Introduction
Hepatocellular carcinoma (HCC), the major type of
liver cancer, is the fifth most frequent neoplasm and the third
most common cause of cancer-related mortality in the world,
particularly in Asia and sub-Saharan Africa (1). Epidemiological studies have
demonstrated that hepatitis B virus (HBV) infection is one of the
key risk factors for HCC (2),
especially in eastern Asia. In western countries where hepatitis B
infections are rare, however, risk factors for HCC include
cirrhosis due to alcohol consumption or hepatitis C virus (HCV)
infection (3,4).
microRNAs (miRNAs) are evolutionarily-conserved,
small, non-coding RNA molecules, approximately 22 nucleotides in
length, which are cleaved from a 70- to 80-nt partially duplexed
precursor (pre-miRNA). miRNAs typically regulate
post-transcriptional gene expression by interacting with sequences
within the 3′-untranslational region (3′-UTR) of the target mRNA
and play important roles in a variety of biological processes,
including cell cycle regulation, cell differentiation,
proliferation and apoptosis (5,6).
Accumulating evidence indicates that miRNAs are extensively
involved in the pathogenesis of several types of human cancer
(7). Approximately 50% of the genes
that encode miRNAs reside in cancer-associated genomic regions or
fragile sites and this fact highlights the importance of miRNAs in
tumorigenesis (8).
miR-96 has been recognized as an oncogenic miRNA
that is upregulated in various types of cancer (9–11). In
HCC, the role of miR-96 remains largely unknown. Recent studies
identified the deregulation of miR-96 as being significantly
associated with different risk factors for HCC, including HBV
infection and alcohol consumption. Moreover, miR-96 was upregulated
in several HCC cell lines (12).
The FOXO subfamily of Forkhead transcription factors
are key tumor suppressors in mammalian cells (13). To date, suppression of FOXOs in
cancer cells was thought to be mainly due to activation of multiple
onco-kinases by a phosphorylation-ubiquitylation-mediated cascade.
Evidence suggests that members of the FOXO family are regulated by
miRNAs. The level of miR-96 was significantly upregulated in
endometrial cancer compared with normal endometrium and
overexpression of miR-96 effectively downregulated forkhead box O1
(FOXO1) expression in endometrial cancer cells (14). The expression of miR-96 was also
markedly upregulated in breast cancer cells and tissues, compared
with normal breast epithelial cells and normal breast tissues
(11). Further study demonstrated
that miR-96 downregulated forkhead box O3a (FOXO3a) expression in
breast cancer cells by directly targeting the FOXO3a 3′-UTR
(9).
Based on these findings, we selected miR-96 for
further investigation and hypothesized that miR-96 could influence
the biological behaviors of HCC cells and affect the expression of
FOXO1 and FOXO3a in HCC cells. Our results indicated that
downregulation of miR-96 effectively upregulated FOXO1 and FOXO3a
expression, and also inhibited cell proliferation and
clonogenicity. We concluded that miR-96 may play an important role
as an oncogenic miRNA in the development and progression of HCC by
regulating the expression of FOXO1 and FOXO3a.
Materials and methods
Cell culture and transfection
HepG2 cells (ATCC, Manassas, VA, USA) were
maintained in Dulbecco’s minimal essential medium (DMEM)
(Invitrogen Inc., USA), supplemented with 10% fetal bovine serum
(FBS) (Gibco, Grand Island, NY, USA) and 1% penicillin/streptomycin
at 37°C under an atmosphere of 5% carbon dioxide. Normal primary
human hepatocytes (PHHCs) were isolated using a modified four-step
retrograde perfusion technique according to the ethical and
institutional guidelines and following the informed consent of the
tissue donors as previously described (15). The study was approved by the
Research Ethics Committee at the Tongji Hospital of Huazhong
University of Science and Technology.
The miR-96 inhibitor, anti-miR-96 (a
chemically-modified single-stranded nucleic acid targeting miR-96)
and the corresponding negative control, anti-miR-NC (a
chemically-modified single-stranded nucleic acid of random
sequence) were obtained from Ambion, Inc. (Austin, TX, USA). Small
interfering RNAs (siRNAs) targeting FOXO1 and FOXO3a and their
negative controls (siR-NC) were purchased from RiboBio Co., Ltd.
(Guangzhou, China). The sequences of the FOXO1-specific siRNA
(siR-FOXO1) (16) and
FOXO3a-specific siRNA (siR-FOXO3a) (17) are listed in Table I. Cells were transfected with siRNA
or miRNA inhibitor using Lipofectamine 2000 (Invitrogen, Carlsbad,
CA, USA) according to the manufacturer’s instruction.
 | Table IsiRNAs for FOXO1 and FOXO3a. |
Table I
siRNAs for FOXO1 and FOXO3a.
| siRNA name | Primer sequence
(5′→3′) |
|---|
| siR-FOXO1 |
| Sense |
GCGGGCUGGAAGAAUUCAAdTdT |
| Antisense |
UUGAAUUCUUCCAGCCCGCdTdT |
| siR-FOXO3a |
| Sense |
GCACAGAGUUGGAUGACGUdTdT |
| Antisense | ACGUCAUCCAACU
CUGUGCdTdT |
Cell proliferation assay
Cell proliferation was assessed at 24, 48 and 72 h
after transfection using the CellTiter 96® AQueous One
Solution Cell Proliferation Assay kit (Promega, Madison, WI, USA)
as previously described (15).
Flow cytometry analysis of cell cycle and
apoptosis
To analyze the effects of siRNA or miRNA inhibitors
on the cell cycle, cells were seeded in 6-well plates. Seventy-two
hours after transfection, cells were collected and fixed in 70%
ethanol at 4°C overnight. After washing and resuspending the fixed
cells in 100 μl phosphate-buffered saline, they were treated with
50 μl RNase (50 mg/ml) at 37°C for 30 min and stained with 200 μl
propidium iodide (KeyGen Biotech. Co., Ltd., Nanjing, China) at 4°C
for 30 min. Cell cycle analysis was performed using a FACSAria cell
sorting system (BD Biosciences, San Jose, CA, USA). For analysis of
apoptosis, cells seeded in 24-well plates were harvested 72 h after
transfection and processed with the Annexin V-FITC Apoptosis
Detection kit (KeyGen Biotech Co., Ltd.) according to the
manufacturer’s instructions.
Cell migration assay
The migration assay was performed by using Transwell
insert chambers (6.5 mm in diameter, 8 μm pore size; Corning, USA).
For the migration assay, 2×105 HepG2 cells, that had
been transfected with siRNA or miRNA inhibitor, were placed into
the upper chamber in serum-free medium in triplicate 12 h after
transfection. The lower chamber was filled with 600 μl DMEM with
10% FBS as the chemoattractant. After incubation for 24 h at 37°C
in a humidified incubator with an atmosphere of 5% carbon dioxide,
non-migrating cells in the upper chambers were removed by using a
cotton swab, and cells that had migrated to the lower surface of
the membrane were fixed with methanol and stained with 0.1% crystal
violet. The cells that had migrated were scored by counting at
least six fields per membrane under a light microscope.
Analysis of clonogenicity in vitro
A colony formation assay was used to assess the
clonogenicity of HepG2 cells transfected with different miRNAs or
siRNAs. To analyze the ability of transfected cells to form
colonies, 100 viable HepG2 transfectants were placed in 24-well
plates 24 h after transfection and were maintained in complete
medium for 2 weeks. Colonies were fixed with methanol and stained
with 0.1% crystal violet in 20% methanol. The anchorage-independent
growth assay was performed in 24-well plates. Eight hundred cells
were trypsinized and suspended in 1 ml complete medium containing
0.3% low-melting-point agarose (Sigma, St. Louis, MO, USA). The
agar-cell mixture was plated on top of a bottom layer with 0.5%
complete medium agar mixture. Two weeks later, colonies were
visualized by staining with 0.05% crystal violet and viable
colonies that contained >50 cells or were >0.1 mm were
counted with an ocular micrometer. Each transfectant was seeded
into wells in triplicate and the experiment was repeated twice.
Real-time quantitative PCR
Detection of miR-96 expression
Total RNA was extracted from cells with the mirVana
miRNA isolation kit (Ambion). Synthesis of cDNA from 10 ng total
RNA was carried out using the TaqMan miRNA Reverse Transcription
kit (Ambion) and cDNA was then subjected to TaqMan Quantitative
Real-Time PCR using an miR-96-specific TaqMan microRNA assay on an
ABI PRISM StepOnePlus Real-Time PCR System (Applied Biosystems,
Foster City, CA, USA). Expression levels of the miRNAs were
normalized to that of snRNA U6, which served as the internal
control.
Detection of mRNA expression
Total RNA was prepared using the mirVana PARIS kit
(Ambion) and the reverse transcription reactions were performed
using the First Strand cDNA Synthesis kit (Fermentas, Burlington,
ON, Canada). Synthesized cDNA was used as a template for PCR and
the quantitative reactions were monitored in real-time using the
SYBR Green dye detections with the SYBR Green Premix Reagent
(Takara Bio, Inc., Shiga, Japan). The primers used for FOXO1,
FOXO3a and β-actin are listed in Table
II and were synthesized by Sangong Biotech Co., Ltd. (Shanghai,
China). β-actin was used as an internal standard for the
quantification of the PCR. The relative expression was calculated
using the comparative threshold cycle (Ct) method.
| Table IIPrimers used for quantitative RT-PCR
with SYBR Green dye detection. |
Table II
Primers used for quantitative RT-PCR
with SYBR Green dye detection.
| Primer name | Primer sequence
(5′→3′) |
|---|
| FOXO1 |
| Sense |
GACAGCCCTGGATCACAGTTT |
| Antisense |
CGGTCATAATGGGTGAGAGTCT |
| FOXO3a |
| Sense |
TTCCGTAAGCAAGCCGTGTA |
| Antisense |
CGAGTCCGAAGTGAGCAGGT |
| β-actin |
| Sense |
AGAGGGAAATCGTGCGTGAC |
| Antisense |
CAATAGTGATGACCTGGCCGT |
Western blotting
Total proteins were harvested with the mirVana PARIS
kit (Ambion) and quantified by a BCA protein assay kit (Pierce,
Rockford, IL, USA). Cell lysates were separated by electrophoresis,
blotted onto membranes (0.45 μm; Millipore) and treated with the
following primary antibodies: rabbit anti-FOXO1 (1:1,000;
Epitomics, Burlingame, CA, USA), rabbit anti-FOXO3a antibody
(1:1,000; Epitomics), rabbit anti-β-actin antibody (1:1,000;
Abmart, Shanghai, China). The blots were then incubated with an
anti-rabbit secondary antibody conjugated to horseradish peroxidase
and visualized by using ECL reagents (Pierce).
Statistical analysis
Data are presented as the means ± SD. Differences
between two groups were evaluated using the Student’s t-test. A
value of P<0.05 was considered to indicate statistically
significant differences.
Results
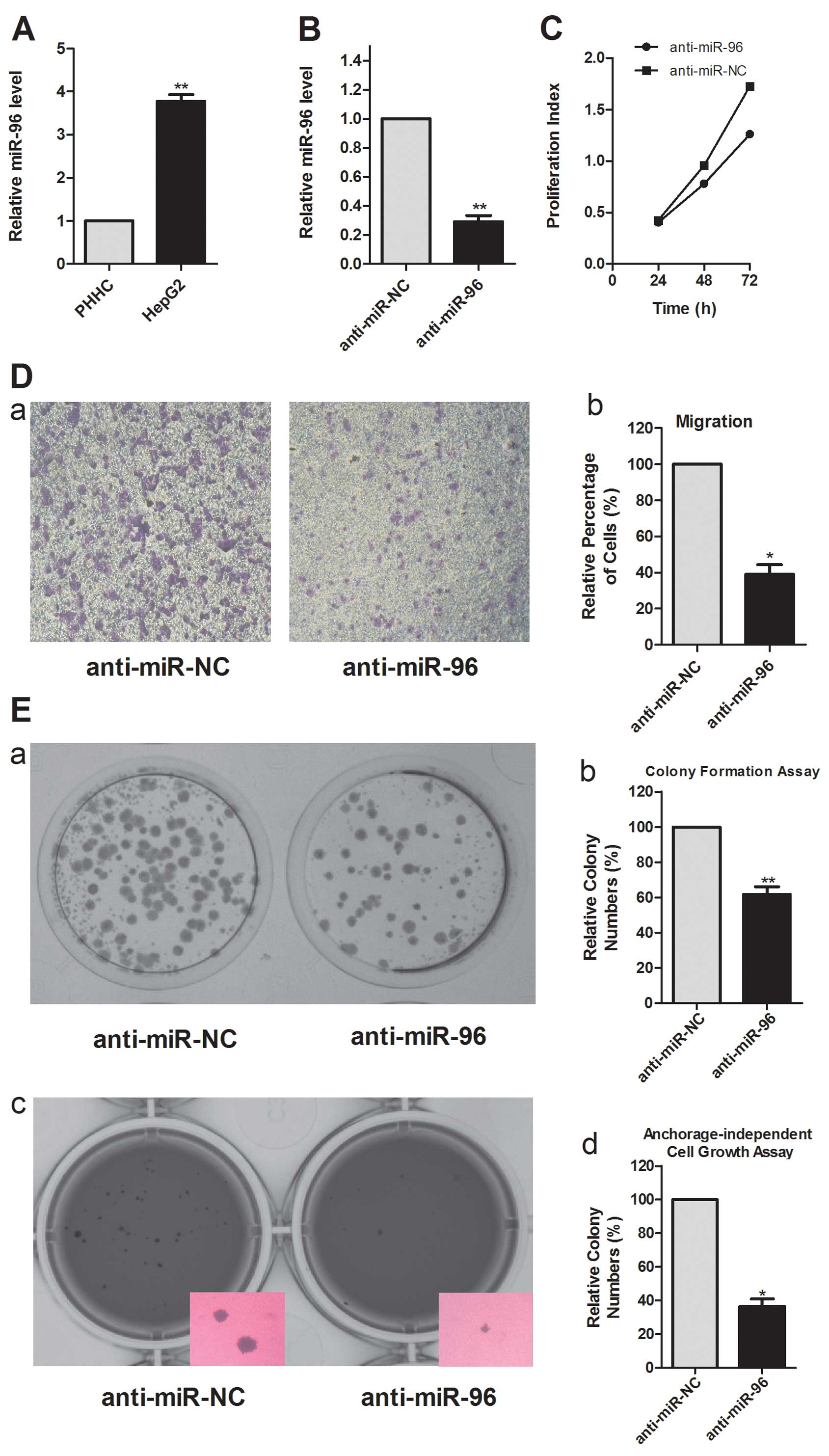
miR-96 is upregulated in HepG2 cells and
is efficiently inhibited by anti-miR-96
To determine whether miR-96 is upregulated in human
HCC, we detected the expression of miR-96 in HepG2 and PHHC cells
by real-time PCR analysis. We found that miR-96 was expressed in
significantly higher levels in HepG2 cells, compared to the levels
in PHHCs (Fig. 1A). This
observation is consistent with previous miRNA microarray data that
suggested a link between the upregulation of miR-96 and progression
to cirrhosis and HCC (12). To
investigate the pathophysiological role of miR-96 in HCC, we
performed loss-of-function analysis using anti-miR-96 to inhibit
endogenous miR-96. miR-96 was inhibited in HepG2 cells by
anti-miR-96 (Fig. 1B). These
results show that miR-96 expression is indeed increased in HCCs and
that anti-miR-96 can effectively block the expression of
miR-96.
miR-96 modulates cell proliferation,
migration and colony formation of HepG2 cells
Sustained cell growth is a hallmark of cancer. In
the HepG2 cell line, transfection with anti-miR-96 markedly
decreased cell proliferation (Fig.
1C). This result indicates that the inhibition of miR-96
reduces the growth of HCC cells and suggests that miR-96 is
necessary for maintaining the proliferation of HCC cells.
We assessed the role of miR-96 in cell migration, a
key determinant of malignant progression and metastasis. Transwell
insert chambers were introduced to investigate the impact of miR-96
on cell motility. HepG2 cells were transfected with either the
miR-96 inhibitor or a negative control, and vertical migration was
assessed. Transfection with anti-miR-96 caused a significant
decrease in cell migration, compared with anti-miR-NC transfection
(Fig. 1D). This result supports a
functional role for miR-96 in mediating cell migration in malignant
hepatocytes.
To assess the function of miR-96 in tumor formation,
colony formation and anchorage-independent growth were measured in
HepG2 cells. Analysis of colony formation showed that cells
transfected with anti-miR-96 displayed much fewer and smaller
colonies than the control transfectants (Fig. 1E). Inhibition of miR-96 also reduced
the anchorage-independent growth of HepG2 cells significantly, as
shown by the decrease in the colony number and size (Fig. 1E).
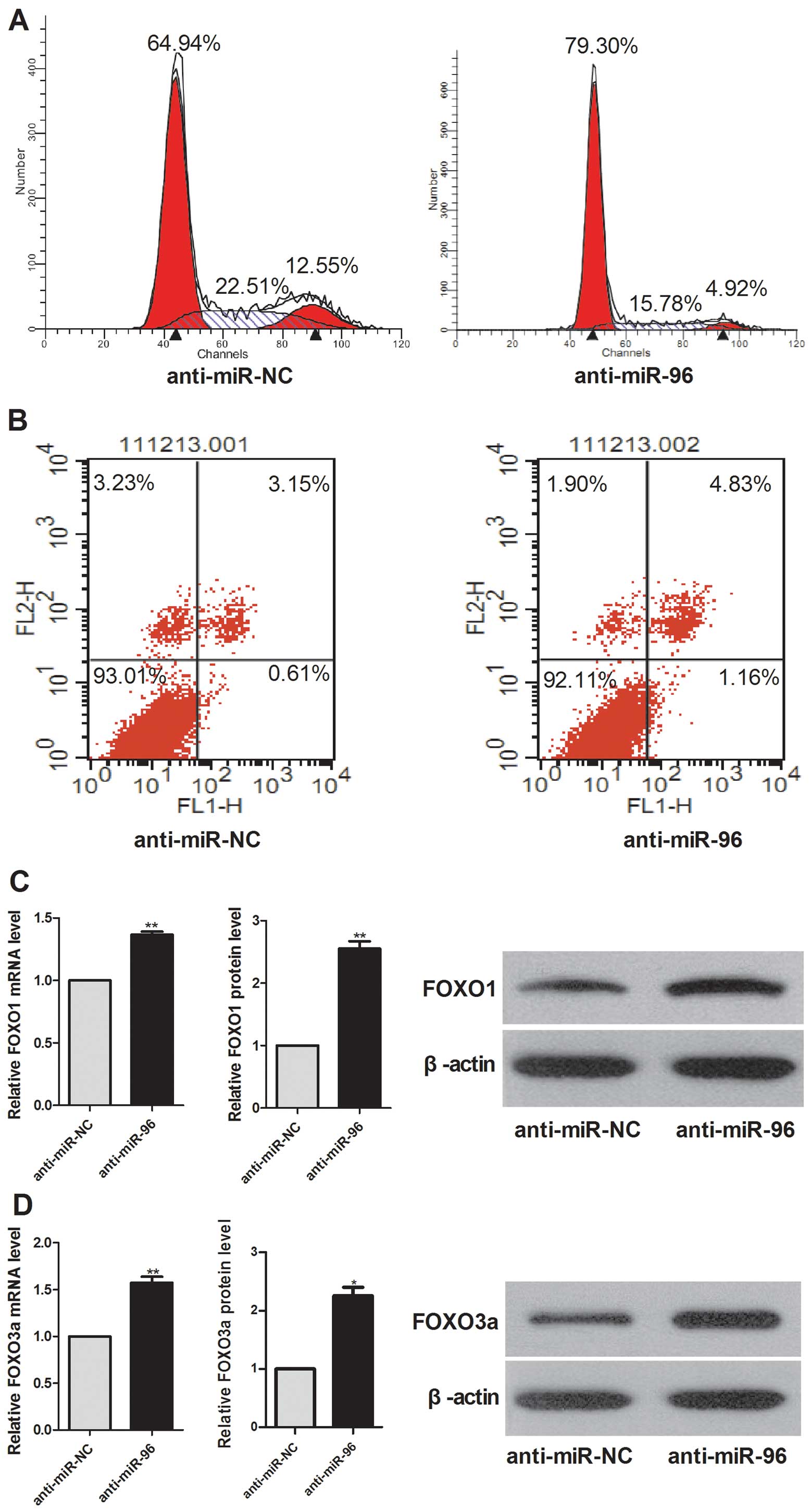
miR-96 affects the cell cycle but does
not induce apoptosis of HepG2 cells
To delineate a possible role of miR-96 in cell
division, cell cycle regulation and apoptosis FACS analysis,
followed by flow cytometry, was carried out. Compared to the
negative control, anti-miR-96 transfection caused significant cell
cycle arrest (Fig. 2A), but did not
induce significant apoptosis (Fig.
2B) in HepG2 cells.
miR-96 inhibits the expression of FOXO1
and FOXO3a
Real-time RT-PCR demonstrated that FOXO1 and FOXO3a
mRNA levels were upregulated in anti-miR-96-transfected HepG2 cells
compared with cells transfected with the negative control and that
the protein levels of FOXO1 and FOXO3a were substantially increased
when miR-96 was inhibited (Fig. 2C and
D). These results indicate that FOXO1 and FOXO3a might be the
targets of miR-96 in HepG2 cells.
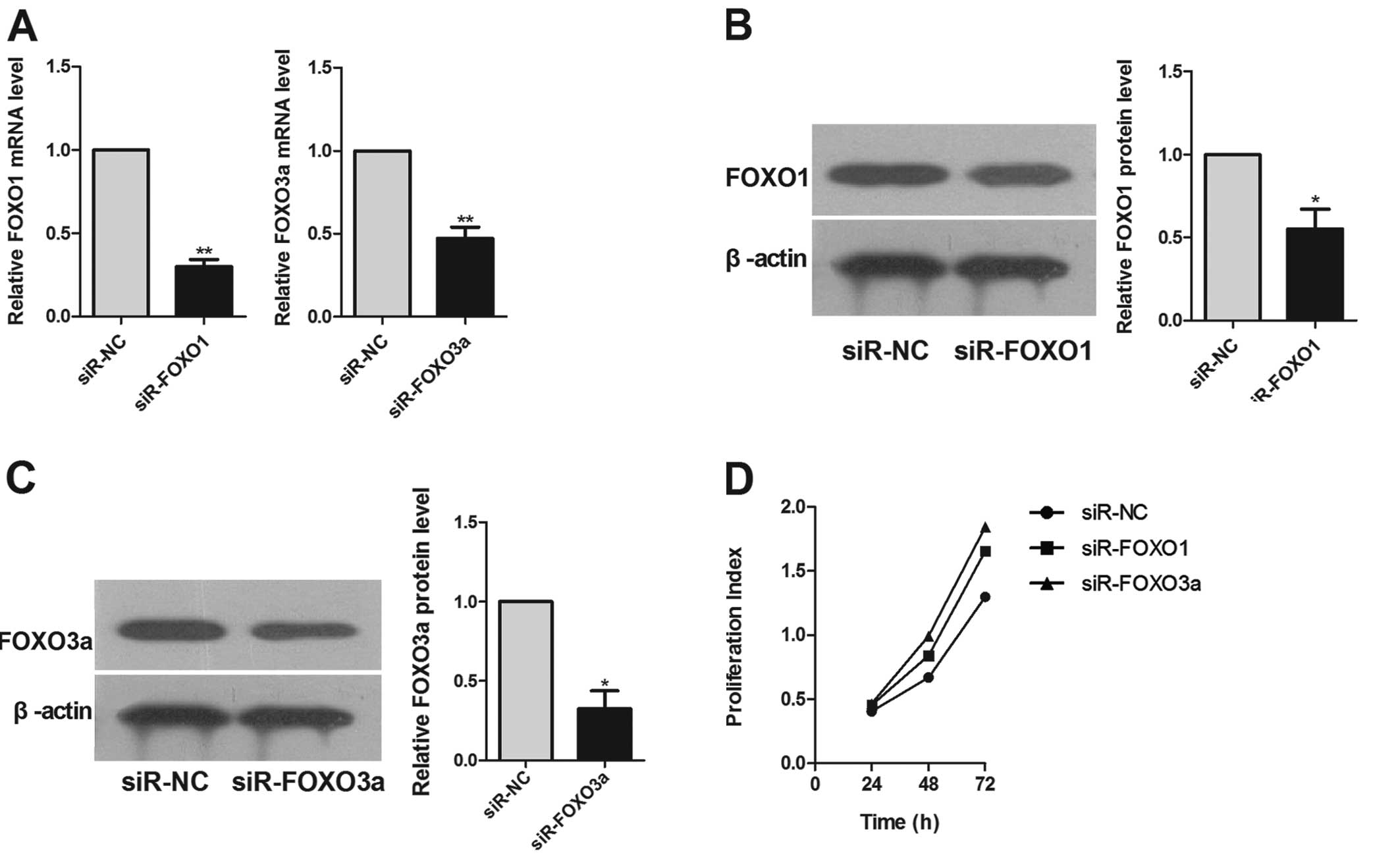
FOXO1 and FOXO3a are efficiently
inhibited by siRNAs
To assess whether siRNAs could alter the expression
of FOXO1 and FOXO3a in HCC cells, we investigated the expression of
FOXO1 and FOXO3a in HepG2 cells transfected with FOXO1- and
FOXO3a-specific siRNAs, or a negative control. Real-time RT-PCR
demonstrated that FOXO1 and FOXO3a mRNA levels were downregulated
in HepG2 cells transfected with the specific siRNAs, but not with
the corresponding negative controls (Fig. 3A) and that the protein level of
FOXO1 and FOXO3a were substantially decreased as well (Fig. 3B and C). These results indicate that
the expression of FOXO1 and FOXO3a are efficiently decreased by
siRNAs in HepG2 cells.
FOXO1 and FOXO3a modulate HepG2 cell
proliferation, migration and colony formation
Transfection of HepG2 cells with siRNAs targeting
FOXO1 and FOXO3a markedly promoted cell proliferation (Fig. 3D) and caused a significant increase
in cell migration, compared with the corresponding negative
controls (Fig. 4A). This result
supports a functional role for FOXO1 and FOXO3a in mediating cell
proliferation and migration in malignant hepatocytes.
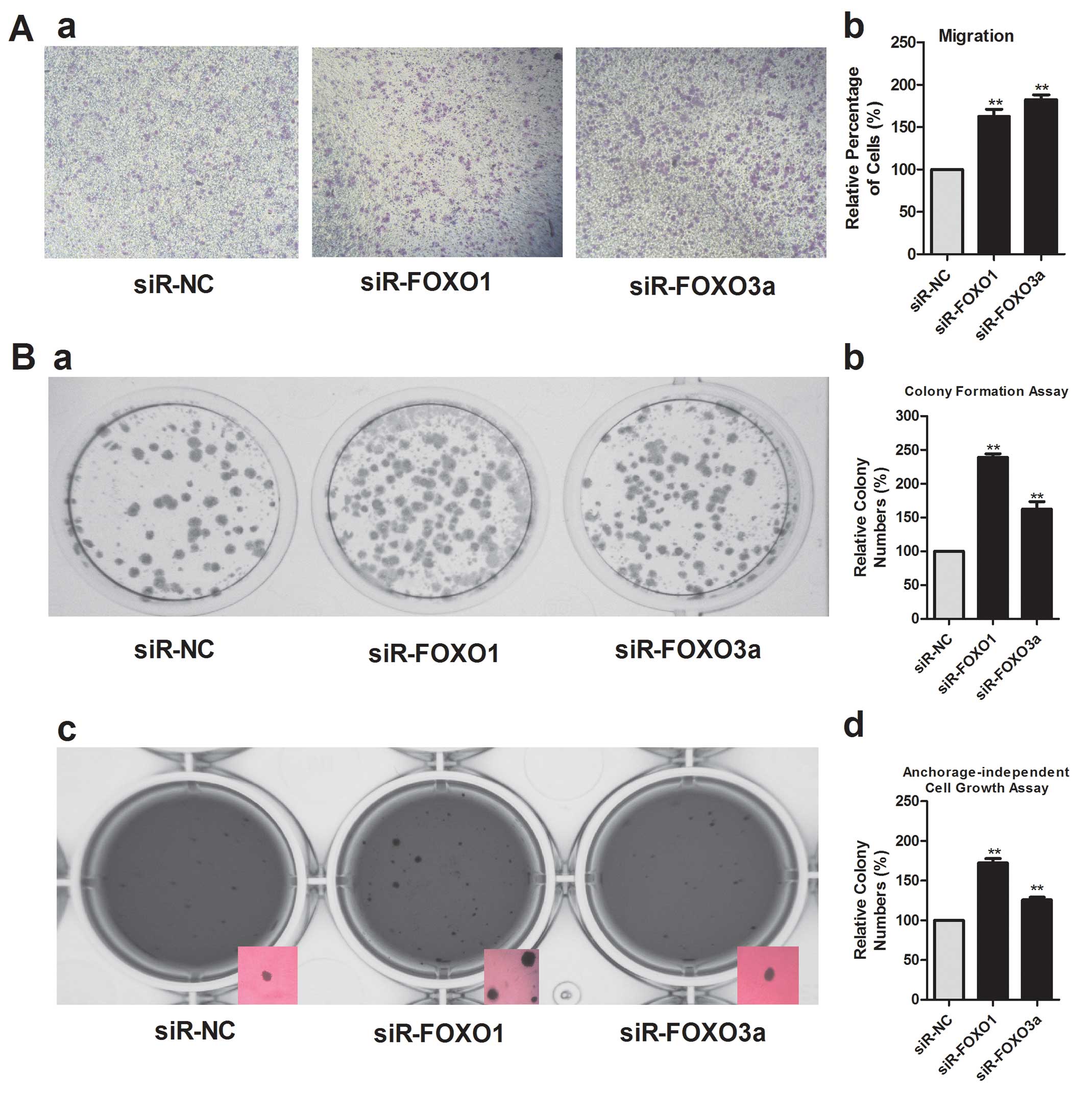
We evaluated the effects of FOXO1 and FOXO3a on the
clonogenicity of HepG2 cells by using a colony formation assay.
Analysis of colony formation showed that downregulation of the
expression of FOXO1 and FOXO3a using siRNAs resulted in the
formation of much fewer and smaller colonies than in control
transfectants (Fig. 4B). Inhibition
of FOXO1 and FOXO3a expression also promoted the
anchorage-independent growth of HepG2 cells significantly, as shown
by the increase in colony number and size (Fig. 4B).
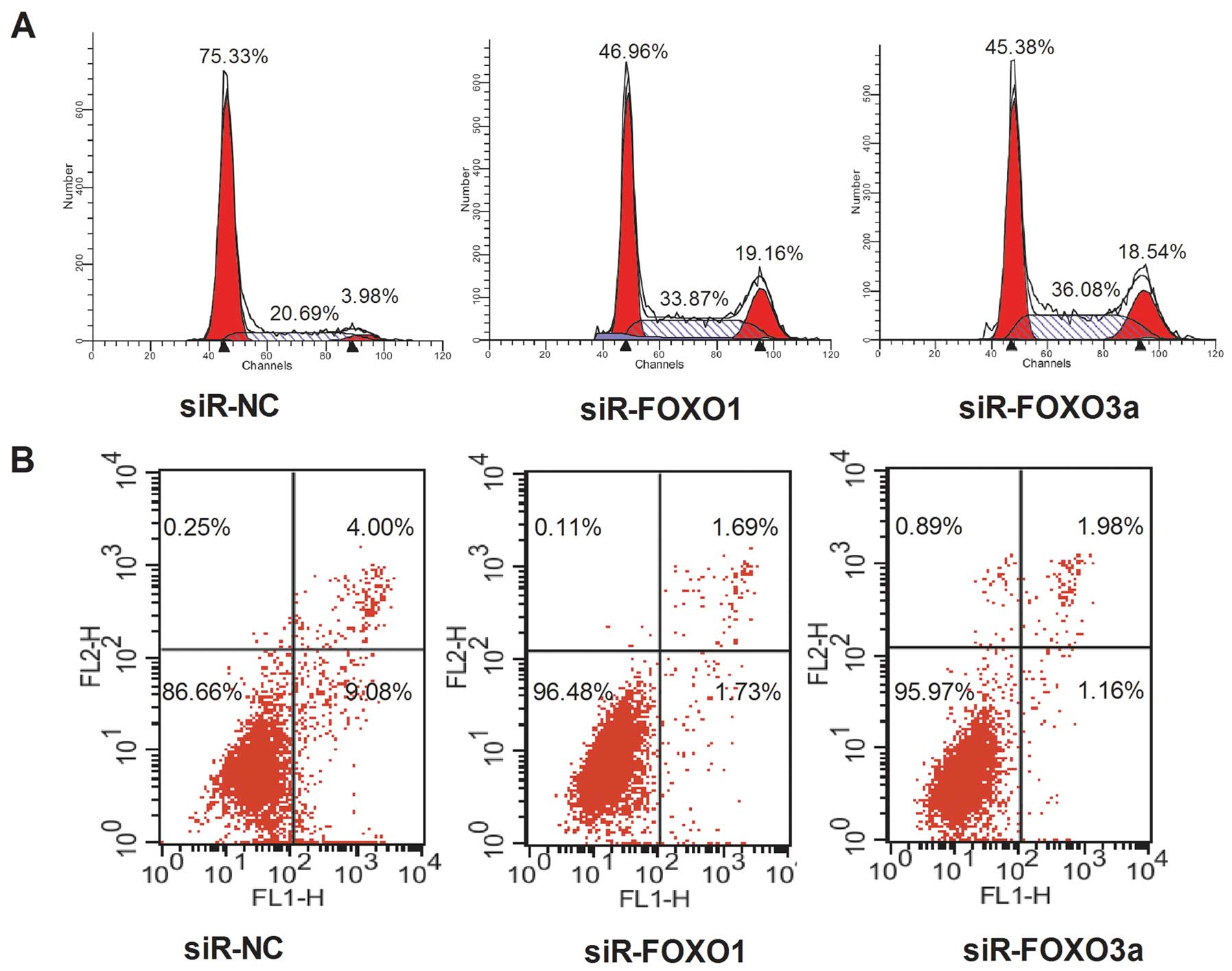
FOXO1 and FOXO3a affect cell cycle and
apoptosis of HepG2 cells
The downregulation of FOXO3a or FOXO1 expression by
transfection with siRNAs altered cell cycle regulation in HepG2
cells, in that the ratio of G0/G1 phase cells decreased and the
fraction of S-phase cells significantly increased, compared with
the cells treated with siR-NC at 72 h (Fig. 5A). Compared to the negative control,
inhibition of FOXO1 and FOXO3a induced less apoptosis (Fig. 5B) in HepG2 cells.
Discussion
miRNAs are involved in critical biological
processes, including development, differentiation, apoptosis, and
proliferation, through imperfect pairing with target messenger RNAs
of protein-coding genes and transcriptional or post-transcriptional
regulation of their expression. Numerous studies have reported that
miRNAs are involved in the development and progression of various
types of human cancer and proposed a role for miRNAs as potential
novel targets for anti-cancer therapies (18–20).
The genesis and growth of HCC is intricately and multifactorially
regulated, and miRNAs may be involved in this process. Several
studies have reported miRNA expression profiling in human HCC cells
and tissues and in experimental models of HCC (21–23).
Consistently, these studies have all shown that specific miRNAs are
abnormally expressed in malignant HCC cells or tissues compared to
non-malignant hepatocytes or tissue. miR-96 is among the miRNAs
that are abnormally increased in HCC compared to non-tumoral tissue
(12).
Our present study shows that miR-96 expression was
markedly upregulated in liver cancer cells, compared with that in
normal liver cells, consistent with results described in a previous
report (12). miR-96 may be
associated with HCC and resides on chromosome 7q32.2 in the
intergenic region between two protein-coding genes. Previously,
miR-96 was identified as an HBV infection-related miRNA (24). miR-96 was also found to be
upregulated in colorectal cancer (25). Furthermore, a previous report
demonstrated that the level of miR-96 was elevated in chronic
myelocytic leukemia (CML) patients compared to the levels in
healthy donor samples, indicating a higher possibility of the
involvement of miR-96 in CML pathogenesis (26). In bladder urothelial carcinoma,
miR-96 was the most significantly upregulated miRNA (27). Using DNA microarrays, it was
demonstrated that miR-96 was significantly and consistently
upregulated in non-small cell lung carcinoma cells (28).
The malignant potential of tumor cells is reflected
by their increased proliferation, migration and colony formation
abilities. We found that miR-96 promotes the proliferation and
migration of HCC cells. Moreover, our results revealed that
colonies formed in soft agar were markedly fewer and smaller when
cells were treated with an inhibitor of miR-96, compared to those
formed by cells treated with a negative control. A similar result
was observed in the colony formation assay. These data indicate
that the malignant potential of HepG2 cells is strengthened by
miR-96.
miRNAs play important regulatory roles in animals
and plants by targeting mRNAs for cleavage or translational
repression. To explore the mechanism by which miR-96 suppressed the
proliferation, migration, and clonogenicity of HCC cells, we used a
bioinformatics analysis to search for genes regulated by miR-96,
using TargetScan (http://www.targetscan.org/) (29) and found that miR-96 might target
FOXO1 and FOXO3a, which are members of the FOXO subfamily. In our
study, we demonstrated that with the inhibition of miR-96, the
expression of FOXO1 and FOXO3a is upregulated. The FOXO subfamily
of Forkhead transcription factors, including FOXO1 (FKHR), FOXO3a
(FKHRL1), FOXO4 (AFX) and FOXO6, contains evolutionarily conserved
transcriptional activators that are characterized by a highly
conserved Forkhead domain with a DNA-binding motif (13). FOXO proteins play pivotal roles in
biological processes, such as apoptosis, cell cycle control,
differentiation, stress response, DNA damage repair, and glucose
metabolism (30). FOXO
transcription factors are considered key tumor suppressors.
The TargetScan program predicted that FOXO1 has the
highest context score (an index for the strength of the bond
between mature miRNA and target sites) to miR-96. FOXO1 could be
the most important HCC-related target gene of miR-96. We
demonstrated that inhibition of miR-96 in liver cancer cells led to
the upregulation of FOXO1, indicating that miR-96 can inhibit the
expression of FOXO1. However, whether miR-96 can directly target
the 3′-UTR of FOXO1 mRNA remains to be confirmed in future
studies.
FOXO3a is also known to control cellular
differentiation, DNA repair, longevity, as well as oxidative
stress. FOXO3a exerts its apoptotic effect by transactivating
apoptotic factors, such as bim (a pro-apoptotic member of the Bcl-2
protein family) and Fas ligand (FasL) (31). In breast cancer cells, miR-96 was
shown to likely promote breast cancer proliferation by directly
targeting the 3′-UTR of the FOXO3a mRNA, consequently reducing the
expression of cyclin-dependent kinase (CDK) inhibitors, and
upregulating the cell cycle regulator cyclin D1 (11). In HCC cells, we demonstrated that
with the reduction of miR-96, FOXO3a was upregulated. These data
suggest that in HCC cells miR-96 might directly target the 3′-UTR
of FOXO3a mRNA and lead to the downregulation of FOXO3a.
Downregulation of FOXO1 in chicken embryo
fibroblasts or inhibition of the transcriptional activity of FOXO3a
protein in human breast cancer cells promotes cell transformation
and tumor progression (10,32). In human breast cancer cells,
inhibition of miR-96 results in downregulation of FOXO1 and FOXO3a,
and thus induces cell proliferation (9,11). Our
study showed that the inhibition of FOXO1 and FOXO3a caused
increased cell proliferation, migration, and colony formation of
HepG2 cells. The inhibition of FOXO1 and FOXO3a also caused
retention of more cells in the S-phase of the cell cycle and
reduction of apoptosis. Thus, miR-96 might modulate the
proliferation and colony formation of HCC cells through FOXO1 and
FOXO3a.
Although miR-96 can downregulate the expression of
FOXO1 and FOXO3a, inhibition of miR-96 caused significant cell
cycle arrest, but did not induce significant apoptosis. The
mechanism underlying this process remains unknown and requires
further exploration.
In conclusion, miR-96 suppresses the expression of
FOXO1 and FOXO3a and promotes the proliferation, migration, and
clonogenicity of liver cancer cells. Further research is needed to
explore the precise molecular mechanism underlying this process.
However, our findings suggest that upregulation of miR-96 may play
an important role in promoting carcinogenesis and progression of
liver cancer.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China, no. 81101824 (X.H.) and
81000874 (C.X.).
References
|
1
|
Srivatanakul P, Sriplung H and Deerasamee
S: Epidemiology of liver cancer: an overview. Asian Pac J Cancer
Prev. 5:118–125. 2004.
|
|
2
|
Lavanchy D: Hepatitis B virus
epidemiology, disease burden, treatment, and current and emerging
prevention and control measures. J Viral Hepat. 11:97–107. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Serag HB and Mason AC: Risk factors for
the rising rates of primary liver cancer in the United States. Arch
Intern Med. 160:3227–3230. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baan R, Straif K, Grosse Y, et al:
Carcinogenicity of alcoholic beverages. Lancet Oncol. 8:292–293.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Girard M, Jacquemin E, Munnich A, Lyonnet
S and Henrion-Caude A: miR-122, a paradigm for the role of
microRNAs in the liver. J Hepatol. 48:648–656. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
O’Connell RM, Rao DS, Chaudhuri AA and
Baltimore D: Physiological and pathological roles for microRNAs in
the immune system. Nat Rev Immunol. 10:111–122. 2010.PubMed/NCBI
|
|
7
|
Skaftnesmo KO, Prestegarden L, Micklem DR
and Lorens JB: microRNAs in tumorigenesis. Curr Pharm Biotechnol.
8:320–325. 2007. View Article : Google Scholar
|
|
8
|
Calin GA, Sevignani C, Dumitru CD, et al:
Human microRNA genes are frequently located at fragile sites and
genomic regions involved in cancers. Proc Natl Acad Sci USA.
101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guttilla IK and White BA: Coordinate
regulation of FOXO1 by miR-27a, miR-96, and miR-182 in breast
cancer cells. J Biol Chem. 284:23204–23216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu MC, Lee DF, Xia W, et al: IkappaB
kinase promotes tumorigenesis through inhibition of forkhead
FOXO3a. Cell. 117:225–237. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin H, Dai T, Xiong H, et al: Unregulated
miR-96 induces cell proliferation in human breast cancer by
downregulating transcriptional factor FOXO3a. PLoS One.
5:e157972010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pineau P, Volinia S, McJunkin K, et al:
miR-221 overexpression contributes to liver tumorigenesis. Proc
Natl Acad Sci USA. 107:264–269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Greer EL and Brunet A: FOXO transcription
factors at the interface between longevity and tumor suppression.
Oncogene. 24:7410–7425. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Myatt SS, Wang J, Monteiro LJ, et al:
Definition of microRNAs that repress expression of the tumor
suppressor gene FOXO1 in endometrial cancer. Cancer Res.
70:367–377. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He XX, Chang Y, Meng FY, et al:
microRNA-375 targets AEG-1 in hepatocellular carcinoma and
suppresses liver cancer cell growth in vitro and in vivo. Oncogene.
31:3357–3369. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Samarin J, Rehm M, Krueger B, Waschke J
and Goppelt-Struebe M: Up-regulation of connective tissue growth
factor in endothelial cells by the microtubule-destabilizing agent
combretastatin A-4. Mol Cancer Res. 7:180–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zrelli H, Matsuoka M, Kitazaki S, Zarrouk
M and Miyazaki H: Hydroxytyrosol reduces intracellular reactive
oxygen species levels in vascular endothelial cells by upregulating
catalase expression through the AMPK-FOXO3a pathway. Eur J
Pharmacol. 660:275–282. 2011. View Article : Google Scholar
|
|
18
|
Gregory RI and Shiekhattar R: microRNA
biogenesis and cancer. Cancer Res. 65:3509–3512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Calin GA and Croce CM: microRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar
|
|
21
|
Murakami Y, Yasuda T, Saigo K, et al:
Comprehensive analysis of microRNA expression patterns in
hepatocellular carcinoma and non-tumorous tissues. Oncogene.
25:2537–2545. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang J, Gusev Y, Aderca I, et al:
Association of microRNA expression in hepatocellular carcinomas
with hepatitis infection, cirrhosis, and patient survival. Clin
Cancer Res. 14:419–427. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang YS, Dai Y, Yu XF, et al: Microarray
analysis of microRNA expression in hepatocellular carcinoma and
non-tumorous tissues without viral hepatitis. J Gastroenterol
Hepatol. 23:87–94. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ladeiro Y, Couchy G, Balabaud C, et al:
microRNA profiling in hepatocellular tumors is associated with
clinical features and oncogene/tumor suppressor gene mutations.
Hepatology. 47:1955–1963. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bandres E, Cubedo E, Agirre X, et al:
Identification by Real-time PCR of 13 mature microRNAs
differentially expressed in colorectal cancer and non-tumoral
tissues. Mol Cancer. 5:292006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Agirre X, Jimenez-Velasco A, San
Jose-Eneriz E, et al: Down-regulation of hsa-miR-10a in chronic
myeloid leukemia CD34+ cells increases USF2-mediated
cell growth. Mol Cancer Res. 6:1830–1840. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han Y, Chen J, Zhao X, et al: microRNA
expression signatures of bladder cancer revealed by deep
sequencing. PLoS One. 6:e182862011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Derynck R, Akhurst RJ and Balmain A:
TGF-beta signaling in tumor suppression and cancer progression. Nat
Genet. 29:117–129. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang H and Tindall DJ: Dynamic FoxO
transcription factors. J Cell Sci. 120:2479–2487. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang JY, Xia W and Hu MC: Ionizing
radiation activates expression of FOXO3a, Fas ligand, and Bim, and
induces cell apoptosis. Int J Oncol. 29:643–648. 2006.PubMed/NCBI
|
|
32
|
Aoki M, Jiang H and Vogt PK: Proteasomal
degradation of the FoxO1 transcriptional regulator in cells
transformed by the P3k and Akt oncoproteins. Proc Natl Acad Sci
USA. 101:13613–13617. 2004. View Article : Google Scholar : PubMed/NCBI
|