Introduction
Pancreatic cancer is a highly lethal disease that is
usually diagnosed at an advanced stage for which there is little or
no effective therapy. It remains the fourth most common cause of
cancer-related death in the Western world (1). Due to the aggressive natural history
of this disease, most patients with pancreatic cancer present with
local invasion or distant metastasis at the time of diagnosis, and
less than 20% of patients are candidates for surgery with curative
intent (2). The overall 5-year
survival rates are reported to be below 5% (3). No adjuvant treatments have shown
efficacy in improving survival to date. Thus, new approaches
including gene therapy are definitely required to improve treatment
results (4,5).
Angiogenesis is necessary for successful tumor
growth (6,7), and inhibition of VEGF represents the
most validated anti-angiogenic approach described thus far
(8,9). VEGF is a key stimulating factor for
angiogenesis of cancer, and it contributes to the malignant
development and metastasis of tumors through many processes. In
addition, VEGF is highly expressed in most human tumors (10–14).
RNA interference (RNAi) has emerged as a powerful
tool to induce lose-of-function phenotypes by post-transcriptional
silencing of gene expression (15,16).
Lentiviral vectors have provided a huge advance in technology and
offer the means to achieve significant levels of gene transfer
in vitro and in vivo(17,18).
In this study, we used the lentiviral vector
mediating RNAi to deliver a specially designed small hairpin RNA
for the human VEGF gene (LV-RNAi) into pancreatic carcinoma cell
line Patu8988 to observe the gene therapeutic effects on
angiogenesis and progression.
Materials and methods
Animals and cell lines
BALB/c nude mice were obtained from the Shanghai
Experimental Animal Center (Shanghai, China) and maintained
according to guidelines of the Animal Research Committee of Soochow
University (Suzhou, China).
Human pancreatic cancer cell line Patu8988 was
provided by Professor Chang-Geng Ruan, Jiangsu Institute of
Hematology, and maintained in RPMI (Roswell Park Memorial
Institute)-1640 medium (Gibco, USA) supplemented with 10% fetal
bovine serum (FBS) at 37°C in a humidified atmosphere containing 5%
CO2. The cultures were passaged 2 or 3 times weekly to
maintain log-phase growth.
Lentiviral vectors for VEGF shRNA
Small hairpin RNA (shRNA) targeting human VEGF
(GenBank, NM 001025366) was designed as follows. The underlined
part in the sense strand is the target sequence of the VEGF gene
which is 19-bp long, the italicized characters in the sense strand
is the loop sequence of the hairpin. According to the sense strand,
the antisense was also synthetized: sense, 5′-GATCCC(G)CCAT
GAACTTTCTGCTGTCTTGATATCCGGACAGCAGAA
AGTTCATGGTTTTTTCCAAC-3′; antisense, 3′-GG(C)GGT
ACTTGAAAGACGACAGAACTATAGGCCTGTCGTCTT
TCAAGTACCAAAAAAGGTTGAGCT-5′.
The recombinant lentivirus gene transfer vector
targeting VEGF pGCSIL-GFP-VEGF (LV-RNAi) encoding the green
fluorescent protein (GFP) sequence was constructed and gifted by Dr
L. Li followed by chemically synthesized shRNAs and lentiviral
vector constructed as previously described (19,20).
The targeting sequence of the shRNA was confirmed by sequencing.
The lentiviral vector pGCSIL-GFP-Negative (LV-NC) containing an
invalid RNAi sequence (GeneChem, Shanghai) was used to monitor
non-specific responses caused by heterologous siRNA. The LV-RNAi
and the LV-NC were prepared to 5×109 Tu/ml (transfection
units/ml).
Lentiviral vector transfection
Cells were subcultured at 5×104
cells/well into 6-well tissue culture plates overnight. The viral
supernatant was then added into cells at a multiplicity of
infection (MOI) of 10 with ENi.S and 5 μg/ml Polybrene. The
infected cells were considered to be the LV-RNAi and the LV-NC
group, respectively, and the Patu8988 cells without infection were
considered as the control group. Flow cytometry was used to detect
the transfection efficiency, and fluorescence microscopy was used
to observe the cells which released fluorescence. The three groups
mentioned above were used in the experiments below.
Real-time quantitative RT-PCR
Total RNA was collected using TRIzol reagent
following the manufacturer’s instructions. The concentration and
purity of the total RNA were detected with an ultraviolet
spectrophotometer and then reversely transcribed into cDNA with
MMLV. Total RNA (2 μg) was converted to cDNA in 40 μl and stored at
−20°C until use. The transcriptional level of VEGF was analyzed
using the MJ Research DNA Engine Opticon 2 System with SYBR-Green
fluorochrome. The GAPDH gene was used as an internal control. PCR
was carried out with cDNA derived from 50 ng of RNA, 1 unit Taq
polymerase and reaction kits in a final volume of 25 μl. Each cycle
of PCR included 15 sec of denaturation at 95°C, 20 sec of primer
annealing at 58°C and 20 sec of extension/synthesis at 72°C. The
primer sequences were as follows: 5′-GCTTTACTGCTGTACCTCCAC-3′
(sense) 5′-TCCAGGGCTTCATCGTTA-3′ (antisense) for VEGF (239 bp);
5′-GCAAGTTCAACGGCACAG-3′ (sense) 5′-GCCAGTAGACTCCACGACAT-3′
(antisense) for GAPDH (140 bp).
Western blotting
Cells were washed twice and lysed on ice. After
centrifugation, the supernatants were collected. Protein
concentrations were determined using the Bio-Rad DC Protein Assay
system. The β-actin gene was used as an internal control. The
following steps were as previously described (20).
Detecting VEGF levels in culture
supernatants by ELISA
Cells were seeded in new cell culture bottles, and
after a 72-h culture supernatants were collected and cell counting
was performed. The expression of VEGF in the collected supernatants
was tested by a human VEGF ELISA kit (R&D Co.) according to the
handbook. VEGF concentration/cell count was considered as the VEGF
expression level. VEGF expression level in samples of the control
group was considered as 1, and the expression of VEGF in the other
groups was calculated by comparison to the control. Duplicate wells
were set, and the entire experiment was repeated twice.
Subcutaneous transplantation model
A total of 18 BALB/c-nu mice, 5-weeks old and 20–24
g in weight, were bred in a specific pathogen-free (SPF) condition
and maintained at a constant humidity and temperature (25–28°C).
All mice underwent subcutaneous injection of a 200-μl cell
suspension of Patu8988 cells (1.0×107) in the
infra-axillary region, respectively. Two weeks later, the animals
were randomly divided into three groups and intratumorally injected
only once with 400 μl normal saline, 400 μl LV-NC or 400 μl
LV-RNAi, respectively. The size of the tumors was measured in a
blinded manner once a week with calipers, and the volume was
determined using the simplified formula of a rotational ellipsoid
(L × W2 × 0.5).
Immunohistochemical staining
Tumors were harvested from mice 5 weeks after
treatment, and VEGF expression and microvessel density (MVD) of the
tumor specimens were determined by immunohistochemistry. The tissue
specimens fixed with formalin solution were embedded in paraffin
wax, serially sectioned at 4 μm and immunohistochemically stained
using the SP method according to the manufacturer’s instructions
for the SP kit. The primary antibodies were diluted to 1:50 for
VEGF (as mentioned above) and 1:100 for CD34 (Santa Cruz
Biotechnology, Santa Cruz, CA, USA). Rectal cancer slides served as
the positive control, and PBS was used to replace the primary
antibodies to serve as the negative control. The expression of VEGF
protein was scored semi-quantitatively. Sections were then
evaluated for the presence of brown diaminobenzidine precipitates
indicative of positive reactivity by microscopy. Ten visual fields
(magnification, ×200) were counted for each section. The brown
staining in or around the nucleus was considered as positive
reactivity for VEGF. CD34 is used as a biomarker in endothelial
cells for the identification of new blood microvessels. One lumen
of blood vessels was assessed as one new blood capillary. The MVD
value was determined based on Weidner’s method (21).
Terminal deoxynucleotidyl
transferase-mediated dUTP nick-end labeling (TUNEL) assay
Apoptotic tumor cells were detected with the TUNEL
method, using an in situ cell death detection kit (Roche
Diagnostics, Mannheim, Germany). The assay was performed according
to the manufacturer’s instructions. Briefly, after routine
deparaffinization and treatment with H2O2
(3%), sections were digested with proteinase K (20 μg/ml, pH 7.4,
12 min) at 25°C and incubated with the reaction mixture (1:40, 60
min) at 37°C. Incorporated fluorescein was detected with
horseradish peroxidase after a 30-min incubation at 37°C and
subsequently dyed with DAB. Cell with brown-colored nuclei were
assessed as positive apoptotic cells, and the number of apoptotic
cells counted for 1,000 tumor cells in one section for at least 5
high power fields, was scored as the apoptotic index (AI).
Orthotopic transplantation pancreatic
cancer model
The establishment of the infra-axillary subcutaneous
transplantation tumor model was performed as described in the
‘Subcutaneous transplantation model’ section. After reaching a
specific volume, the tumors were resected under aseptic environment
and washed twice in antibiotic-containing RPMI-1640 to prevent
possible infection. Necrotic tissues were removed, and the
remaining viable tumor tissues were cut into small pieces of 1
mm3. Five-week-old BALB/c-nu mice, weighing 20–24 g,
were anesthetized with urethane (4 ml/kg) by intramuscular
injection. After the abdominal skin was sterilized, an incision was
made in the upper left abdomen and the pancreas was exposed. Tumor
pieces were attached to the pancreas using absorbable sutures. The
pancreas was then returned to the peritoneum, and the abdominal
wall and the skin were closed with silk sutures, respectively. The
animals were allowed to recover for 24 h. Eighteen surviving mice
were randomly divided into three groups (n=6) and intraperitoneally
injected only once with 400 μl normal saline, 400 μl LV-NC or 400
μl LV-RNAi, respectively. All of the mice were sacrificed 6 weeks
after treatment. Tumors were harvested from mice, and the volume
was determined as previously described. Liver metastasis was also
observed.
Statistical analysis
Statistical analysis was carried out using SAS 9.0
statistical software. Data are presented as the means ± standard
deviation (SD). The Student’s t-test or ANOVA was used to compare
the means of different groups. Chi-square test was used to compare
categorical variables and clinical pathological correlation. The
relationships among VEGF and MVD were investigated by Spearman-rank
correlation. A P<0.05 was considered to indicate a statistically
significant difference.
Results
Sequencing result and transfection
efficiency of the lentiviral vector
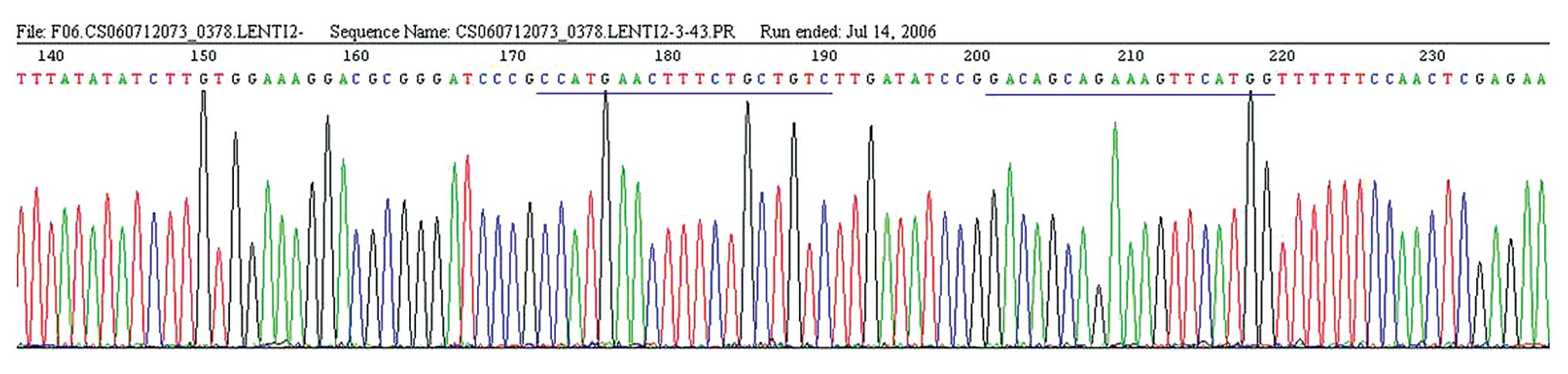
The result of sequencing for the recombinant vector
confirmed that the target sequences were constructed to the
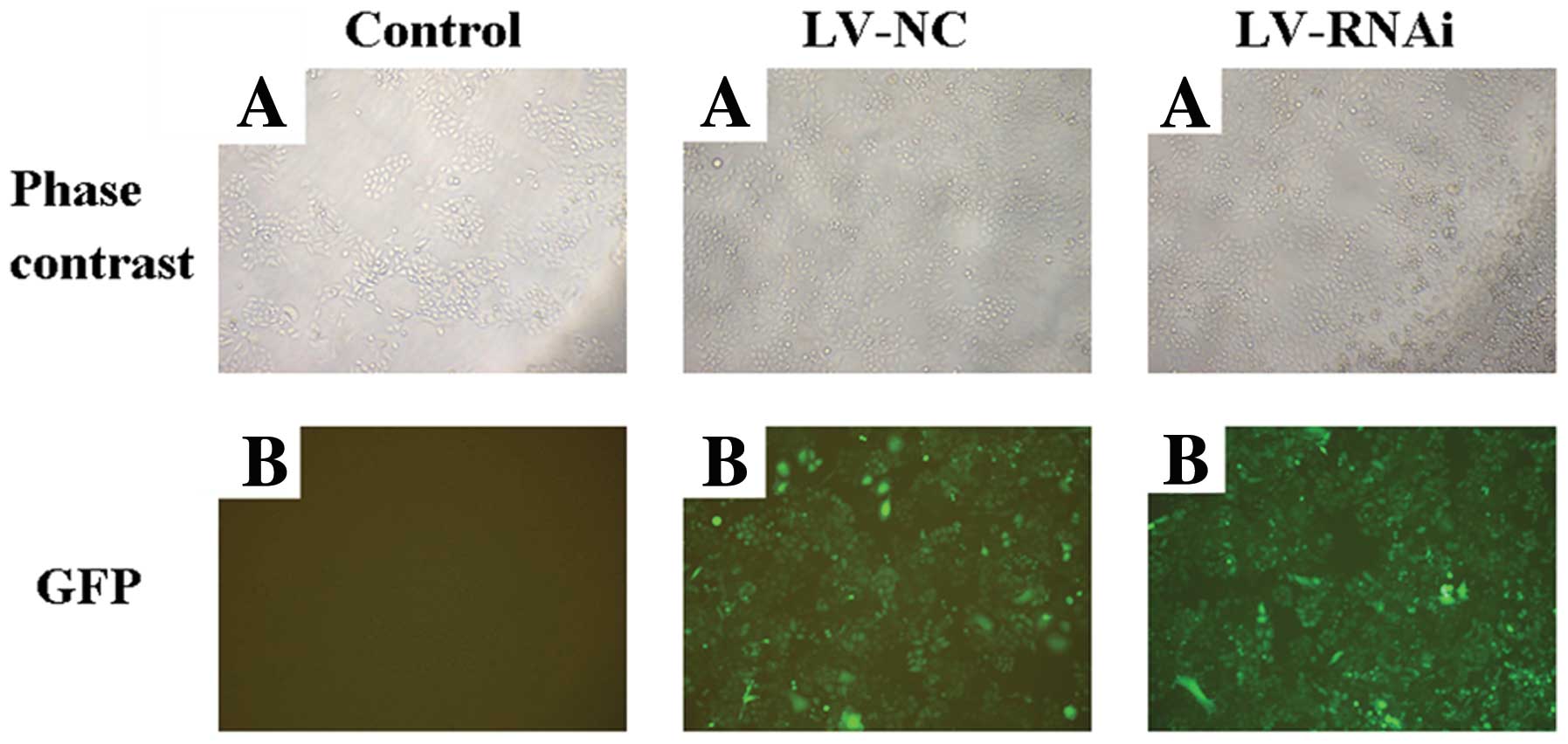
lentivirus system pGCSIL-GFP successively (Fig. 1). We used a lentiviral vector system
to express shRNAs directed against VEGF. In addition, GFP was as
used as a reporter gene. After a single exposure of Patu8988 cells
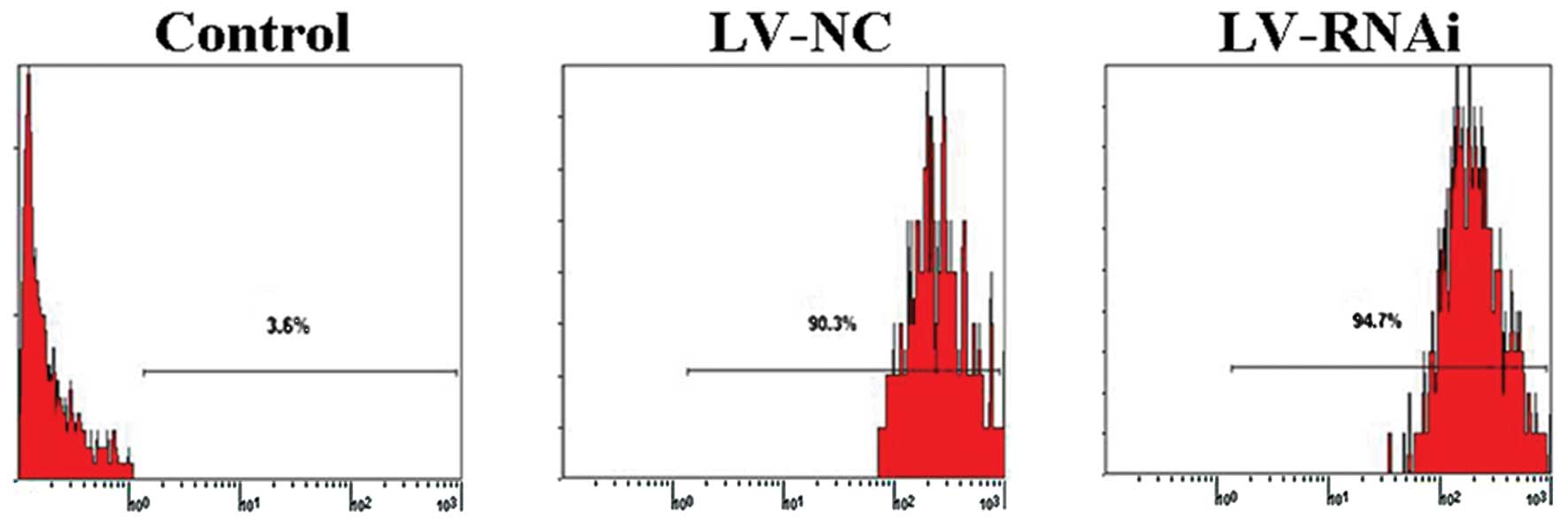
to the encoding lentivirus at MOI of 10 for 120 h, the cells were
examined by fluorescence microscopy (Fig. 2) and flow cytometry (Fig. 3). A high percentage (>90%) of
transfectants expressed GFP, indicating high and stable
transfection of the lentiviral vector system.
Assessment of the VEGF silencing effect
by RQ-PCR and western blotting
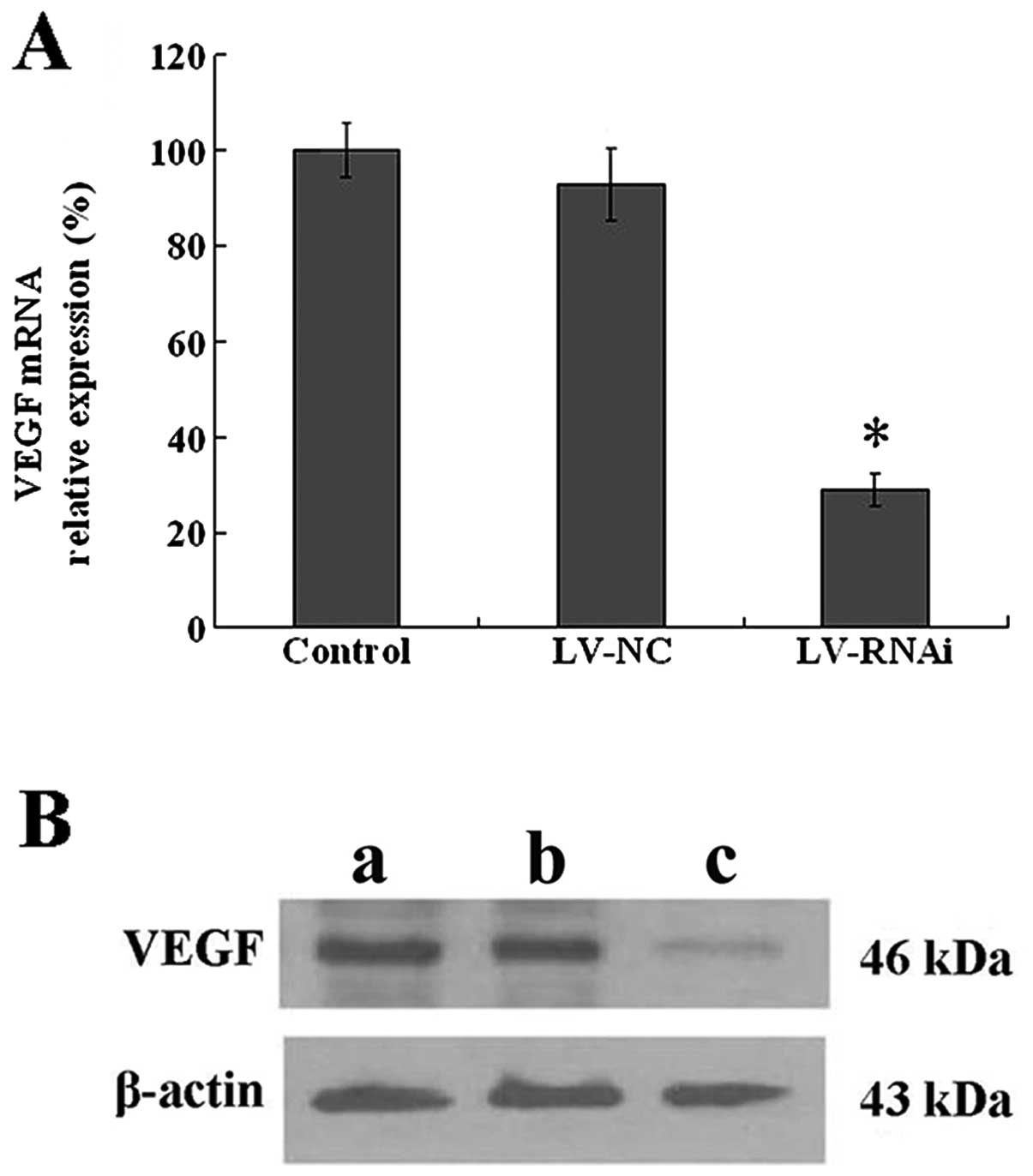
To detect the effect of VEGF silencing, real-time
RQ-PCR and western blot analysis were performed to determine the
mRNA and protein levels of VEGF following transfection. As shown in
Fig. 4A, the relative VEGF mRNA
expression (0.29±0.04, P<0.05) was significantly decreased in
the LV-RNAi group compared with the LV-NC (0.93±0.07) and the
control group (1.00±0.06), while no significant differences were
noted between the LV-NC and control group (P>0.05). As shown in
Fig. 4B, a 46-kDa protein band,
VEGF protein, was detected in the control and the LV-NC group, but
was weakly expressed in the LV-RNAi group.
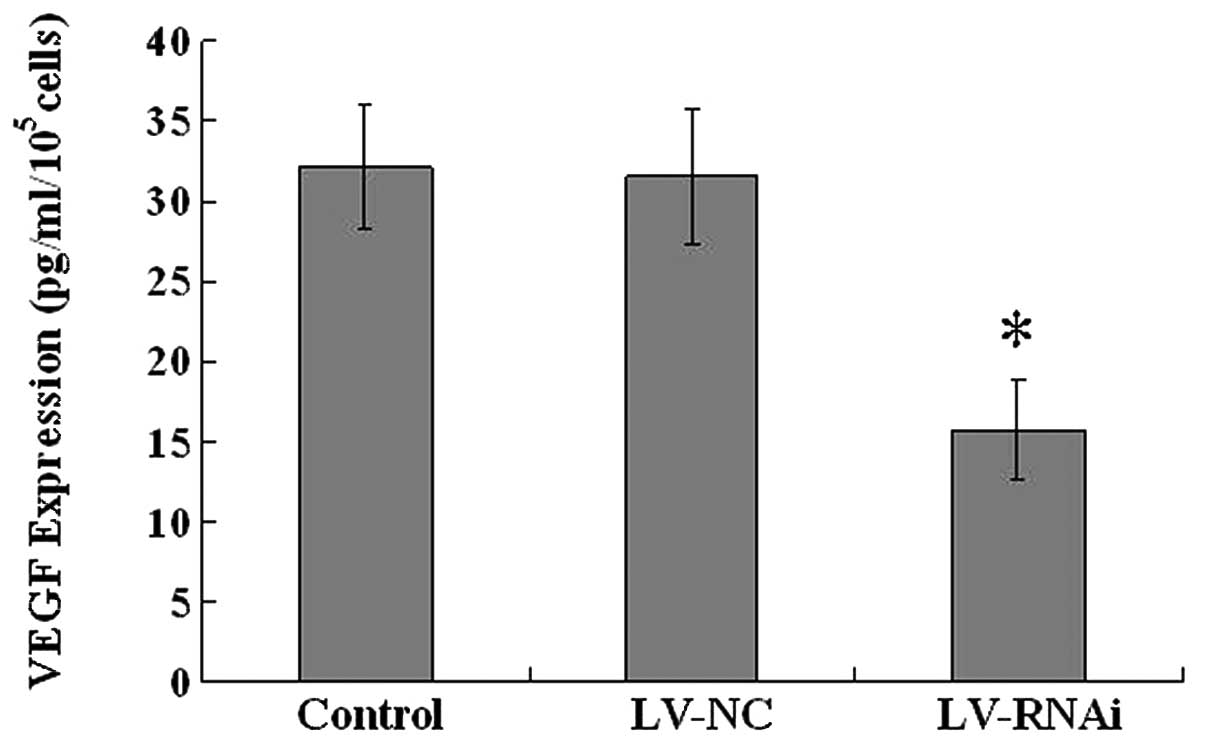
VEGF expression levels in vitro
To detect the downregulation effect on VEGF
expression, ELISA assay was performed. VEGF concentration/cell
counting was considered as the VEGF expression level (Fig. 5). The VEGF expression level in the
culture supernatants of the LV-RNAi group was 15.7±3.06
pg/ml/105cells, and in comparison to the control group
(32.16±3.90 pg/ml/105cells), it was obviously inhibited
(P<0.05), with a high inhibition efficiency (51.18%). There were
no significant differences between the LV-NC and the control
group.
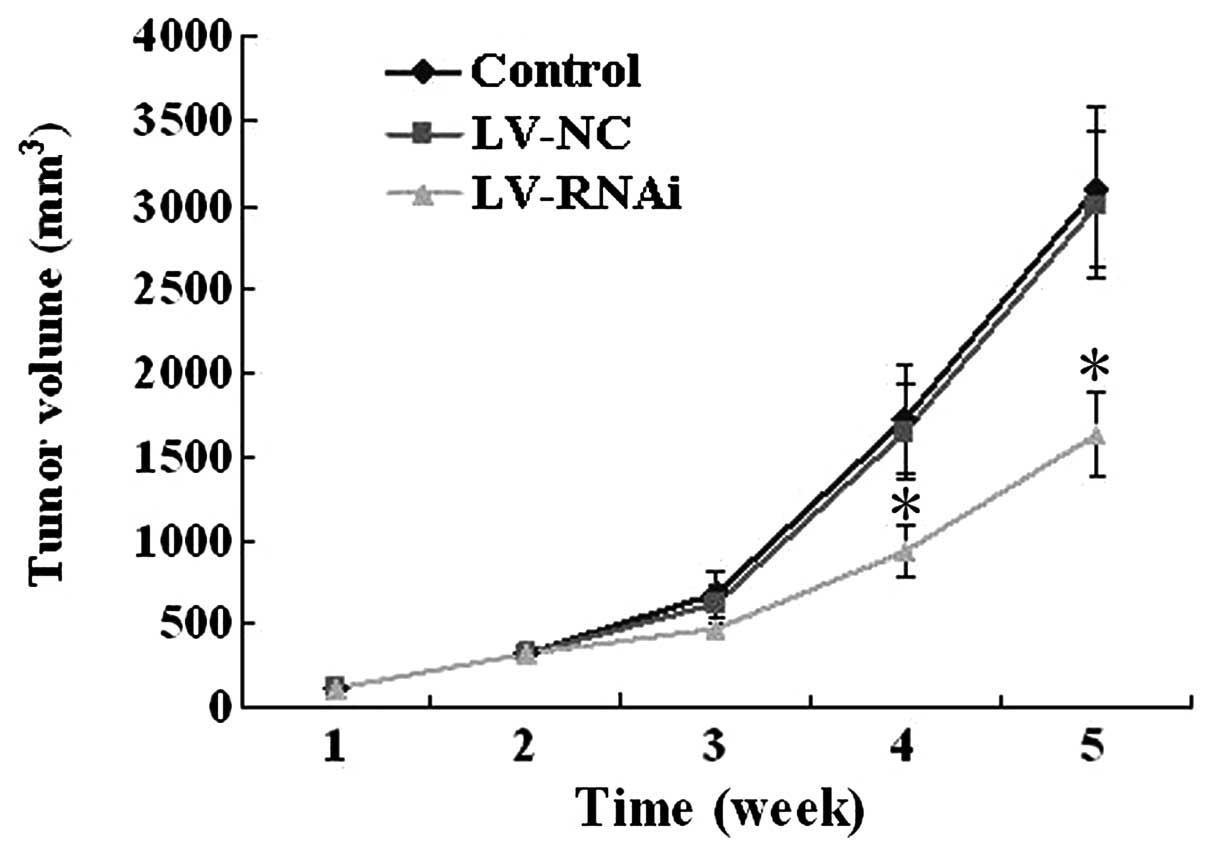
Tumor growth in the subcutaneous
transplantation model
All of the 18 mice developed detectable tumors at
the beginning of this experiment. Inhibition of growth was observed
more significantly in mice after treatment with LV-RNAi for 5
weeks, when compared to the LV-NC (3,000±430 mm3) or
control group (3,100±480 mm3). The average tumor volume
(1,630±250 mm3) in the LV-RNAi group was significantly
lower than the other two groups (P<0.05) (Fig. 6). No significant differences were
noted between the LV-NC and the control group.
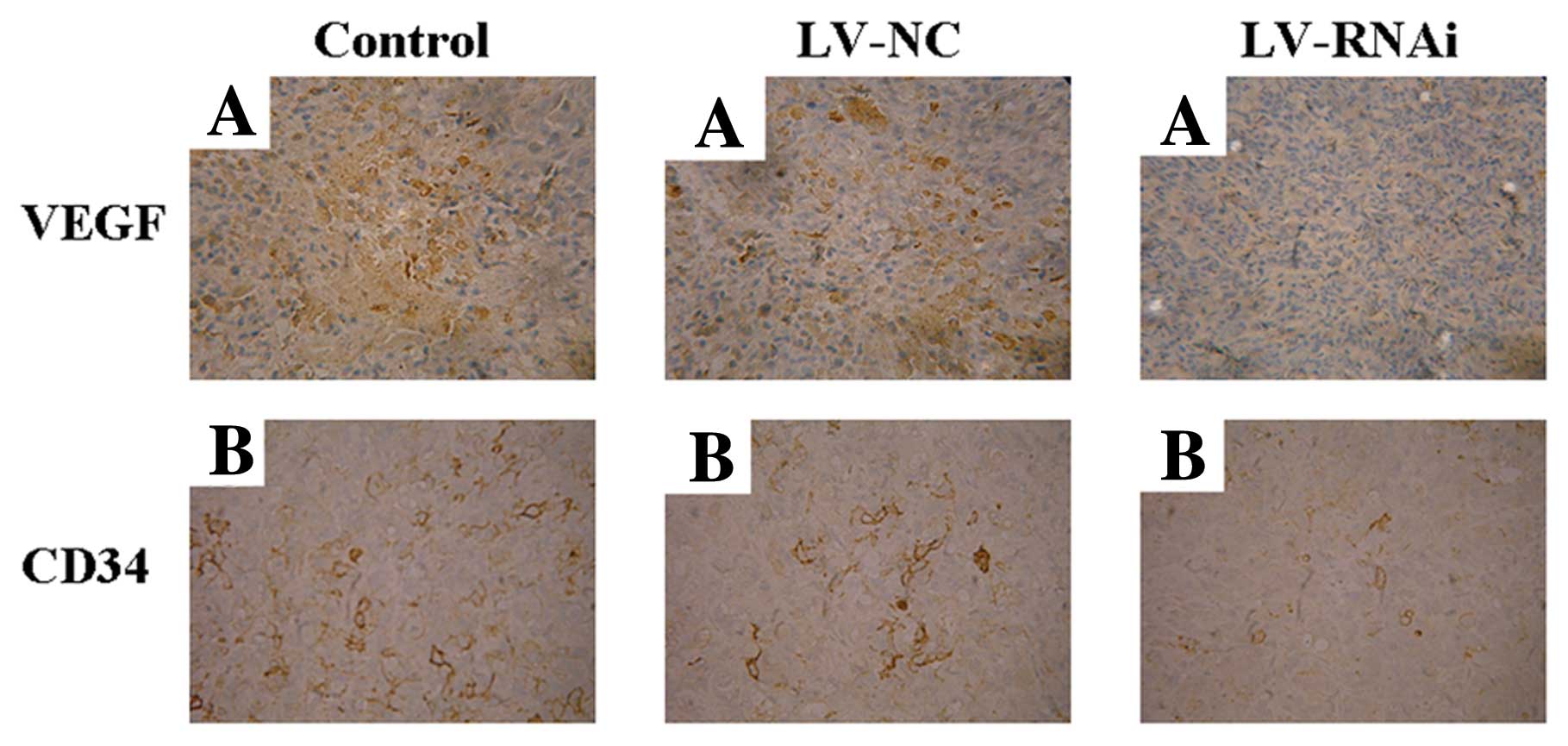
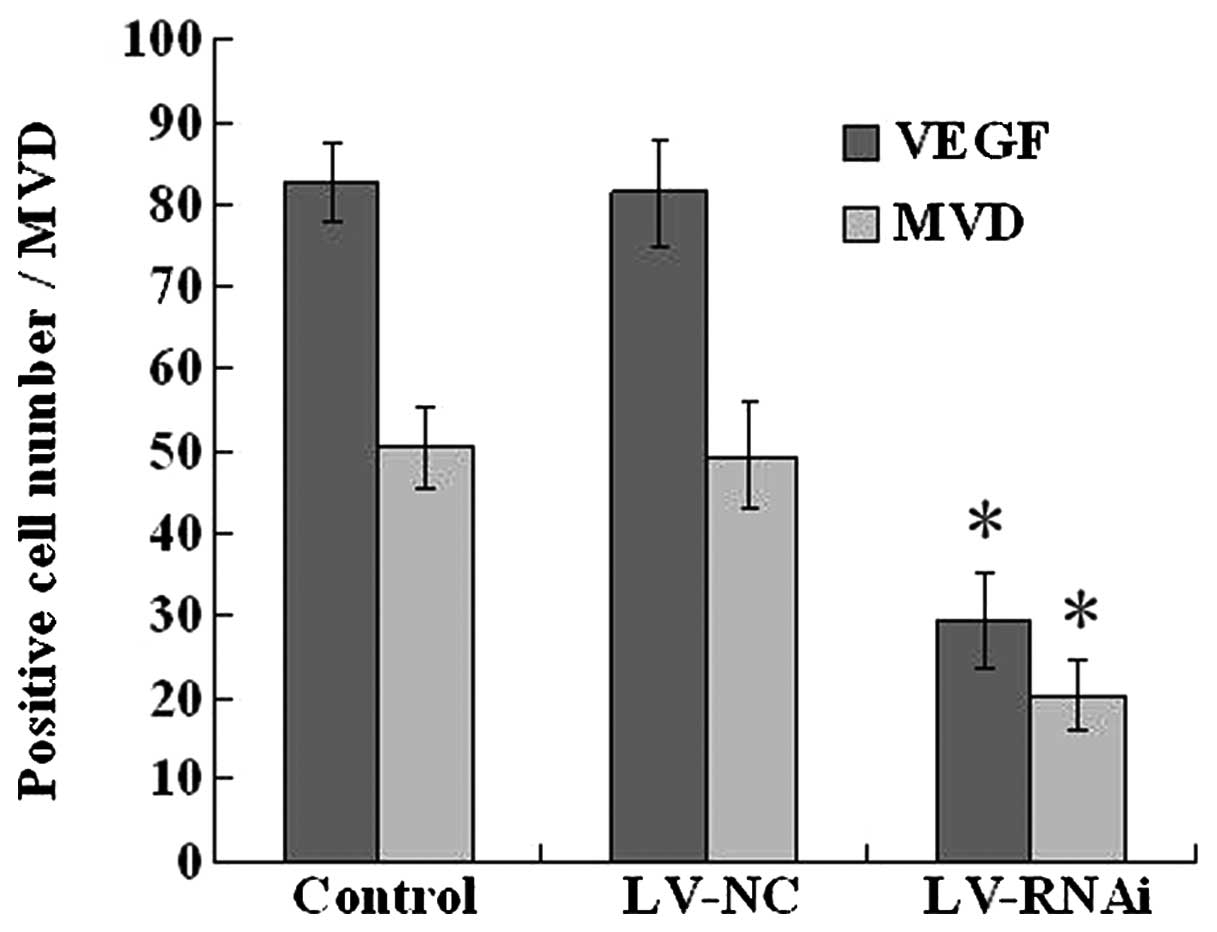
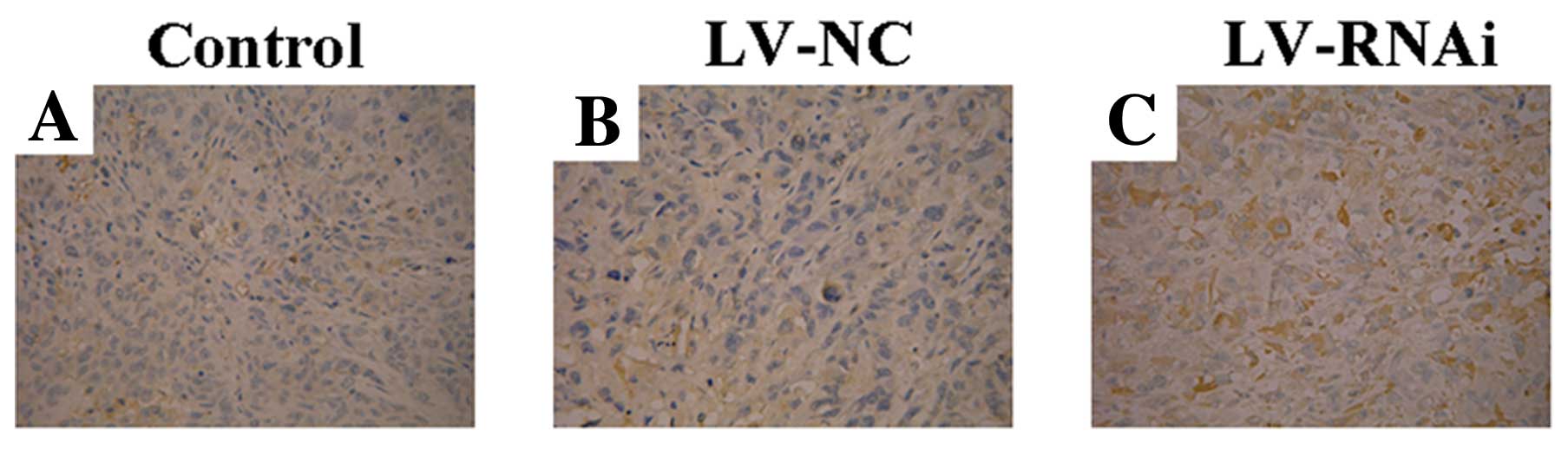
Evaluation of expression of VEGF and CD34
by immunohistochemistry
In order to demonstrate the mechanism of the
anti-angiogenic effect of LV-RNAi, the expression of VEGF and CD34
was assessed by immunohistochemistry in the nude mouse transplanted
tumors after treatments. The LV-RNAi group exhibited downregulation
of VEGF expression and a decrease in the MVD when compared to the
LV-NC and control groups (P<0.05) (Figs. 7 and 8). There were no significant differences
between the LV-NC and control group (P>0.05).
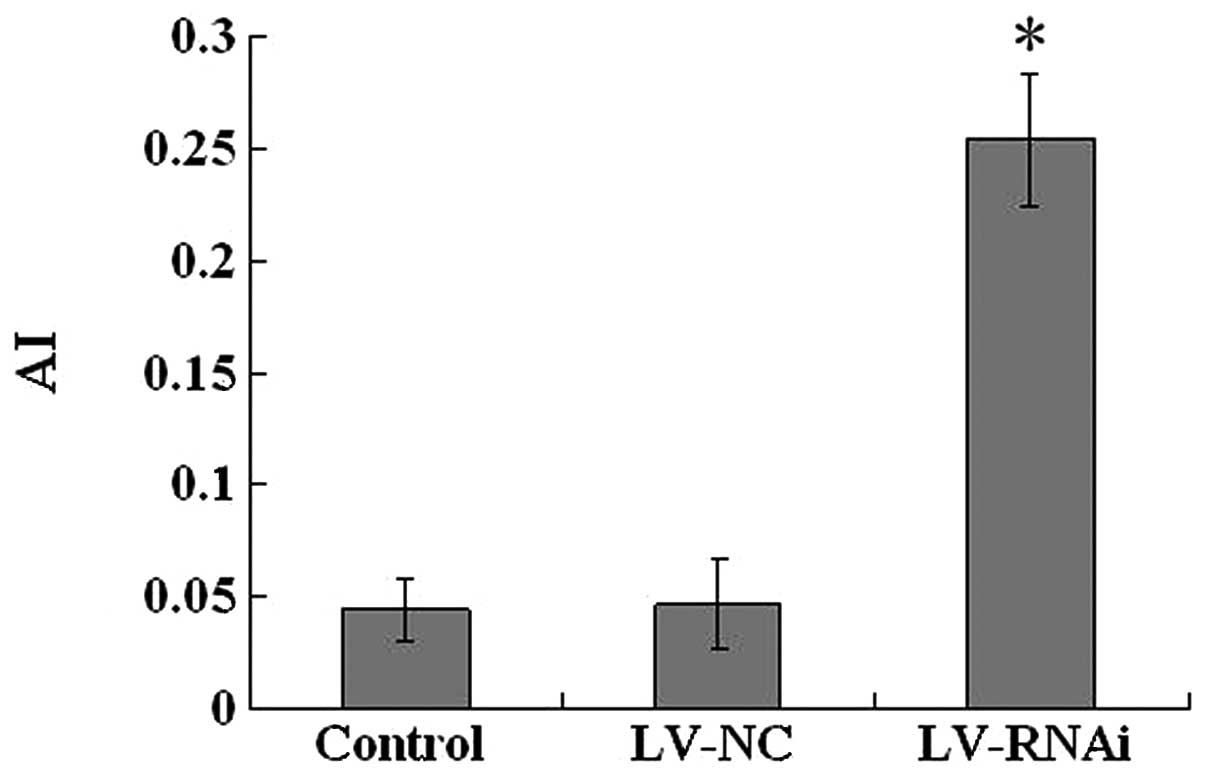
Apoptosis assay by TUNEL staining
The number of positive apoptotic tumor cells
exhibiting brown nuclei in the TUNEL assay was determined (Fig. 9). Based on the TUNEL assay, we found
that increased numbers of apoptotic pancreatic carcinoma cells were
present in the subcutaneously transplanted tumors treated with
LV-RNAi. When compared to the LV-NC (0.047±0.020) or control group
(0.044±0.014), the AI (0.254±0.029) in the LV-RNAi group was
significantly higher than that in the former two groups (P<0.05)
(Fig. 10). These results indicate
that inhibition of VEGF gene expression caused apoptotic cell death
in pancreatic carcinoma cells in vivo.
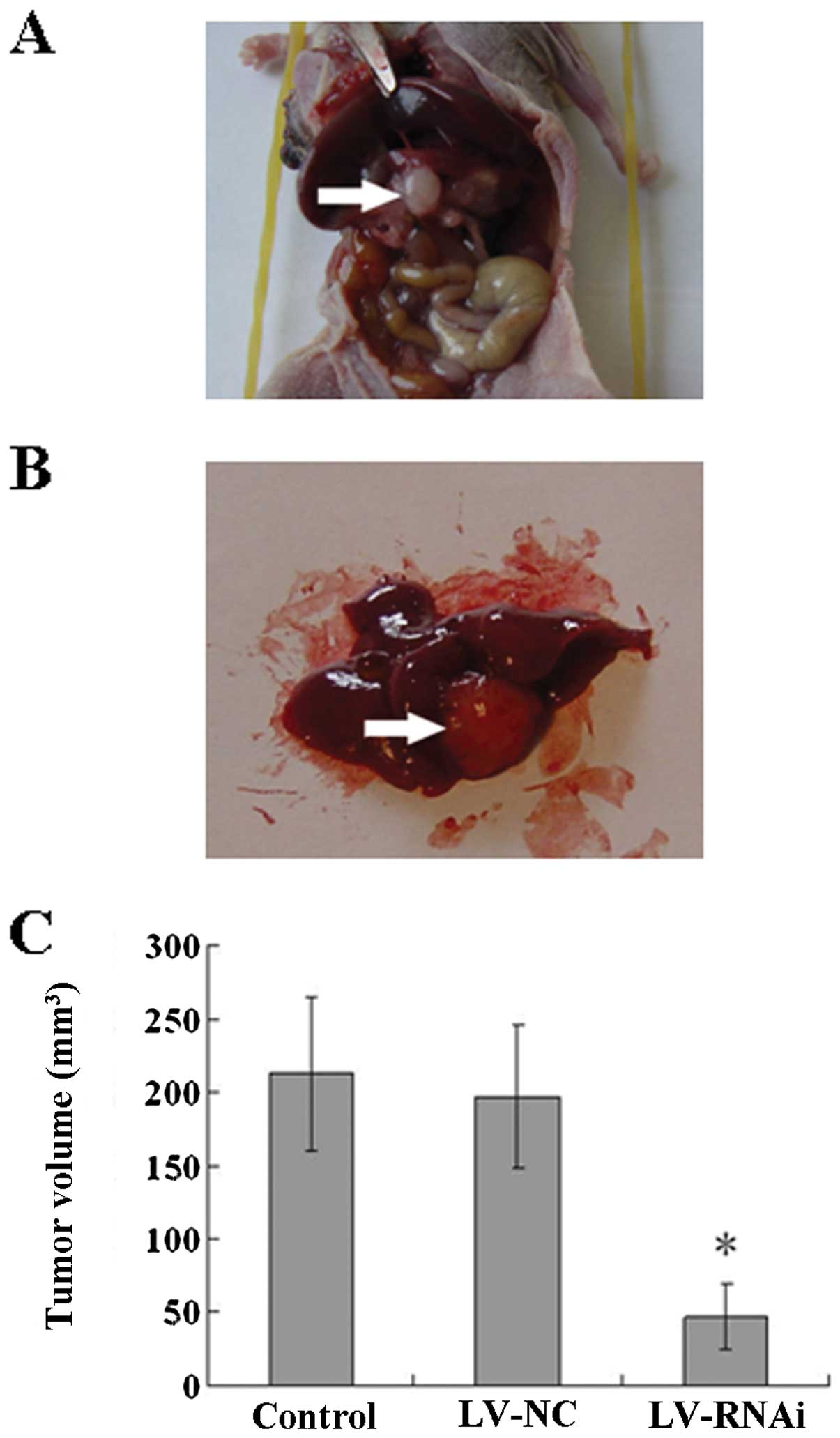
Tumor growth and liver metastasis in the
orthotopic transplantation pancreatic cancer model
All of the 18 mice developed orthotopic
transplantation pancreatic carcinoma tumors in this experiment
(Fig. 11A). Liver metastasis was
detected (Fig. 11B) in the
orthotopic transplantation pancreatic cancer model. Inhibition of
growth was significantly observed in mice following treatment with
LV-RNAi for 5 weeks, when compared to the mice treated with LV-NC
(197±49 mm3) or normal saline (213±52 mm3).
The average tumor volume (47±22 mm3, P<0.05) in the
former group was significantly lower than that in the latter two
groups (Fig. 11C). The number of
cases of liver metastasis in the LV-RNAi group (1/6, 16.67%) was
less than that in the LV-NC (3/6, 50.00%) or the control group
(4/6, 66.67%), while there was no significant differences between
the LV-NC and the control group (P=0.58).
Discussion
Pancreatic adenocarcinoma is one of the deadliest
human malignancies, accounting for more than 20% of
gastrointestinal cancer-related deaths (22,23).
At the time of diagnosis, the disease has often progressed to an
advanced stage at which surgical resection is often not a viable
option and at which time tumors are highly resistant to
conventional chemotherapy and radiation treatments (24,25).
The resistance of pancreatic adenocarcinoma to conventional
treatment strategies has led to a search for novel targeted
therapies that may be useful in eradicating this disease. Gene
therapy has been recently emphasized for its contribution to a more
favorable patient prognosis (26).
In this study, we chose a lentiviral vector since it displays high
efficiency in gene delivery and is expressed long-term. In
addition, the vector can integrate genes into non-dividing cells
with little immunologic reaction (17,18).
Our results showed that a high gene transduction efficiency
(>90%) was achieved at day 5 after exposure to lentiviral
vectors, suggesting gene integration.
Small hairpin RNA (shRNA) mimic natural RNAi in ways
that synthetic siRNA oligonucleotides do not (27). shRNA expression vector systems have
been established to induce RNA interference (RNAi) in mammalian
cells (28). Although these vectors
provide certain advantages over chemically synthesized siRNAs, some
disadvantages remain, including transient shRNA expression and low
transfection efficiency, especially in non-dividing primary cells.
To overcome these limitations, shRNA delivery systems using
retroviral vectors (29),
adenoviral vectors (30) and, more
recently, lentiviral vectors (31)
have been reported and proven safe for humans. Lentiviral vectors
encoding antisense targeting sequences have been used in clinical
trials with no obvious side effects (32,33).
Lentivirus-delivered shRNAs are capable of specific, highly stable
and functional silencing of gene expression in a variety of human
cells including primary non-dividing cells and also in transgenic
mice (34,35).
Tumor growth relies on angiogenesis, the formation
of new blood vessels, to receive an adequate supply of oxygen and
nutrients (6,7). In the absence of a blood vascular
network, tumors are restrained in size due to limits in the
diffusion of oxygen. Therefore, angiogenesis is an important
process for growth, progression and metastasis of solid tumors
(36). VEGF plays a central role in
tumor angiogenesis; it is expressed in most tumors, often at
substantially increased levels (37), which makes it a critical target for
cancer therapy (38–42). Experiments with neutralizing
antibodies and other inhibitors have demonstrated that blockade of
the VEGF pathway is sufficient to significantly suppress
angiogenesis associated with solid tumor growth in many models.
Subcutaneous and orthotopic models have been used to test the
effects of inhibitors of the VEGF/VEGFR pathway on the growth of a
variety of tumor cell lines (43).
Angiogenesis in pancreatic carcinoma is based on the same
fundamental principles of activation, proliferation and migration
of endothelial cells. The expression of VEGF and CD34 in pancreatic
carcinomas was demonstrated to be significantly higher than that in
normal pancreatic tissue samples, respectively (44,45).
In the present study, we used lentivirus-mediated
shRNA expression system targeting the VEGF gene to downregulate
gene expression. After successful lentivirus-mediated VEGF RNA
interference, the mRNA and protein expression of the VEGF gene was
virtually knocked down in vitro as detected by real-time
RT-PCR, western blot analysis and ELISA, respectively. Decreased
expression of VEGF in the human pancreatic carcinoma cell line and
in the xenografted tumors contributed to decreased angiogenesis,
growth and metastasis. CD34 is a cell surface marker of progenitor
cells and is frequently used as a new vessel marker and an
indicator of microvessel density in tissues (46,47).
Immunostaining assays revealed VEGF and CD34 in tumors were
significantly decreased after LV-RNAi transfection. As shown in the
subcutaneous and orthotopic xenografted pancreatic cancer in mice,
downregulation of VEGF was found to lead to the suppression of
cancer growth, resulting in reduced tumor size.
Additionally, overexpression of VEGF has been shown
to be associated with enhanced tumorigenicity and tumor metastatic
potential (8,48). Evidence now indicates that the
direct receptor-mediated effects of VEGF on tumors such as VEGF
induction of phosphatidylinositol-3′-kinase (PI3K) and Akt and
enhancement of cell survival (49,50),
affect clinical outcomes more decisively than do changes in blood
flow and/or oxygenation (51). As
is now recognized for co-expression of the kinase-impaired HER3
with HER2, (52) co-expression of
kinase-defective VEGFR-1 with active VEGFR-2 increases Akt
signaling (53) thus enhancing cell
survival and tumor invasion (48).
The pro-apoptotic effect of VEGF signaling inhibition is also
apparent in vitro(54),
confirming that the pro-apoptotic action is at least in part
independent of the blood supply. In the TUNEL assay of the
subcutaneous xenografted tumors, we found that decreased VEGF
expression increased cell apoptosis of pancreatic carcinoma cells.
Moreover, VEGF-dependent signaling cascades were found to increase
cell motility via Src or Fak inducible tyrosine phosphorylation of
adhesion substrates (55) and such
motility is directly inducible in vitro by VEGF (56). In the orthotopic transplantation
pancreatic cancer model, we found that decreased VEGF expression
reduced the liver metastasis rate, although there was no
significant difference detected by statistics which may have been
due to the small sample size. Thus, more animal experiments nust be
performed to further confirm the inhibitory effects of metastasis
mediated by lentivirus-mediated shRNA interference targeting VEGF
in pancreatic cancer in vivo. Therefore, the anticancer
effects induced by lentivirus-mediated shRNA interference targeting
VEGF require further investigation, and this will be the focus of
our interest in future research.
In conclusion, our findings indicate that
lentivirus-mediated shRNA interference targeting VEGF potently
suppressed angiogenesis, growth and increased cell apoptosis in the
Patu8988 pancreatic cancer cell line in vivo. Our findings
support the theory that lentivirus-mediated shRNA interference
targeting VEGF may be a promising mothod for the treatment of
pancreatic carcinoma.
Acknowledgements
This study was supported by a grant from the
Post-graduate Scientific Research Innovation Project of the
Education Department of Jiangsu Province (no. CXZZ11_0125), China,
and the Science and Technology Research Project of the Science and
Technology Bureau of Suzhou City (no. SYS201120), China.
References
|
1
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics, 2007. CA Cancer J Clin. 57:43–66. 2007. View Article : Google Scholar
|
|
2
|
Neoptolemos JP, Cunningham D, Friess H, et
al: Adjuvant therapy in pancreatic cancer: historical and current
perspectives. Ann Oncol. 14:675–692. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar
|
|
4
|
Ghaneh P, Costello E and Neoptolemos JP:
Biology and management of pancreatic cancer. Postgrad Med J.
84:478–497. 2008. View Article : Google Scholar
|
|
5
|
Pan X, Sheng W, Zhu Q, et al: Inhibition
of pancreatic carcinoma growth by adenovirus-mediated human
interleukin-24 expression in animal model. Cancer Biother
Radiopharm. 23:425–434. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Folkman J: Tumor angiogenesis: a possible
control point in tumor growth. Ann Intern Med. 82:96–100. 1975.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kerbel RS: Tumor angiogenesis. New Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dai J and Rabie A: VEGF: an essential
mediator of both angiogenesis and endochondral ossification. J Dent
Res. 86:937–950. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Midgley R and Kerr D: Bevacizumab: current
status and future directions. Ann Oncol. 16:999–1004. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Weidner N, Folkman J, Pozza F, et al:
Tumor angiogenesis: a new significant and independent prognostic
indicator in early-stage breast carcinoma. J Natl Cancer Inst.
84:1875–1887. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Olson TA, Mohanraj D, Carson LF, et al:
Vascular permeability factor gene expression in normal and
neoplastic human ovaries. Cancer Res. 54:276–280. 1994.PubMed/NCBI
|
|
12
|
Seo Y, Baba H, Fukuda T, et al: High
expression of vascular endothelial growth factor is associated with
liver metastasis and a poor prognosis for patients with ductal
pancreatic adenocarcinoma. Cancer. 88:2239–2245. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
El-Houseini ME, Abdel-Azim SA, El-Desouky
GI, et al: Clinical significance of vascular endothelial growth
factor (VEGF) in sera of patients with pediatric malignancies. J
Egypt Natl Canc Inst. 16:57–61. 2004.PubMed/NCBI
|
|
14
|
Rutkowski P, Kamińska J, Kowalska M, et
al: Cytokine and cytokine receptor serum levels in adult bone
sarcoma patients: correlations with local tumor extent and
prognosis. J Surg Onco1. 84:151–159. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Merritt WM, BarEli M and Sood AK: The
dicey role of dicer: implications for RNAi therapy. Cancer Res.
70:2571–2574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Agrawal N, Dasaradhi PV, Mohmmed A, et al:
RNA interference: biology, mechanism, and applications. Microbiol
Mol Biol Rev. 67:657–685. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Buchschacher GL and Wong-Staal F:
Development of lentiviral vectors for gene therapy for human
diseases. Blood. 95:2499–2504. 2000.PubMed/NCBI
|
|
18
|
Naldini L: Medicine. A comeback for gene
therapy. Science. 326:805–806. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li L, Zhang R, Cen JN, et al: Construction
and identification of lentiviral vector-mediated RNA interference
of VEGF gene. Soochow Univ J Med Sci. 28:20–22. 2008.(abstract in
English).
|
|
20
|
Li L, Zhang R, Cen JN, et al:
Lentivirus-mediated RNA interference targeting vascular endothelial
growth factor gene enhances the sensitivity of K562 cells to STI
571. Chin J Pathophysiol. 25:1122–1126. 2009.(abstract in
English).
|
|
21
|
Weidner N: Intratumor microvessel density
as a prognostic factor in cancer. Am J Pathol. 147:9–19.
1995.PubMed/NCBI
|
|
22
|
Korc M: Pathways for aberrant angiogenesis
in pancreatic cancer. Mol Cancer. 2:2–8. 2003. View Article : Google Scholar
|
|
23
|
Shi X, Friess H, Kleeff J, et al:
Pancreatic cancer: factors regulating tumor development,
maintenance and metastasis. Pancreatol. 1:517–524. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Keleg S, Büchler P, Ludwig R, et al:
Invasion and metastasis in pancreatic cancer. Mol Cancer. 2:142003.
View Article : Google Scholar
|
|
25
|
MacKenzie MJ: Molecular therapy in
pancreatic adenocarcinoma. Lancet Oncol. 5:541–549. 2004.
View Article : Google Scholar
|
|
26
|
Jimeno A and Hidalgo M: Molecular
biomarkers: their increasing role in the diagnosis,
characterization, and therapy guidance in pancreatic cancer. Mol
Cancer Ther. 5:787–796. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Snove O and Rossi JJ: Expressing short
hairpin RNAs in vivo. Nat Methods. 3:689–695. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Brummelkamp TR, Bernards R and Agami R: A
system for stable expression of short interfering RNAs in mammalian
cells. Science. 296:550–553. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Brummelkamp TR, Bernards R and Agami R:
Stable suppression of tumorigenicity by virus-mediated RNA
interference. Cancer Cell. 2:243–247. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xia H, Mao Q, Paulson HL, et al:
siRNA-mediated gene silencing in vitro and in vivo. Nat Biotechnol.
20:1006–1010. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qin XF, An DS, Chen IS, et al: Inhibiting
HIV-1 infection in human T cells by lentiviral-mediated delivery of
small interfering RNA against CCR5. Proc Natl Acad Sci USA.
100:183–188. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Manilla P, Rebello T, Afable C, et al:
Regulatory considerations for novel gene therapy products: a review
of the process leading to the first clinical lentiviral vector. Hum
Gene Ther. 16:17–25. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bank A, Dorazio R and Leboulch P: A phase
I/II clinical trial of beta-globin gene therapy for
beta-thalassemia. Ann NY Acad Sci. 1054:308–316. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nishitsuji H, Ikeda T, Miyoshi H, et al:
Expression of small hairpin RNA by lentivirus-based vector confers
efficient and stable gene-suppression of HIV-1 on human cells
including primary non-dividing cells. Microbes Infect. 6:76–85.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rubinson DA, Dillon CP, Kwiatkowski AV, et
al: A lentivirus-based system to functionally silence genes in
primary mammalian cells, stem cells and transgenic mice by RNA
interference. Nat Genet. 33:401–406. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Folkman J: Tumor angiogenesis: therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim KJ, Li B, Winer J, et al: Inhibition
of vascular endothelial growth factor-induced angiogenesis
suppresses tumor growth in vivo. Nature. 362:841–844. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sullivan LA and Brekken RA: The VEGF
family in cancer and antibody-based strategies for their
inhibition. MAbs. 2:165–175. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dvorak HF: Vascular permeability
factor/vascular endothelial growth factor: a critical cytokine in
tumor angiogenesis and a potential target for diagnosis and
therapy. J Clin Oncol. 20:4368–4380. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Roskoski R Jr: Vascular endothelial growth
factor (VEGF) signaling in tumor progression. Critical Rev Oncol
Hematol. 62:179–213. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Presta LG, Chen H, O’Connor SJ, et al:
Humanization of an anti-VEGF monoclonal antibody for the therapy of
solid tumors and other disorders. Cancer Res. 57:4593–4599.
1997.PubMed/NCBI
|
|
42
|
Ferrara N: Vascular endothelial growth
factor: basic science and clinical progress. Endocr Rev.
25:581–611. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ferrara N: VEGF-A: a critical regulator of
blood vessel growth. Eur Cytokine Netw. 20:158–163. 2009.PubMed/NCBI
|
|
44
|
Itakura J, Ishiwata T, Shen B, et al:
Concomitant over-expression of vascular endothelial growth factor
and its receptors in pancreatic cancer. Int J Cancer. 85:27–34.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sun HC, Qiu ZJ, Liu J, et al: Expression
of hypoxia-inducible factor-1 alpha and associated proteins in
pancreatic ductal adenocarcinoma and their impact on prognosis. Int
J Oncol. 30:1359–1367. 2007.PubMed/NCBI
|
|
46
|
Krause DS, Fackler MJ, Civin CI, et al:
CD34: structure, biology, and clinical utility. Blood. 87:1–13.
1996.PubMed/NCBI
|
|
47
|
Nielsen JS and McNagny KM: Novel functions
of the CD34 family. J Cell Sci. 121:3683–3692. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Girling JE and Rogers PA: Regulation of
endometrial vascular remodelling: role of the vascular endothelial
growth factor family and the angiopoietin-TIE signalling system.
Reproduction. 138:883–893. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wu Y, Hooper AT, Zhong Z, et al: The
vascular endothelial growth factor receptor (VEGFR-1) supports
growth and survival of human breast carcinoma. Int J Cancer.
119:1519–1529. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Graells J, Vinyals A, Figueras A, et al:
Overproduction of VEGF concomitantly expressed with its receptors
promotes growth and survival of melanoma cells through MAPK and
PI3K signaling. J Invest Dermatol. 123:1151–1161. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Mercurio AM, Lipscomb EA and Bachelder RE:
Non-angiogenic functions of VEGF in breast cancer. J Mammary Gland
Biol Neoplasia. 10:283–290. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sergina NV, Rausch M, Wang D, et al:
Escape from HER-family tyrosine kinase inhibitor therapy by the
kinase-inactive HER3. Nature. 445:437–441. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wu Y, Zhong Z, Huber J, et al:
Anti-vascular endothelial growth factor receptor-1 antagonist
antibody as a therapeutic agent for cancer. Clin Cancer Res.
12:6573–6584. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Krause S, Förster Y, Kraemer K, et al:
Vascular endothelial growth factor antisense pretreatment of
bladder cancer cells significantly enhances the cytotoxicity of
mitomycin C, gemcitabine and cisplatin. J Urol. 174:328–331. 2005.
View Article : Google Scholar
|
|
55
|
Lesslie DP, Summy JM, Parikh NU, et al:
Vascular endothelial growth factor receptor-1 mediates migration of
human colorectal carcinoma cells by activation of Src family
kinases. Br J Cancer. 94:1710–1717. 2006.PubMed/NCBI
|
|
56
|
Wey JS, Fan F, Gray MJ, et al: Vascular
endothelial growth factor receptor-1 promotes migration and
invasion in pancreatic carcinoma cell lines. Cancer. 104:427–438.
2005. View Article : Google Scholar : PubMed/NCBI
|