Introduction
Gastric cancer is a major public health problem.
Despite a decrease in incidence in developed countries, it is the
fourth most common cancer worldwide and is the second leading cause
of cancer-related mortality (1,2).
Although it is curable if detected early, most gastric cancers are
asymptomatic until an advanced stage, when curative therapeutic
approaches are not optimal (3).
Therefore, to improve early diagnosis and develop new therapeutic
strategies in gastric cancer, a better understanding of the
molecular biology of gastric cancer development and progression is
required. A variety of molecules, including chemokines and their
receptors and growth factors, have been suggested to be responsible
for the metastatic propensity of gastric cancer cells (3–5).
However, the underlying molecular mechanisms that guide the
directional migration and metastasis of cancer cells towards target
tissues have yet to be established.
Chemokines and their receptors, particularly
chemokine (C-X-C motif) ligand 12 (CXCL12) and its receptor
chemokine (C-X-C motif) receptor 4 (CXCR4), have gained attention
for their functional roles in the development and maintenance of
the hematopoietic and immune systems, as well as in cancer
metastasis and progression (4,6–8). CXCR4
has been demonstrated to be upregulated in a variety of different
cancer cell lines and in human cancers including breast, prostate,
ovarian, lung, and gastric cancer. It directs the migration,
invasion and dissemination of tumor cells to specific sites that
are rich in CXCL12, including lymph nodes, bone, the lungs and body
cavities (9,10). The CXCR4/CXCL12 axis is also
involved in cancer cell proliferation at the primary or secondary
site, or both, suggesting it might be the key determinant of
overall disease progression. Indeed, in colorectal, breast, and
gastric cancer, high expression of CXCR4 is associated with more
aggressive tumor biology and poor survival outcome. CXCR4 is
therefore considered a potential therapeutic target, and CXCR4
antagonists and other inhibitors have been shown to reduce tumor
growth and metastasis in experimental models (9).
Given the critical role of CXCR4 in cancer
development and progression, microenvironmental factors that
modulate its levels may impact the process of tumor expansion.
Intratumoral hypoxia is a common feature of solid tumors and an
important microenvironmental factor that drives aggressive behavior
in cancer (9,11). After being exposed to hypoxia,
several types of cancer cells increase their synthesis of a protein
called hypoxia-inducible factor (HIF), which in turn binds to and
transactivates target genes (12).
HIF-1 is a heterodimeric transcription factor composed of a HIF-1α
subunit, which is rapidly degraded by ubiquitination and subsequent
passage through the proteasomal pathway, a process that is
inhibited under hypoxic conditions, and a HIF-1β subunit that is
constitutively expressed. Accumulating evidence suggests that
HIF-1α plays a role in the regulation of CXCR4 expression and
function. Moreover, the CXCR4 promoter contains four potential
hypoxia-response elements upstream of the transcriptional start
site, as well as one intra-intronic site (12–14).
CXCR4 is upregulated on monocytes and endothelial cells, as well as
tumor-associated macrophages and cancer cell lines, including
melanoma and breast cancer cell lines, under hypoxic conditions
(7,15–17).
However, the role of hypoxia in regulating CXCR4 in gastric cancer
remains poorly understood.
In the present study, we investigated the effect of
hypoxia on CXCR4 expression and biological activity in gastric
cancer cells and found that it promotes CXCR4 expression and
facilitates tumor cell migration and invasion by a HIF-1α-dependent
mechanism.
Materials and methods
Cell line and cultures
Human KATO III gastric cancer cell lines, purchased
from the Korean Cell Line Bank (Seoul, Korea), were used in this
study. Cells were cultured in RPMI-1640 medium (Gibco BRL Life
Technologies, Grand Island, NY, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco BRL Life Technologies), 100 U/ml
penicillin G, 100 μg/ml streptomycin (Sigma Aldrich, St. Louis, MO,
USA) and 1 mmol/l L-glutamine (Gibco BRL Life Technologies). Cells
were grown at 37°C in a humidified atmosphere containing 5%
CO2. For hypoxic exposure, cells were incubated with 5%
CO2 and 1% O2 (v/v), balanced with
N2 gas, at 37°C for the indicated periods of time.
Cell-surface expression of CXCR4
After being cultured under either normoxic or
hypoxic conditions for the indicated periods of time, cells were
incubated with phycoerythrin (PE)-conjugated monoclonal anti-CXCR4
(12G5; BD Pharmingen, San Diego, CA, USA) at 4°C for 30 min and
were analyzed using a Coulter Elite flow cytometer (Coulter
Electronics Ltd., Hialeah, FL, USA). A PE-conjugated mouse IgG
isotype-matched monoclonal antibody (dilution 1:50; Jackson
ImmunoResearch Laboratories, Inc., West Grove, PA, USA) was used as
a negative control.
Reagents
Human recombinant CXCL12/stromal cell derived
factor-1 (SDF-1) was purchased from R&D Systems, Inc.
(Minneapolis, MN, USA), and cobalt chloride (CoCl2) and
AMD3100 were purchased from Sigma-Aldrich.
shRNA targeting HIF-1α mRNA in gastric
cancer cells
Knockdown of HIF-1α in gastric cancer cells was
achieved through lentivirus-mediated transduction of HIF-1α
mRNA-specific shRNA using Mission RNAi system clones
(Sigma-Aldrich). To generate stable transfectants, lentiviral
vectors and packaging vectors were cotransfected into HEK293T cells
using Lipofectamine (Invitrogen, Carlsbad, CA, USA), according to
the manufacturer’s instructions. The following day, virus harvested
from the supernatant was added to KATO III cells together with 10
μg/ml polybrene. After 24 h, the medium was removed and replaced
with fresh medium containing 8 μg/ml puromycin. Puromycin-resistant
clones were selected after culture for 1 week in the presence of
puromycin. HIF-1α expression levels were analyzed by western
blotting.
Wound healing, migration and invasion
assays
Cells were grown to confluence in 6-well culture
plates. A wound was created with a sterile pipette tip at an angle
of ~30°. After washing, the culture medium was replaced. Cell
migration was monitored with a microscope for 24–48 h. For
Transwell migration and invasion experiments, cells
(4×105 per well) were loaded into the upper chamber of a
24-well Transwell plate containing an 8-μm microporous membrane
(Corning-Costar, Cambridge, MA, USA) and were allowed to migrate
into the lower chamber, which contained CXCL12 (200 ng/ml) or
AMD3100 (10 μM/ml) or medium alone (RPMI-1640 supplemented with 10%
FBS) at 37°C in normoxic or hypoxic conditions for 24 h (migration)
or 48 h (invasion). For the migration assay, the lower surface of
the filter was coated with 10 μg of gelatin; for the invasion
assay, the upper side was coated with 12 μg of reconstituted
basement membrane substance (Matrigel; BD Biosciences, Bedford, MA,
USA). Cells were fixed and stained with hematoxylin and eosin.
Non-migrating cells on the upper surface of the filter were removed
by wiping with a cotton swab. Chemotaxis was quantified by counting
the cells that migrated to the lower side of the filter under an
optical microscope. Six random fields were counted for each
assay.
Western blot analysis
Western blotting was used to detect protein
molecules. Cells were starved in serum-free medium for 12 h and
then stimulated with cytokines or CXCR4 antagonists. The cells were
collected by centrifugation, washed in phosphate-buffered saline,
and lysed by the addition of RIPA buffer (5 M NaCl, NP-40, sodium
deoxycholate, 1 M Tris-HCl (pH 7.4), 0.5 M EDTA (pH 8.0). Equal
amounts of protein from each sample were separated by
electrophoresis on 10% SDS-polyacrylamide gels and transferred to
polyvinylidene fluoride membranes (Amersham Life Science, Arlington
Heights, IL, USA). The membranes were blocked for 1 h in
Tris-buffered saline (TBS) containing 5% (w/v) milk and 0.1%
Tween-20, and were then incubated overnight at 4°C with primary
mouse or rabbit monoclonal antibody (Cell Signaling Technology
Inc., Danvers, MA, USA). The blots were washed with TBS containing
Tween-20, incubated with anti-mouse or anti-rabbit secondary
antibody for 2 h, and developed using West-Zol Plus (iNtRON
Biotechnology, Seoul, Korea). The following antibodies were used:
anti-CXCR4 monoclonal antibody (12G5; Thermo Scientific, Rockford,
IL, USA) and anti-HIF-1α polyclonal antibody (BD Biosciences).
Statistical analysis
Results are expressed as the means ± standard
deviation (SD) of at least three experiments. Data were analyzed
using Student’s t-test (for paired samples). P<0.05 was
considered to indicate statistically significant differences.
Results
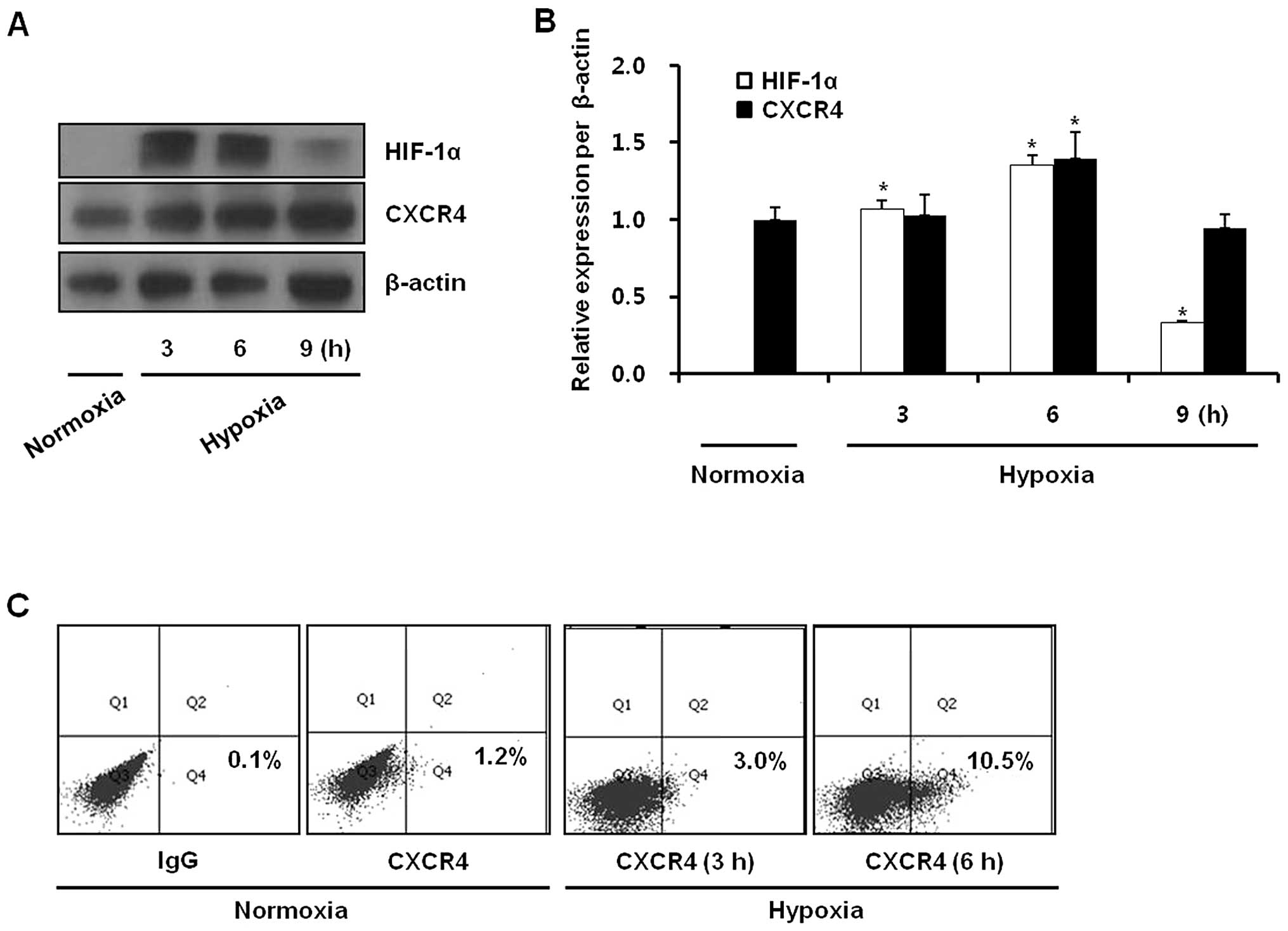
Hypoxia upregulates CXCR4 protein levels
and cell surface expression
To investigate the effect of hypoxia on CXCR4
expression in gastric cancer cells, KATO III cells were exposed to
either normoxia (21% O2) or hypoxia (1% O2)
for 3, 6 or 9 h and the expression of HIF-1α and CXCR4 proteins was
analyzed by western blotting. Hypoxia upregulated CXCR4 and HIF-1α
protein levels. CXCR4 protein levels were increased at 6 h and then
showed a decrease that corresponded to a change in HIF-1α protein
levels (Fig. 1A and B). Hypoxia
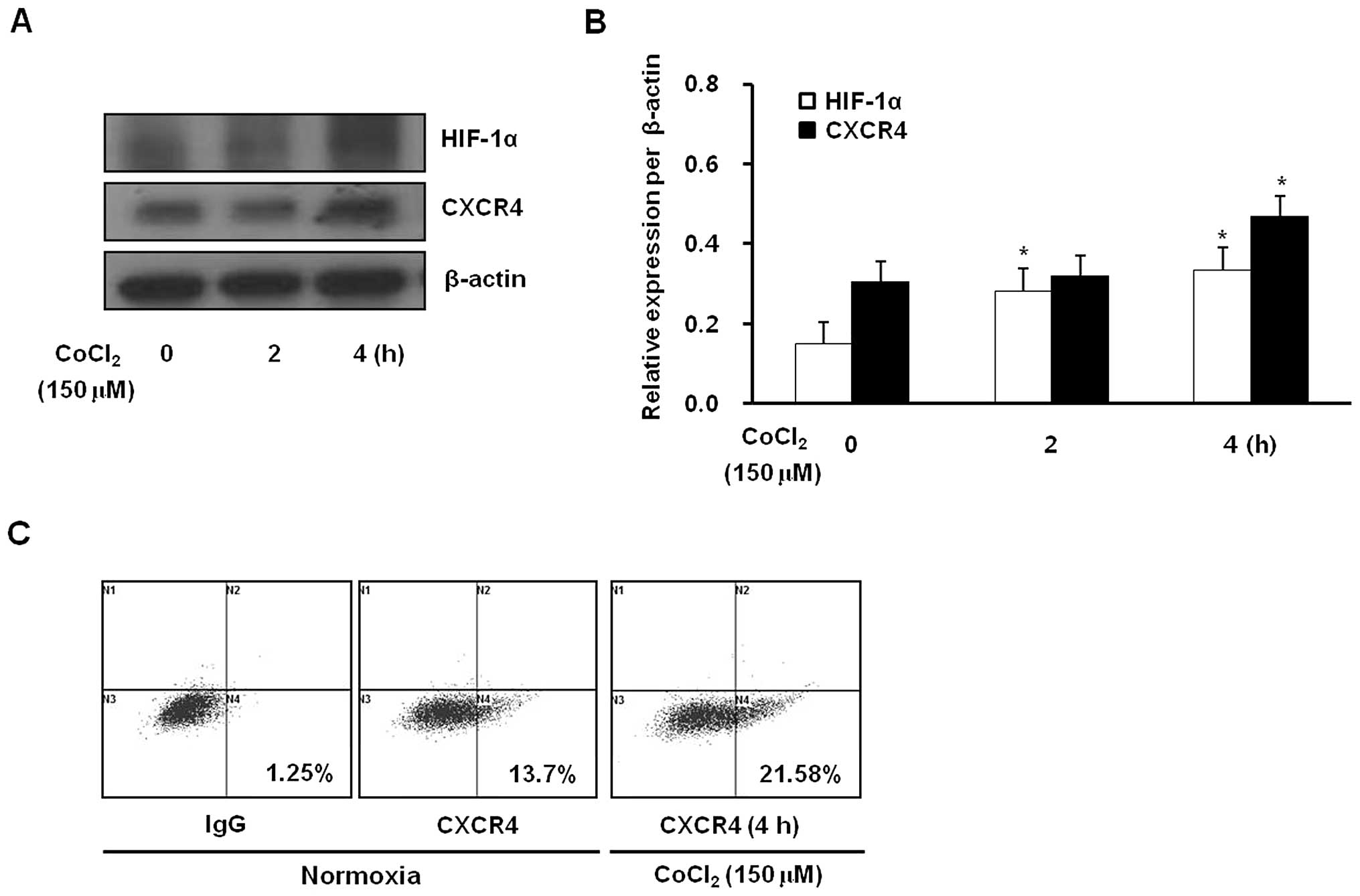
also increased cell surface expression of CXCR4 (Fig. 1C). In addition, the hypoxia-mimetic
agent CoCl2 increased CXCR4 protein levels and surface
expression, responses that were accompanied by an increase in
HIF-1α levels (Fig. 2). These
observations were consistent with the above hypoxic stimulation
results. These data provide evidence that CXCR4 expression in
gastric cancer cells is upregulated by hypoxia.
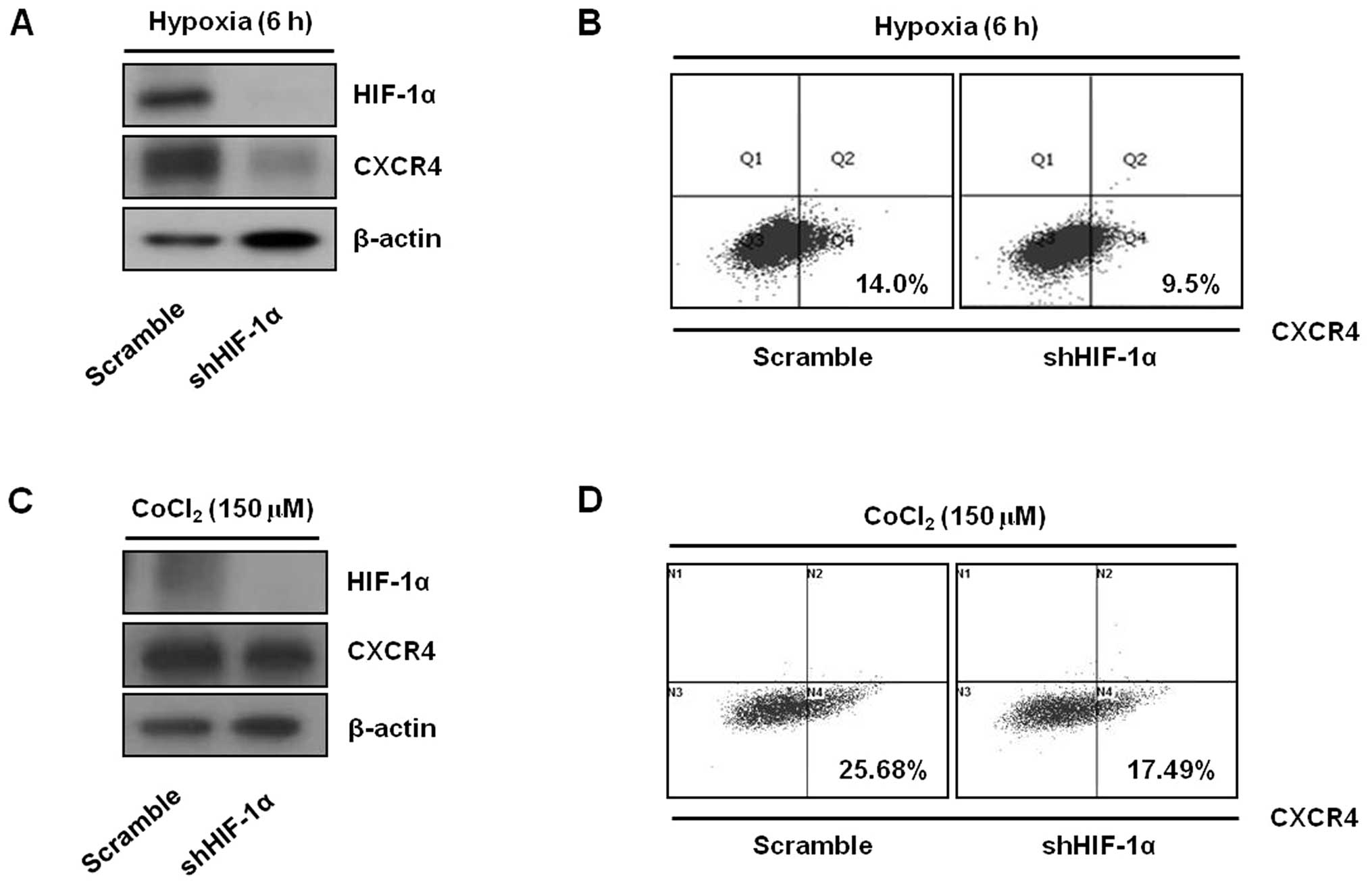
HIF-1α is involved in the induction of
CXCR4 expression by hypoxia
To further explore whether HIF-1α was involved in
the upregulation of CXCR4 expression by hypoxia, we generated KATO
III cell clones with stable knockdown of HIF-1α. Specific depletion
of HIF-1α in these cells was accomplished by lentivirus-mediated
transduction and expression of a HIF-1α mRNA-specific shRNA. We
effectively inhibited HIF-1α expression using this system (Fig. 3A and C). When exposed to hypoxic
conditions, cells transfected with a lentivirus containing scramble
sequences showed upregulation of CXCR4 protein levels and surface
expression, as expected (Fig. 3A and
B). By contrast, hypoxia-induced CXCR4 upregulation under
hypoxic conditions was blocked in cells transfected with shRNA
directed against HIF-1α (Fig. 3A and
B). Similarly, CoCl2-induced upregulation of CXCR4
protein levels and surface expression was inhibited by abrogation
of HIF-1α (Fig. 3C and D).
Collectively, these data suggest that the induction of CXCR4
expression by hypoxia is mediated by HIF-1α.
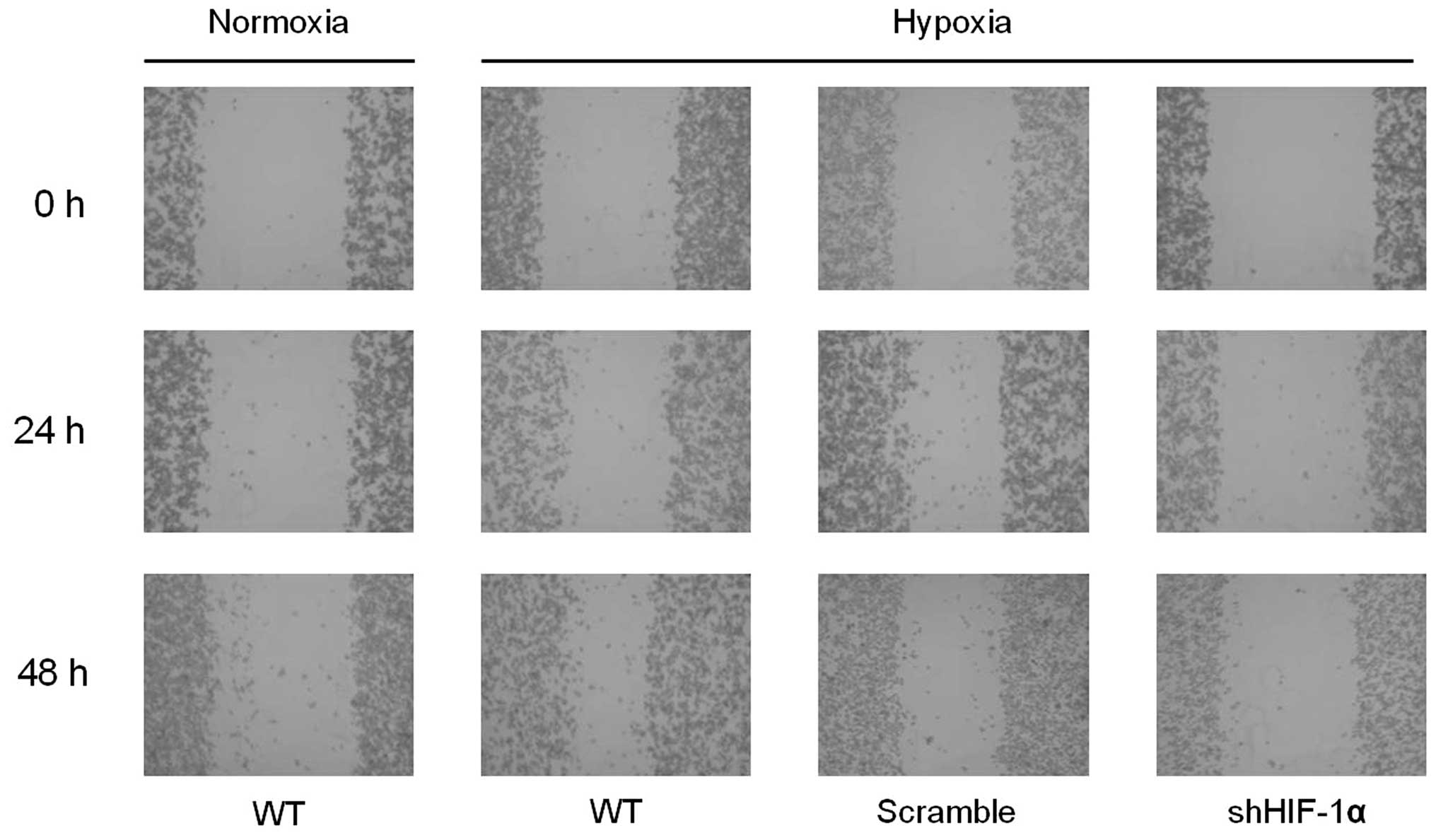
Hypoxia promotes the migration and
invasion of gastric cancer cells
To examine the effect of hypoxia on the metastatic
ability of gastric cancer cells, we performed wound healing and
Transwell migration and invasion assays. The ability to heal wounds
was assessed in cells exposed to normoxia and hypoxia. Hypoxia was
shown to facilitate wound healing in gastric cancer cells. However,
the facilitation of wound healing by hypoxia was abrogated by
knockdown of HIF-1α (Fig. 4). When
exposed to hypoxia for 24 h, KATO III cells showed significantly
increased migration in response to CXCL12 compared to cells exposed
to normoxia. These effects were blocked by treatment with AMD3100
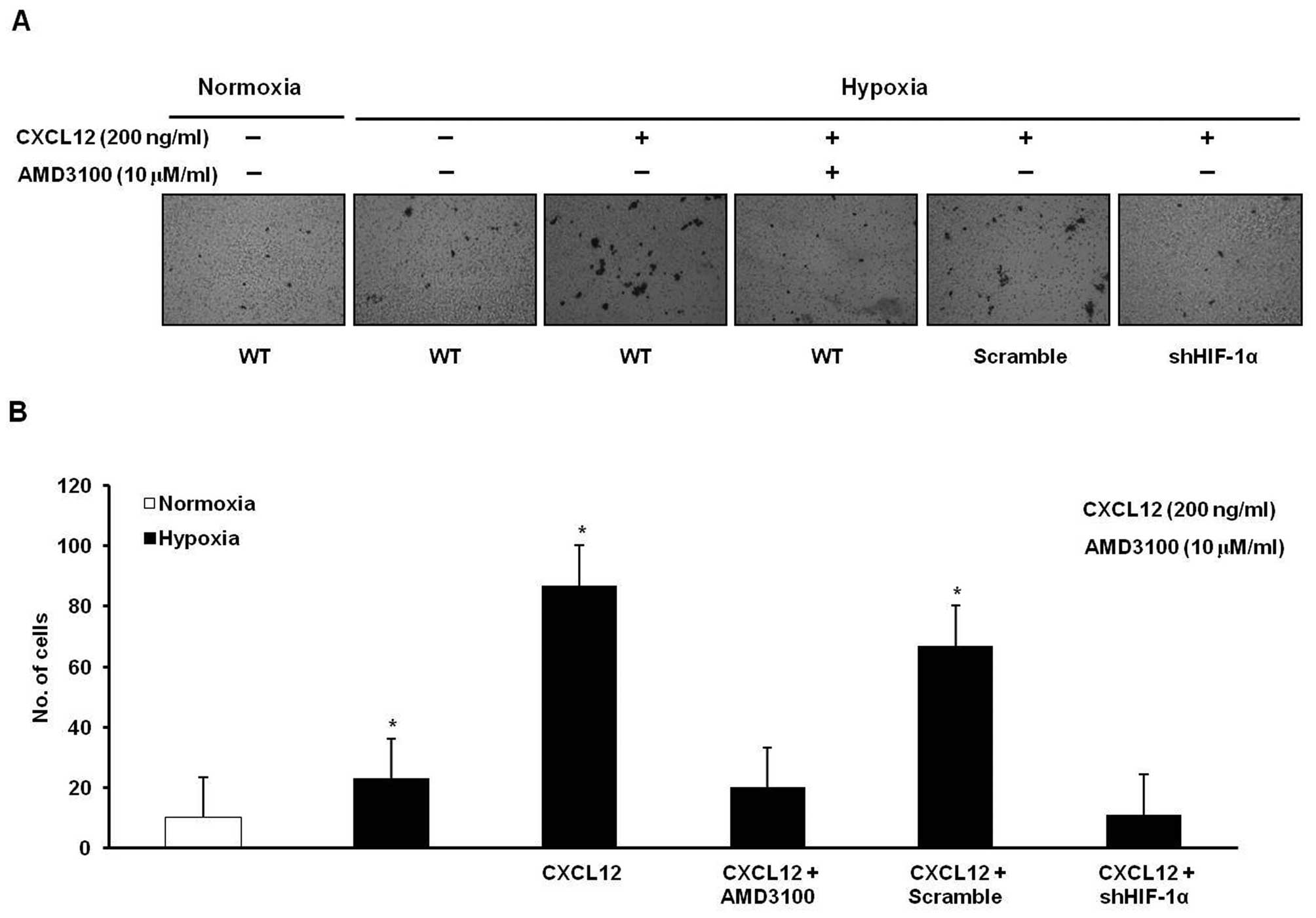
or knockdown of HIF-1α (Fig. 5).
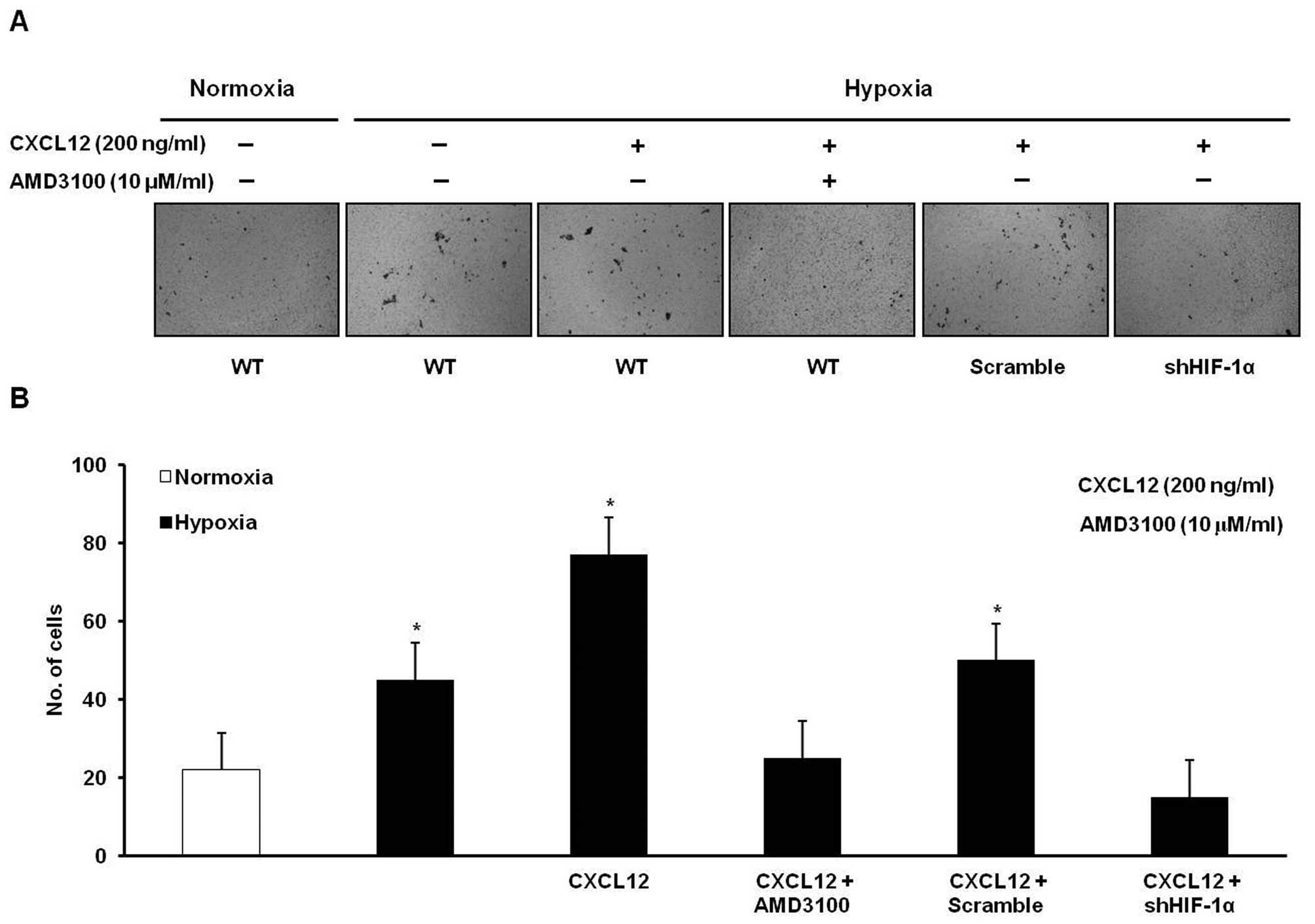
Invasion analysis similarly showed that a hypoxia-induced increase
in invasion in cells stimulated with CXCL12 was blocked by
treatment with AMD3100 or knockdown of HIF-1α (Fig. 6). These results suggest that hypoxia
may induce gastric cancer cell migration and invasion in a HIF-1α-
and CXCR4-dependent manner.
Discussion
To the best of our knowledge, the present study is
the first to provide evidence that hypoxia upregulates CXCR4
protein levels and cell membrane expression in gastric cancer
cells, responses that were dependent on activation of HIF-1α and
implicated in increased tumor cell migration and invasion.
Chemokines are 8- to 10-kDa chemoattractant
cytokines that not only control leukocyte trafficking, but also
play critical roles in the migration and metastasis of cancer cells
(originating from primary tumors) with corresponding chemokine
receptors to certain organs. Notably, the chemokine CXCL12
activates its receptor, CXCR4, which is involved in cancer cell
migration and invasion, and thus promotes organ-specific
localization of distant metastases from various carcinomas
(18). In gastric cancer,
accumulating evidence suggests that CXCR4 protein levels and cell
membrane expression in vitro and in vivo are altered
(4,19–22).
The differential expression of CXCR4 in gastric cancer cells was
also shown by gene expression profiling (23). Overexpression of CXCR4 in gastric
cancer cells was associated with the development of peritoneal
carcinomatosis, which is induced by dissemination of cancer cells
into the peritoneal cavity (20,24).
Numerous studies have demonstrated that strong expression of CXCR4
is correlated with aggressive tumor characteristics such as deep
invasion, lymph node, and liver metastasis (4,20,25–28).
Furthermore, several pre-clinical investigations demonstrated that
blocking of CXCR4 signaling reduced the size and number of tumor
metastases (20,29,30).
Therefore, a better understanding of the mechanism by which CXCR4
expression and biological activity are regulated in gastric cancer
is important for the development of improved treatment
strategies.
In contrast to normal organs, oxygen homeostasis in
solid tumors is deregulated and the pO2 decreases to
very low levels as the tumors grow (15). Although hypoxia is cytotoxic to both
normal and cancer cells, some cancer cells acquire characteristics
that allow them to survive and grow under hypoxic conditions (such
as, expression of angiogenic factors, glycolytic enzymes and stress
proteins). In addition, these ischemic conditions contribute to the
aggressive metastatic phenotypes of tumor cells, and metastasis,
which is characterized by migration, seeding and growth of
satellite lesions in specific organs, is commonly considered to be
the final stage of cancer (11,31–34).
Experimental and clinical data provided evidence for a relationship
between intratumoral hypoxia and aggressive behavior in solid
tumors, including gastric cancer (35–39).
Several recent studies provide evidence that hypoxia, primarily
acting through HIF-1α, is involved in the upregulation of CXCR4 in
carcinomas and hematologic malignancies such as lung, breast cancer
and myeloma, with HIF-1α binding directly to the CXCR4 promoter
(15,40–45).
In this study, hypoxia increased CXCR4 protein levels and cell
surface expression by activating HIF-1α, thereby supporting
previous findings from other cancer cell lines and suggesting that
tumor hypoxia plays a crucial role in the regulation of CXCR4
(15,16,41).
Next, we investigated whether hypoxia-induced
upregulation of CXCR4 in gastric cancer cells was related to
increased metastatic potential. Gastric cancer cells exposed to
hypoxia showed significantly increased migration and invasion in
response to CXCL12 treatment compared with those exposed to
normoxia. The increase in migratory ability due to hypoxia was
abrogated by knockdown of HIF-1α and inhibition of CXCR4. These
results are in accordance with previous reports in different types
of cancer (16,46) and suggest that upregulation of CXCR4
by HIF-1α in response to hypoxia plays a biological role in gastric
cancer cell migration. We previously reported that functional CXCR4
plays an important role in cancer cell migration during normoxia in
gastric and gallbladder cancers (18,21),
suggesting that the CXCR4/CXCL12 axis may be significantly involved
in cancer progression and may be a potential target for
therapeutics that block the interaction between CXCL12 and CXCR4,
or that inhibit downstream signaling, in the treatment of cancer.
To date, several antagonists have been used to target CXCR4,
including AMD3100, T22 and ALX40-4C (10). Among them, AMD3100 (trade name
Plerixafor) was shown to be well-tolerated and has been approved by
the Food and Drug Administration of the United States for use in
non-Hodgkin’s lymphoma and multiple myeloma to mobilize stem cells
for collection prior to autologous transplantation (47). AMD3100 is currently being tested
against hematologic malignancies such as acute myeloid leukemia in
early clinical trials in combination with chemotherapy with the
rationale that disrupting the interaction between leukemic cells
and the bone marrow microenvironment will increase the cytotoxic
effect of chemotherapy (48).
In gastric cancer, several pre-clinical
investigations demonstrated that monoclonal antibodies and specific
low-molecular weight antagonists of CXCR4 showed antitumor activity
in vitro and in vivo. CXCL12-induced cancer cell
migration was significantly reduced by treatment with a
neutralizing anti-CXCR4 antibody or AMD3100. AMD3100 reduced tumor
growth and malignant ascites formation in a xenograft model
(20,21,29,49).
Taken together, these findings suggest that hypoxia may increase
CXCR4 activity in gastric cancer cells by activating HIF-1α,
thereby resulting in increased migratory ability. This suggests
that the hypoxia-HIF-1α-CXCR4 axis might be a potential therapeutic
target.
In conclusion, hypoxia upregulates CXCR4 protein
levels and cell membrane expression in gastric cancer cells in a
HIF-1α-dependent manner. The upregulation of CXCR4 plays roles in
cancer cell migration and invasion, which were reduced by knockdown
of HIF-1α or inhibition of CXCR4. Our results collectively suggest
that disruption of the hypoxia-HIF-1α-CXCR4 axis is a potential
therapeutic strategy for the treatment of gastric cancer.
Acknowledgements
This study was supported in part by the Basic
Science Research Program through the National Research Foundation
of Korea (NRF) funded by the Ministry of Education, Science and
Technology (KRF-2009-0076540, KRF-2009-0067256).
References
|
1
|
Kamangar F, Dores GM and Anderson WF:
Patterns of cancer incidence, mortality, and prevalence across five
continents: defining priorities to reduce cancer disparities in
different geographic regions of the world. J Clin Oncol.
24:2137–2150. 2006. View Article : Google Scholar
|
|
2
|
Lee HJ, Cho do Y, Park JC, et al: Phase II
trial of biweekly paclitaxel plus infusional 5-fluorouracil and
leucovorin in patients with advanced or recurrent inoperable
gastric cancer. Cancer Chemother Pharmacol. 63:427–432. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cheng WL, Wang CS, Huang YH, Tsai MM,
Liang Y and Lin KH: Overexpression of CXCL1 and its receptor CXCR2
promote tumor invasion in gastric cancer. Ann Oncol. 22:2267–2276.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee HJ, Huang SM, Kim HY, et al:
Evaluation of the combined expression of chemokine SDF-1 alpha and
its receptor CXCR4 as a prognostic marker for gastric cancer. Exp
Ther Med. 2:499–504. 2011.PubMed/NCBI
|
|
5
|
Shen Z, Seppanen H, Vainionpaa S, et al:
IL10, IL11, IL18 are differently expressed in CD14(+) TAMs and play
different role in regulating the invasion of gastric cancer cells
under hypoxia. Cytokine. 59:352–357. 2012.PubMed/NCBI
|
|
6
|
Song IC, Liang ZL, Lee JC, et al:
Expression of stromal cell-derived factor-1α is an independent risk
factor for lymph node metastasis in early gastric cancer. Oncol
Lett. 2:1197–1202. 2011.
|
|
7
|
Fiegl M, Samudio I, Clise-Dwyer K, Burks
JK, Mnjoyan Z and Andreeff M: CXCR4 expression and biologic
activity in acute myeloid leukemia are dependent on oxygen partial
pressure. Blood. 113:1504–1512. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Muller A, Homey B, Soto H, et al:
Involvement of chemokine receptors in breast cancer metastasis.
Nature. 410:50–56. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Richard CL, Tan EY and Blay J: Adenosine
upregulates CXCR4 and enhances the proliferative and migratory
responses of human carcinoma cells to CXCL12/SDF-1alpha. Int J
Cancer. 119:2044–2053. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun X, Cheng G, Hao M, et al:
CXCL12/CXCR4/CXCR7 chemokine axis and cancer progression. Cancer
Metastasis Rev. 29:709–722. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Li Z, Zhang H, et al: HIF-1alpha
and HIF-2alpha correlate with migration and invasion in gastric
cancer. Cancer Biol Ther. 10:376–382. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang X, Li C, Chen Y, et al: Hypoxia
enhances CXCR4 expression favoring microglia migration via
HIF-1alpha activation. Biochem Biophys Res Commun. 371:283–288.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schioppa T, Uranchimeg B, Saccani A, et
al: Regulation of the chemokine receptor CXCR4 by hypoxia. J Exp
Med. 198:1391–1402. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Krishnamachary B, Berg-Dixon S, Kelly B,
et al: Regulation of colon carcinoma cell invasion by
hypoxia-inducible factor 1. Cancer Res. 63:1138–1143.
2003.PubMed/NCBI
|
|
15
|
Ishikawa T, Nakashiro K, Klosek SK, et al:
Hypoxia enhances CXCR4 expression by activating HIF-1 in oral
squamous cell carcinoma. Oncol Rep. 21:707–712. 2009.PubMed/NCBI
|
|
16
|
Cronin PA, Wang JH and Redmond HP: Hypoxia
increases the metastatic ability of breast cancer cells via
upregulation of CXCR4. BMC Cancer. 10:2252010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schutyser E, Su Y, Yu Y, et al: Hypoxia
enhances CXCR4 expression in human microvascular endothelial cells
and human melanoma cells. Eur Cytokine Netw. 18:59–70.
2007.PubMed/NCBI
|
|
18
|
Lee HJ, Lee K, Lee DG, et al: Chemokine
(C-X-C motif) ligand 12 is associated with gallbladder carcinoma
progression and is a novel independent poor prognostic factor. Clin
Cancer Res. 18:3270–3280. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kwak MK, Hur K, Park DJ, et al: Expression
of chemokine receptors in human gastric cancer. Tumour Biol.
26:65–70. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yasumoto K, Koizumi K, Kawashima A, et al:
Role of the CXCL12/CXCR4 axis in peritoneal carcinomatosis of
gastric cancer. Cancer Res. 66:2181–2187. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee HJ, Kim SW, Kim HY, et al: Chemokine
receptor CXCR4 expression, function, and clinical implications in
gastric cancer. Int J Oncol. 34:473–480. 2009.PubMed/NCBI
|
|
22
|
Pituch-Noworolska A, Drabik G, Szatanek R,
et al: Immunophenotype of isolated tumour cells in the blood, bone
marrow and lymph nodes of patients with gastric cancer. Pol J
Pathol. 58:93–97. 2007.PubMed/NCBI
|
|
23
|
Sun XJ, Sun KL, Zheng ZH, et al: Gene
expression patterns in gastric cancer. Zhonghua Yi Xue Yi Chuan Xue
Za Zhi. 23:142–146. 2006.PubMed/NCBI
|
|
24
|
Hashimoto I, Koizumi K, Tatematsu M, et
al: Blocking on the CXCR4/mTOR signalling pathway induces the
anti-metastatic properties and autophagic cell death in peritoneal
disseminated gastric cancer cells. Eur J Cancer. 44:1022–1029.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tsuboi K, Kodera Y, Nakanishi H, et al:
Expression of CXCL12 and CXCR4 in pT3-stage gastric cancer does not
correlate with peritoneal metastasis. Oncol Rep. 20:1117–1123.
2008.PubMed/NCBI
|
|
26
|
Iwasa S, Yanagawa T, Fan J and Katoh R:
Expression of CXCR4 and its ligand SDF-1 in intestinal-type gastric
cancer is associated with lymph node and liver metastasis.
Anticancer Res. 29:4751–4758. 2009.PubMed/NCBI
|
|
27
|
Arigami T, Natsugoe S, Uenosono Y, et al:
CCR7 and CXCR4 expression predicts lymph node status including
micrometastasis in gastric cancer. Int J Oncol. 35:19–24. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao BC, Wang ZJ, Mao WZ, et al:
CXCR4/SDF-1 axis is involved in lymph node metastasis of gastric
carcinoma. World J Gastroenterol. 17:2389–2396. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ding YL, Zhang JL, Tang SF, Fu QY and Li
ZT: Effect of chemokine stromal cell derived factor-1 and its
receptor CXCR4 on the peritoneal carcinometastasis of gastric
cancer. Zhonghua Yi Xue Za Zhi. 88:202–205. 2008.(In Chinese).
|
|
30
|
Koizumi K, Kato S, Sakurai H, Hashimoto I,
Yasumoto K and Saiki I: Therapeutics target of CXCR4 and its
downstream in peritoneal carcinomatosis of gastric cancer. Front
Biosci (Schol Ed). 4:269–276. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Noda S, Yashiro M, Nshii T and Hirakawa K:
Hypoxia upregulates adhesion ability to peritoneum through a
transforming growth factor-beta-dependent mechanism in diffuse-type
gastric cancer cells. Eur J Cancer. 46:995–1005. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hockel M, Schlenger K, Hockel S and Vaupel
P: Hypoxic cervical cancers with low apoptotic index are highly
aggressive. Cancer Res. 59:4525–4528. 1999.PubMed/NCBI
|
|
33
|
Zhong H, De Marzo AM, Laughner E, et al:
Overexpression of hypoxia-inducible factor 1alpha in common human
cancers and their metastases. Cancer Res. 59:5830–5835.
1999.PubMed/NCBI
|
|
34
|
Hynes RO: Metastatic potential: generic
predisposition of the primary tumor or rare, metastatic variants-or
both? Cell. 113:821–823. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Griffiths EA, Pritchard SA, Welch IM,
Price PM and West CM: Is the hypoxia-inducible factor pathway
important in gastric cancer? Eur J Cancer. 41:2792–2805. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hockel M, Schlenger K, Aral B, Mitze M,
Schaffer U and Vaupel P: Association between tumor hypoxia and
malignant progression in advanced cancer of the uterine cervix.
Cancer Res. 56:4509–4515. 1996.PubMed/NCBI
|
|
37
|
Generali D, Berruti A, Brizzi MP, et al:
Hypoxia-inducible factor-1alpha expression predicts a poor response
to primary chemoendocrine therapy and disease-free survival in
primary human breast cancer. Clin Cancer Res. 12:4562–4568. 2006.
View Article : Google Scholar
|
|
38
|
Bachtiary B, Schindl M, Potter R, et al:
Overexpression of hypoxia-inducible factor 1alpha indicates
diminished response to radiotherapy and unfavorable prognosis in
patients receiving radical radiotherapy for cervical cancer. Clin
Cancer Res. 9:2234–2240. 2003.
|
|
39
|
Schindl M, Schoppmann SF, Samonigg H, et
al: Overexpression of hypoxia-inducible factor 1alpha is associated
with an unfavorable prognosis in lymph node-positive breast cancer.
Clin Cancer Res. 8:1831–1837. 2002.PubMed/NCBI
|
|
40
|
Pan J, Mestas J, Burdick MD, et al:
Stromal derived factor-1 (SDF-1/CXCL12) and CXCR4 in renal cell
carcinoma metastasis. Mol Cancer. 5:562006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Phillips RJ, Mestas J, Gharaee-Kermani M,
et al: Epidermal growth factor and hypoxia-induced expression of
CXC chemokine receptor 4 on non-small cell lung cancer cells is
regulated by the phosphatidylinositol 3-kinase/PTEN/AKT/mammalian
target of rapamycin signaling pathway and activation of hypoxia
inducible factor-1alpha. J Biol Chem. 280:22473–22481. 2005.
|
|
42
|
Shim H, Lau SK, Devi S, Yoon Y, Cho HT and
Liang Z: Lower expression of CXCR4 in lymph node metastases than in
primary breast cancers: potential regulation by ligand-dependent
degradation and HIF-1alpha. Biochem Biophys Res Commun.
346:252–258. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim SW, Kim HY, Lee HJ, Yun HJ, Kim S and
Jo DY: Dexamethasone and hypoxia upregulate CXCR4 expression in
myeloma cells. Leuk Lymphoma. 50:1163–1173. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sun X, Wei L, Chen Q and Terek RM:
CXCR4/SDF1 mediate hypoxia induced chondrosarcoma cell invasion
through ERK signaling and increased MMP1 expression. Mol Cancer.
9:172010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dunn LK, Mohammad KS, Fournier PG, et al:
Hypoxia and TGF-beta drive breast cancer bone metastases through
parallel signaling pathways in tumor cells and the bone
microenvironment. PloS One. 4:e68962009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zagzag D, Lukyanov Y, Lan L, et al:
Hypoxia-inducible factor 1 and VEGF upregulate CXCR4 in
glioblastoma: implications for angiogenesis and glioma cell
invasion. Lab Invest. 86:1221–1232. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mahaseth H and Kaufman J: Optimizing stem
cell collection through CXCR4 antagonists. Front Biosci (Schol Ed).
4:611–619. 2012.PubMed/NCBI
|
|
48
|
Gangadhar T, Nandi S and Salgia R: The
role of chemokine receptor CXCR4 in lung cancer. Cancer Biol Ther.
9:409–416. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Iwanaga T, Iwasaki Y, Ohashi M, Nunobe S
and Iwagami S: Establishment of a CXCR4-expressing gastric cancer
cell line in nude mice and the effect of AMD 3100 on tumor
regression. Gan To Sagaku Ryoho. 34:1917–1919. 2007.(In
Japanese).
|