Introduction
Prostate cancer (PCa) is one of the most commonly
diagnosed male cancers in Western countries and is a leading cause
of cancer mortality (1). Radical
prostatectomy and/or local radiotherapy are the standard treatment
for patients with organ-confined PCa. However, in 15–30% of these
patients, PCa progresses as metastatic disease (2).
Epigenetic inactivation of genes in cancer cells is
largely based on transcriptional silencing by aberrant CpG
methylation of CpG-rich promoter regions. Disruption in the DNA
methylation machinery resulting in global hypomethylation and
regional hypermethylation is well documented in most types of
cancer affecting gene activity. Genes with unmethylated CpG islands
are competent for regulated transcription, responding to signaling
cues by recruiting trans-activating factors that modify nucleosome
and chromatin structure, using histone acetyltransferases and
histone methyltransferases to promote transcript synthesis by RNA
polymerases. By contrast, genes with methylated CpG islands tend to
be incompetent for expression, tightly wound around nucleosomes in
a repressive chromatin structure maintained by histone deacetylases
(HDACs) and other enzymes. Thus, somatic increases in CpG island
methylation in cancer cells have been associated with gene
silencing and heterochromatinization. Bedford and van Helden in
1987 (3) observed that DNA
methylation levels were significantly lower in prostates with
benign prostatic hyperplasia (BPH) and metastatic tumors. By
contrast, non-metastatic prostate tumor DNA had a 5-methylcytosine
content essentially the same as in normal tissue.
Prostatic carcinogenesis has been associated with
the loss of glutathione S-transferase-π (GSTP1) by methylation
evident in some 5–10% of proliferative inflammatory atrophy (PIA)
lesions, the earliest prostate cancer precursors, and in <70% of
prostatic intraepithelial neoplasia lesions (4,5). Human
prostate cancer cells devoid of GSTP1 better activate heterocyclic
amine carcinogens, such as those found in overcooked meats, to
mutagenic species than cells capable of GSTP1 expression (6). GSTP1 silencing early during the
pathogenesis of prostate cancer, with the resultant loss of
enzymatic protection against reactive chemical species, may offer
an explanation for the well-known sensitivity of human prostatic
carcinogenesis to dietary and lifestyle habits (7–9).
Three major enzymes are responsible for DNA
methylation in eukaryotes: DNMT1, DNMT3a and DNMT3b (10–12).
DNMT1 has been implicated primarily in the maintenance of
methylation patterns that occurs during cellular replication and it
preferentially methylates hemi-methylated DNA. It has been the most
extensively studied maintenance methyltransferase and is abundant
in tumor cells and tissues. DNMT3a and DNMT3b are known to be de
novo methylators of CpG sites, which have higher
methyltransferase activity for unmethylated DNA than DNMT1 and can
contribute to de novo methylation during embryogenesis and
tumorigenesis. In the transgenic mouse model of PCa (TRAMP), DNMT
expression increases during the progressive stages of PCa (13,14).
The DNMT inhibitor 5-azacytidine (5-aza-CR) as well as its deoxy-
derivative, 5-aza-2′-deoxycytidine (5-aza-CdR), prevents prostatic
disease progression and the development of lymph node metastases in
this model (15). Demethylating
agents have been shown to be effective in the treatment of
myelodysplastic syndromes (16).
While in vitro experiments and animal models have shown that
5-aza-CR has antitumor activities in several types of cancer,
including prostate cancer (17–21),
clinical trials with 5-aza-CR (Vidaza) or its deoxy derivative
5-aza-CdR (decitabine) for the treatment of solid tumors have shown
no significant effects due to the high side-effects (22,23).
The majority of genes activated by demethylating agents, such as
Rb, p16, E-cadherin, APC, VHL, retinoic acid receptor β, BRCA1 and
DLC-1, are, however, tumor-suppressor genes and are related to
chemo-resistance (21,24) in different types of cancer cells. We
previously reported that prostate tumor cells may acquire androgen
independence upon hormone therapy through epigenetic mechanisms
involving modifications of DNMT expression and activity both in
patients treated with bicalutamide as neoadjuvant hormone therapy
(NHT) and in vitro systems (25–27).
In addition, it has been shown that upon castration, the TRAMP
mouse develops ‘castration-resistant’ prostate tumors similar to
those seen with the recurrence of human prostate tumor growth after
androgen-deprivation therapy. Treatment with 5-aza-CdR was found to
increase survival of TRAMP mice and delayed prostate cancer
progression, including the recurrence of prostate tumor growth
after castration (12,13). We also demonstrated that azacitidine
was able to amplify or restore bicalutamide treatment (26,27) or
chemotherapy (21). In the present
study, we analyzed the expression of DNMT1, DNMT3a and DNMT3b in
primary culture from BPH and PCa human tissue samples as well as in
neoplastic and non-neoplastic prostate epithelial cells.
Materials and methods
Reagents
All the materials for tissue culture were purchased
from Hyclone (Cramlington, NE, USA). Plasticware was obtained from
Nunc (Roskilde, Denmark). Azacitidine (Vidaza®) was
obtained in collaboration with Celgene Corp. (Summit, NJ, USA).
Antibodies against cytokeratin 8 (K8), 5 (K5) and 14 (K14) and PSA,
vimentin (VMNT) and AMACR were purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA). Anti-DNMT1, DNMT3a and DNMT3b
antibodies were purchased from BioCarta (Hamburg, Germany). GSPT1
monoclonal antibody (ALX-804-510) was purchased from Enzo Life
Sciences (Vinci-Biochem Srl Florence, Italy).
Cell lines
We used non-tumor prostate epithelial cell lines
[BPH-1, RWPE-1 and EPN (28)] and 9
PCa cells lines [CWR22 (29), 22Rv1
(29), LAPC-4 (30), LNCaP, VCaP (31), DuCaP (31), C4-2B (32), DU145 and PC3]. PC3 and LNCaP cell
derivatives (PCM, PC3-Pro4, PC3-LN4, LNCaP-Pro5 and LNCaP-Ln5) were
kindly provided by Dr I.J. Fidler and Dr C.A. Pettaway (33). Cells were cultured as previously
described.
Primary tumor cultures
The primary tumor cultures of PCa were established
from specimens of consenting patients undergoing radical
prostatectomy. A wedge-shaped specimen of the fresh prostate was
removed within 1 h of surgery. Frozen sections of a part of this
tissue were used to confirm the prostatic origin and for diagnosis.
We analysed 10 tissues harvested from BPH and 40 tissues derived
from patients with clinically localized tumors surgically treated
with prostatectomy as previously described (34–36).
Tissue samples were minced and cultured in DMEM.
Preparation of cell lysates and western
blot analysis
Cells were washed with cold PBS and immediately
lysed with 1 ml lysis buffer (50 mM HEPES, pH 7.5, 150 mM NaCl, 10%
gly- cerol, 1% Triton X-100, 1 mM EDTA, 1 mM EGTA, 50 mM NaF, 1 mM
sodium orthovanadate, 30 mM p-nitrophenyl phosphate, 10 mM sodium
pyrophosphate, 1 mM phenylmethylsulfonyl fluoride, 10 μg/ml
aprotinin and 10 μg/ml leupeptin). The nuclear extracts were
collected as described: cells were scraped in 1 ml PBS-EDTA and
centrifuged at 3,000 rpm. Pellets were resuspended in 1 ml harvest
buffer containing 10 mM HEPES, pH 7.9, 50 mM NaCl, 0.5 M sucrose,
0.1 mM EDTA and 0.5% Triton X-100 and incubated on ice for 5 min.
Cells were pelleted at 1,000 rpm with a table top swinging
microfuge, washed and resuspended in 1 ml of 10 mM HEPES, pH 7.9
containing 10 mM KCl, 0.1 mM EDTA and 0.1 mM EGTA. Lysates were
electrophoresed in 7% SDS-PAGE and separated proteins were
transferred to nitrocellulose and probed with the appropriate
antibodies using the conditions recommended by the suppliers.
DNA methyltransferase activity assay
DNMT activity was evaluated by a colorimetric
EpiQuik DNMT Activity Assay kit (BioVision, Mountain View, CA, USA)
in nuclear extracts of cells treated with 5-Aza-CR according to the
manufacturer’s instructions.
Statistical analysis
Continuous variables were summarized as mean and
standard deviation (SD) or as median and 95% CI for the median. For
continuous variables not normally distributed, statistical
comparisons between control and treated groups were established
using the Kruskal-Wallis tests. For continuous variables normally
distributed, statistical comparisons between control and treated
groups were established by the ANOVA test or by the Student’s
t-test for unpaired data (for two comparisons).
Results
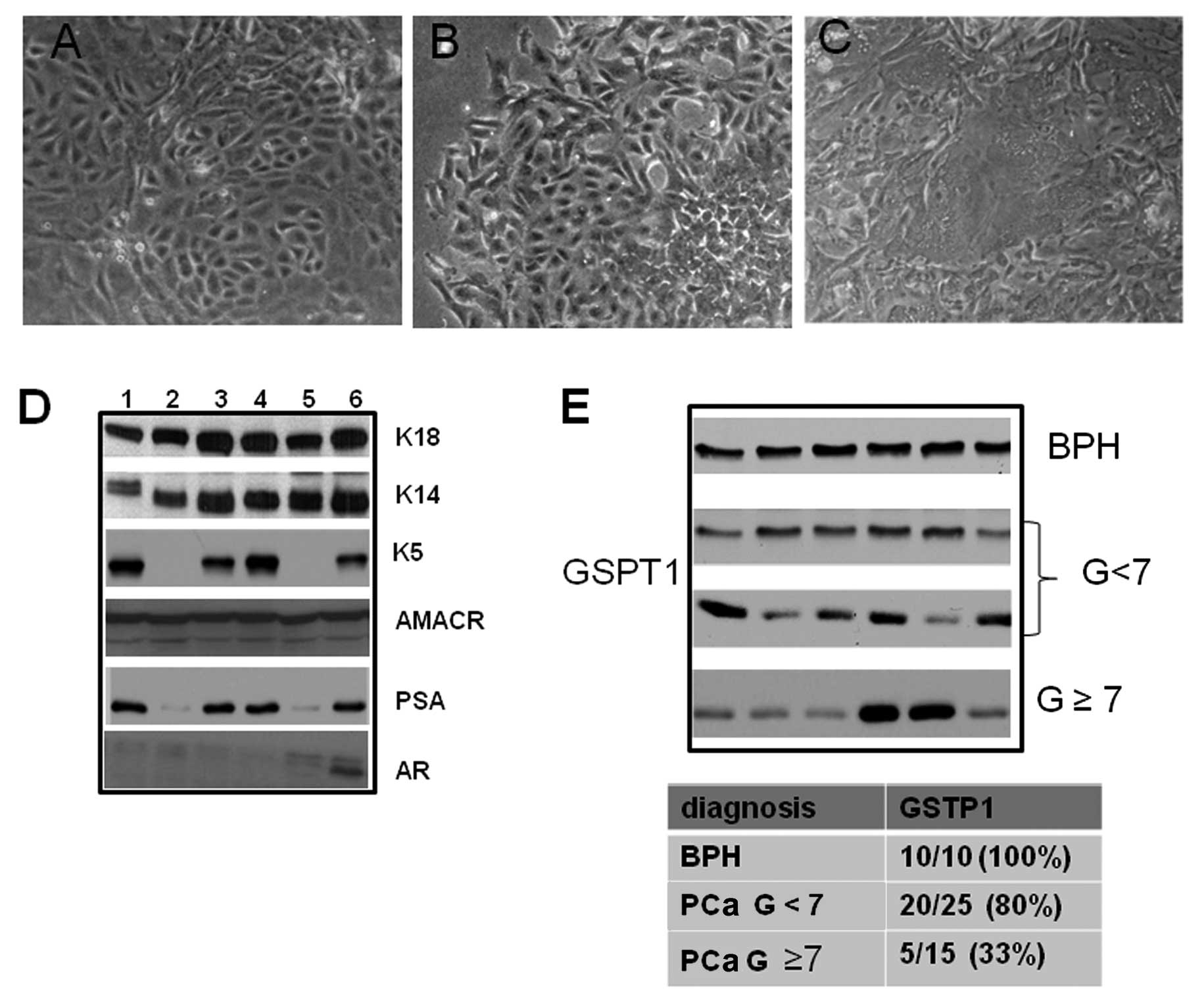
Characterization of primary epithelial cancer cell
cultures was performed as previously described (43–45). Organoids
and small cell aggregates similar to acini attached within 2–5 days
of primary culture and small outgrowths appeared with cells forming
polygonal colonies of various grades of architecture. A
morphological aspect of three prostate epithelial cultures with
small polygonal cells containing dark granules and large polyclonal
cells are shown in Fig. 1. Some
cells formed small compact isles and acquired a wavy pattern.
Moreover, we observed that some cells formed irregular colonies
showing senescent signs such as multinuclear aspect and flattened
epithelial sheets with a final tendency to intersect. Cells were
positive for K18, which are typical of prostatic lumenal cells, and
for K14, typical of basal cells with variable levels of K5, typical
for intermediated/amplifying cells. Low expression of AR and PSA
characterized our primary cell cultures whereas AMACR was present
in ~70% of primary cultures. Only AMACR positive cultures were used
in our study. In Fig. 1D we show
the expression of the above-mentioned markers in AMACR positive
cultures. A densitometric analysis of the GSTP1 expression
(Fig. 1E) allowed us to observe
that 10/10 (100%) of BPH cultures were GSTP1 positive whereas 62.5%
(25/40) of PCa cultures showed GSTP1 protein. Following
randomization in two groups based on the Gleason score, 5/25 (20%)
cultures containing low/absent levels of GSTP1 were grouped in the
Gleason score <7 whereas 10/15 (66.7%) with low/absent levels of
GSTP1 were in the Gleason score ≥7 group. In addition, we showed
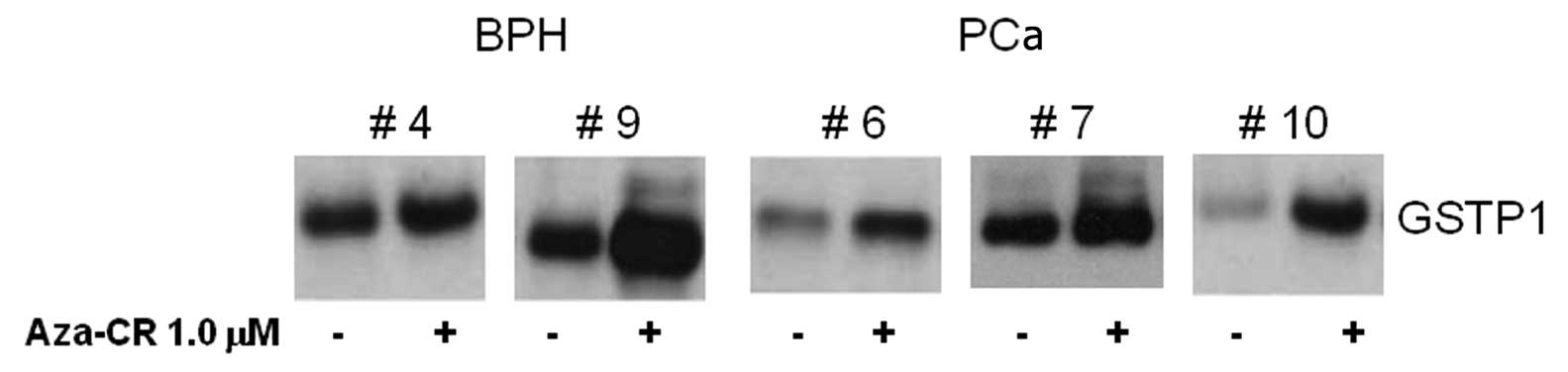
that 5-aza-CR was able to re-express GSTP1 in human primary cell
cultures (Fig. 2), suggesting that
GSTP1 may be considered a potentially useful biomarker for the
methylation status of prostate cancer cells, which is in accordance
with previous reports (11–14).
DNMT activity and expression in primary
cultures derived from BPH and PCa tissue samples
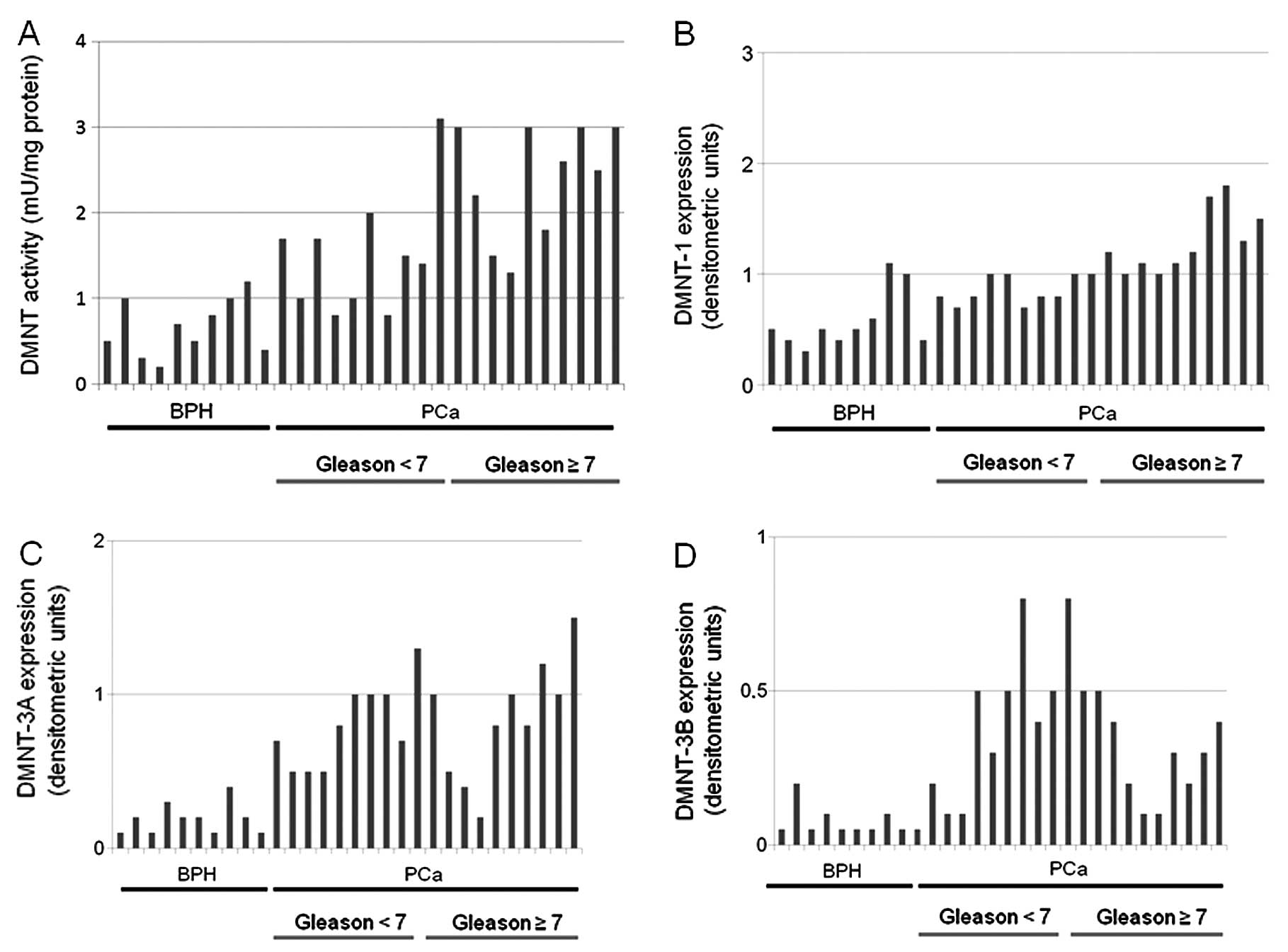
We also demonstrated that DNMT activity was higher
in primary cultures derived from PCa compared to BPH tissue
samples. In Fig. 3A we show single
values of DNMT activity in representative BPH (10/10), PCa Gleason
<7 (10/25) and PCa Gleason ≥7 (10/15). The differences between
the values of DNMT activity observed in BPH-derived cultures
(0.68±0.30 mU/mg of protein) were significantly lower compared with
those observed in PCa-derived cultures (1.90±0.81 mU/mg of protein,
P<0.001). DNMT activity was significantly higher in cultures
derived from Gleason score ≥7 (2.31±0.70 mU/mg of protein) compared
to those observed in Gleason score <7 derived cultures
(1.49±0.72 mU/mg of protein, P<0.005). The evaluation of the
levels of DNMT1 also shows that PCa-derived cultures resulted in
higher levels of protein compared to BPH cultures. In Fig. 3B we showed single values of DNMT1
protein expression in representative BPH (10/10), PCa Gleason <7
(10/25) and PCa Gleason ≥7 (10/15). The differences between the
values of arbitrary densitometric units for DNMT1 protein observed
in BPH-derived cultures (0.56±0.27 ADU) were significantly lower
compared with those observed in PCa-derived cultures (1.09±0.33
ADU, P<0.001). The DNMT1 protein levels were significantly
higher in cultures derived from Gleason score ≥7 (1.31±0.32 ADU)
compared to those observed in Gleason score <7 derived cultures
(0.86±0.13 ADU, P<0.001). Evaluation of the levels of DNMT3a
also shows that PCa-derived cultures resulted in higher levels of
protein compared to BPH cultures. In Fig. 3C we show single values of DNMT1
protein expression in representative BPH (10/10), PCa Gleason <7
(10/25) and PCa Gleason ≥7 (10/15). The differences between the
values of arbitrary densitometric units for DNMT3a protein observed
in BPH-derived cultures (0.25±0.14 ADU) were significantly lower
compared with those observed in PCa-derived cultures (0.83±0.30
ADU, P<0.001). The DNMT3a protein levels were similar in
cultures derived from Gleason score ≥7 (0.82±0.25 ADU) compared to
those observed in Gleason score <7-derived cultures (0.84±0.36
ADU, NS). Evaluation of the levels of DNMT3b also show that
PCa-derived cultures produced higher levels of protein compared to
BPH cultures. In Fig. 3D we show
single values of DNMT3b protein expression in representative BPH
(10/10), PCa Gleason <7 (10/25) and PCa Gleason ≥7 (10/15). The
differences between the values of arbitrary densitometric units for
DNMT3b protein observed in BPH-derived cultures (0.06±0.02 ADU)
were significantly lower compared with those observed in
PCa-derived cultures (0.43±0.17 ADU, P<0.001). The DNMT3a
protein levels were statistically lower in cultures derived from
Gleason score ≥7 (0.34±0.14 ADU) compared to those observed in
Gleason score <7-derived cultures (0.52±0.16 ADU, NS).
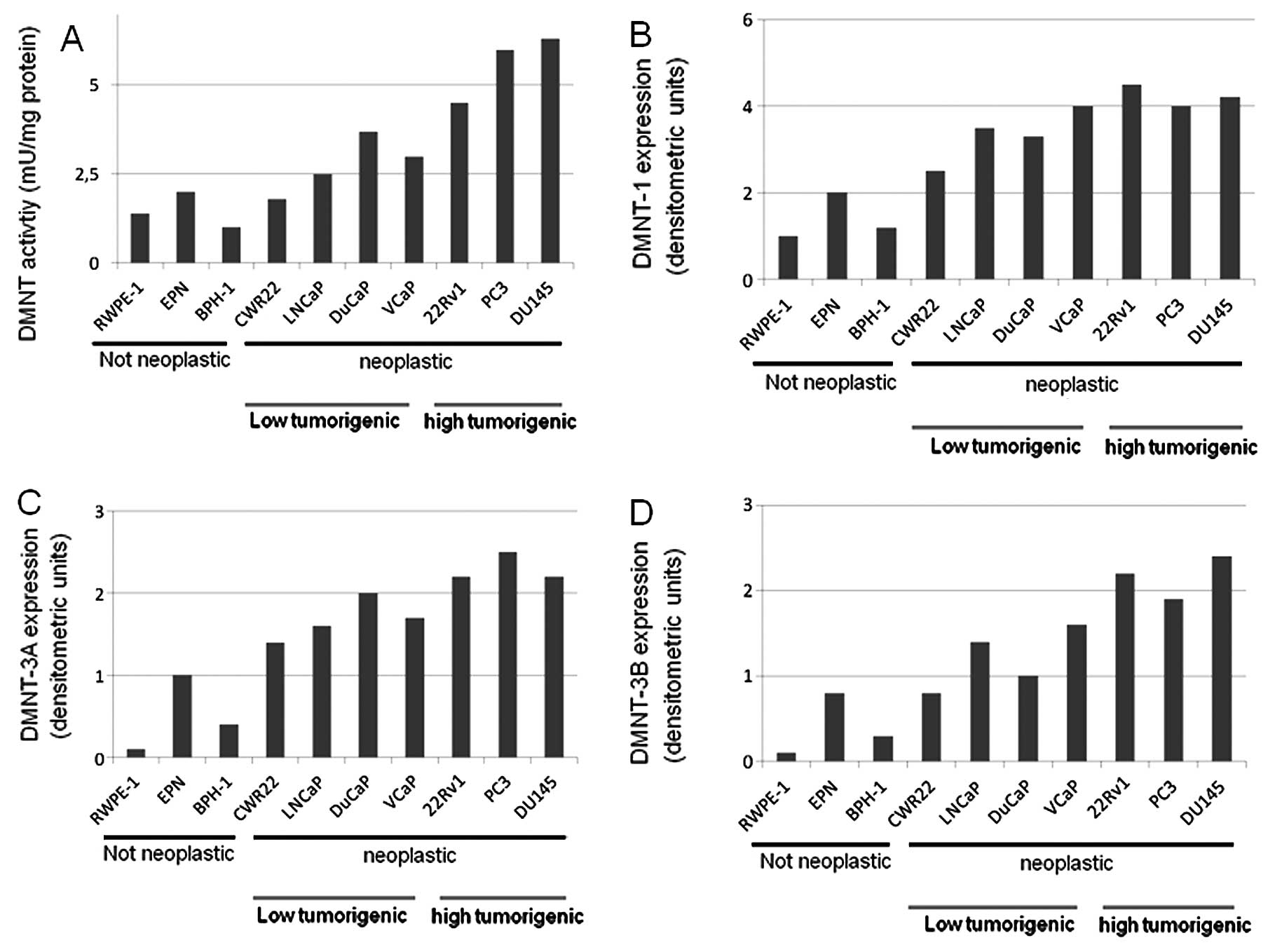
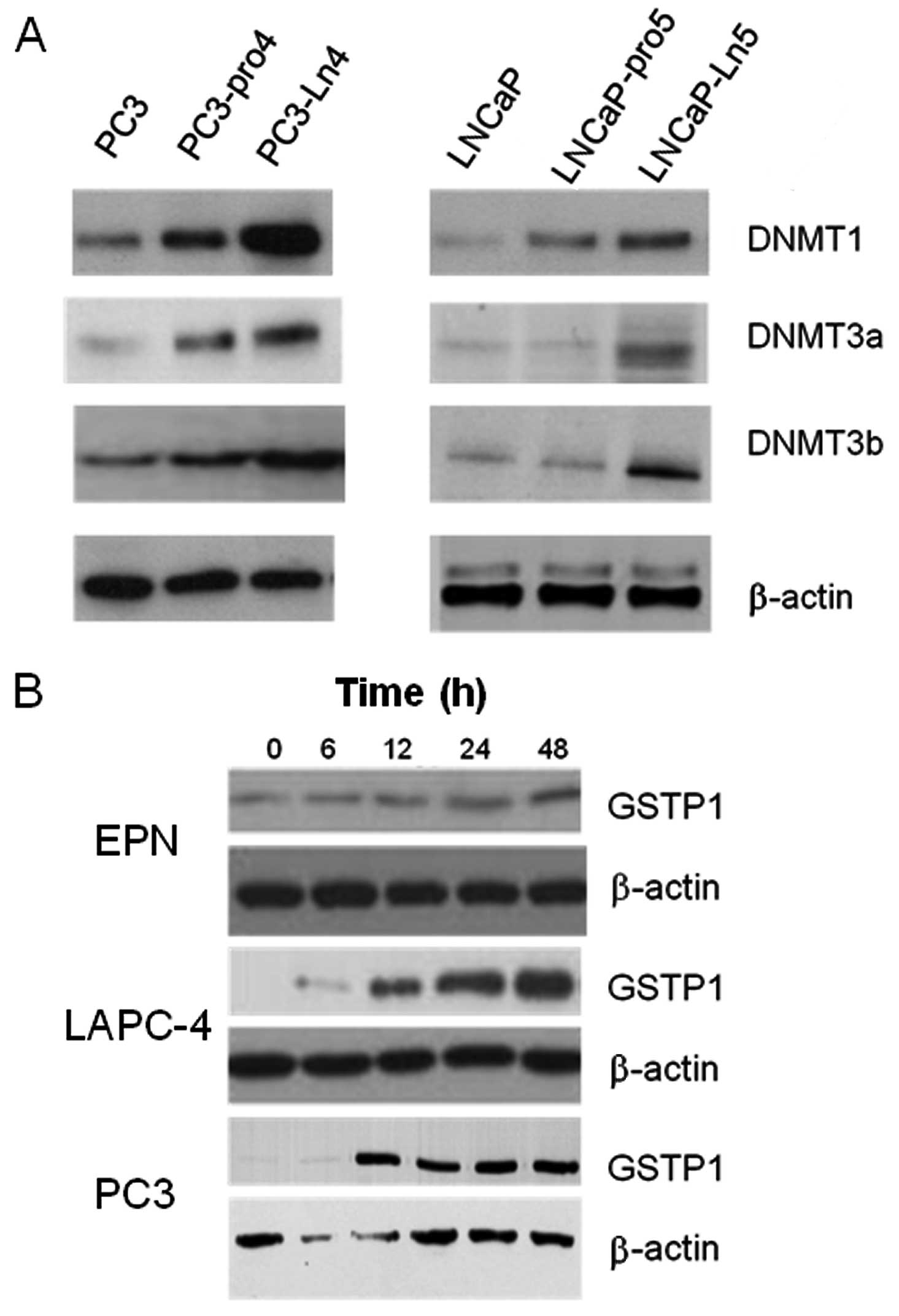
Higher DNMT activity and expression are
observed in more aggressive PCa cell lines
Next, we analyzed DNMT activity and DNMT expression
in different prostate cancer cell lines with different tumorigenic
and metastatic capacities. We demonstrated that DNMT activity
(Fig. 4A), as well as the
expression of DNMT1 (Fig. 4B),
DNMT3a (Fig. 4C) and DNMT3b
(Fig. 4D) were higher in neoplastic
vs. non-neoplastic prostate epithelial cell lines and this was in
agreement with previous findings (41). Conversely, only DNMT1 expression was
higher in aggressive/androgen-independent (PC3, DU145 and 22Rv1
cell lines) compared to low tumorigenic/androgen-dependent (CWR22,
LAPC4, LNCaP, DuCaP and VCaP) prostate cancer cells. The levels of
DNMT3a and DNMT3b were higher in more tumorigenic cells compared to
low tumorigenic cells but the differences were not significant. The
comparison in PC3 and LNCaP cell derivatives possessing higher
metastatic potential shows that DNMT1, DNMT3A and DNMT3b were
significantly higher (Fig. 5A) in
more metastatic PC3-LN4 and LNCaP-Ln5 cell derivatives compared to
parental cells.
GSTP1 may be re-expressed or up-modulated
by azacitidine
GSTP1 was expressed at high levels only in RWPE-1
and BPH1 cells, EPN cells expressed low levels of this protein
whereas all prostatic cancer cell lines analyzed were negative.
However, azacitidine was able to increase GSTP1 expression in EPN
cells and to restore protein expression in PCa cell lines. In
Fig. 5B we show the effects of
aza-CR on 3 representative cell lines (EPN, LAPC-4 and PC3).
Discussion
In recent years, several inhibitors of DNMTs have
been developed and evaluated in pre-clinical models and in clinical
trials. Among these, 5-azacytidine (5-aza-CR, Vidaza) and
5-aza-deoxycytidine (5-aza-dC, decitabine) have received Food and
Drug Administration (FDA) approval for the treatment of
myelodysplastic syndromes (MDS) (37,38),
and these agents and others are being tried alone and in
combination with other drugs as cancer therapeutic agents. One
major disadvantage of Vidaza or decitabine is that they are
nucleoside analogs, whose mechanism of action involves
incorporation of the aza-modified base into DNA during DNA
synthesis with subsequent covalent trapping of the DNMTs. As with
other nucleoside analogs, these drugs can have significant
cytotoxicity and can lead to major adverse effects when
administered to patients. While these compounds are effective in
the treatment of hematologic conditions, clinical trials in solid
tumors and in prostate cancer have shown limited or no efficacy
(22,25). This may be attributed to
inappropriate dose regimens leading to the above-mentioned
toxicity-related adverse events. However, as with several
anti-target compounds one of the possible obstacles encountered
with Vidaza or decitabine, could be the inability to select
patients to be enrolled in the clinical studies as well as the
inability to monitor the efficacy of the drug if not the conclusion
of the study. Based on our findings from the study as well as from
a previous report (39), we propose
monitoring the expression of DNMT1, DNMT3a and DNMT3b in prostate
tissues obtained from biopsy of radical prostatectomy, and
evaluating the levels of GSTP before and during the treatment,
taking into consideration that GSTP1 (a molecular hallmark of human
prostate cancer, the expression of which epigenetic silencing
reduces during prostate carcinogenesis and progression) may be
re-expressed after 5-aza-CR treatment and is easily measured in a
patient’s serum and urine (40–42) or
circulating tumor cells. In this study, we demonstrated that DNMT
activity and the levels of DNMT1, DNMT3a and DNMT3b were
significantly higher in primary cultures derived from PCa compared
to BPH. Greater differences in the protein levels were, however,
observed for DNMT3a and DNMT3b. Next, we demonstrated that DNMT
activity and DNMT1 enzyme expression were not statistically higher
in cultures derived from Gleason score ≥7 compared to those
observed in cultures derived from lower Gleason scores, whereas the
levels of DNMT3a and DNMT3b were statistically higher in Gleason
scores >7. This is in agreement with findings previously
observed in human PCa tissue in which we demonstrated that DNMT
expression, particularly for DNMT3a and DNMT3b levels, varied with
tumor differentiation (34). We
then demonstrated that DNMT activity was higher in aggressive
androgen-independent compared to low tumorigenic androgen-dependent
prostate cancer cells and this was in agreement with previous
results (43). In addition, we
demonstrated that GSTP1 may be a target for azacitidine treatment
since exposure of human prostate cancer cells to 1 mM aza-CR for
1–7 days caused a concentration- and time-dependent re-expression
of GSTP1, which was correlated with DNMT inhibition. Taken
together, our results support that GSTP1 may be considered a useful
marker of DNMTi treatment efficacy which will facilitate its use as
a biomarker in future clinical trials. The ability to track
efficacy of the drug using tissue biopsies or circulating tumor
cells at earlier time-points could greatly assist future clinical
trials. In addition, GSTP1 has the potential to improve the
assessment of drug efficacy, thus reducing both the duration and
cost of a clinical trial, and, secondly, it may improve the welfare
of patients in clinical trials by minimizing unnecessary
exposure.
References
|
1
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics. CA Cancer J Clin. 56:106–130. 2006.
|
|
2
|
Kirby R: Management of clinically
localized prostate cancer by radical prostatectomy followed by
watchful waiting. Nat Clin Pract Urol. 2:298–303. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bedford M and van Helden PD:
Hypomethylation of DNA in pathological conditions of the human
prostate. Cancer Res. 47:5274–5276. 1987.PubMed/NCBI
|
|
4
|
Nakayama M, Bennett CJ, Hicks JL, Epstein
JI, Platz EA, Nelson WG and De Marzo AM: Hypermethylation of the
human glutathione S-transferase-π gene (GSTP1) CpG island is
present in a subset of proliferative inflammatory atrophy lesions
but not in normal or hyperplastic epithelium of the prostate: a
detailed study using laser-capture microdissection. Am J Pathol.
163:923–933. 2003.
|
|
5
|
Brooks JD, Weinstein M, Lin X, Sun Y, Pin
SS, Bova GS, Epstein JI, Isaacs WB and Nelson WG: CG island
methylation changes near the GSTP1 gene in prostatic
intraepithelial neoplasia. Cancer Epidemiol Biomarkers Prev.
7:531–536. 1998.PubMed/NCBI
|
|
6
|
Nelson CP, Kidd LC, Sauvageot J, Isaacs
WB, De Marzo AM, Groopman JD, Nelson WG and Kensler TW: Protection
against 2-hydroxyamino-1-methyl-6-phenylimidazo[4,5-b]pyridine
cytotoxicity and DNA adduct formation in human prostate by
glutathione S-transferase P1. Cancer Res. 61:103–109. 2001.
|
|
7
|
Lin X, Tascilar M, Lee WH, Vles WJ, Lee
BH, Veeraswamy R, Asgari K, Freije D, et al: GSTP1 CpG island
hypermethylation is responsible for the absence of GSTP1 expression
in human prostate cancer cells. Am J Pathol. 159:1815–1826. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Henderson CJ, Smith AG, Ure J, Brown K,
Bacon EJ and Wolf CR: Increased skin tumorigenesis in mice lacking
π-class glutathione S-transferases. Proc Natl Acad Sci USA.
95:5275–5280. 1998.
|
|
9
|
De Marzo AM, Marchi VL, Epstein JI and
Nelson WG: Proliferative inflammatory atrophy of the prostate:
implications for prostatic carcinogenesis. Am J Pathol.
155:1985–1992. 1999.PubMed/NCBI
|
|
10
|
Kano M, Bell DW, Haber DA and Li E: DNA
methyltransferases Dnmt3a and Dnmt3b are essential for de novo
methylation and mammalian development. Cell. 99:247–257. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu R, Wang X, Chen ZF, Sun DF, Tian XQ, et
al: Inhibition of the extracellular signal-regulated
kinase/mitogen-activated protein kinase pathway decreases DNA
methylation in colon cancer cells. J Biol Chem. 282:12249–12259.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bai T, Tanaka T, Yukawa K and Umesaki N: A
novel mechanism for acquired cisplatin-resistance: Suppressed
translation of death-associated protein kinase mRNA is insensitive
to 5-aza-2′-deoxycitidine and trichostatin in cisplatin-resistant
cervical squamous cancer cells. Int J Oncol. 28:497–508.
2006.PubMed/NCBI
|
|
13
|
Gravina GL, Festuccia C, Marampon F, Popov
VM, Pestell RG, Zani BM and Tombolini V: Biological Rationale for
the use of DNA methyltransferase inhibitors as new strategy for
modulation of tumor response to chemotherapy and radiation. Mol
Cancer. 9:305–321. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zorn CS, Wojno KJ, McCabe MT, Kuefer R,
Gschwend JE, et al: 5-aza-2′-deoxycytidine delays
androgen-independent disease and improves survival in the
transgenic adenocarcinoma of the mouse prostate mouse model of
prostate cancer. Clin Cancer Res. 13:2136–2143. 2007.
|
|
15
|
Morey Kinney SR, Smiraglia DJ, James SR,
Moser MT, Foster BA and Karpf AR: Stage-specific alterations of DNA
methyltransferase expression, DNA hypermethylation, and DNA
hypomethylation during prostate cancer progression in the
transgenic adenocarcinoma of mouse prostate model. Mol Cancer Res.
6:1365–1374. 2008.
|
|
16
|
Stewart DJ, Issa JP, Kurzrock R, Nunez MI,
Jelinek J, et al: Decitabine effect on tumor global DNA methylation
and other parameters in a phase I trial in refractory solid tumors
and lymphomas. Clin Cancer Res. 15:3881–3888. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Iwata H, Sato H, Suzuki R, Yamada R,
Ichinomiya S, Yanagihara M, et al: A demethylating agent enhances
chemosensitivity to vinblastine in a xenograft model of renal cell
carcinoma. Int J Oncol. 38:1653–1661. 2011.PubMed/NCBI
|
|
18
|
Perry AS, Foley R, Woodson K and Lawler M:
The emerging roles of DNA methylation in the clinical management of
prostate cancer. Endocr Relat Cancer. 13:357–377. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pulukuri SM and Rao JS: Activation of
p53/p21Waf1/Cip1 pathway by 5-aza-2′-deoxycytidine inhibits cell
proliferation, induces pro-apoptotic genes and mitogen-activated
protein kinases in human prostate cancer cells. Int J Oncol.
26:863–871. 2005.
|
|
20
|
Festuccia C, Gravina GL, D’Alessandro AM,
Millimaggi D, Di Rocco C, Dolo V, et al: Downmodulation of dimethyl
transferase activity enhances tumor necrosis factor-related
apoptosis-inducing ligand-induced apoptosis in prostate cancer
cells. Int J Oncol. 33:381–388. 2008.
|
|
21
|
Festuccia C, Gravina GL, D’Alessandro AM,
Muzi P, Millimaggi D, Dolo V, et al: Azacitidine improves antitumor
effects of docetaxel and cisplatin in aggressive prostate cancer
models. Endocr Relat Cancer. 16:401–413. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
van Groeningen CJ, Leyva A, O’Brien AM,
Gall HE and Pinedo HM: Phase I and pharmacokinetic study of
5-aza-2′-deoxycytidine (NSC 127716) in cancer patients. Cancer Res.
46:4831–4836. 1986.
|
|
23
|
Thibault A, Figg WD, Bergan RC, Lush RM,
Myers CE, et al: A phase II study of 5-aza-2′deoxycytidine
(decitabine) in hormone independent metastatic (D2) prostate
cancer. Tumori. 84:87–89. 1998.
|
|
24
|
Kastl L, Brown I and Schofield AC: Altered
DNA methylation is associated with docetaxel resistance in human
breast cancer cells. Int J Oncol. 36:1235–1241. 2010.PubMed/NCBI
|
|
25
|
Gravina GL, Marampon F, Piccolella M,
Motta M, Ventura L, Pomante R, Popov VM, Zani BM, Pestell RG,
Tombolini V, Jannini EA and Festuccia C: Hormonal therapy promotes
hormone-resistant phenotype by increasing DNMT activity and
expression in prostate cancer models. Endocrinology. 152:4550–4561.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gravina GL, Festuccia C, Millimaggi D,
Dolo V, Tombolini V, de Vito M, Vicentini C and Bologna M: Chronic
azacitidine treatment results in differentiating effects,
sensitizes against bicalutamide in androgen-independent prostate
cancer cells. Prostate. 68:793–801. 2008. View Article : Google Scholar
|
|
27
|
Gravina GL, Marampon F, Di Staso M,
Bonfili P, Vitturini A, Jannini EA, Pestell RG, Tombolini V and
Festuccia C: 5-Azacitidine restores and amplifies the bicalutamide
response on preclinical models of androgen receptor expressing or
deficient prostate tumors. Prostate. 70:1166–1178. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sinisi AA, Chieffi P, Pasquali D,
Kisslinger A, Staibano S, Bellastella A and Tramontano D: EPN: a
novel epithelial cell line derived from human prostate tissue. In
Vitro Cell Dev Biol Anim. 23:165–172. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tepper CG, Boucher DL, Ryan PE, et al:
Characterization of a novel androgen receptor mutation in a
relapsed CWR22 prostate cancer xenograft and cell line. Cancer Res.
62:6606–6612. 2002.PubMed/NCBI
|
|
30
|
Craft N, Shostak Y, Carey M, et al: A
mechanism for hormone-independent prostate cancer through
modulation of androgen receptor signaling by the HER-2/neu tyrosine
kinase. Nat Med. 5:280–288. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Korenchuk S, Lehr JE, MClean L, Lee YG,
Whitney S, Vessella R, Lin DL and Pienta KJ: VCaP, a cell-based
model system of human prostate cancer. In Vivo. 15:163–168.
2001.PubMed/NCBI
|
|
32
|
Lin DL, Tarnowski CP, Zhang J, Dai J, Rohn
E, Patel AH, Morris MD and Keller ET: Bone metastatic
LNCaP-derivative C4–2B prostate cancer cell line mineralizes in
vitro. Prostate. 47:212–218. 2001.
|
|
33
|
Pettaway CA, Pathak S, Greene G, Ramirez
E, Wilson MR, Killion JJ and Fidler IJ: Selection of highly
metastatic variants of different human prostatic carcinomas using
orthotopic implantation in nude mice. Clin Cancer Res. 2:1627–1636.
1996.PubMed/NCBI
|
|
34
|
Festuccia C, Vincentini C, di Pasquale AB,
Aceto G, Zazzeroni F, Miano L and Bologna M: Plasminogen activator
activities in short-term tissue cultures of benign prostatic
hyperplasia and prostatic carcinoma. Oncol Res. 7:131–138.
1995.PubMed/NCBI
|
|
35
|
Festuccia C, Angelucci A, Gravina GL, Muzi
P, Miano R, Vicentini C and Bologna M: Epithelial and prostatic
marker expression in short-term primary cultures of human prostate
tissue samples. Int J Oncol. 26:1353–1362. 2005.PubMed/NCBI
|
|
36
|
Gravina GL, Biordi L, Martella F, Flati V,
Ricevuto E, Ficorella C, Tombolini V and Festuccia C: Epigenetic
modulation of PTEN expression during antiandrogenic therapies in
human prostate cancer. Int J Oncol. 35:1133–1139. 2009.PubMed/NCBI
|
|
37
|
Kaminskas E, Farrell A, Abraham S, Baird
A, Hsieh LS, Lee SL, Leighton JK, et al: Approval summary:
azacitidine for treatment of myelodysplastic syndrome subtypes.
Clin Cancer Res. 11:3604–3608. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kantarjian H, Issa JP, Rosenfeld CS,
Bennett JM, Albitar M, DiPersio J, Klimek V, et al: Decitabine
improves patient outcomes in myelodysplastic syndromes: results of
a phase III randomized study. Cancer. 106:1794–1803. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chiam K, Centenera MM, Butler LM, Tilley
WD and Bianco-Miotto T: GSTP1 DNA methylation and expression status
is indicative of 5-aza-2′-deoxycytidine efficacy in human prostate
cancer cells. PLoS One. 6:e256342011.PubMed/NCBI
|
|
40
|
Mulder TP, Peters WH, Wobbes T, Witteman
BJ and Jansen JB: Measurement of glutathione S-transferase P1-1 in
plasma: pitfalls and significance of screening and follow-up of
patients with gastrointestinal carcinoma. Cancer. 80:873–880. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Oude Ophuis MB, Mulder TP, Peters WH and
Manni JJ: Plasma glutathione S-transferase P1-1 levels in patients
with head and neck squamous cell carcinoma. Cancer. 82:2434–2438.
1998.
|
|
42
|
Prior C, Guillen-Grima F, Robles JE,
Rosell D, Fernandez-Montero JM, Agirre X, et al: Use of a
combination of biomarkers in serum and urine to improve detection
of prostate cancer. World J Urol. 28:681–686. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Patra SK, Patra A, Zhao H and Dahiya R:
DNA methyltransferase and demethylase in human prostate cancer. Mol
Carcinog. 33:163–171. 2002. View
Article : Google Scholar : PubMed/NCBI
|