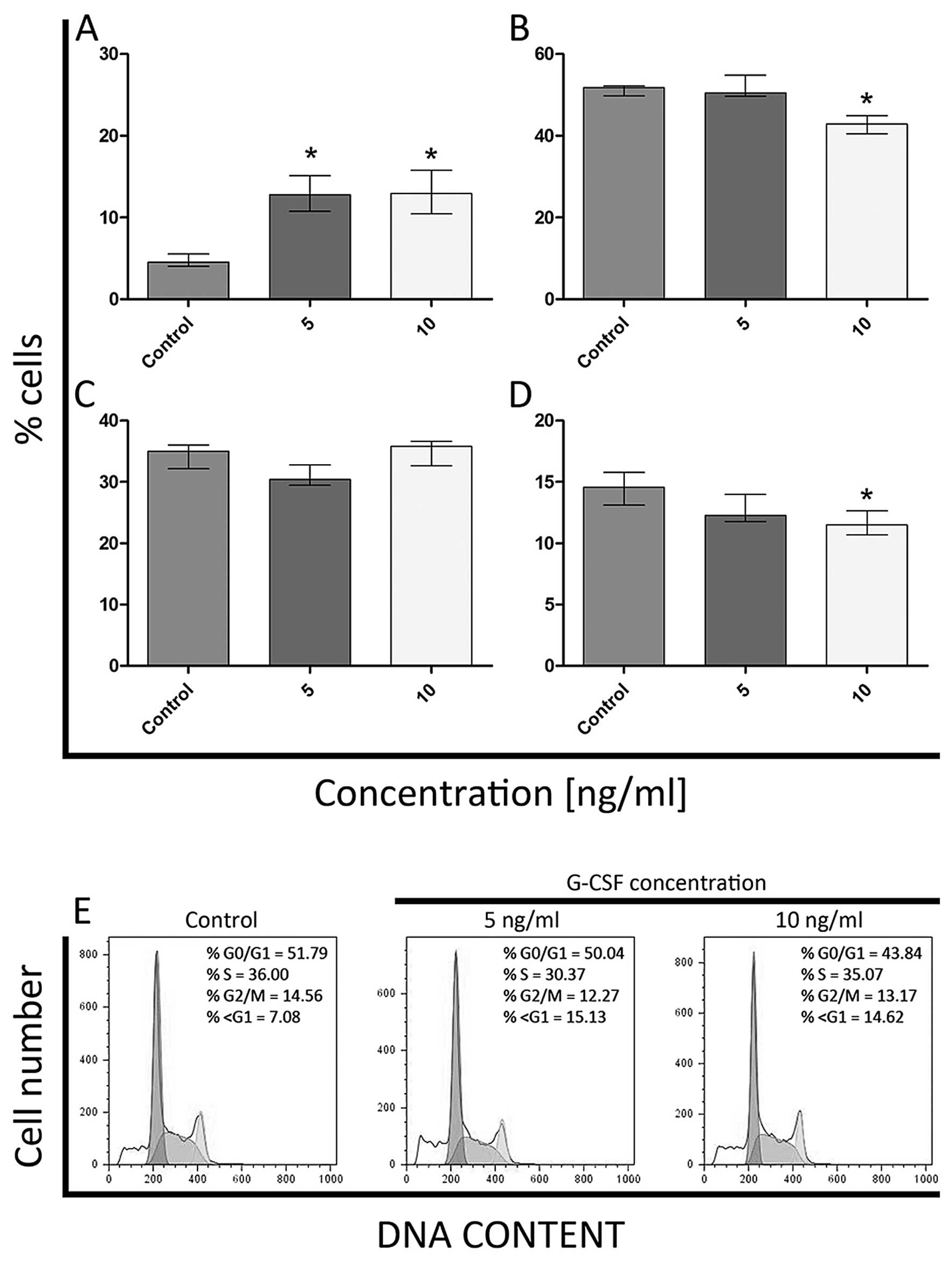
8D). As far as K562 cell line is concerned, a significant
increase in the percentage of cells in sub-G1 phase was observed
following treatment with 5 ng/ml of G-CSF (Fig. 9A). Even though the number of cells
classified as G0/G1 and G2/M was decreased after treatment with 10
ng/ml of G-CSF, the percentage of cells with DNA content typical of
sub-G1 increased, in comparison to the control. These differences
were statistically significant (Fig.
9A, B and D). Moreover, the statistically significant
differences in the percentage of cells in S phase were not observed
(Fig. 9C).
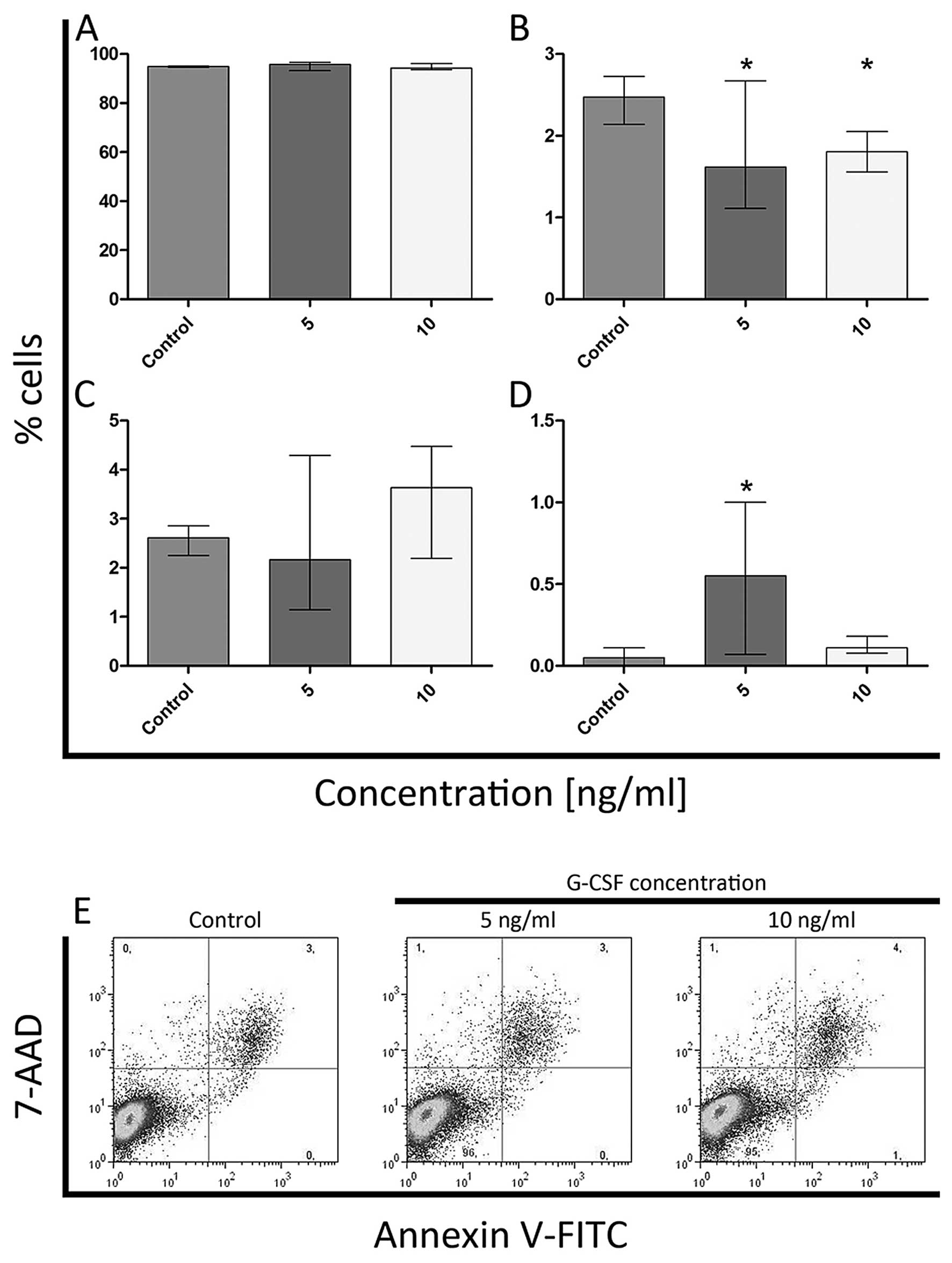
Annexin V and 7-AAD
double-staining
The flow cytometric analysis indicated a decrease in
the number of early apoptotic cells after treatment of HL-60 cells
with both doses of G-CSF (Fig.
10B). At the 5 ng/ml concentration of G-CSF, there was also
seen a significant increase in the number of necrotic cells (only
7-AAD positive) (Fig. 10D).
However, the statistically significant differences in the
percentage of viable and late apoptotic cells were not observed
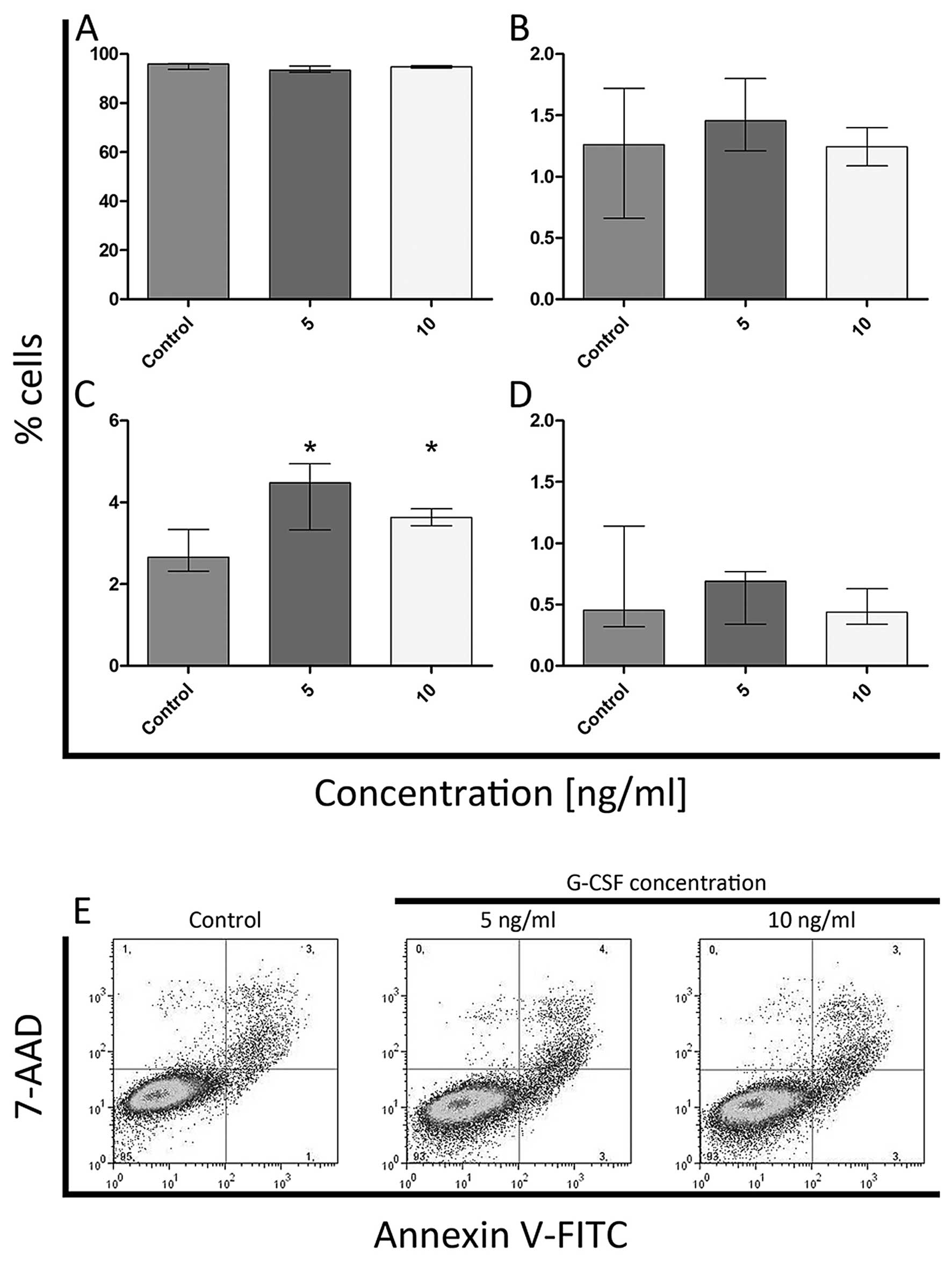
(Fig. 10A and C). In K562 cell
line, an increase in the percentage of late apoptotic cells was
noted after treatment with both doses of G-CSF (Fig. 11C) but the statistically
significant differences in the percentage of viable, early
apoptotic and necrotic cells were not observed (Fig. 11A, B and D).
Discussion
In the present study, we showed the changes of
F-actin distribution in HL-60 cells after treatment with
non-glycosylated G-CSF. Although, K562 cell line has no detectable
G-CSF receptors, changes in the organization and fluorescence
intensity of F-actin in isolated nuclei were observed. Moreover,
the fluorescence microscopic results showed that stimulation of
HL-60 cell line with G-CSF caused F-actin reorganization, including
the formation of aggregates and short fibers in the cytoplasm and
its conglomeration at the cell surface. Similar results were
reported by Veselska et al, who observed the same structures
of F-actin during the incubation of HL-60 cells with ATRA and PMA
(34). We suggest that observed
reorganization of filamentous actin is associated with
differentiation process and that surface-associated structures may
participate in the apoptotic bleb formation. Such an explanation is
reasonable because, in our experiment, some HL-60 cell nuclei
showed apoptotic features and the significant increase in the
percentage of cells in sub-G1 phase after treatment with both doses
of G-CSF was shown. Furthermore, the G1 phase arrest is considered
to be a marker of cell differentiation, which was confirmed in the
present study during treatment of HL-60 cells with 5 ng/ml of G-CSF
(35,36). After differentiation, the cells were
most probably switched to apoptotic pathway. An accumulation of
F-actin in the apoptotic cells in HL-60 cells after treatment with
ATRA and doxorubicin was noticed (37,38).
It is well known now, that the processes of differentiation and
apoptosis are connected with the reorganization of the
cytoskeleton, but the mechanism of this linkage still remains
unclear (30,34,39–41).
Some scientists consider that the differentiation process is
associated with actin polymerization whereas others suggest its
relation to depolymerization (8,42,43).
Moreover, we observed statistically significant increase of F-actin
level in HL-60 cell line after exposure to 5 ng/ml of G-CSF. This
observation was confirmed by flow cytometry and transmission
electron microscopy. Moreover, our previous study also showed the
increase of F-actin level in HL-60 cells after treatment with G-CSF
(41). Furthermore, the increase of
F-actin level in neutrophils was shown by Carulli et al and
Gomez-Cambronero et al as a result of G-CSF treatment
(8,43). Similarly, Chodniewicz and Zhelev
also noted the higher content of actin filaments in granulocytes
incubated with GM-CSF (42).
However, the opposite results were observed by Kutsuna et al
in human neutrophils. They observed a cytokine-induced actin
depolymerization (30). We are in
agreement with the authors who suggested that F-actin
polymerization and depolymerization depends on the cytokine doses
and the time of incubation (8,30,42,43).
In the past decade, many studies have been focused
on the development of new nanomaterials and their interactions with
cells (44). Here, we showed the
usefulness of semiconductor CdSe/ZnS nanocrystals in the
localization of F-actin at the ultrastructural level. The results
presented in this paper are the ultrastructural evidence of
phalloidin-based localization of F-actin using transmission
electron microscopy. Up to now, the localization of cytoskeletal
proteins in TEM was performed by a direct or indirect method using
antibodies. However, there was no evidence concerning
phalloidin-based localization of actin filaments. Our previous data
showed the presence of F-actin in the cell nucleus, that was
determined only by using fluorescence techniques, including flow
cytometer analysis of isolated nuclei and laser scanning confocal
microscopy (45,46). In the present study, we combined
pre- and post-embedding methods and that was the key to successful
detection of F-actin at the ultrastructural level. Moreover, the
use of colloidal gold for phalloidin-based method of F-actin
localization, made it impossible to localize actin filaments or
showed very weak delectability (data not shown). As regards the
results obtained by using fluorescence microscopy and flow
cytometry, we showed changes in F-actin distribution at the
ultrastructural level in the area of the cytoplasm and cell nucleus
after treatment of HL-60 and K562 cell lines with different G-CSF
doses.
In 2010, Xu et al demonstrated the nuclear
translocation of β-actin, one of the actin isoform, during the
macrophage differentiation of HL-60 cells. It was shown that this
actin isoform is translocated from the cytoplasm to the nucleus and
plays an important role in the regulation of the transcription
process (29). It is known, that
actin and actin-binding proteins are involved in chromatin
remodeling and gene transcription. In our previous study, we
observed that actin was associated with the heterochromatin during
apoptosis (38,46,47).
Moreover, Zhao et al showed that F-actin is directly
involved in the chromatin reorganization (48). Influence of actin on the
translocation of transcriptional RNA was presented by Hofmann et
al and Widlak et al (24,49).
They described the participation of F-actin in the processing of
retroviral RNA and transport of protein kinase inhibitor (PKI).
Influence of actin on the regulation of RNAP II-mediated
transcription was observed by many other researchers (24,29,50).
In the present study we observed the increase of F-actin level
after treatment of HL-60 cells with 5 ng/ml concentration of G-CSF
in cytometric analysis, and a higher labeling of nuclear F-actin
under transmission electron microscope was observed. However,
different observations were made during the analyses with the use
of K562 cell line. Even though these cells do not express the
receptor for G-CSF, some changes were seen in the present study in
comparison to the control. We observed a statistically significant
increase in the percentage of cells in sub-G1 phase following
treatment with 5 and 10 ng/ml concentration of G-CSF. There was
also an increase in the percentage of late apoptotic cells after
treatment with both doses of G-CSF. On the other hand, the
statistically significant difference in the cytoplasmic F-actin
level was not detected, but its nuclear level was decreased. We
believe that a decreased level of F-actin content in the cell
nuclei may be due to the reorganization of the chromatin, which can
be supported by the increase of the population of cells in the
subG1 phase and in the percentage of late apoptotic cells.
It is known that G-CSF has no detectable effect on
some of the hematopoietic cell lines such as U-937, WEHI-3B and
K562 (33). However, it has been
also shown that granulocyte-colony stimulating factors could
promote a time- and dose-dependent increase in ROS production.
Additionally, Zhu et al suggested that the G-CSF-induced
Lyn-PI3KAkt pathway controls ROS production in a myeloid cell line
(51). However, Kitagawa et
al indicated the G-CSF-enhanced the effect of Ara-C in U937 and
WEHI-3B (52). They found that
G-CSF mobilized G0/G1-phase cells into the S phase in U937 cells.
On the basis of our present studies, we suggest that G-CSF can act
by a receptor-independent pathway or through another homologous
receptor. As shown by Chow et al the G-CSF receptor reveals
46% homology in sequence to gp130, the IL-6 receptor signal
transducer, and has an identical domain structure (53). Moreover, the overall structure of
the receptor for G-CSF is similar to the leptin and LIF receptor
(54,55).
In summary, we conclude that the G-CSF-based
reorganization of actin filaments is involved in the
differentiation process in HL-60 cell line. Moreover, its
polymerization and depolymerization depends mostly on G-CSF
concentration. As far as K562 cell line is concerned, we observed
changes at the flow cytometry, fluorescence and transmission
electron microscopy level. Although, K562 cell line has no
detectable G-CSF receptors, in our studies it was shown that the
G-CSF-induced statistically significant effect on the cell cycle,
apoptosis and F-actin fluorescence intensity in the cell nuclei. We
suppose that these changes are associated with the G-CSF
receptor-independent pathway or its binding to other similar
receptors. However, in order to find the exact mechanism of its
influence on the cells without G-CSF receptors, further
investigations are needed. Moreover, we showed here that CdSe/ZnS
quantum dots are useful in the localization of subcellular
structures at the ultrastructural level and that our
phalloidin-based method allows determination of the presence of
F-actin in the cell nucleus by transmission electron
microscopy.
Acknowledgements
This study was supported by research task within the
framework of the statutory activities no. 585 (Nicolaus Copernicus
University in Torun, Collegium Medicum in Bydgoszcz, Poland).
References
|
1
|
Oppenheim JJ: Cytokines: past, present and
future. Int J Hematol. 74:3–8. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Basu S, Dunn A and Ward A: G-CSF: function
and modes of action. Int J Mol Med. 10:3–10. 2002.PubMed/NCBI
|
|
3
|
Beekman R and Touw IP: G-CSF and its
receptor in myeloid malignancy. Blood. 115:5131–5136. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Franzke A: The role of G-CSF in adaptive
immunity. Cytokine Growth Factor Rev. 17:235–244. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Leavey PJ, Sellins KS, Thurman G, et al:
In vivo treatment with granulocyte colony-stimulating factor
results in divergent effects on neutrophil functions measured in
vitro. Blood. 92:4366–4374. 1998.PubMed/NCBI
|
|
6
|
Suzuki S, Kobayashi M, Chiba K, et al:
Autocrine production of epithelial cell-derived neutrophil
attractant-78 induced by granulocyte colony-stimulating factor in
neutrophils. Blood. 99:1863–1865. 2002.PubMed/NCBI
|
|
7
|
Bradstock KF: The use of hematopoietic
growth factors in the treatment of acute leukemia. Curr Pharm Des.
8:343–355. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Carulli G, Mattii L, Azzara A, et al:
Actin polymerization in neutrophils from donors of peripheral blood
stem cells: divergent effects of glycosylated and nonglycosylated
recombinant human granulocyte colony-stimulating factor. Am J
Hematol. 81:318–323. 2006. View Article : Google Scholar
|
|
9
|
Håkansson L, Höglund M, Jönsson UB,
Torsteinsdottir I, Xu X and Venge P: Effects of in vivo
administration of G-CSF on neutrophil and eosinophil adhesion. Br J
Haematol. 98:603–611. 1997.PubMed/NCBI
|
|
10
|
Ripa RS and Kastrup J: G-CSF therapy with
mobilization of bone marrow stem cells for myocardial recovery
after acute myocardial infarction - a relevant treatment? Exp
Hematol. 36:681–686. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ku NO, Zhou X, Toivola DM and Omary MB:
The cytoskeleton of digestive epithelia in health and disease. Am J
Physiol. 277:G1108–G1137. 1999.PubMed/NCBI
|
|
12
|
Bursch W, Hochegger K, Torok L, Marian B,
Ellinger A and Hermann RS: Autophagic and apoptotic types of
programmed cell death exhibit different fates of cytoskeletal
filaments. J Cell Sci. 113:1189–1198. 2000.PubMed/NCBI
|
|
13
|
Pollard TD: The cytoskeleton, cellular
motility and the reductionist agenda. Nature. 422:741–745. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ndozangue-Touriguine O, Hamelin J and
Bréard J: Cytoskeleton and apoptosis. Biochem Pharmacol. 76:11–18.
2008. View Article : Google Scholar
|
|
15
|
Hightower RC and Meagher RB: The molecular
evolution of actin. Genetics. 114:315–332. 1986.PubMed/NCBI
|
|
16
|
Akisaka T, Yoshida H, Inoue S and Shimizu
K: Organization of cytoskeletal F-actin, G-actin, and gelsolin in
the adhesion structures in cultured osteoclast. J Bone Miner Res.
16:1248–1255. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
White S, Williams P, Wojcik KR, Sun S,
Hiemstra PS, Rabe KF and Dorscheid DR: Initiation of apoptosis by
actin cytoskeletal derangement in human airway epithelial cells. Am
J Respir Cell Mol Biol. 24:282–294. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ascough KR: Endocytosis: Actin in the
driving seat. Curr Biol. 14:R124–R126. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carlier MF and Pantaloni D: Control of
actin assembly dynamics in cell motility. J Biol Chem.
282:23005–23009. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pederson T and Aebi U: Actin in the
nucleus: what form and what for? J Struct Biol. 140:3–9. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Visa N and Percipalle P: Nuclear functions
of actin. Cold Spring Harb Perspect Biol. 2:a0006202010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Olave IA, Reck-Peterson SL and Crabtree
GR: Nuclear actin and actin-related proteins in chromatin
remodeling. Annu Rev Biochem. 71:755–781. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Blessing CA, Ugrinova GT and Goodson HV:
Actin and ARPs: action in the nucleus. Trends Cell Biol.
14:435–442. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hofmann WA, Stojiljkovic L, Fuchsova B, et
al: Actin is part of pre-initiation complexes and is necessary for
transcription by RNA polymerase II. Nat Cell Biol. 6:1094–1101.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Miralles F and Visa N: Actin in
transcription and transcription regulation. Curr Opin Cell Biol.
18:261–266. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen M and Shen X: Nuclear actin and
actin-related proteins in chromatin dynamics. Curr Opin Cell Biol.
19:326–330. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pederson T: As functional nuclear actin
comes into view, is it globular, filamentous, or both? J Cell Biol.
180:1061–1064. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gieni RS and Hendzel MJ: Actin dynamics
and functions in the interphase nucleus: moving toward an
understanding of nuclear polymeric actin. Biochem Cell Biol.
87:283–306. 2009.PubMed/NCBI
|
|
29
|
Xu YZ, Thuraisingam T, Morais DA,
Rola-Pleszczynski M and Radzioch D: Nuclear translocation of
β-actin is involved in transcriptional regulation during macrophage
differentiation of HL-60 cells. Mol Biol Cell. 21:811–820.
2010.
|
|
30
|
Kutsuna H, Suzuki K, Kamata N, et al:
Actin reorganization and morphological changes in human neutrophils
stimulated by TNF, GM-CSF and G-CSF: the role of MAP kinases. Am J
Physiol Cell Physiol. 286:55–64. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
El-Sonbaty SS, Watanabe M, Hochito K,
Yamaguchi K, Matsuda I and Tsuchiya H: Exogenously expressed
granulocyte colony-stimulating factor (G-CSF) receptor on K562
cells can transduce G-CSF-triggered growth and differentiation
signals. Int J Hematol. 61:61–68. 1995. View Article : Google Scholar
|
|
32
|
Saito K, Nakamura Y, Waga K, et al: Mature
and immature myeloid cells decrease the granulocyte
colony-stimulating factor level by absorption of granulocyte
colony-stimulating factor. Int J Hematol. 67:145–151. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Brandstetter T, Ninci E, Bettendorf H, et
al: Granulocyte colony stimulating factor (G-CSF) receptor gene
expression of ovarian carcinoma does not correlate with G-CSF
caused cell proliferation. Cancer. 91:1372–1383. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Veselska R, Zitterbart K, Jelinkova S,
Neradil J and Svoboda A: Specific cytoskeleton changes during
apoptosis accompanying induced differentiation of HL-60 myeloid
leukemia cells. Oncol Rep. 10:1049–1058. 2003.
|
|
35
|
Godyn JJ, Xu H, Zhang F, Kolla S and
Studzinski GP: A dual block to cell cycle progression in HL60 cells
exposed to analogues of vitamin D3. Cell Prolif. 27:37–46. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Erenpreisa J and Cragg MS: Mitotic death:
a mechanism of survival? Cancer Cell Int. 1:12001. View Article : Google Scholar
|
|
37
|
Zitterbart K and Veselska R: Effect of
retinoic acid on the actin cytoskeleton in HL-60 cells. Neoplasma.
48:456–461. 2001.PubMed/NCBI
|
|
38
|
Grzanka A, Grzanka D and Orlikowska M:
Cytoskeletal reorganization during process of apoptosis induced by
cytostatic drugs in K-562 and HL-60 leukemia cell lines. Biochem
Pharmacol. 66:1611–1617. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Atencia R, Garcia-Sanz M, Perez-Yarza G,
Asumendi A, Hilario E and Arechaga J: A structural analysis of
cytoskeleton components during the execution chase of apoptosis.
Protoplasma. 198:163–169. 1997. View Article : Google Scholar
|
|
40
|
Brown SB, Bailey K and Savill J: Actin is
cleaved during constitutive apoptosis. Biochem J. 323:233–237.
1997.PubMed/NCBI
|
|
41
|
Grzanka A, Izdebska M, Litwiniec A,
Grzanka D and Safiejko-Mroczka B: Actin filament reorganization in
HL-60 leukemia cell line after treatment with G-CSF and GM-CSF.
Folia Histochem Cytobiol. 45:191–197. 2007.PubMed/NCBI
|
|
42
|
Chodniewicz D and Zhelev DV: Novel pathway
of F-actin polymerization in the human neutrophil. Blood.
102:2251–2258. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gomez-Cambronero J, Horn J, Paul CC and
Baumann MA: Granulocyte-macrophage colony-stimulating factor is a
chemoattractant cytokine for human neutrophils: involvement of the
ribosomal p70 S6 kinase signaling pathway. J Immunol.
171:6846–6855. 2003. View Article : Google Scholar
|
|
44
|
Mailänder V and Landfester K: Interaction
of nanoparticles with cells. Biomacromolecules. 10:2379–2400.
2009.
|
|
45
|
Izdebska M, Grzanka D, Gackowska L, Żuryń
A and Grzanka A: The influence of Trisenox on action organization
in HL-60 cells. Cent Eur J Biol. 4:351–361. 2009. View Article : Google Scholar
|
|
46
|
Grzanka D, Marszałek A, Gagat M, Izdebska
M, Gackowska L and Grzanka A: Doxorubicin-induced F-actin
reorganization in cofilin-1 (nonmuscle) down-regulated CHO AA8
cells. Folia Histochem Cytobiol. 48:377–386. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Izdebska M, Grzanka A, Ostrowski M, Żuryń
A and Grzanka D: Effect of arsenic trioxide (Trisenox) on actin
organization in K-562 erythroleukemia cells. Folia Histochem
Cytobiol. 47:453–459. 2009.PubMed/NCBI
|
|
48
|
Zhao K, Wang W, Rando OJ, Xue Y, Swiderek
K, Kuo A and Crabtree GR: Rapid and phosphoinositol-dependent
binding of the SWI/SNF-like BAF complex to chromatin after T
lymphocyte receptor signaling. Cell. 95:625–636. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Widlak P, Palyvoda O, Kumala S and Garrard
WT: Modeling apoptotic chromatin condensation in normal cell
nuclei. Requirement for intranuclear mobility and actin
involvement. J Biol Chem. 277:21683–21690. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sjolinder M, Bjork P, Soderberg E, Sabri
N, Farrants AK and Visa N: The growing pre-mRNA recruits actin and
chromatin-modifying factors to transcriptionally active genes.
Genes Dev. 19:1871–1884. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhu QS, Xia L, Mills GB, Lowell CA, Touw
IP and Corey SJ: G-CSF induced reactive oxygen species involves
Lyn-PI3-kinase-Akt and contributes to myeloid cell growth. Blood.
107:1847–1856. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kitagawa J, Hara T, Tsurumi H, Kanemura N,
Kasahara S, Shimizu M and Moriwaki H: Cell cycle-dependent priming
action of granulocyte colony-stimulating factor (G-CSF) enhances in
vitro apoptosis induction by cytarabine and etoposide in leukemia
cell lines. J Clin Exp Hematop. 50:99–105. 2010. View Article : Google Scholar
|
|
53
|
Chow DC, Brevnova L, He XL, Martick MM,
Bankovich A and Garcia KC: A structural template for gp130-cytokine
signaling assemblies. Biochim Biophys Acta. 1592:225–235. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gearing DP, Thut CJ, VandeBos T, et al:
Leukemia inhibitory factor receptor is structurally related to the
IL-6 signal transducer, gp130. EMBO J. 10:2839–2848.
1991.PubMed/NCBI
|
|
55
|
Tartaglia LA, Dembski M, Weng X, et al:
Identification and expression cloning of a leptin receptor, OB-R.
Cell. 83:1263–1271. 1995. View Article : Google Scholar : PubMed/NCBI
|