Introduction
The incidence and mortality of esophageal cancer
rank high on the global cancer list, particularly in developing
countries (1). Esophageal squamous
cell carcinoma (ESC) is one of the most common cancers in China.
Due to the lack of early symptoms, rapid progression, ineffective
chemotherapy and frequent recurrence after surgery, ESC is
characterized by high mortality rate with a 5-year survival rate of
less than 10% (2). Although
multi-modality therapy and neoadjuvant therapy have been employed,
the survival rate of ESC patients has not improved for many years.
The molecular events underlying esophageal cancer development
remain undefined. Therefore, there is an urgent need to understand
the factors associated with prognosis, therapy and drug resistance
of ESCs.
Cell cycle progression is tightly regulated by
checkpoint systems which play an essential role in maintaining DNA
replication and genome integrity. During mitosis, the mitotic
checkpoint complex exerts strict surveillance on the fidelity of
the mitotic process by sensing the proper tension between
kinetochores of paired chromatids and the attachment of bipolar
microtubules to kinetochores (3).
BubR1, one of the mitotic checkpoint complex proteins, plays dual
roles in sensing unaligned chromatids at the kinetochore and in
suspending E3 ubiquitin ligase function in the cytoplasm (4,5). In
this way, the mitotic spindle checkpoint ensures that
sister-chromatids can be evenly distributed into daughter cells to
sustain their genetic integrity. Otherwise, BubR1 is activated and
hyperphosphorylated to arrest the mitotic process by inhibiting
metaphase-anaphase transition. Consequently, prolonged mitotic
arrested cells exit mitosis and undergo post-mitotic apoptosis.
Abnormal expression of mitotic checkpoint proteins, such as Mad2
and Bub1, has been widely reported to be related to the chromosomal
instability of cells and tumor predisposition (6,7).
Overexpression of BubR1 has been reported in several types of
malignancies, including breast, bladder, gastric, ovarian, kidney
and esophageal squamous cell cancers (8–13),
while mutations of the Bub1b gene are rarely observed
(14). Depletion of BubR1 protein
or loss of function can cause spindle checkpoint failure and
further disturb the normal mitotic process, which finally results
in chromosomal instability and aneuploidy (11,15).
Furthermore, haplo-insufficiency of BubR1 in mice can trigger
tumorigenesis and senescence (16,17).
Therefore, it is likely that abnormal expression of BubR1 may be
involved in tumor progression and chemotherapeutic
responsiveness.
Paclitaxel is a potent chemotherapeutic agent
approved to treat ovarian, breast and lung cancers. As an adjuvant
chemotherapy drug, paclitaxel has recently been used to potentiate
the efficacy of 5-fluorouracil and cisplatin in esophageal cancer
therapy. Several studies have shown that the cytotoxic effects of
paclitaxel depend on the proper and timely activation of mitotic
checkpoints (18–20). Defective mitotic spindle checkpoint
by gradually reducing BubR1 in SKOV3-TR30 cells was found to lead
to paclitaxel resistance through decreasing the mitotic index and
suppressing cell death (21).
However, the depletion or inactivation of BubR1 in several cancer
cell lines somehow leads to an increase in their sensitivity to
paclitaxel (22). Thus, it remains
unclear how the expression status of BubR1 in cancer cells would
affect the development of paclitaxel resistance. Nonetheless, it
was reported that esophageal squamous cell cancer TE1 cells with
relatively low BubR1 expression show more sensitivity to paclitaxel
than TE2 cells (13).
This study was designed to thoroughly evaluate the
role of mitotic checkpoint protein BubR1 in the development of
paclitaxel chemoresistance in esophageal cancer cells. Using
quantitative real-time PCR analysis on 50 samples of paired
esophageal squamous cell cancer (ESC) tissues and adjacent
non-cancerous tissues, we found that 72% (36 of 50) of the analyzed
ESC samples exhibited a high expression level of BubR1, which was
also confirmed in ESC cell lines. ESC cells with a high level of
BubR1 expression were less sensitive to the anti-microtubule drugs
paclitaxel and nocodazole. Recombinant adenovirus-mediated enforced
expression of BubR1 in relatively sensitive ESC cell lines resulted
in increased resistance to paclitaxel. Conversely, RNAi-mediated
knockdown of BubR1 restored the ESC cell sensitivity to paclitaxel.
Cell cycle analysis indicated that the sub-G1 cell population
increased in the ESC cells having a reduced BubR1 level. Our
results suggest that upregulation of BubR1 expression may be
associated with ESC resistance to paclitaxel treatment. Thus, BubR1
may serve as a potential chemosensitizing target to overcome
chemoresistance in ESC.
Materials and methods
Esophageal cancer samples
The use of clinical esophageal cancer samples was
approved by the Institutional Ethics Committees of Chongqing
Medical University (Chongqing, China) and Chuanbei Medical College
(Sichuan, China). The patients consented to the use of the
surgically resected samples for this study. Tissue specimens (n=50)
from freshly resected esophageal squamous cell cancer and adjacent
non-cancerous tissues were obtained at the Affiliated Hospital of
Chuanbei Medical College. Samples were maintained in liquid
nitrogen and used for RNA extraction using TRIzol agent
(Invitrogen).
Cell culture and chemicals
ESC cell lines ECA-109, KYSE150 and KYSE180 were
cultured in RPMI-1640 medium (Hyclone) supplemented with 10% fetal
bovine serum (FBS, Hyclone), and the breast cancer cell line MCF-7
was cultured in DMEM (Hyclone) in 10% FBS. All cells were
maintained in 1% penicillin/streptomycin in 5% CO2 at
37°C. Paclitaxel and nocodazole were purchased from Sangon Biotech
(Shanghai, China).
Recombinant adenovirus construction and
amplifications
Recombinant adenovirus-expressing hBubR1 was
generated as previously described (23). Briefly, the coding region for BubR1
was amplified by PCR and subcloned into a shuttle vector for
recombinant adenovirus generation using the AdEasy system (24). Then, recombinant adenoviruses were
packaged and amplified in HEK-293 cells. High titer of adenovirus
expression of both GFP and BubR1 was obtained, namely AdBubR1,
while adenovirus only expressing GFP (i.e., AdGFP) was used as a
negative control.
RNA interference
Oligo cassettes for short hairpin RNA (shRNA)
targeting BubR1 mRNA (NM_001211.4) were synthesized by
GenePharma (Shanghai, China) and cloned into the pU6/GFP/Neo
vector. Four pairs of shRNA oligos targeting 606–626, 1518–1538,
1745–1765 and 2564–2584 nucleotides of BubR1 mRNA were constructed.
A scramble shRNA cassette was constructed as the negative control.
The shRNA vectors were transfected into cells using Lipofectamine
2000 (Invitrogen) according to the manufacturer’s instructions. The
efficacy of RNA interference was verified by western blot analysis
and RT-PCR assays of transfected cells.
RNA extraction and quantitative real-time
PCR
Total RNA of the clinical samples was isolated using
TRIzol reagent. RNA quality and integrity were verified using the
NanoDrop 1000 spectrophotometer (Thermo Scientific, Inc.) and RNA
gel electrophoresis. Two micrograms of total RNA of each sample was
reverse transcribed to cDNA with random hexamer primers and the
Prime Script kit (Takara). Quantitative PCR was performed using
Rotor Gene 6000 (Corbett Life Science, Qiagen). Each sample was
analyzed in triplicate. PCR primers were as follows: BubR1,
5′-ACGTTATTAGAAAGAGCTGTAG-3′ (forward) and
5′-CATATCCAAAGGCTCATTGC-3′ (reverse); GAPDH,
5′-CAGCGACACCCACTCCTC-3′ (forward) and 5′-TGAGGTCCACCACCCTGT-5′
(reverse). Gene expression was quantified using SYBR-Green I dye
real-time detection system. Thermal cycling conditions consisted of
an initial 2 min at 95°C, followed by 40 cycles for 20 sec at 95°C
and 20 sec at 57°C. Quantification was based on the relative
expression ratio of BubR1 in esophageal cancer tissue to adjacent
cancerous tissue after normalizing with internal GAPDH
expression.
Western blot analysis
Cells were lysed with RIPA lysis buffer (BiYunTian,
Shanghai, China) plus 1 mmol PMSF on ice for 30 min, and
centrifuged at 13000 × g at 4°C for an additional 30 min. Protein
concentrations were determined using the NanoDrop 1000
spectrophotometer (Thermo Scientific, Inc.). Each sample containing
30 μg total proteins was resolved by 8% SDS-PAGE and transferred
onto PVDF membranes (Millipore). Primary BubR1 antibody (1:2000,
mouse monoclonal antibody, BD Biosciences) and anti-β tubulin
(1:1000, Santa Cruz Biotechnology) were diluted in TBST buffer,
respectively, and applied on the membranes for overnight incubation
at 4°C. The PVDF membranes were incubated with horseradish
peroxidase-conjugated secondary antibody (goat; ZhongShan
Goldenbridge Biotechnology, China) for 1 h at room temperature.
After extensive washes with TBST, the protein bands of interest
were visualized using space chemoluminescence reagent (Pierce) in
the ChemiDoc™ XRS+ system (Bio-Rad, USA).
Cell viability assay
Cytotoxicity was determined by MTT assays (Promega).
Briefly, 5000 cells/well were seeded into 96-well plates in 100 μl
culture medium. The medium was then replaced by various
concentrations of paclitaxel. At 24, 48 and 72 h after treatment,
20 μl of 5 mg/ml MTT reagent was added to each well and incubated
for 4 h at 37°C. MTT formazan precipitates were completely
dissolved in DMSO with agitation. Optical density of each sample
was measured at 490 nm with a plate reader (Sunrise Remote; Tecan,
Switzerland).
Mitotic index analysis
ESC cells were treated with 0.2 μg/ml nocodazole for
the indicated time points and harvested via trypsinization. Cells
were suspended in a hypotonic solution containing 0.075 M KCL in
PBS to swell for 15 min at 37°C. After centrifugation, pellets were
fixed in three changes of ice cold newly prepared methanol/acetic
acid (3:1), and then incubated in fixative at 4°C overnight.
Metaphase spreads were prepared by dropping cell pellets onto glass
slides and staining with Giemsa and after the slides were air dried
at 37°C. At least 500 cells were counted in triplicate for each
sample. Mitotic and interphase cells were scored, and the mitotic
index was determined by dividing the number of mitotic cells by the
number of total counted cells.
Cell cycle analysis with FACS
Approximately 1×105 cells were plated in
6-cm dishes for 24 h. Paclitaxel was added (final concentration of
0.1 μM). Both floating and adherent cells were harvested at the
indicated time points by trypsinization, washed 3 times with PBS
and then fixed with 70% ethanol/PBS while gently vortexing the
tube. Cells were fixed at 4°C overnight and subjected to flow
cytometric analysis.
Statistical analysis
The clinical features of the groups with different
BubR1 expression were analyzed by the χ2 test (2×2 or
RxC contingency tables). Analysis of BubR1 relative expression was
performed using real-time quantitative PCR and 2−ΔΔCT
method. Other comparisons were carried out using the independent
sample t-test. A P-value <0.05 was considered to indicate a
statistically significant difference. All data were analyzed using
SPSS 19.0.
Results
High expression of BubR1 in clinical
esophageal cancer samples
We collected 50 cases of clinical ESC samples and
analyzed the relative expression of BubR1 between the cancer
tissues and adjacent non-cancerous tissues by quantitative RT-PCR.
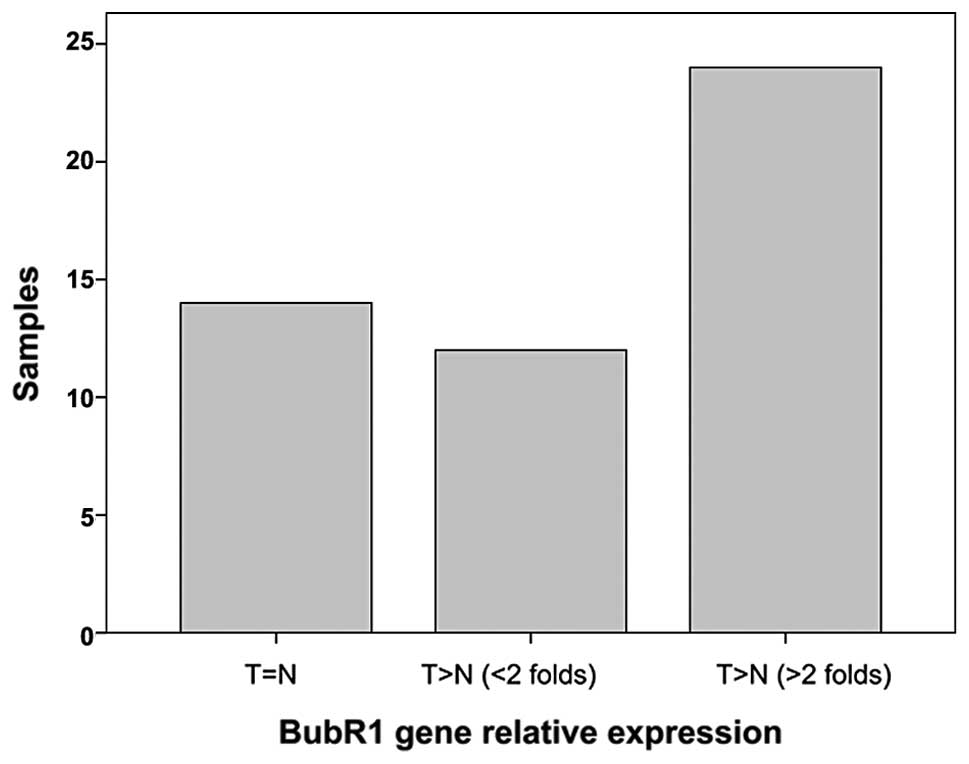
As shown in Fig. 1, BubR1 was
significantly overexpressed in the ESC cancer compared to the
adjacent non-cancerous tissues. Seventy-two percent (36 of 50) of
the samples showed relatively high BubR1 transcription (P<0.05),
and 64% (23 of 36) of those samples exhibited at least a 2-fold
higher BubR1 level than that the in non-cancerous tissues.
Previous studies have demonstrated that upregulation
of BubR1 in ovarian and tonsillar cancers was associated with lymph
node metastasis, tumor grade and stage (11,25).
We examined the relationship between the clinical histopathological
features and BubR1 expression in the ESC cases (Table I). No significant correlations were
found between BubR1 expression levels and the clinical
histopathological parameters.
 | Table IPatient characteristics and BubR1
expression. |
Table I
Patient characteristics and BubR1
expression.
| BubR1 expression
(n=50) | |
|---|
|
| |
|---|
| Features | Unchanged (n=14) | Increased (n=36) | P-value |
|---|
| Age (years), mean ±
SD | 60.3±7.9 | |
| Gender | | | 0.341 |
| Male | 12 | 27 | |
| Female | 2 | 9 | |
| Histopathologic
grading | | | 0.681 |
| Well
differentiation | 8 | 18 | |
| Moderate
differentiation | 5 | 12 | |
| Poor
differentiation | 1 | 6 | |
| Lymphatic
invasion | | | 0.522 |
| Negative | 8 | 22 | |
| Positive | 6 | 14 | |
| Stage | | | 0.811 |
| I | 1 | 1 | |
| II | 6 | 16 | |
| III | 5 | 16 | |
| IV | 2 | 3 | |
| Location of
tumor | | | 0.196 |
| Upper | 0 | 1 | |
| Middle | 9 | 30 | |
| Lower | 5 | 5 | |
Correlation of the high expression of
BubR1 in ESC cell lines with their resistance to anti-microtubule
drugs
Mitotic checkpoint protein BubR1 has been reportedly
involved in the cytotoxic effects of anti-microtubule drugs. We
tested whether high BubR1 expression in ESC cells correlates with
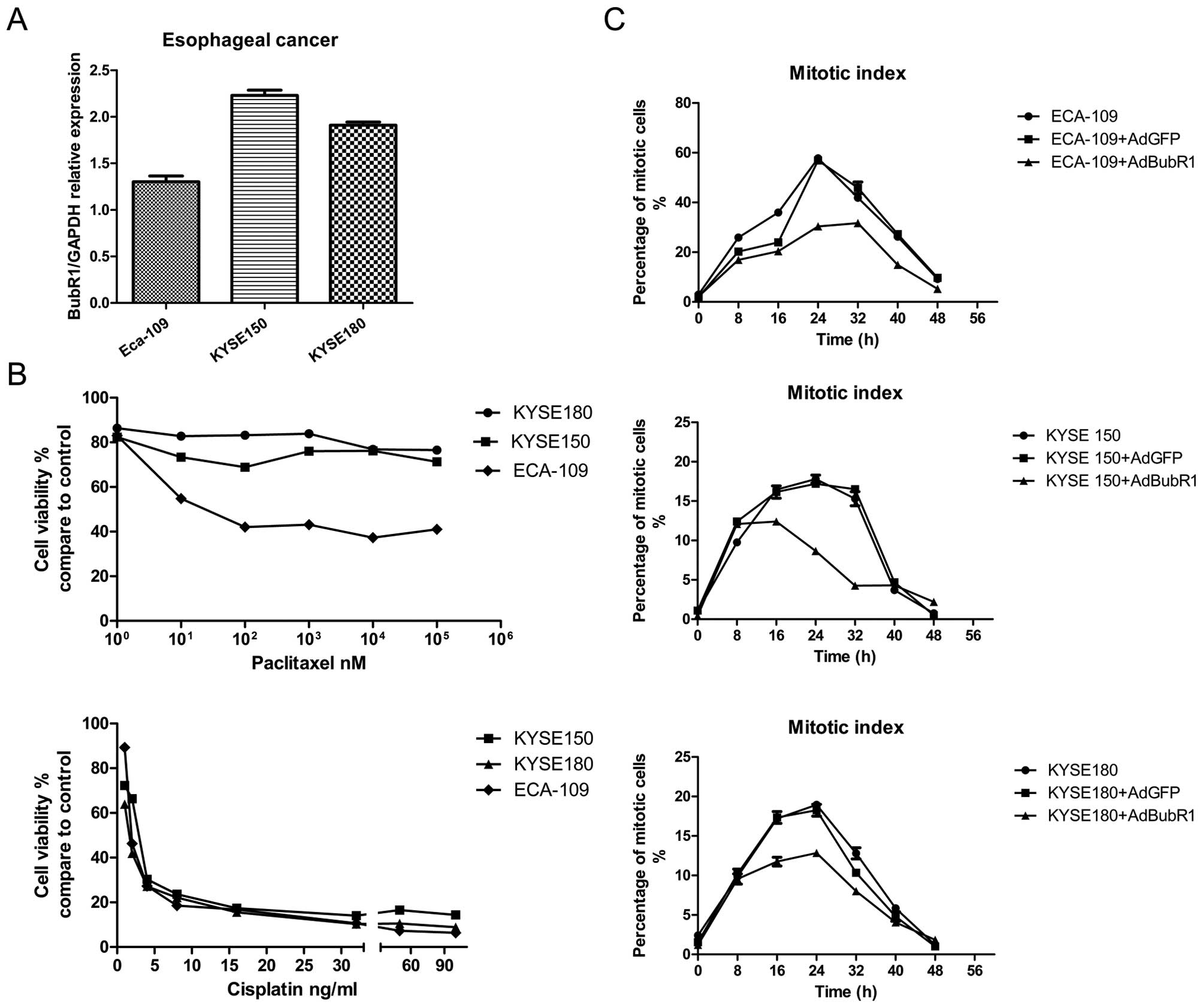
their paclitaxel sensitivity. We first determined the BubR1 mRNA
levels in three ESC cell lines using quantitative RT-PCR (Fig. 2A). Consistent with the results from
the clinical samples, all three cell lines exhibited a high level
of BubR1 expression. Compared with ECA-109, the KYSE150 and KYSE180
cell lines exhibited relatively higher expression of BubR1. Drug
sensitivity assay demonstrated that KYSE150 and KYSE180 cells were
less responsive to paclitaxel than ECA-109 (Fig. 2B). Cisplatin is clinically used for
esophageal cancer treatment and served as a positive control. The
responsiveness of cancer cells to paclitaxel is potentially linked
to mitotic progression and the functional mitotic checkpoint. The
mitotic index analysis showed that much fewer mitotic cells were
noted in the KYSE150 and KYSE180 cell lines when treated with the
anti-microtubule agent nocodazole, suggesting that the paclitaxel
insensitivity and mitosis progression may be linked in these ESC
cell lines. Accordingly, when BubR1 was overexpressed via an
adenoviral vector AdBubR1, all three cell lines showed a severe
disrupted mitotic progression (Fig.
2C). Thus, high BubR1 expression may be closely associated with
paclitaxel resistance.
Restoration of paclitaxel sensitivity by
RNAi-mediated BubR1 knockdown in esophageal cancer cells
To further explore the possibility that a high BubR1
level may lead to paclitaxel insensitivity, we constructed
expression vectors that contained four different shRNA pairs
targeting the BubR1 gene and a control shRNA with a scramble
sequence, respectively. These shRNA vectors were transfected into
ECA-109 cells and the interference effects were determined by
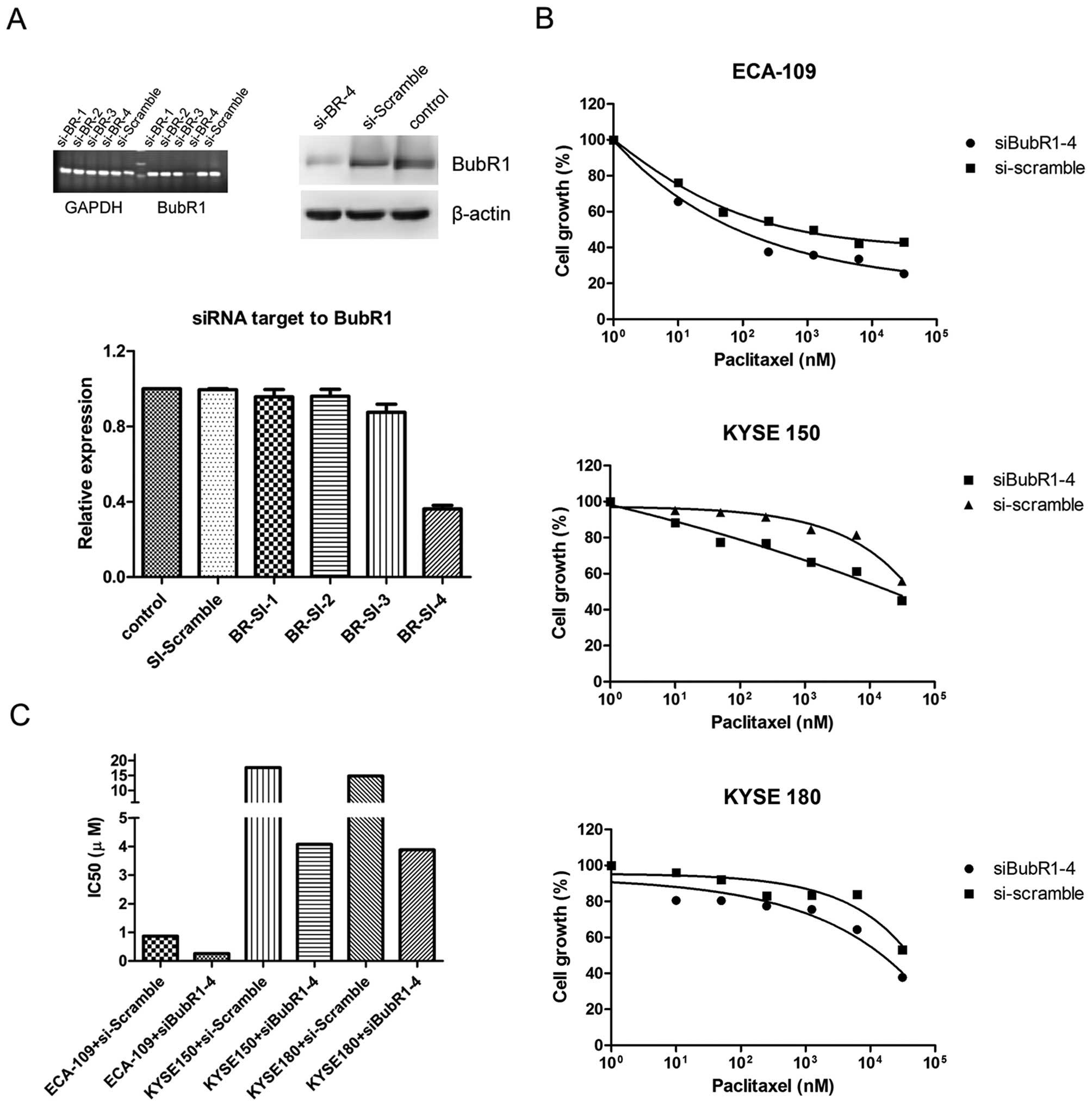
quantitative PCR and western blot assays. As shown in Fig. 3A, the BubR1 transcription level was
significantly reduced in the si-BubR1-4 lane, which was also
confirmed by western blot analysis. The three ESC lines were
transfected with si-BubR1-4 or si-Scramble plasmids. At 24 h, the
transfected cells were treated with paclitaxel. BubR1 suppression
in the three cell lines led to a moderate restoration of drug
sensitivity, and the respective drug IC50 values were
reduced (Fig. 3C), (i.e., ECA-109,
from 0.86 to 0.26 μM; KYSE150, from 17.68 to 4.07 μM; KYSE 180,
from 14.82 to 3.88 μM; P<0.05). We next sought to determine
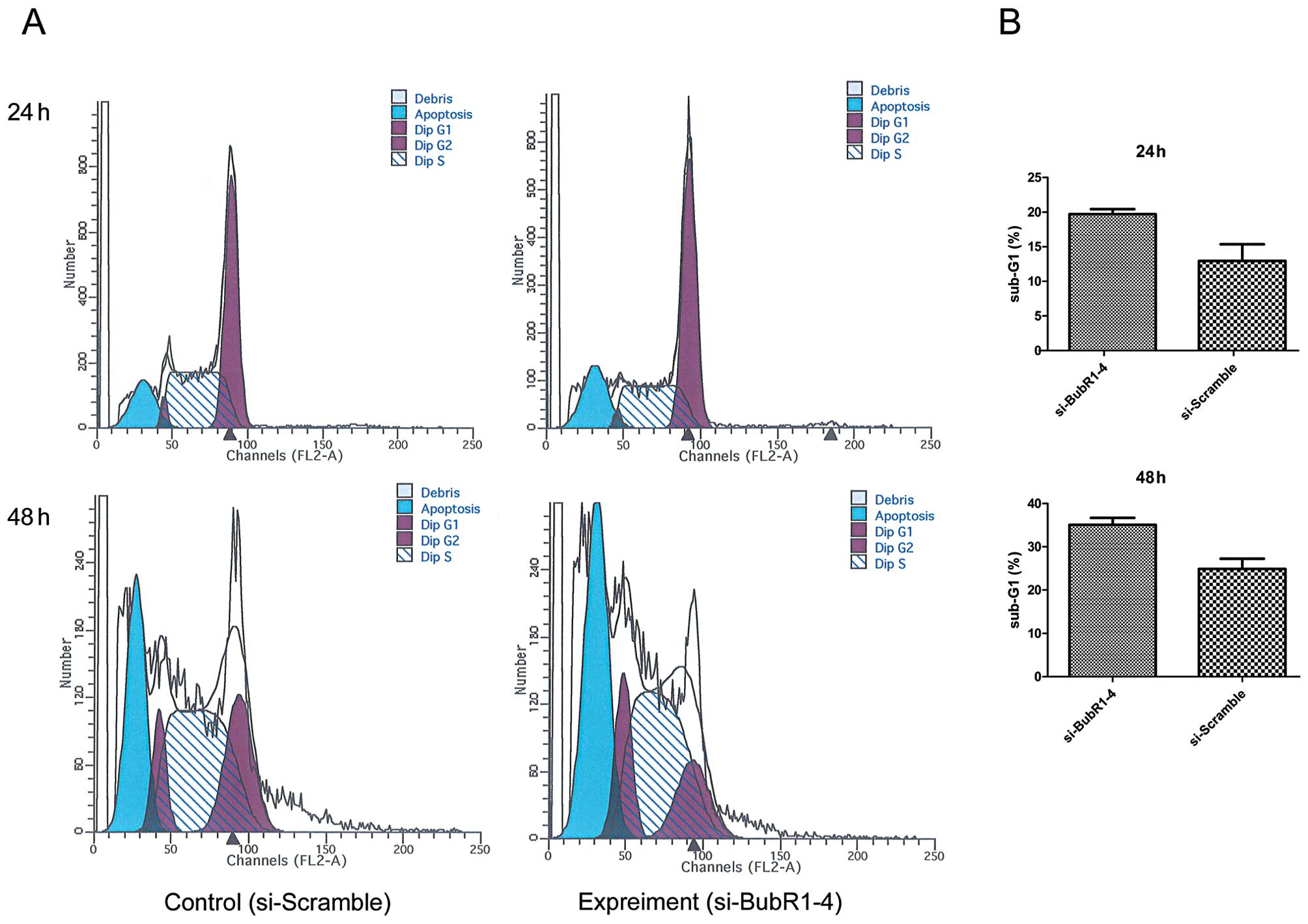
whether increased drug sensitivity was associated with cell death.
ECA-109 cells transfected with siRNA were treated with 100 nM
paclitaxel for 24 or 48 h, respectively, followed by the FACS
assay. FACS results showed that cell death was more severe in the
si-BubR1-4 group (Fig. 4).
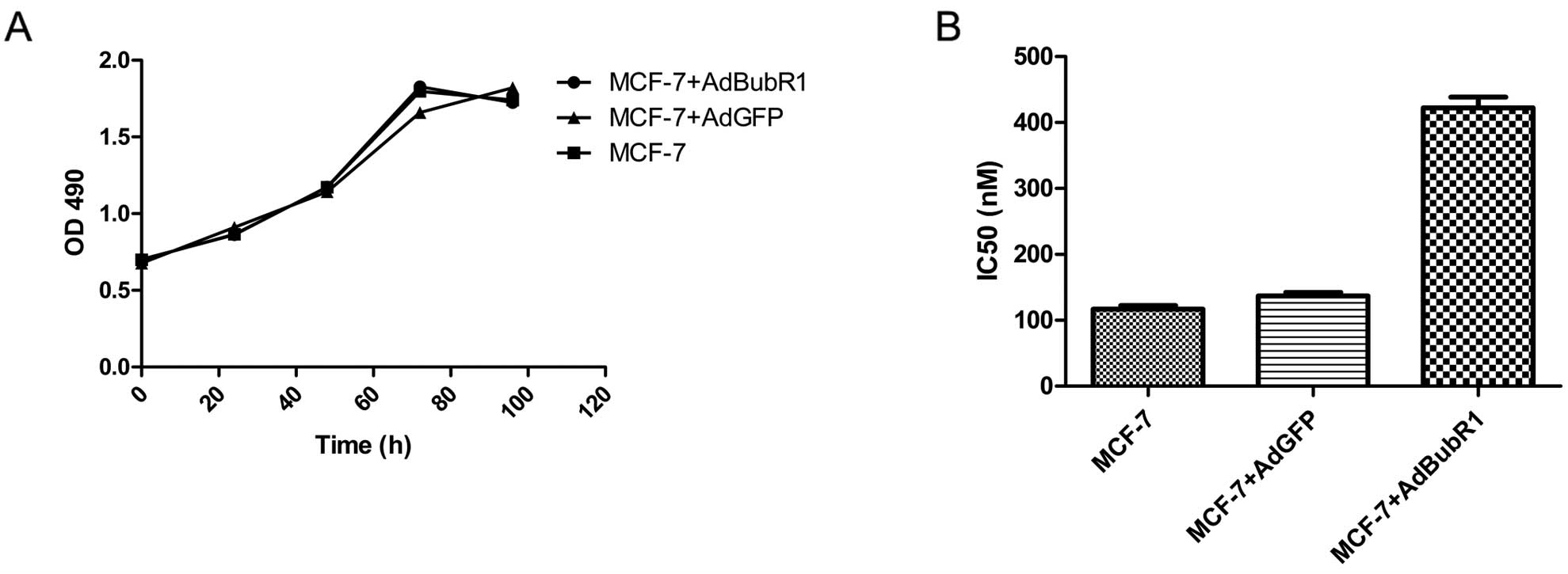
Induction of drug resistance by forced
expression of BubR1
As most ESC cell lines have an elevated BubR1 level
and are insensitive to anti-microtubule agents, we further tested
whether forced expression of BubR1 would confer drug resistance in
the cancer cells that were relatively sensitive to paclitaxel. We
transduced the breast cancer cell line MCF-7 with AdBubR1. The
infected cells were subjected to cell proliferation and drug
responsiveness assays. As shown in Fig.
5, overexpression of BubR1 did not affect MCF-7 cell
proliferation, but significantly increased the paclitaxel
resistance of the infected MCF-7 cells.
Discussion
Spindle assembly checkpoint (SAC), orchestrated by
multiple proteins, is a highly conversed surveillance mechanism
that exists in mitosis. BubR1 is one of the most important mitotic
phosphoproteins which serves both as a sensor to coordinate with
CENP-E for proper attachment between microtubules and the
kinetochore, and as an executor to delay the onset of the anaphase
by halting APC/C activity. Several clinical studies have indicated
that abnormal BubR1 expression is prevalent in cancers and is
somehow associated with tumor progression and prognosis (8–13).
BubR1 upregulation in esophageal cancer has also been reported and
cells with high BubR1 expression exhibit relatively low sensitivity
to paclitaxel. Here, we found that BubR1 expression was greatly
upregulated in clinical esophageal squamous cell cancer samples
compared with respective adjacent non-cancerous tissues, although
statistical analysis failed to identify any correlations between
BubR1 expression and tumor progression, or lymphatic invasion. It
is conceivable that BubR1 is not sufficient to serve as an
independent prognostic factor, but may be potentially valuable in
combination with other factors, as shown in the cases of breast and
colon cancer (8,26). While hBUB1b mutations are
rarely detected in clinical cancers, it remains to be determined
what causes BubR1 overexpression in a variety of cancers. However,
several findings may shed light on this issue. At the early stage
of esophageal cancer development, aberrant expression of cell cycle
regulators has previously been found in addition to oncogene
mutations. Analysis of the hBUB1b promoter region in lung
cancer cell lines found multiple transcription factor binding sites
(27). p53 and Myc proteins have
been found to interact with the hBUB1b promoter and regulate
its transcription (28). In
addition, the status of hBUB1b promoter methylation in
tumors may indirectly regulate BubR1 expression (29). Thus, BubR1 overexpression is likely
the consequence of an interaction of many regulatory signaling
complexes.
High BubR1 expression in gastric cancer was
significantly correlated with DNA aneuploidy (10). We found that forced BubR1
overexpression in esophageal cancer cel lines resulted in disrupted
mitotic progression. Consistent with a previous study (13), BubR1 overexpression is easily
detected in esophageal squamous cell cancer samples. Moreover,
esophageal squamous cell cancer cell lines with high BubR1
expression are seemingly insensitive to paclitaxel.
As a potent anti-microtubule drug, paclitaxel which
requires a functional mitotic checkpoint is one of the most
commonly used chemotherapeutic drugs in the clinic. However, its
application and anticancer efficacy are limited by its rapidly
developed drug resistance. The events that ensure that cells enter
into mitosis and that the mitotic checkpoint is properly activated,
no matter how long it lasts, guarantee anti-microtubule drug
efficiency (19,30). Therefore, paclitaxel sensitivity
will be affected under the condition of mitotic checkpoint failure
caused by abnormal checkpoint protein expression. Although BubR1
suppression severely impairs the mitotic checkpoint and confers
drug resistance to cancer cells, forced expression of BubR1 also
leads to mitotic checkpoint dysfunction and reduces drug
sensitivity, as shown in our studies.
Conversely, an RNAi-mediated depletion of BubR1 in
esophageal cancer cells contributes to a modest restoration of
paclitaxel sensitivity, as shown by a reduction in IC50
values and accumulation of the sub-G1 fraction. In cell lines
derived from breast and ovarian cancers, which also exhibit high
BubR1 expression in clinical samples, a decreased BubR1 level is
associated with augmented paclitaxel-induced cell death (20,21).
Thus, it is conceivable that either suppression or promotion of
BubR1 expression can disrupt the function of the mitotic checkpoint
and may subsequently diminish the effects of paclitaxel. In the
case of esophageal squamous cell cancer, overexpression of BubR1 is
seemingly a dominant event, conferring cancer cells with
insensitivity to paclitaxel. Thus, future studies should be
directed to determine the role of BubR1 expression in the
development of drug resistance against anti-microtubule drugs.
In summary, our study revealed that mitotic
checkpoint BubR1 is overexpressed in clinical esophageal squamous
cell cancer tissues and in esophageal squamous cell cancer cell
lines. Esophageal squamous cell cancer cells with high BubR1
expression show resistance to the anti-microtubule drug paclitaxel,
while knockdown of BubR1 significantly sensitizes the
responsiveness of these cells to paclitaxel. Thus, our findings
strongly suggest that BubR1 may serve as a potential
chemosensitizing target to overcome the chemoresistance of ESC.
Acknowledgements
The reported study was supported in part by research
grants from the Special Academic Foundation of Doctor Degrees of
Chongqing Medical University funded by the Ministry of Education of
China (No. 20115503110009), and was supported by grants 30872770
from the National Natural Science Foundation of China
(NSFC30872770), Natural Science Foundation Project of CQ CSTC
(CSTC, 2011BB5131) and the National Ministry of Education
Foundation of China (KJ120327).
References
|
1
|
Parkin DM: Global cancer statistics in the
year 2000. Lancet Oncol. 2:533–543. 2001.PubMed/NCBI
|
|
2
|
Stathopoulos GP and Tsiaras N:
Epidemiology and pathogenesis of esophageal cancer: management and
its controversial results (Review). Oncol Rep. 10:449–454.
2003.PubMed/NCBI
|
|
3
|
Musacchio A and Salmon ED: The
spindle-assembly checkpoint in space and time. Nat Rev Mol Cell
Biol. 8:379–393. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bolanos-Garcia VM and Blundell TL: BUB1
and BUBR1: multifaceted kinases of the cell cycle. Trends Biochem
Sci. 36:141–150. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Elowe S, Dulla K, Uldschmid A, Li X, Dou Z
and Nigg EA: Uncoupling of the spindle-checkpoint and
chromosome-congression functions of BubR1. J Cell Sci. 123:84–94.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sotillo R, Hernando E, Díaz-Rodríguez E,
Teruya-Feldstein J, Cordón-Cardo C, Lowe SW and Benezra R: Mad2
overexpression promotes aneuploidy and tumorigenesis in mice.
Cancer Cell. 11:9–23. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ricke RM, Jeganathan KB and van Deursen
JM: Bub1 overexpression induces aneuploidy and tumor formation
through Aurora B kinase hyperactivation. J Cell Biol.
193:1049–1064. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuan B, Xu Y, Woo JH, et al: Increased
expression of mitotic checkpoint genes in breast cancer cells with
chromosomal instability. Clin Cancer Res. 12:405–410. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yamamoto Y, Matsuyama H, Chochi Y, et al:
Overexpression of BUBR1 is associated with chromosomal instability
in bladder cancer. Cancer Genet Cytogen. 174:42–47. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ando K, Kakeji Y, Kitao H, et al: High
expression of BUBR1 is one of the factors for inducing DNA
aneuploidy and progression in gastric cancer. Cancer Sci.
101:639–645. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee YK, Choi E, Kim MA, Park PG, Park NH
and Lee H: BubR1 as a prognostic marker for recurrence-free
survival rates in epithelial ovarian cancers. Br J Cancer.
101:504–510. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pinto M, Vieira J, Ribeiro FR, et al:
Overexpression of the mitotic checkpoint genes BUB1 and BUBR1 is
associated with genomic complexity in clear cell kidney carcinomas.
Cell Oncol. 30:389–395. 2008.PubMed/NCBI
|
|
13
|
Tanaka K, Mohri Y, Ohi M, et al: Mitotic
checkpoint genes, hsMAD2 and BubR1, in oesophageal squamous cancer
cells and their association with 5-fluorouracil and cisplatin-based
radiochemotherapy. Clin Oncol (R Coll Radiol). 20:639–646. 2008.
View Article : Google Scholar
|
|
14
|
Hanks S, Coleman K, Reid S, et al:
Constitutional aneuploidy and cancer predisposition caused by
biallelic mutations in BUB1B. Nat Genet. 36:1159–1161. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dai W, Wang Q, Liu T, et al: Slippage of
mitotic arrest and enhanced tumor development in mice with BubR1
haploinsufficiency. Cancer Res. 64:440–445. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baker DJ, Perez-Terzic C, Jin F, et al:
Opposing roles for p16Ink4a and p19Arf in senescence and ageing
caused by BubR1 insufficiency. Nat Cell Biol. 10:825–836. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baker DJ, Jeganathan KB, Cameron JD, et
al: BubR1 insufficiency causes early onset of aging-associated
phenotypes and infertility in mice. Nat Genet. 36:744–749. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim M, Murphy K, Liu F, Parker SE, Dowling
ML, Baff W and Kao GD: Caspase-mediated specific cleavage of BubR1
is a determinant of mitotic progression. Mol Cell Biol.
25:9232–9248. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sudo T, Nitta M, Saya H and Ueno NT:
Dependence of paclitaxel sensitivity on a functional spindle
assembly checkpoint. Cancer Res. 64:2502–2508. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee EA, Keutmann MK, Dowling ML, Harris E,
Chan G and Kao GD: Inactivation of the mitotic checkpoint as a
determinant of the efficacy of microtubule-targeted drugs in
killing human cancer cells. Mol Cancer Ther. 3:661–669.
2004.PubMed/NCBI
|
|
21
|
Fu Y, Ye D, Chen H, Lu W, Ye F and Xie X:
Weakened spindle checkpoint with reduced BubR1 expression in
paclitaxel-resistant ovarian carcinoma cell line SKOV3-TR30.
Gynecol Oncol. 105:66–73. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamada HY and Gorbsky GJ: Inhibition of
TRIP1/S8/hSug1, a component of the human 19S proteasome, enhances
mitotic apoptosis induced by spindle poisons. Mol Cancer Ther.
5:29–38. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shin HJ, Baek KH, Jeon AH, et al: Dual
roles of human BubR1, a mitotic checkpoint kinase, in the
monitoring of chromosomal instability. Cancer Cell. 4:483–497.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo J, Deng Z-L, Luo X, et al: A protocol
for rapid generation of recombinant adenoviruses using the AdEasy
system. Nat Protoc. 2:1236–1247. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hannisdal K, Burum-Auensen E, Schjolberg
A, De Angelis PM and Clausen OP: Correlation between reduced
expression of the spindle checkpoint protein BubR1 and bad
prognosis in tonsillar carcinomas. Head Neck. 32:1354–1362. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rao CV, Yang YM, Swamy MV, et al: Colonic
tumorigenesis in
BubR1+/−ApcMin/+ compound
mutant mice is linked to premature separation of sister chromatids
and enhanced genomic instability. Proc Natl Acad Sci USA.
102:4365–4370. 2005.PubMed/NCBI
|
|
27
|
Seike M, Gemma A, Hosoya Y, et al: The
promoter region of the human BUBR1 gene and its expression analysis
in lung cancer. Lung Cancer. 38:229–234. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Menssen A, Epanchintsev A, Lodygin D, et
al: c-MYC delays prometaphase by direct transactivation of MAD2 and
BubR1: identification of mechanisms underlying c-MYC-induced DNA
damage and chromosomal instability. Cell Cycle. 6:339–352. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Park HY, Jeon YK, Shin HJ, et al:
Differential promoter methylation may be a key molecular mechanism
in regulating BubR1 expression in cancer cells. Exp Mol Med.
39:195–204. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu YC, Yen WY and Yih LH: Requirement of a
functional spindle checkpoint for arsenite-induced apoptosis. J
Cell Biochem. 105:678–687. 2008. View Article : Google Scholar : PubMed/NCBI
|