Introduction
Gliomas are the most common form of primary human
brain tumors, and the incidence rate is approximately 40–50% among
all brain tumors. Malignant gliomas are associated with a poorly
prognosis and survival rate because of their highly invasive
behavior, which results in incomplete tumor resection and a major
obstacle to any therapeutic remedies including surgery,
radiotherapy and chemotherapy (1).
In recent years molecular-targeted therapies are a new strategy for
the treatment of gliomas to block cell signaling that governs cell
growth, survival and migration (2).
Epidermal growth factor receptor (EGFR) gene amplification and
overexpression are a striking feature of high-grade gliomas
(3). EGFR inhibitors were found to
promote significant clinical response in a subset of malignant
glioma patients (4,5).
Eps8 is a substrate of both the receptor and
non-receptor tyrosine kinases (6,7). Eps8
increased the mitogenic response to EGF and contributes to
EGF-dependent cellular tranformation (6,8,9). Eps8,
Abi-1 and Sos1 form a tricomplex and induce Rac-specific guanine
nucleotide exchange factor (GEF) activity, and enhance
Rac-dependent actin remodeling (10–12).
Further studies showed that the C-terminal effector region of Eps8
binds with F-actin and localizes to main sites of actin remodeling,
such as lamellipodia, filopodia and membrane ruffles (10,11).
Recently, many studies have shown that Eps8 functions in various
solid tumors, including squamous cell carcinoma, pancreatic cancer
and cervical cancer, as well as colon cancer and pituitary tumors
(13–18). Moreover, high expression and
concomitant tyrosine phosphorylation of Eps8 were detected in tumor
cell lines (9). Eps8 increases cell
growth and motility, by the upregulation of FOXM1, FAK, MMP-9, ERK,
Akt, cyclins D1, D3 and E, and the downregulation of P53 and
p21Waf1/Cip1 in cancer cells (13–15,17,18).
Additionally, the Sp1 inhibitor mithramycin downregulates Eps8
expression and inhibits human epithelial carcinoma cell
proliferation and migration, which clearly suggests the Eps8
knockdown may exert an anticancer ability (19).
Although recent studies imply a critical role for
Eps8 in the development of many malignancies, the expression and
function of Eps8 in gliomas have not been fully elucidated. In this
study, we attempted to investigate the expression of Eps8 in glioma
tissues and cell lines, as well as the role of Eps8 in glioma
malignancy, including cell proliferation and survival.
Overexpression of Eps8 significantly enhanced the growth of the
glioma cells, whereas inhibition of Eps8 by a lentiviral RNAi
system had an opposite effect. Our results demonstrated that Eps8
serves as a novel growth regulator in glioma cells and could be a
novel target for the diagnosis and treatment of gliomas.
Materials and methods
Immunohistochemistry
A total of 53 brain tumors including 10 WHO grade
III and 43 WHO grade IV gliomas was examined. Patient
characteristics are listed in Table
I. Seven adjacent normal human brain tissues were used as the
control. The study was approved by the Hunan Normal University
Human Ethics Committee, and informed consent was obtained from all
patients. The immunohistochemistry was analyzed on formalin-fixed
and paraffin-embedded samples. Endogenous peroxidase was blocked
with 3% H2O2 in methanol for 10 min. The
mouse monoclonal anti-Eps8 antibody (BD Biosciences, CA, USA) at a
1:500 dilution or mouse control IgG was added at 4°C overnight. The
sections were incubated with HRP-conjugated goat anti-mouse
secondary antibody (Sigma-Aldrich Corp., St. Louis, MO, USA) for 30
min and then with 3,3-diaminobenzidine
(DAB)/H2O2 for 5 min. The sections were
counterstained with hematoxylin, mounted and photographed using an
optical microscope. The percentage of tumor cells stained was
scored as: 0 (no cell staining), + or 1 (≤30%), ++ or 2 (31–60%),
and +++ or 3 (61–100%). Some samples with a staining between two
score values were given 0.5.
 | Table IPatient characteristics. |
Table I
Patient characteristics.
| No. of patients
(%) |
|---|
| Total number | 53 |
| Age, range in years
(average) | 30–65 (75) |
| Gender |
| Male | 39 (73.6) |
| Female | 14 (26.4) |
| Grade of
glioma |
| III | 10 (18.9) |
| IV | 43 (81.1) |
| Normal tissue | 7 |
Cell culture
The human neuroblastoma cell line SH-SY5Y was
cultured in a 1:1 mixture of F12 and Eagle’s minimum essential
medium (EMEM) supplemented with non-essential amino acids,
glutamine, sodium pyruvate, penicillin, streptomycin, and 10% fetal
bovine serum (FBS). HEK293FT and 293T cells, as well as human
glioma A172, U251, and SHG-44 cells were cultured in complete
Dulbecco’s modified Eagle’s medium (DMEM) containing penicillin,
streptomycin and 10% fetal bovine serum. Cells were maintained in a
humidified atmosphere of 5% CO2 and 95% air at 37°C.
Plasmid constructs and establishment of
U251 cell lines that express GFP and GFP-Eps8
To generate the pEGFP-C3-Eps8 plasmid, full-length
human Eps8 was released by EcoRI and XhoI digestion
of a Myc-tagged human Eps8 expression vector pCMV-Myc-Eps8
(20) and subcloned into the
EcoRI and SalI sites of vector pEGFP-C3. U251 cells
were transfected with pEGFP-C3-Eps8 or pEGFP-C3 control plasmids
using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA). Stably
transfected cells were isolated by G418 screening (21,22).
The transfection efficiency was detected using a fluorescence
microscope (Zeiss Axioskop 2, LLC, USA).
Western blotting
Samples were denatured in sample buffer [2% sodium
dodecyl sulfate (SDS), 10% glycerol, 2.5% β-mercaptoethanol, and
bromophenyl blue] and heated to 100°C for 5 min before gel
electrophoresis. Samples were then separated on a 10% SDS-PAGE gel
and transferred to a polyvinylidene difluoride (PVDF) membrane. The
blots were probed with rabbit polyclonal anti-GFP antibody, mouse
monoclonal antibodies against β-catenin, β-actin, Akt,
phosphorylated Akt, ERK and phosphorylated ERK (Santa Cruz
Biotechnology, Santa Cruz, CA, USA). HRP-conjugated goat
anti-rabbit and goat anti-mouse secondary antibodies (Sigma) were
used for detection. The signal was visualized with SuperSignal West
Femto Maximum Sensitivity Substrate (Pierce Biotechnology,
Rockford, IL, USA).
Eps8 RNAi lentivirus generation
Four siRNA sequences targeting Eps8 (NM_004447.5)
were from the positions (siRNA1, 93–111; siRNA2, 246–264; siRNA3,
678–696; siRNA4, 1974–1992) relative to the start codon and
synthesized by Shanghai Integrated Biotech Solutions Co. Ltd.,
China. After testing knockdown efficiencies, stem-loop DNA
oligonucleotides were synthesized by Shanghai GeneChem Co. Ltd.
(shRNA1 sense, 5′-CCG GAC GGA CAG AGA ACA TGG TTC ACT CGA GTG AAC
CAT GTT CTC TGT CCG TTT TTT G-3′; antisense, 5′-AAT TCA AAA AAC GGA
CAG AGA ACA TGG TTC ACT CGA GTG AAC CAT GTT CTC TGT CCG T-3′;
shRNA2 sense, 5′-CCG GAC CAC TGT TGA TGA TGG AAT ACT CGA G TA TTC
CAT CAT CAA CAG TGG TTT TTG-3′; antisense, 5′-AAT TCA AAA AAC CAC
TGT TGA TGA TGG AAT ACT CGA GTA TTC CAT CAT CAA CAG TGG T-3′) and
cloned into the lentivirus-based RNAi vector pGCSIL-GFP. A
nontargeting stem-loop DNA was also inserted into the pGCSIL-GFP
vector as a negative control (NC). Lentiviral particles were
prepared as described previously (23). SHG-44 cells were infected with
Eps8-RNAi-lentivirus or NC-GFP-lentivirus and detected on day 6 by
a fluorescence microscope and western blot analysis.
Cell proliferation assays
For cell survival assays, 5,000 cells stably
expressing GFP-Eps8 or GFP and parental U251 cells were plated in
triplicate in 6-well plates in complete medium containing 400 μg/ml
G418. For SHG-44 cells, 5,000 cells expressing Eps8-RNAi-LV or
NC-GFP-LV were plated in triplicate in 6-well plates. After 8–10
days, cell numbers were counted with a hemocytometer after trypan
blue staining of viable cells.
For the cell viability assay, 3,000 cells were
seeded in octuplicate in 96-well plates. On days 1, 4, 8 and 10 or
days 1, 3, 5 and 8, cells were treated with 1 mg/ml
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenytetrazolium bromide (MTT)
at 37°C for 4 h. Then 100 μl dimethylsulfoxide (DMSO) was added to
the medium to dissolve the formazan crystals. The absorbency value
at 490 nm in each well was obtained using a spectrophotometer
(UV-2102C, Changsha, China).
The liquid colony formation was carried out. 1,000
cells were seeded in triplicate in 6-well plates, and cells were
grown for 2 weeks in a humidified incubator at 37°C with 5%
CO2. Colonies were fixed with methanol, stained with
Giemsa (BBI International, Cardiff, UK) and photographed with a
digital camera (Olympus, Tokyo, Japan). Only colonies containing
over 30 cells were counted.
Statistical analysis
Data are presented as the means ± SD from at least
three independent experiments. The significance of difference was
assessed using the Student’s t-test. Values of p<0.05 were
considered statistically significant.
Results
Expression of Eps8 in glioma tissues and
cell lines
The expression level of Eps8 was examined in 10 WHO
grade III, 43 WHO grade IV gliomas and 7 adjacent normal human
brain tissues by immunohistochemical analysis using mouse
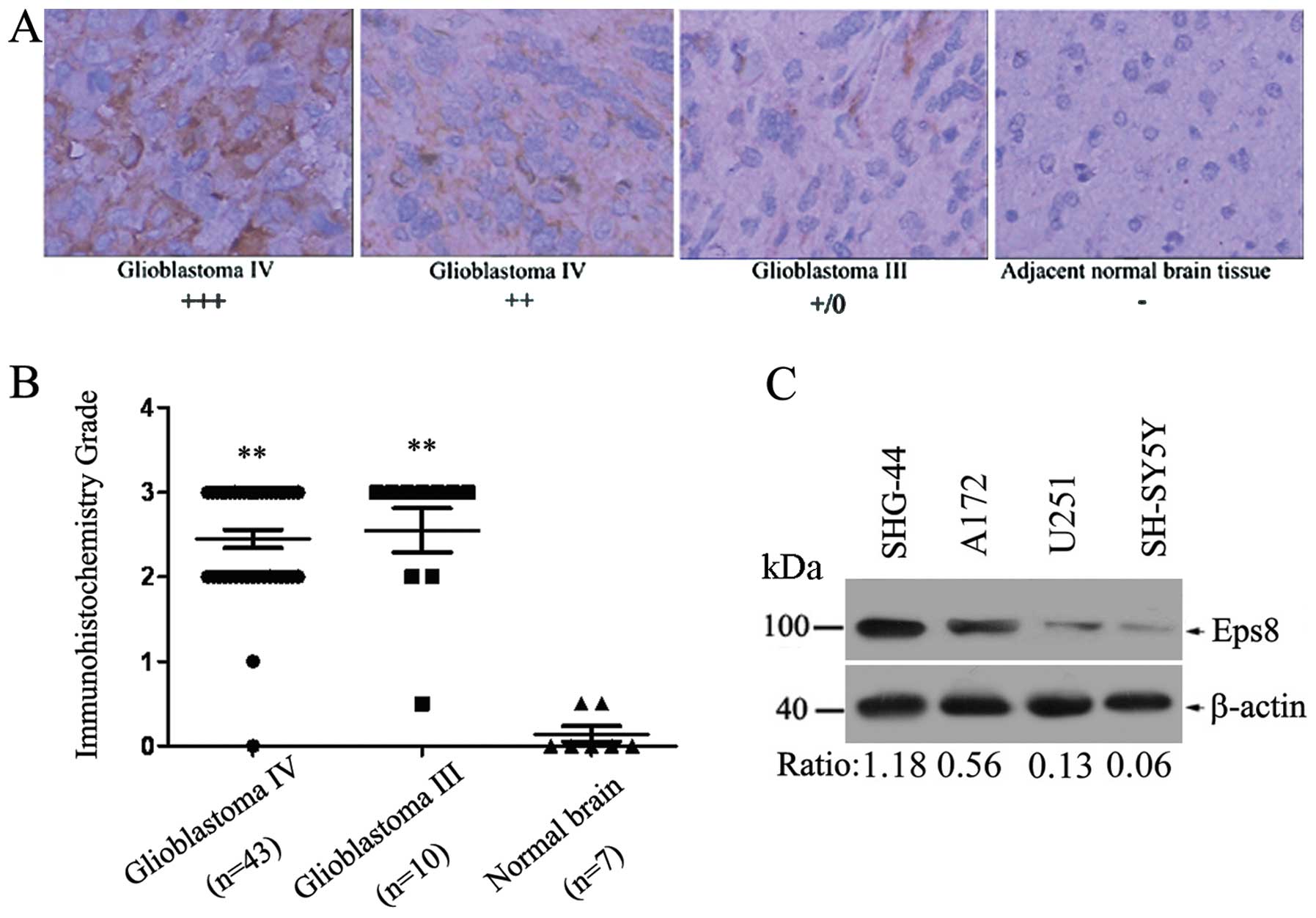
monoclonal Eps8-specific antibody. We found that Eps8 was
completely localized in the cytoplasm (Fig. 1A). Eps8 expression was detected in
30 (56.6%) of the 53 brain tumors with strong staining (3+), 20
(37.7%) of the 53 gliomas were moderately stained (2+), and 3
(5.7%) were weakly stained or negative for Eps8 expression (+/0),
which showed Eps8 was significantly highly expressed in the gliomas
(P<0.001; Student’s t-test). A complete loss of Eps8 expression
was observed in normal brain tissues (Fig. 1B). Therefore, Eps8 expression was
markedly increased in the human glioma tissues when compared with
expression in the normal brain tissues.
We next analyzed the expression of Eps8 protein in
three human glioma cell lines (A172, U251 and SHG-44) and human
neuroblastoma cell line SH-SY5Y. Higher expression of Eps8 protein
was evident in the SHG-44 cells when compared with that in the A172
and U251 cells. SH-SY5Y cells demonstrated a slight level of Eps8
protein (Fig. 1C). Thus, the level
of Eps8 expression differed in the two glioma cell lines. U251 and
SHG-44 cell lines were further selected to overexpress and
knockdown Eps8 protein, respectively.
Eps8 overexpression enhances the
proliferation of the U251 human glioma cell line
To further investigate the role of Eps8 in human
glioma cells, we constructed the pEGFP-C3/Eps8 plasmid and
established U251 cell lines stably overexpressing GFP/Eps8 and GFP
by G418 screening (Fig. 2A). The
level of Eps8 overexpression was demonstrated by western blotting.
As shown in Fig. 2B, U251-GFP and
U251-GFP/Eps8 cells were found to express the target proteins at
high levels.
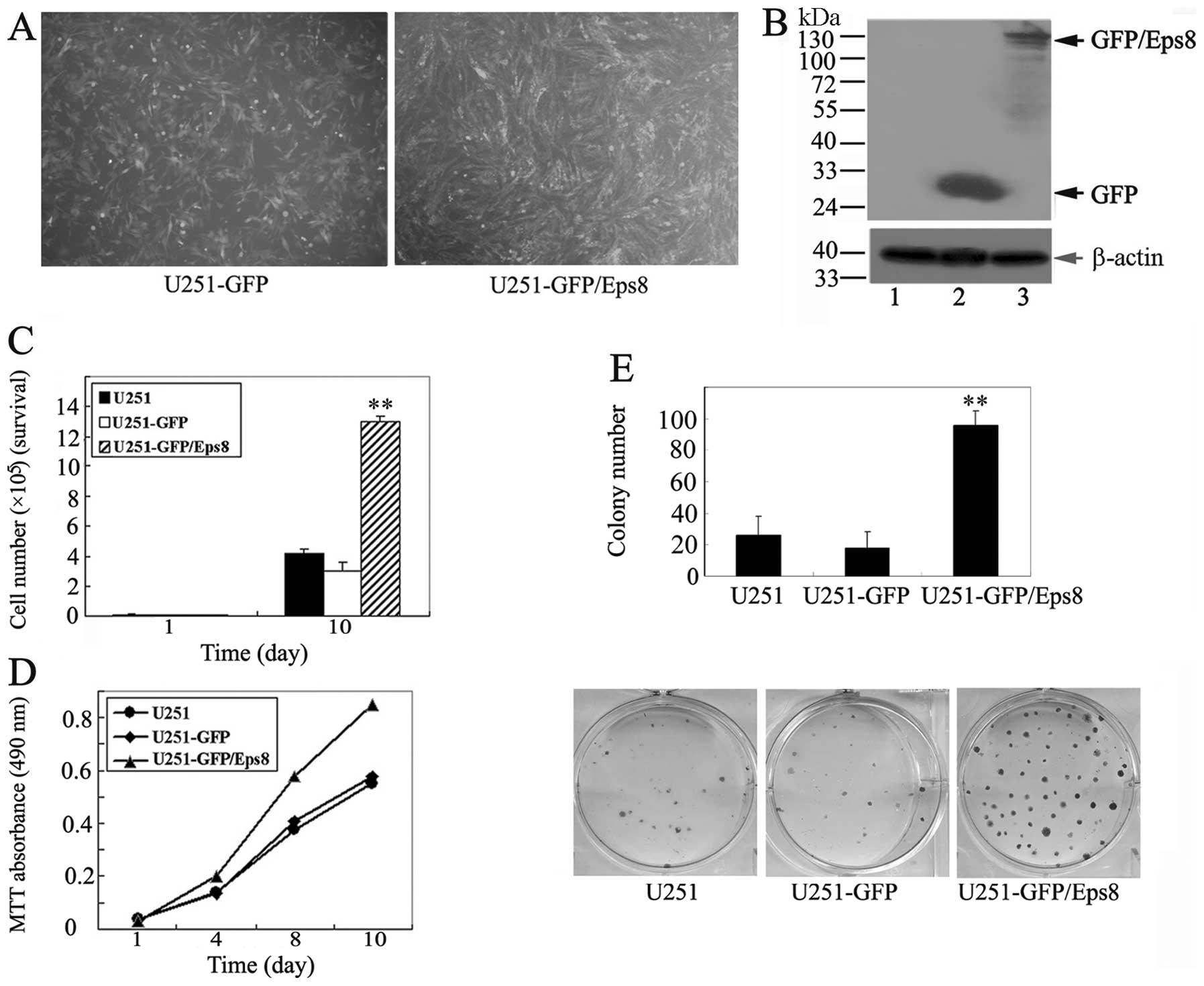 | Figure 2Effects of Eps8 overexpression on U251
glioma cell survival and proliferation. (A) Immunofluorescence
assay confirming the establishment of U251 cells stably expressing
GFP/Eps8 and GFP. (B) Western blot assay of GFP and GFP/Eps8
expression in U251 cells using anti-GFP antibodies. β-actin was
served as a loading control. Lane 1, U251 cells; lane 2,
GFP-transfected U251 cells; lane 3, GFP/Eps8-transfected U251
cells. (C) Cell survival assay of GFP/Eps8-transfected U251 cells,
GFP-transfected U251 cells and parental U251 cells. Cells (5,000)
were plated into 6-well plates in triplicate under the same
condition, grown in RPMI-1640 with 10% FBS for 10 days and stained
with trypan blue in PBS, and viable cells were counted. (D) MTT
assays of mock or transfected U251 cells. Cells (3,000) were plated
in octuplicate in 96-well plates and grown in RPMI-1640 with 10%
FBS. The absorbance was analyzed following 1, 4, 8 and 10 days. (E)
Liquid colony formation analysis of mock or transfected U251 cells.
Cells (1,000) were seeded in triplicate in 6-well plates, and grown
for 2 weeks. Colonies were fixed with methanol, stained with
Giemsa, images (lower panel) were captured and the number of
colonies (upper panel) were counted. These data represent at least
three independent experiments with similar results.
**p<0.01, compared with parental and control
cells. |
We next examined whether Eps8 overexpression in U251
glioma cells promotes cell proliferation. Cells were plated in
triplicate in 6-well plates, and the cell number was counted on day
10. We found that the overexpression of Eps8 in U251 cells resulted
in an increased cellular growth compared with the controls
(Fig. 2C). Similarly, we tested the
cell viability using MTT assay, Eps8 overexpression resulted in a
striking increase in the number of viable cells (Fig. 2D). Furthermore, the liquid colony
formation assay demonstrated a great increase in the numbers of
colonies of the Eps8-overexpressing U251 cells (Fig. 2E). Taken together, these data
suggest that Eps8 contributes to glioma cell survival and
proliferation in vitro.
Eps8 knockdown antagonizes the
proliferation of the SHG-44 human glioma cell line
The above results indicate that Eps8 significantly
promotes U251 cellular proliferation. To gain further supporting
evidence, we knocked down Eps8 expression using shRNA and
determined whether shRNA-mediated Eps8 knockdown inhibits the
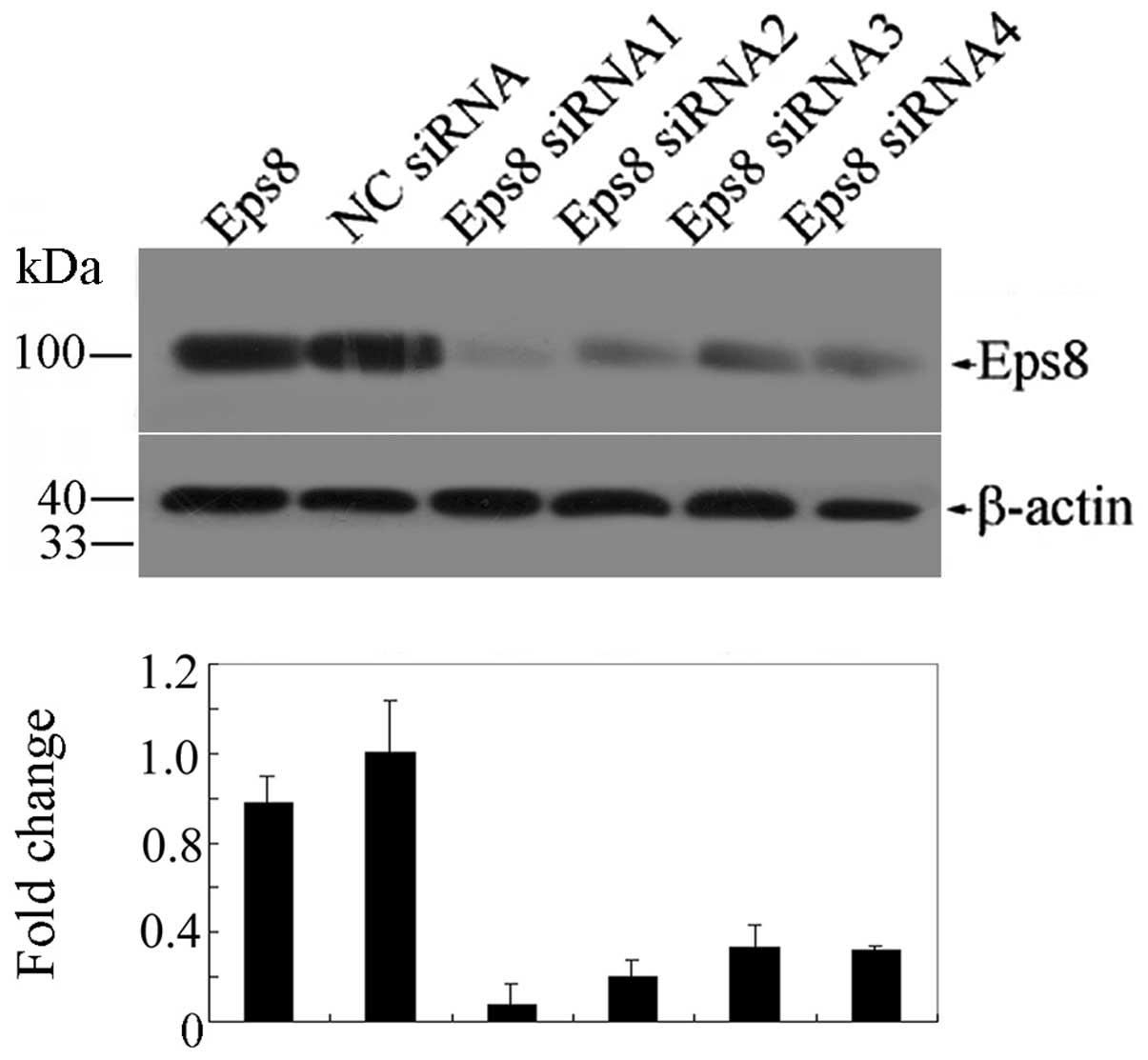
proliferative effects. Four siRNA sequences targeting different
regions of Eps8 mRNA were constructed and their abilities to knock
down Eps8 expression in transfected HEK293FT cells were
demonstrated by western blotting. As shown in Fig. 3, the four sequences knocked down the
Eps8 gene to different levels, especially siRNA1 and siRNA2.
Consequently, siRNA1 and siRNA2 were designed to
construct the lentivirus-based RNAi vector pGCSIL-Eps8. Packaging
plasmids and pGCSIL-Eps8 were cotransfected into the 293T cells.
The fluorescence intensity was markedly increased 4 days after
transfection and the transfection efficiency was nearly 90% in the
SHG-44 cells (Fig. 4A). Meanwhile,
the expression of Eps8 proteins was detected by western blot
analysis. Eps8 shRNA1 and shRNA2 significantly suppressed the
expression of Eps8 protein when compared with the negative control
shRNA (Fig. 4B).
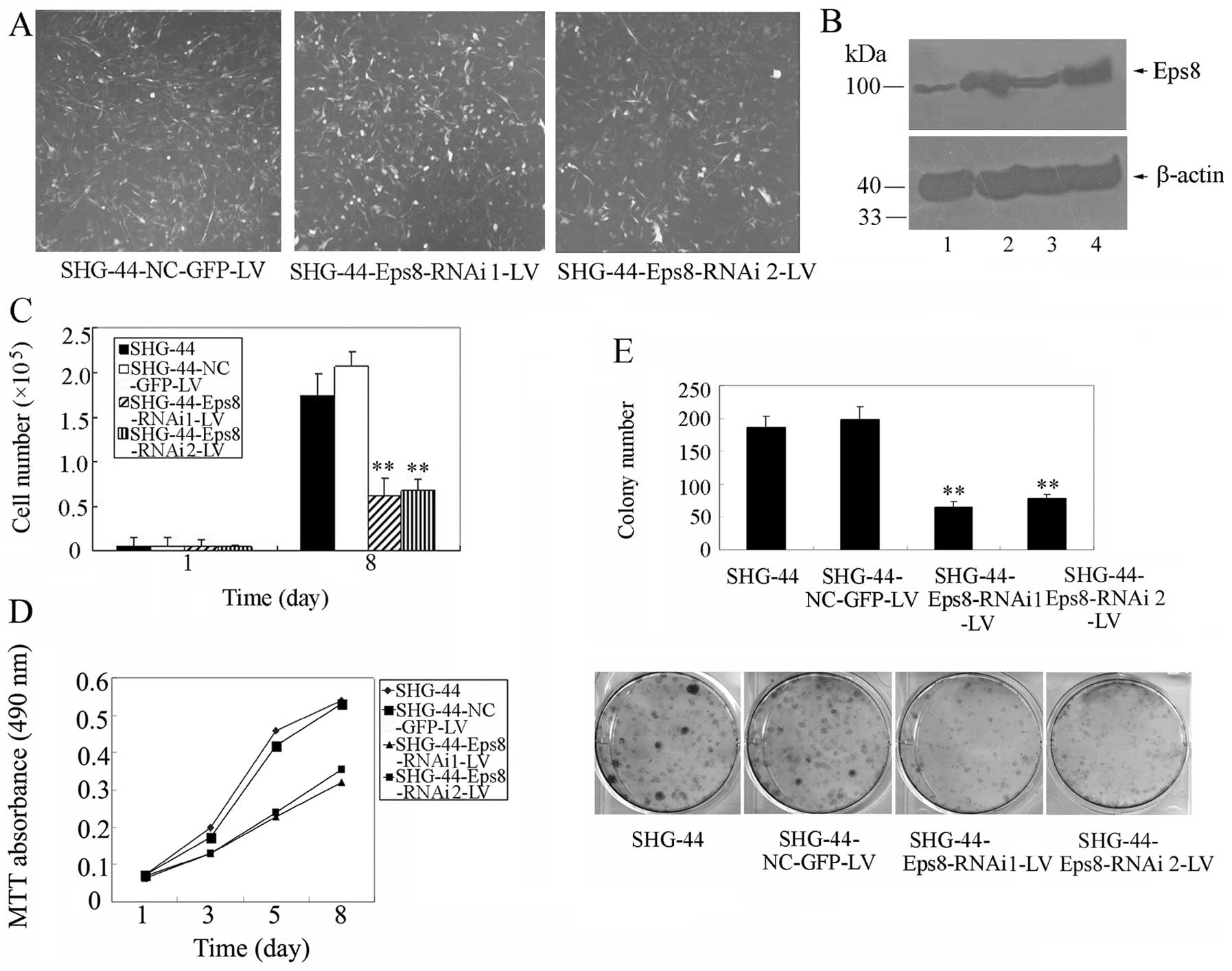 | Figure 4Effects of Eps8 knockdown on SHG-44
glioma cell survival and proliferation. (A) Immunofluorescence
assays for the measurement of the transduction efficiency of SHG-44
cells infected with shRNA1 or shRNA2. (B) The Eps8 knockdown
efficiency was detected by western blot analysis with mouse
monoclonal anti-Eps8 antibody. β-actin was used as a loading
control for total lysate samples. Lane 1, Eps8 shRNA 1-transfected
SHG-44 cells; lane 2, SHG-44 cells; lane 3, Eps8 shRNA2-transfected
SHG-44 cells; lane 4, GFP NC-transfected SHG-44 cells. (C) Cell
survival assay of Eps8 shRNA-transfected SHG-44 cells, GFP
NC-transfected SHG-44 cells and parental SHG-44 cells as described
in Materials and methods. (D) MTT assays of mock or transfected
SHG-44 cells. SHG-44 cells (3,000) were plated and grown, and the
absorbance was analyzed on days 1, 3, 5 and 8. (E) Liquid colony
formation analysis of mock or transfected SHG-44 cells. SHG-44
cells (2,000) were seeded and grown for 2 weeks. Colonies were
fixed with methanol, stained with Giemsa, images (lower panel) were
captured and the number of colonies were counted (upper panel).
These data represent at least three independent experiments with
similar results. **p<0.01, compared with parental and
control cells. |
Given that Eps8 is a critical regulator of glioma
cell proliferation, we examined the effect of Eps8 on SHG-44 glioma
cell proliferation. As shown in Figs.
4C and D, Eps8 knockdown markedly inhibited the proliferation
of SHG-44 cells, whereas control shRNA exhibited no effect. The
colony formation assays showed that deletion of Eps8 expression
produced a significant decrease in the clonogenicity of SHG-44
cells, while control shRNA produced no effect (Fig. 4E). These data clearly indicate that
Eps8 knockdown reduces glioma cell proliferation.
Eps8 affects the expression of
phosphorylated ERK, Akt and β-catenin in glioma cell lines and
tissues
Since Eps8 regulates many genes involved in the
development of human cancers, we determined to investigate whether
Eps8 influences these target genes in glioma cell lines and
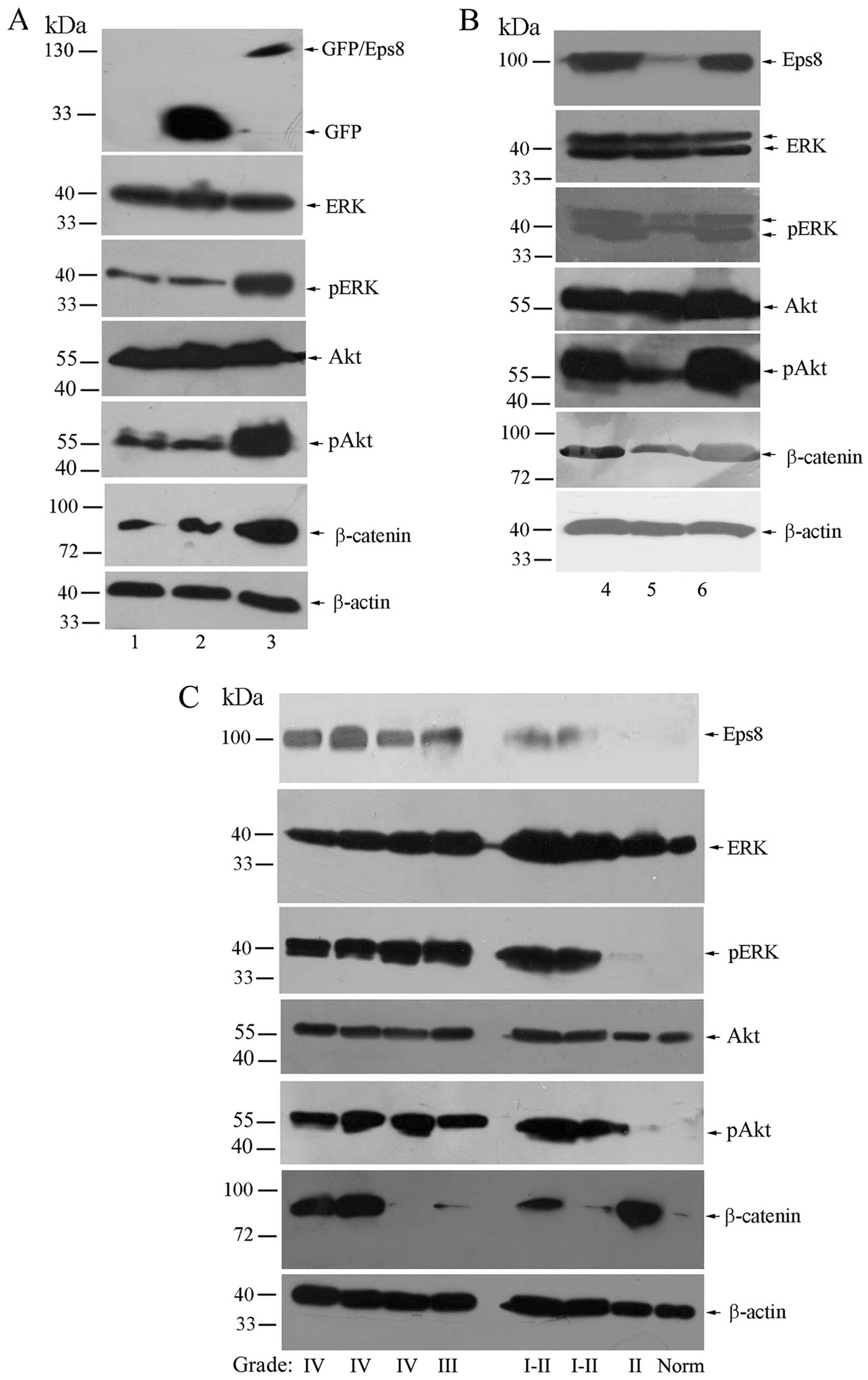
tissues. As shown in Fig. 5A, Eps8
overexpression had no effect on ERK and Akt protein levels, while
it increased the levels of phosphorylated ERK and Akt expression.
Moreover, the level of β-catenin was concomitantly enhanced in
Eps8-transfected cells. Likely, Eps8 knockdown decreased the
expression of phosphorylated ERK, phosphorylated Akt and β-catenin,
but not ERK or Akt (Fig. 5B).
We then analyzed the expression of Eps8 and target
genes in normal brain tissues and gliomas. Eps8 had a significantly
higher expression in high grade gliomas (WHO III-IV) than in low
grade gliomas (WHO I-II) and normal brain tissues. In addition, the
expression of Eps8 revealed broadly paralleled expression of target
genes in the gliomas and normal brain tissues, particularly
phosphorylated ERK and phosphorylated Akt. However, the levels of
β-catenin were partially correlated with Eps8 expression in certain
samples (Fig. 5C). Taken together,
these data show that Eps8 regulates cell proliferation and
survival, at least in part, by affecting phosphorylated ERK and
Akt/β-catenin activities.
Discussion
The human Eps8 family includes Eps8 and three
related proteins Eps8L1, Eps8L2, and Eps8L3. Eps8Ls display 27–42%
identity to Eps8. However, Eps8L2 and Eps8 display the highest
degree of amino acid identity within the family, which explains for
the lack of overt phenotype in Eps8 null mice. Only Eps8 is
specifically expressed in the brain of the 16.5 dpc mouse embryo
among the Eps8 family (24). In
addition, it has also been reported that Eps8 is enriched in the
growth cone and filopodia of developing hippocampal neurons
(25). Thus, Eps8 is likely to
function in the process of neurodevelopment.
Previous data showed that Eps8 is important in
regulating the progression of certain types of human cancers.
However, the role of Eps8 in human gliomas has not been previously
reported. In the present study, we provide evidence concerning the
expression of Eps8 in glioma tissues and cell lines. As assessed by
immunohistochemical analysis, Eps8 expression was detectable in the
majority of the glioma specimens examined. More importantly, Eps8
protein was significantly higher in 56.6% of high grade (III and
IV) glioma tissues compared to normal brain tissues. Accordingly, 3
glioma cell lines were used to demonstrate that the protein level
of Eps8 was upregulated in glioma specimens, which was consistent
with previous evidence that high expression of Eps8 is noted in
tumor cell lines and tissues (9).
Thus, Eps8 expression may clinically designate high grade malignant
gliomas.
The evidence presented here also verified the effect
of Eps8 on glioma cell proliferation. We established a stable cell
line overexpressing Eps8 using U251 cells which have low
constitutive expression of Eps8. We used this cell line to
investigate the effect of Eps8 overexpression on glioma cellular
growth in vitro. Eps8 overexpression markedly enhanced cell
proliferation. Conversely, we constructed lentivirus-based RNAi
vector pGCSIL-Eps8 to knock down Eps8 expression in the SHG-44 cell
line, which exhibits high expression of Eps8. Depletion of Eps8
reversed the growth phenotype of Eps8-overexpressing glioma cells.
Our data strongly indicate that Eps8 expression is critical for the
cell proliferation of malignant gliomas and is possibly involved in
malignant progression in human gliomas.
Eps8, Abi-1 and Sos1 are known to form a tricomplex
and induce Rac-specific guanine nucleotide exchange factor (GEF)
activity, transduce signals from Ras to Rac and enhance
Rac-dependent actin remodeling (10–12).
Additional, Eps8 activates Ras and Raf. Raf then activates MEK,
which in turn activates ERK (26).
ERK is known to be a critical regulator that mainly participates in
the stimulation of cell proliferation and survival (27). We demonstrated that Eps8 plays an
important role in the growth of glioma cells by regulating the
expression of phosphorylated ERK.
β-catenin functions as a core member of canonical
Wnt signaling. When Wnt binds to its receptor, transcription factor
β-catenin accumulates in the cytoplasm and then translocates to the
nucleus where it subsequently transactivates Wnt pathway downstream
targets known to be crucial in cancer develoment (28,29).
β-catenin is an oncogene in a broad range of human cancers
(29,30). For example, overexpression of
β-catenin contributes to glioma progression (31,32).
Eps8 was reported to trigger increased proliferation through
activation of PI3K via Akt (16,33).
Moreover, β-catenin can be transactivated via the PI3K/Akt pathway
(34,35). Therefore, our finding that Eps8
knockdown in glioma cells downregulates the overall level of
phosphorylated Akt and β-catenin and vice versa, suggesting that
Eps8 may effectively regulate Akt/β-catenin signaling in
gliomas.
In summary, this study reveals the biological
significance of Eps8 expression in gliomas. We report that Eps8 is
highly expressed in malignant gliomas, which correlates with glioma
cell proliferation. We found that Eps8 mediates cell proliferation
and survival of glioma cells, at least in part, by affecting
phosphorylated ERK and Akt/β-catenin activities. These data suggest
that Eps8 may be a potential therapeutic target against human
gliomas.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (nos. 30900827, 81071656, 81071696,
81172112), the Opening Fund of the Key Laboratory of Protein
Chemistry and Developmental Biology of the Ministry of Education,
Hunan Normal University.
References
|
1
|
Giese A, Bjerkvig R, Berens ME and
Westphal M: Cost of migration: invasion of malignant gliomas and
implications for treatment. J Clin Oncol. 21:1624–1636. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu TJ, LaFortune T, Honda T, Ohmori O,
Hatakeyama S, Meye T, Jackson D, de Groot J and Yung WK: Inhibition
of both focal adhesion kinase and insulin-like growth factor-I
receptor kinase suppresses glioma proliferation in vitro and in
vivo. Mol Cancer Ther. 6:1357–1367. 2007. View Article : Google Scholar
|
|
3
|
Ekstrand AJ, James CD, Cavenee WK, Seliger
B, Pettersson RF and Collins VP: Genes for epidermal growth factor
receptor, transforming growth factor alpha, and epidermal growth
factor and their expression in human gliomas in vivo. Cancer Res.
51:2164–2172. 1991.PubMed/NCBI
|
|
4
|
Nakamura JL: The epidermal growth factor
receptor in malignant gliomas: pathogenesis and therapeutic
implications. Expert Opin Ther Targets. 11:463–472. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mellinghoff IK, Wang MY, Vivanco I,
Haas-Kogan DA, Zhu S, Dia EQ, Lu KV, Yoshimoto K, Huang JH, Chute
DJ, Riggs BL, Horvath S, Liau LM, Cavenee WK, Rao PN, Beroukhim R,
Peck TC, Lee JC, Sellers WR, Stokoe D, Prados M, Cloughesy TF,
Sawyers CL and Mischel PS: Molecular determinants of the response
of glioblastomas to EGFR kinase inhibitors. N Engl J Med.
353:2012–2024. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fazioli F, Minichiello L, Matoska V,
Castagnino P, Miki T, Wong WT and Di Fiore PP: Eps8, a substrate
for the epidermal growth factor receptor kinase, enhances
EGF-dependent mitogenic signals. EMBO J. 12:3799–3808.
1993.PubMed/NCBI
|
|
7
|
Maa MC, Lai JR, Lin RW and Leu TH:
Enhancement of tyrosyl phosphorylation and protein expression of
eps8 by v-Src. Biochim Biophys Acta. 1450:341–351. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Castagnino P, Biesova Z, Wong WT, Fazioli
F, Gill GN and Di Fiore PP: Direct binding of eps8 to the
juxtamembrane domain of EGFR is phosphotyrosine- and
SH2-independent. Oncogene. 10:723–729. 1995.PubMed/NCBI
|
|
9
|
Matoskova B, Wong WT, Salcini AE, Pelicci
PG and Di Fiore PP: Constitutive phosphorylation of eps8 in tumor
cell lines: relevance to malignant transformation. Mol Cell Biol.
15:3805–3812. 1995.PubMed/NCBI
|
|
10
|
Scita G, Nordstrom J, Carbone R, Tenca P,
Giardina G, Gutkind S, Bjarnegard M, Betsholtz C and Di Fiore PP:
Eps8 and E3B1 transduce signals from Ras to Rac. Nature.
401:290–293. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Scita G, Tenca P, Areces LB, Tocchetti A,
Frittoli E, Giardina G, Ponzanelli I, Sini P, Innocenti M and Di
Fiore PP: An effector region in Eps8 is responsible for the
activation of the Rac-specific GEF activity of Sos-1 and for the
proper localization of the Rac-based actin-polymerizing machine. J
Cell Biol. 154:1031–1044. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen H, Wu X, Pan ZK and Huang S:
Integrity of SOS1/EPS8/ABI1 tri-complex determines ovarian cancer
metastasis. Cancer Res. 70:9979–9990. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang H, Teh MT, Ji Y, Patel V,
Firouzabadian S, Patel AA, Gutkind JS and Yeudall WA: EPS8
upregulates FOXM1 expression, enhancing cell growth and motility.
Carcinogenesis. 31:1132–1141. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen YJ, Shen MR, Chen YJ, Maa MC and Leu
TH: Eps8 decreases chemosensitivity and affects survival of
cervical cancer patients. Mol Cancer Ther. 7:1376–1385. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Maa MC, Lee JC, Chen YJ, Chen YJ, Lee YC,
Wang ST, Huang CC, Chow NH and Leu TH: Eps8 facilitates cellular
growth and motility of colon cancer cells by increasing the
expression and activity of focal adhesion kinase. J Biol Chem.
282:19399–19409. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang H, Patel V, Miyazaki H, Gutkind JS
and Yeudall WA: Role for EPS8 in squamous carcinogenesis.
Carcinogenesis. 30:165–174. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang W, Wang L, Liu Y, Xu J, Zhu G, Cang
H, Li X, Bartlam M, Hensley K, Li G, Rao Z and Zhang XC: Structure
of human lanthionine synthetase C-like protein 1 and its
interaction with Eps8 and glutathione. Genes Dev. 23:1387–1392.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Welsch T, Endlich K, Giese T, Buchler MW
and Schmidt J: Eps8 is increased in pancreatic cancer and required
for dynamic actin-based cell protrusions and intercellular
cytoskeletal organization. Cancer Lett. 255:205–218. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang TP, Chiou HL, Maa MC and Wang CJ:
Mithramycin inhibits human epithelial carcinoma cell proliferation
and migration involving downregulation of Eps8 expression. Chem
Biol Interact. 183:181–186. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Luo C, Ding XF, Sun YB and Han M:
Subcellular location of EPS8 by its expression and preparation of
antiserum. J Nat Sci Hunan Normal University. 31:100–104. 2008.
|
|
21
|
Mangravite LM, Lipschutz JH, Mostov KE and
Giacomini KM: Localization of GFP-tagged concentrative nucleoside
transporters in a renal polarized epithelial cell line. Am J
Physiol Renal Physiol. 280:F879–F885. 2001.PubMed/NCBI
|
|
22
|
Schmid JA, Birbach A, Hofer-Warbinek R,
Pengg M, Burner U, Furtmuller PG, Binder BR and de Martin R:
Dynamics of NF kappa B and Ikappa Balpha studied with green
fluorescent protein (GFP) fusion proteins. Investigation of GFP-p65
binding to DNa by fluorescence resonance energy transfer. J Biol
Chem. 275:17035–17042. 2000. View Article : Google Scholar
|
|
23
|
Lois C, Hong EJ, Pease S, Brown EJ and
Baltimore D: Germline transmission and tissue-specific expression
of transgenes delivered by lentiviral vectors. Science.
295:868–872. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Offenhauser N, Borgonovo A, Disanza A,
Romano P, Ponzanelli I, Iannolo G, Di Fiore PP and Scita G: The
eps8 family of proteins links growth factor stimulation to actin
reorganization generating functional redundancy in the Ras/Rac
pathway. Mol Biol Cell. 15:91–98. 2004. View Article : Google Scholar
|
|
25
|
Menna E, Disanza A, Cagnoli C, Schenk U,
Gelsomino G, Frittoli E, Hertzog M, Offenhauser N, Sawallisch C,
Kreienkamp HJ, Gertler FB, Di Fiore PP, Scita G and Matteoli M:
Eps8 regulates axonal filopodia in hippocampal neurons in response
to brain-derived neurotrophic factor (BDNF). PLoS Biol.
7:e10001382009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hecquet C, Lefevre G, Valtink M, Engelmann
K and Mascarelli F: Activation and role of MAP kinase-dependent
pathways in retinal pigment epithelial cells: ERK and RPE cell
proliferation. Invest Ophthalmol Vis Sci. 43:3091–3098.
2002.PubMed/NCBI
|
|
27
|
Wortzel I and Seger R: The ERK cascade:
distinct functions within various subcellular organelles. Genes
Cancer. 2:195–209. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Uthoff SM, Eichenberger MR, McAuliffe TL,
Hamilton CJ and Galandiuk S: Wingless-type frizzled protein
receptor signaling and its putative role in human colon cancer. Mol
Carcinog. 31:56–62. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Barker N and Clevers H: Catenins, Wnt
signaling and cancer. Bioessays. 22:961–965. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Polakis P: Wnt signaling and cancer. Genes
Dev. 14:1837–1851. 2000.
|
|
31
|
Liu XWL, Zhao S, Ji X, Luo Y and Ling F:
β-Catenin overexpression in malignant glioma and its role in
proliferation and apoptosis in glioblastoma cells. Med Oncol.
28:608–614. 2011.
|
|
32
|
Zhang JHK, Shi Z, Zou J, Wang Y, Jia Z,
Zhang A, Han L, Yue X, Liu N, Jiang T, You Y, Pu P and Kang C: High
β-catenin/Tcf-4 activity confers glioma progression via direct
regulation of AKT2 gene expression. Neuro Oncol. 13:600–609.
2011.
|
|
33
|
Xu M, Shorts-Cary L, Knox AJ,
Kleinsmidt-DeMasters B, Lillehei K and Wierman ME: Epidermal growth
factor receptor pathway substrate 8 is overexpressed in human
pituitary tumors: role in proliferation and survival.
Endocrinology. 150:2064–2071. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kanneganti M, Mino-Kenudson M and
Mizoguchi E: Animal models of colitis-associated carcinogenesis. J
Biomed Biotechnol. 2011:3426372011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nava P, Koch S, Laukoetter MG, Lee WY,
Kolegraff K, Capaldo CT, Beeman N, Addis C, Gerner-Smidt K,
Neumaier I, Skerra A, Li L, Parkos CA and Nusrat A:
Interferon-gamma regulates intestinal epithelial homeostasis
through converging beta-catenin signaling pathways. Immunity.
32:392–402. 2010. View Article : Google Scholar
|