Introduction
Hepatocellular carcinoma (HCC) is one of the most
deadly malignancies worldwide. Surgical resection, liver
transplantation, percutaneous ethanol injection (PEI) and
radiofrequency ablation (RFA) may offer curative opportunities to
some selected patients (1). Most
HCC patients are diagnosed at an advanced stage. In this case,
localregional therapies such as transarterial chemoembolization
(TACE) and drug-eluting beads (DEB) are the only feasible options
(2). TACE is a standard of care for
patients with intermediate stage disease and includes infusing
chemotherapeutics (doxorubicin, mitomycin C and cisplatin) mixed
with lipiodol via the transarterial route into the tumor (3). With conventional chemoregimens such as
doxorubicin or cisplatin, TACE has shown objective response rates
of 16-61% and prolonged patient survival (1). Nevertheless, HCC is highly resistant
to chemoregimens (4) and the
majority of patients die of relentless disease recurrence. To this
end, optimizing drug delivery methods and enhancing drug efficacy
by targeted medicine may improve clinical outcomes.
The Hippo signaling pathway, which has previously
been discovered in Drosophila, plays an important role in
organ-size control during embryonic development and it is conserved
as a tumor-suppressor pathway in mammals (5). The core components of the Hippo
pathway act in a kinase cascade, in which kinases large tumor
suppressor homolog 1/2 (LATS1/2) are phosphorylated by mammalian
Ste20-like kinase 1/2 (MST1/2), and this phosphorylation is coupled
by salvador homolog 1 (SAV1 or WW45) (6). The cascade promotes phosphorylation
and cytoplasmic retention of the Yes-associated protein (YAP,
YAP1), leading to ubiquitin-mediated degradation of YAP (7) and attenuated ability to enhance
transcription of genes including connective tissue growth factor
(CTGF) (8) and AXL (9). YAP activation can override cell-cell
contact inhibition and promote cellular growth (10), which result in malignant
transformation of mammary cells (11,12)
and hepatocytes (9). A transgenic
mouse model demonstrated that YAP overexpression causes a marked
increase in liver size and eventually liver tumor formation
(13). Furthermore, amplification
of the YAP gene and overexpression of YAP have been found in
HCC (14), lung, colon and breast
cancers (15). Previously, we found
that YAP overexpression is an independent prognostic marker
associated with poor disease-free survival and overall survival in
HCC (16). In this study, we
further examined the role of modifying HCC cell sensitivity to
doxorubicin, one of the frequently used therapeutic agents in
TACE.
Materials and methods
Cell lines
The human HCC cell lines PLC/PRF/5, Hep3B and Huh7
were used as described previously (17) and maintained in DMEM (Invitrogen,
Carlsbad, CA, USA) supplemented with 10% FBS (Invitrogen) at 37°C
in a humidified incubator in a 5% CO2 atmosphere.
Ectopic expression of YAP1 in Huh7 and
Hep3B cells
The Huh7 cell line was transfected with the
full-length YAP1 complementary DNA (NM_006106.3) plasmid
(pcDNA3.1-YAP1) (9) or pcDNA3.1
(Invitrogen) empty vector (control) using Lipofectamine 2000
reagent (Invitrogen), and maintained in complete medium containing
300 μg/ml G418 (Sigma-Aldrich, St. Louis, MO, USA). Two separate
neomycin-resistant clones (Huh7-YAP1a, Huh7-YAP1b) with high
expression levels of YAP were selected for further studies. Hep3B
cells were transiently transfected with the pcDNA3.1-YAP1 or
pcDNA3.1 empty vector using Lipofectamine 2000. Twenty-four hours
post-transfection, Hep3B-YAP1 cells were subjected to a
chemosensitivity assay.
Transient knockdown of YAP1 in HCC
cells
YAP1 Stealth RNAi siRNA (catalogue nos. HSS115942
and HSS115944; Invitrogen) and non-targeting siRNA (Silencer
Negative control no. 1siRNA, siCon, Invitrogen) were transfected at
100 pmol into semiconfluent PLC/PRF/5 and Huh7 cells. Cells after
transfection were denoted as PLC-siYAP1a, PLC-siYAP1b, PLC-siCon,
Huh7-siYAP1a, Huh7-siYAP1b and Huh7-siCon, respectively.
MTT assay
Cell viability was determined by 3-(4,
5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay. Cells were seeded at 1×103 cells/well onto
96-well microplates in triplicates followed by exposure to various
concentrations of doxorubicin. To evaluate the involvement of the
phosphatidylinositol 3-kinase (PI3-K)/Akt pathway and
mitogen-activated protein (MAP) kinase pathway, Huh7-YAP1b cells
were pretreated with either the PI3-K inhibitor LY294002 (25 μM;
Sigma-Aldrich) or the MEK1/2 inhibitor U0126 (25 μM; Promega,
Madison, WI, USA) dissolved in dimethyl sulphoxide (DMSO,
Sigma-Aldrich) for 12 h before exposure to 1.5 μg/ml doxorubicin.
At the indicated times, 20 μl of MTT solution (5 mg/ml;
Sigma-Aldrich) was added and incubated for 5 h before the end of
the experiments. At the end of incubation, the culture medium was
removed, and 200 μl DMSO was added to dissolve the MTT formazan
crystals at room temperature. Color development was measured at
OD570 with OD655 used as the reference wavelength in a microplate
reader (model 680; Bio-Rad Laboratories, Carlsbad, CA). Cell
survival of untreated cells was set to 100% and survival of treated
cells was expressed as a percentage relative to this value.
Western blotting
RIPA buffer containing protease inhibitor (Roche
Hong Kong Ltd., Hong Kong) and phosphatase inhibitor (Roche) was
used to extract the total protein from cultured cells. For
SDS-PAGE, 20 μg of protein was electrophoresed onto SDS
polyacrylamide gel, and then electro-transferred onto a PVDF
membrane (0.4 μm; Merck Millipore, Billerica, MA). After blocking,
the membrane was probed with one of the following primary
antibodies at a 1:1000 dilution: rabbit monoclonal antibody against
human p44/42 MAP kinase, rabbit monoclonal antibody against human
phospho-p44/42 MAP kinase (Thr202/Tyr204), rabbit monoclonal
antibody against Akt, rabbit monoclonal antibody against
phosphor-Akt (S473), rabbit polyclonal antibody against cleaved
PARP (Cell Signaling Technology Inc., Danvers, MA, USA), rabbit
polyclonal antibody against human YAP (H-125), mouse monoclonal
antibody against Bcl-xL, mouse monoclonal antibody against Bax
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). Mouse
monoclonal antibody against human β-actin (Sigma-Aldrich) was use
at a 1:5000 dilution as the reference control. HRP-conjugated
antibodies against IgG (Invitrogen) (1:10,000 dilution) were used
as the secondary antibody. Immunoreactivity signals were amplified
using ECL detection reagents (GE Healthcare, Hong Kong). Bcl-xL and
Bax protein expression was measured semi-quantitatively using Image
J software version 1.38 (National Institute of Health). The signal
was converted to ratio by analyzing the protein band intensities of
Bcl-xL relative to Bax after normalization with β-actin within the
same sample.
TUNEL assay
Cell apoptosis was evaluated using
ApopTag® In Situ Apoptosis Detection kits (Millipore).
For induction of apoptosis, Huh7 and PLC/PRF/5 cells grown on
coverslips in 6-well plates were tranfected with siRNA (scramble
siRNA or siYAP1) and cultured in complete medium containing 1 μg/ml
doxorubicin on the next day for a duration of 48 h. Briefly, for
detection of apoptosis, cells were fixed in 1% paraformaldehyde and
then post-fixed in ethanol/acetic acid. After quenching endogenous
peroxidase, the cells were further incubated with equilibration
buffer and TdT enzyme. The reaction was halted by agitation with
stop/wash buffer. After incubation with anti-digoxigenin peroxidase
conjugate, the cells were incubated with peroxidase substrate for
color development and then counterstained with Mayer’s hematoxylin.
Apoptotic cells were examined and counted under the microscope at a
×200 magnification from 3 random fields. The experiments were
repeated twice.
Statistical analysis
The SPSS statistical package for Window version 13
(SPSS, Chicago, IL, USA) was used for data analysis. Independent
Student’s t-test was used to assess the effects of YAP as
determined in the MTT assay, TUNEL assay and by the protein ratios
of Bcl-xL to Bax. P-value <0.05 was considered to indicate a
statistically significant difference.
Results
Overexpression of YAP confers resistance
to doxorubicin in HCC cells
Previously, we demonstrated that YAP has a strong
effect on cell growth (9).
Therefore, in this study we investigated whether YAP-overexpressing
HCC cells could overcome growth-inhibition when challenged with
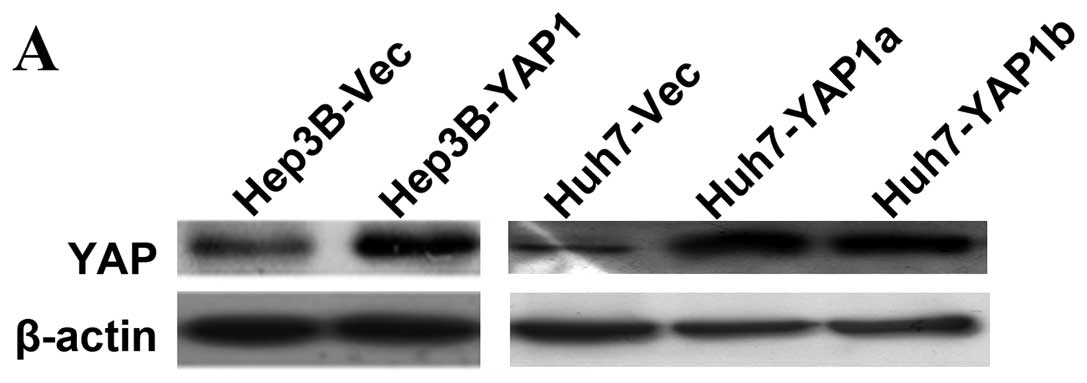
doxorubicin. Western blotting showed YAP was overexpressed in both
stable transfectants (Huh7-YAP1a and Huh7-YAP1b) and transient
transfectants (Hep3B-YAP1, Fig.
1A). Cell viability was determined by MTT assays at 48 and 72 h
after doxorubicin treatment. As shown in Fig. 1B and C, Huh7-YAP1 and Hep3B-YAP1
cells had significantly higher viability than Huh7-Vec and
Hep3B-Vec cells, respectively, at 48 and 72 h when treated with
different concentrations of doxorubicin. The half maximal
inhibitory concentrations (IC50) at 48 h increased from
0.83 μg/ml and 1.14 μg/ml in Huh7-vec and Hep3B-Vec cells to 1.02
μg/ml and 1.73 μg/ml, respectively, in Huh7-YAP1 and Hep3B-YAP1
cells. These data indicate that YAP decreases doxorubicin
sensitivity of HCC cells.
Silencing YAP increased HCC cell
sensitivity to doxorubicin
We showed that YAP overexpression desensitizes HCC
cells to doxorubicin; therefore, we further explored the effects of
suppressing endogenous YAP expression on doxorubicin sensitivity of
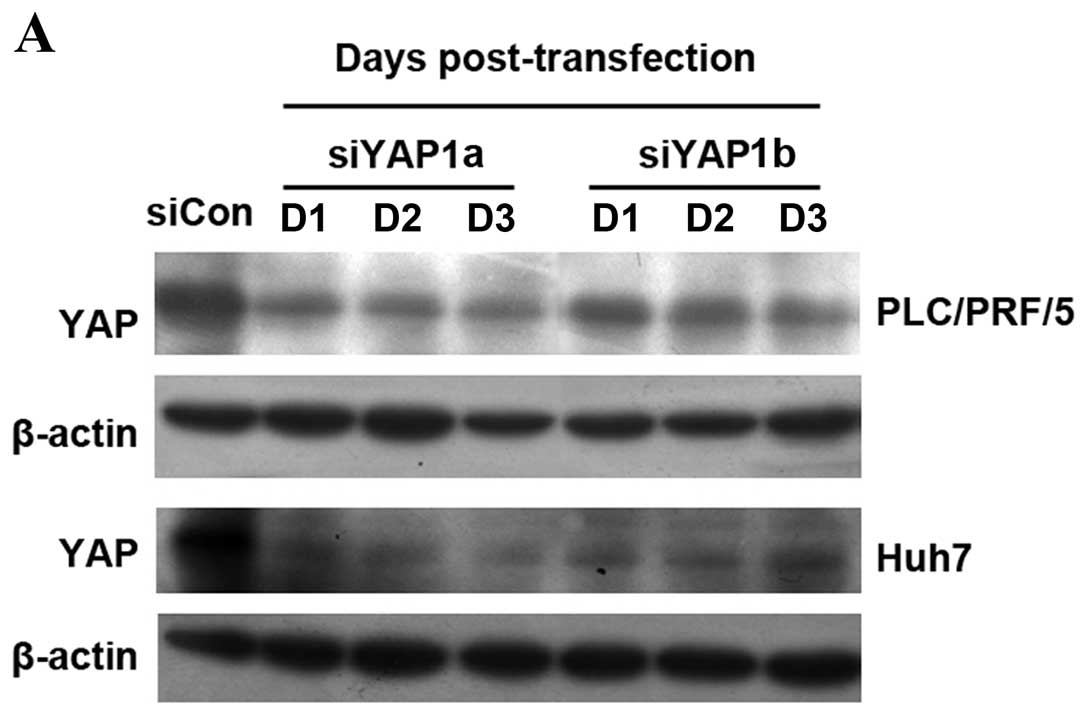
HCC cells. By using RNA interference, YAP expression in Huh7 and
PLC/PRF/5 cells was continuously downregulated from day 1 to day 3
post-transfection (Fig. 2A).
Twenty-four hours after transient transfection with siYAP1a and
siYAP1b, Huh7 and PLC/PFR/5 cells were treated with various
concentrations of doxorubicin for 48 h and the cell viability was
assessed by MTT assays. As shown in Fig. 2B, Huh7 and PLC/PRF/5 cells with
decreased YAP expression showed significantly lower cell viability
when comparing to the scramble siRNA-transfected controls. To
evaluate whether knockdown of YAP is auxiliary to doxorubicin
treatment on cell apoptosis, TUNEL assay was performed.
Downregulation of YAP expression significantly increased the number
of apoptotic cells in the PLC/PRF/5 and Huh7 cells following
treatment with doxorubicin (1 μg/ml) for 48 h (Fig. 2C).
MAP kinase pathway activation is involved
in YAP-mediated chemoresistance to doxorubicin
The effect of YAP on doxorubicin-treated HCC cell
apoptosis was further investigated by western blotting. After
exposure to 1.5 μg/ml of doxorubicin, Huh7-YAP1b cells showed less
cleaved PARP when compared to the Huh7-Vec cells (Fig. 3A) from day 1 to day 3. Meanwhile,
modest upregulation of phosphorylated Akt and notably increased
phosphorylated ERK1/2 were observed in the Huh7-YAP1b cells when
compared to their controls (Fig.
3A), without altered total protein levels of Akt or ERK1/2.
Although Bcl-xL and Bax were also induced by YAP overexpression,
semi-quantitative western blotting showed that the ratios of Bcl-xL
to Bax were significantly upregulated in Huh7-YAP1b cells when
compared to Huh7-Vec cells after exposure to the same concentration
of doxorubicin (Fig. 3B; p<0.01
on day 2 and 3). In line with these results, when exposed to 1 and
1.5 μg/ml doxorubicin for 48 h, PLC-siYAP1 cells showed increased
cleaved PARP, modestly decreased phosphorylated Akt, notably
decreased phosphorylated ERK1/2 and Bcl-xL expression when compared
to PLC-siCon cells after exposure to 1 and 1.5 μg/ml doxorubicin
for 48 h (Fig. 3C).
Semi-quantitative analysis showed the ratios of Bcl-xL to Bax were
significantly reduced in PLC-siYAP cells when compared to PLC-siCon
cells (Fig. 3D, p<0.01 for both
two dosages). These results indicate that suppression of YAP
enhances doxorubicin-induced apoptosis in HCC cells.
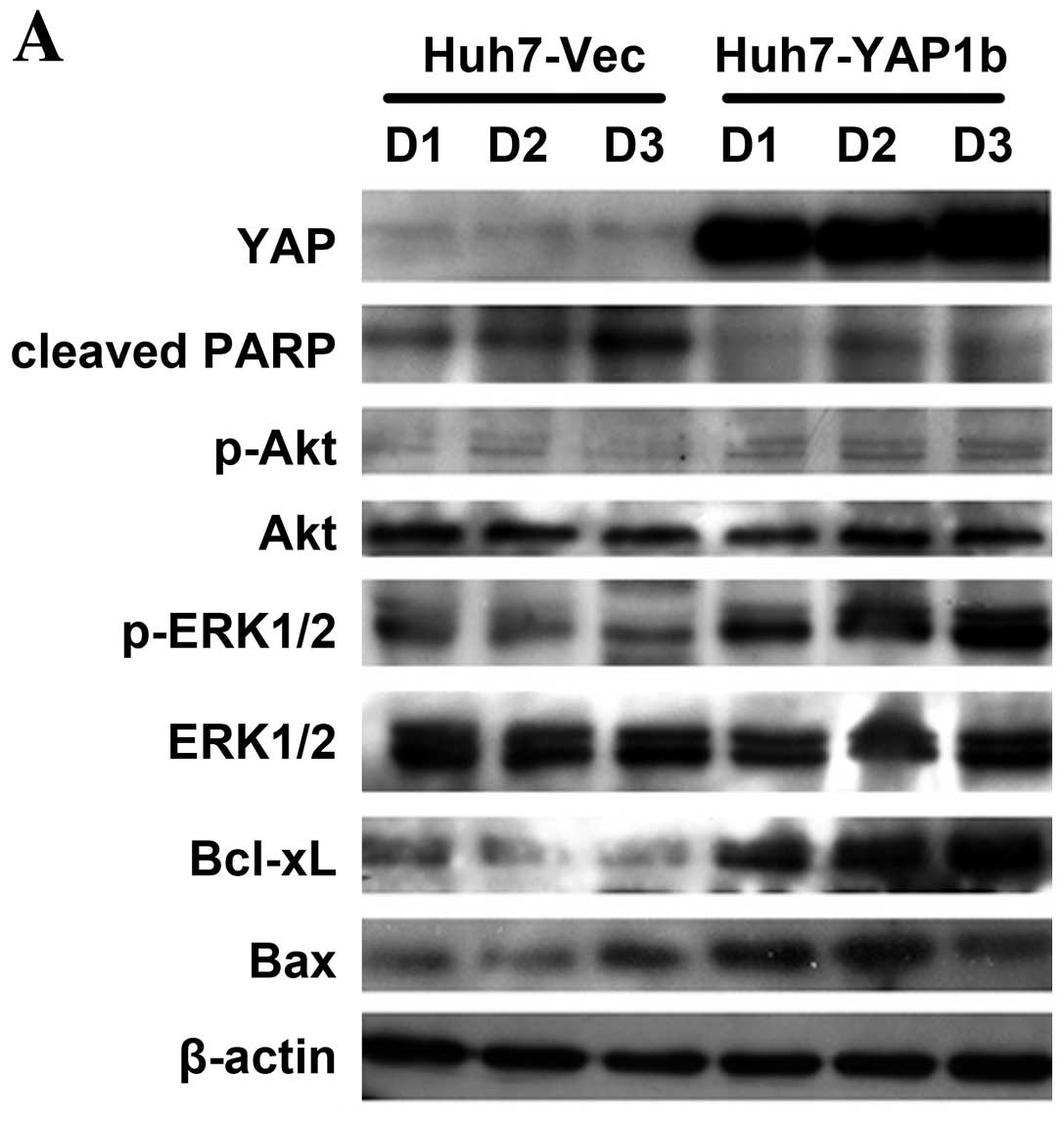 | Figure 3MAP kinase pathway mediates
YAP-conferred doxorubicin resistance in HCC cells. (A) When exposed
to 1.5 μg/ml doxorubicin, Huh7-YAP1b cells showed reduced cleaved
PARP, modestly increased activated Akt, significantly upregulated
phosphorylated ERK1/2 levels, elevated Bcl-xL and Bax expression
compared to Huh7-Vec cells. D1, D2 and D3 indicate the doxorubicin
treatment time points at day 1, 2 and 3. (B) The ratios of relative
protein levels of Bcl-xL over Bax significantly increased in the
Huh7-YAP1b cells when compared to the Huh7-Vec cells at day 2 and
3. Data at each time point are derived from 2 independent
experiments and shown as means ± SD. (C) When exposed to 1 or 1.5
μg/ml doxorubicin for 48 h, PLC-siYAP1a cells exhibited increased
cleaved PARP, modestly reduced phosphorylated Akt, significantly
decreased phosphorylated ERK1/2 levels and Bcl-xL expression. (D)
The ratios of the relative protein levels of Bcl-xL over Bax
significantly decreased in PLC-siYAP1 than in PLC-siCon when the
cells were stimulated with 1 and 1.5 μg/ml doxorubicin. Data at
each doxorubicin concentration were derived from 2 independent
experiments shown as means ± SD. (E) Huh7-YAP1b cells were
pretreated with DMSO, 25 μM LY294002 or 25 μM U0126, followed by
stimulation with doxorubicin. Cell viability was measured by MTT
assay. Comparisons were carried out by Student’s t-test between the
viability rates of YAP-overexpressing cells and those of control
cells at the indicated concentration of doxorubicin. Data are
derived from 3 independent experiments and shown as means ± SD.
*P<0.05; **P<0.01. (F) When exposed to
1.5 μg/ml doxorubicin, only U0126-pretreated Huh7-YAP1b cells
showed increased cleaved PARP, significantly reduced phosphorylated
ERK1/2 level and decreased Bcl-xL expression compared to the
DMSO-pretreated cells. LY294002-pretreated Huh7-1b cells showed a
significantly decreased phosphorylated Akt level, but not
significantly altered cleaved PARP and Bcl-xL expression
levels. |
As increased phosphorylated ERK1/2 and Akt was
observed in YAP-overexpressing cells, we investigated whether
activation of the MAP kinase or Akt signaling pathway plays roles
in mediating doxorubicin resistance in YAP-overexpressing HCC
cells. Pretreatment with the MAP kinase inhibitor U0126, but not
the PI3-K inhibitor LY294002, significantly decreased cell
viability in Huh7-YAP1b cells following exposure to doxorubicin
(Fig. 3E). Moreover, when treated
with both U0126 and doxorubicin, Huh7-YAP1b cells showed markedly
reduced Bcl-xL expression, which was in line with the decreased
phosphorylated ERK1/2 (Fig. 3F).
These findings suggest that activation of the MAP kinase pathway is
involved in YAP-dependent doxorubicin resistance in HCC cells.
Discussion
HCC is highly resistant to chemotherapy drugs.
Currently, systemic chemotherapy fails to prolong patient survival
time at large (4), and the response
rate to conventional chemotherapeutics is only approximately 25%
(18,19). Over the past 10 years, several
targeted agents, such as cetuximab, gefitinib and sorafenib, have
been evaluated in advanced HCC; nevertheless, the response rates to
these drugs were only modest and their effect was to mainly promote
disease stabilization (20).
Recently, concurrent treatment of unresectable HCC with
conventional TACE and sorafenib showed promising efficacy with
manageable safety (21). Therefore,
overcoming one of the major impediments of current treatment -
chemoresistance, could rely on a combination of targeted therapies
to improve chemocytotoxicity. In this study, we investigated the
significant role of YAP in influencing doxorubicin sensitivity of
HCC cells. We found that overexpression of YAP in HCC cells
conferred doxorubicin resistance whereas by suppressing its
endogenous expression the effect was attenuated. We suggest that
this is partially attributed to YAP-induced activation of the MAP
kinase pathway.
Aberrant overexpression of YAP has been described in
many types of human cancers (15).
The oncogenic properties of YAP include the induction of
epithelial-to-mesenchymal transition (EMT), promoting resistance to
apoptosis, enhancement of anchorage-independent growth capability
and increasing metastatic potential (6). Previous studies have also shown that
YAP is implicated in promoting chemoresistance to cisplatin in
ovarian cancer (22,23) and radioresistance in medulloblastoma
(24). Herein, we showed that YAP
expression is associated with doxorubicin sensitivity in HCC cells.
Both transient and stable overexpression of YAP1 enhanced cell
viability against doxorubicin in HCC cells. Compared to the
controls, Huh7-YAP1 cells showed less cleaved PARP at various drug
treatment time-points, indicating that YAP overexpression inhibited
doxorubicin-induced apoptosis in HCC cells. In line with this,
underexpression of YAP in both Huh7 and PLC/PRF/5 cells attenuated
doxorubicin resistance as evidenced by the reductions in their
IC50 values, increased cleaved PARP and increased
numbers of apoptotic cells in the presence of doxorubicin. These
results support that YAP mediates HCC cell chemoresistance to
doxorubicin. Therefore, targeting YAP could be used in combination
with doxorubicin to enhance chemotoxicity. Recently, dobutamine was
identified as a YAP inhibitor, prohibiting YAP nuclear
translocation and therefore inhibiting YAP-dependent gene
transcription (25). Based on these
findings, whether synergistic antitumor effects exist for HCC
requires further investigation.
Our study also revealed that YAP-dependent
doxorubicin resistance was strongly associated with increased
expression of Bcl-xL. Both Bcl-xL and Bax belong to the Bcl-2
family and are involved in the regulation of apoptosis. Bax
accelerates apoptosis by stimulating cytochrome c release,
which can be inhibited by Bcl-xL (26). Hence, an elevated ratio of
Bcl-xL/Bax is recognized as a phenomenon of cell fate under
apoptotic damage (27). Upon
apoptotic stimulation from DNA-damaging agents such as cisplatin
and doxorubicin, p73-mediated Bax expression was enhanced by YAP
overexpression and promoted apoptosis in breast and colon cancer
cell lines (28,29). Along with these findings, we found
that Bax expression was upregulated in Huh7-YAP1 cells and
attenuated in PLC-siYAP1 cells, which may be due to the
transcriptional activation by the YAP/p73 complex. However,
YAP-induced Bax expression did not increase apoptosis in the
YAP-overexpressing HCC cells. Compared to the controls, enhanced
cell viability and less cleaved PARP were observed at a series of
doxorubicin-treated time-points in Huh7-YAP1 cells. YAP-mediated
induction of Bcl-xL expression significantly elevated Bcl-xL/Bax
ratios, which were associated with enhanced cell viability
following DNA damage. These findings that YAP plays opposing roles
in different types of cancer cells in response to DNA-damaging
agents suggest that its proapoptotic or antiapoptotic activities
may be cell-type dependent.
Previously, we found that YAP overexpression in HCC
cells caused activation of the Akt and MAP kinase pathways
(9), which are closely associated
with chemoresistance (30-32). Herein, we showed that Huh7-YAP1
cells exhibited modestly phosphorylated Akt and notably
phosphorylated ERK1/2 after doxorubicin treatment when compared to
Huh7-Vec cells. Consistently, the levels of phosphorylated Akt,
phosphorylated ERK1/2 and Bcl-xL were decreased in
siYAP1-transfected cells than that in the controls. By using a
pathway-specific inhibitor, we demonstrated the involvement of the
MAP kinase pathway in YAP-conferred doxorubicin resistance. Only
the MEK1/2 inhibitor, U0126, reduced Bcl-xL levels and sensitized
Huh7-YAP1b cells to doxorubicin-induced apoptosis but not the PI3-K
inhibitor LY294002. The alteration of ERK1/2 phosphorylation levels
was also strongly correlated with the expression levels of Bcl-xL.
Inhibition of ERK1/2 activities was found to downregulate Bcl-xL
expression without affecting Bax expression (33). These results suggest that
YAP-conferred doxorubicin resistance to HCC cells was mediated by
activation of the MAP kinase pathway and upregulation of Bcl-xL
expression.
In conclusion, we provide evidence that YAP
overexpression plays an important role in the chemoresistance of
HCC cells to doxorubicin, which, at least partially, depends on
activation of the MAP kinase pathway. Targeting YAP could sensitize
cells to doxorubicin-induced cell death, suggesting that YAP may be
a therapeutic target for HCC in combination with conventional
TACE.
Acknowledgements
The study was supported by grants from the National
Natural Science Foundation of China to Dr Z. Xu (grant no.
81000880) and Dr J.M. Luk (grant no. 81128080); a grant from the
Technology Foundation for Selected Overseas Chinese Scholar,
Ministry of Personnel of China to Dr Z. Xu and a grant from the
Jiangsu Provincial 12th Five-Year Program on Developing Health by
Technology and Education Project to Dr J. Chen.
References
|
1
|
Meza-Junco J, Montano-Loza AJ, Liu DM, et
al: Locoregional radiological treatment for hepatocellular
carcinoma; Which, when and how? Cancer Treat Rev. 38:54–62. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huppert P: Current concepts in
transarterial chemoembolization of hepatocellular carcinoma. Abdom
Imaging. 36:677–683. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Llovet JM and Bruix J: Systematic review
of randomized trials for unresectable hepatocellular carcinoma:
Chemoembolization improves survival. Hepatology. 37:429–442. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Asghar U and Meyer T: Are there
opportunities for chemotherapy in the treatment of hepatocellular
cancer? J Hepatol. 56:686–695. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yin M and Zhang L: Hippo signaling: a hub
of growth control, tumor suppression and pluripotency maintenance.
J Genet Genomics. 38:471–481. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan SW, Lim CJ, Chen L, et al: The Hippo
pathway in biological control and cancer development. J Cell
Physiol. 226:928–939. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao B, Li L, Tumaneng K, Wang CY and Guan
KL: A coordinated phosphorylation by Lats and CK1 regulates YAP
stability through SCF(beta-TRCP). Genes Dev. 24:72–85. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Urtasun R, Latasa MU, Demartis MI, et al:
Connective tissue growth factor autocriny in human hepatocellular
carcinoma: oncogenic role and regulation by epidermal growth factor
receptor/yes-associated protein-mediated activation. Hepatology.
54:2149–2158. 2012. View Article : Google Scholar
|
|
9
|
Xu MZ, Chan SW, Liu AM, et al: AXL
receptor kinase is a mediator of YAP-dependent oncogenic functions
in hepatocellular carcinoma. Oncogene. 30:1229–1240. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao B, Wei X, Li W, et al: Inactivation
of YAP oncoprotein by the Hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Overholtzer M, Zhang J, Smolen GA, et al:
Transforming properties of YAP, a candidate oncogene on the
chromosome 11q22 amplicon. Proc Natl Acad Sci USA. 103:12405–12410.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang X, Su L and Ou Q: Yes-associated
protein promotes tumour development in luminal epithelial derived
breast cancer. Eur J Cancer. 48:1227–1234. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dong J, Feldmann G, Huang J, et al:
Elucidation of a universal size-control mechanism in
Drosophila and mammals. Cell. 130:1120–1133. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zender L, Spector MS, Xue W, et al:
Identification and validation of oncogenes in liver cancer using an
integrative oncogenomic approach. Cell. 125:1253–1267. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Steinhardt AA, Gayyed MF, Klein AP, et al:
Expression of Yes-associated protein in common solid tumors. Hum
Pathol. 39:1582–1589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu MZ, Yao TJ, Lee NP, et al:
Yes-associated protein is an independent prognostic marker in
hepatocellular carcinoma. Cancer. 115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu LX, Lee NP, Chan VW, et al: Targeting
cadherin-17 inactivates Wnt signaling and inhibits tumor growth in
liver carcinoma. Hepatology. 50:1453–1463. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ikeda M, Okusaka T, Ueno H, et al: Hepatic
arterial infusion chemotherapy with epirubicin in patients with
advanced hepatocellular carcinoma and portal vein tumor thrombosis.
Oncology. 72:188–193. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Park SH, Lee Y, Han SH, et al: Systemic
chemotherapy with doxorubicin, cisplatin and capecitabine for
metastatic hepatocellular carcinoma. BMC Cancer. 6:32006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Avila MA, Berasain C, Sangro B and Prieto
J: New therapies for hepatocellular carcinoma. Oncogene.
25:3866–3884. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park JW, Koh YH, Kim HB, et al: Phase II
study of concurrent transarterial chemoembolization and sorafenib
in patients with unresectable hepatocellular carcinoma. J Hepatol.
56:1336–1342. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang P, Bai Y, Song B, et al:
PP1A-mediated dephosphorylation positively regulates YAP2 activity.
PLoS One. 6:e242882011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang JM, Nagatomo I, Suzuki E, et al: YAP
modifies cancer cell sensitivity to EGFR and survivin inhibitors
and is negatively regulated by the non-receptor type protein
tyrosine phosphatase 14. Oncogene. Jun 11–2012.(Epub ahead of
print).
|
|
24
|
Fernandez LA, Squatrito M, Northcott P, et
al: Oncogenic YAP promotes radioresistance and genomic instability
in medulloblastoma through IGF2-mediated Akt activation. Oncogene.
31:1923–1937. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bao Y, Nakagawa K, Yang Z, et al: A
cell-based assay to screen stimulators of the Hippo pathway reveals
the inhibitory effect of dobutamine on the YAP-dependent gene
transcription. J Biochem. 150:199–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shimizu S, Narita M and Tsujimoto Y: Bcl-2
family proteins regulate the release of apoptogenic cytochrome c by
the mitochondrial channel VDAC. Nature. 399:483–487. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee TL, Yeh J, Friedman J, et al: A signal
network involving coactivated NF-kappaB and STAT3 and altered p53
modulates BAX/BCL-XL expression and promotes cell survival of head
and neck squamous cell carcinomas. Int J Cancer. 122:1987–1998.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Basu S, Totty NF, Irwin MS, Sudol M and
Downward J: Akt phosphorylates the Yes-associated protein, YAP, to
induce interaction with 14-3-3 and attenuation of p73-mediated
apoptosis. Mol Cell. 11:11–23. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lapi E, Di Agostino S, Donzelli S, et al:
PML, YAP, and p73 are components of a proapoptotic autoregulatory
feedback loop. Mol Cell. 32:803–814. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Naci D, El Azreq MA, Chetoui N, et al:
α2β1 integrin promotes chemoresistance against doxorubicin in
cancer cells through extracellular signal-regulated kinase (ERK). J
Biol Chem. 287:17065–17076. 2012.
|
|
31
|
Yang CL, Jiang FQ, Xu F and Jiang GX:
ADAM10 overexpression confers resistance to doxorubicin-induced
apoptosis in hepatocellular carcinoma. Tumour Biol. 33:1535–1541.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiao M and Nan KJ: Activation of PI3
kinase/Akt/HIF-1α pathway contributes to hypoxia-induced
epithelial-mesenchymal transition and chemoresistance in
hepatocellular carcinoma. Int J Oncol. 40:461–468. 2012.
|
|
33
|
Boucher MJ, Morisset J, Vachon PH, Reed
JC, Laine J and Rivard N: MEK/ERK signaling pathway regulates the
expression of Bcl-2, Bcl-X(L), and Mcl-1 and promotes survival of
human pancreatic cancer cells. J Cell Biochem. 79:355–369. 2000.
View Article : Google Scholar : PubMed/NCBI
|