Introduction
Autotaxin is an extracellular phosphodiesterase and
a cell autocrine motility-stimulating factor initially isolated
from human malignant melanoma A2058 cell medium and purified by
M.L. Stracke. Autotaxin (ATX) is a 125-kDa glycoprotein reported to
stimulate both random or directed motility of melanoma cells
(1).
Protein sequence analysis indicates that there
exists a significant homology between ATX and hemolytic
phospholipase D on the cell surface (2). Hydrolysis of lysophosphatidylcholine
by ATX in lysophosphatidic acid occurs when ATX is released into
the extracellular space with deletion of amino acid (2,3).
Recently, ATX was found to be closely linked to angiogenesis in
embryo development. In previous studies, ATX expression was
investigated in thyroid carcinoma (4), malignant melanoma (5), breast carcinoma (6–8),
non-small cell lung carcinoma (9),
renal cell carcinoma (10) and
neuroblastoma (11), while the role
of ATX in hepatic carcinoma (12)
has been documented in China. Matrix metalloproteinases MMP2 and
MMP3, reported to be closely associated with gastric cancer
infiltration and lymphatic metastasis, are used as molecular
markers for detecting and evaluating the prognosis of gastric
cancer (13,14).
In our in vitro study, the plasmid vectors
containing shRNA fragments targeting ATX were transfected into the
human gastric cancer cell line AGS, to prove that shRNA effectively
and consistently inhibits ATX mRNA and protein expression in AGS
cells and reduces their abilities for proliferation, migration and
invasion. Our study also compared and observed in vivo the
inhibitory effect of shRNA on the growth of human gastric cancer
xenografts in nude mice. These observations indicate the
relationship between ATX and gastric cancer infiltration and
metastasis, thereby providing the laboratory data for clarifying
the molecular mechanisms of gastric cancer development and
treatment.
Materials and methods
Human cell lines and culture
The human gastric cancer cell line AGC was cultured
in Dulbecco’s modified Eagle’s medium (DMEM), supplemented with 10%
foetal bovine serum (Invitrogen, Carlsbad, CA, USA) and incubated
at 37°C in 5% CO2.
RNA extraction, cDNA synthesis and
RT-PCR
Total RNA was extracted from cell pellets by TRIzol
(Invitrogen) and reverse transcribed into cDNA using Reverse
Transcriptase (Applied Biosystems, Foster City, CA, USA) according
to the manufacturer’s instructions. The target gene expression was
determined by RT-PCR using specific primers of target genes (5′-GTT
GCA AGG AAA CCT TTG GA-3′; 5′-AAC TTC CTC TGG CAT GGT TG-3′). GAPDH
served as an internal control for total cDNA content. Samples were
amplified using the ABI Prism 7700 Sequence Detection system
(Applied Biosystems).
Cell proliferation assay
The cells were cultured in well plates
(~2×103/well). After culture for 24, 48, 72 and 96 h, 20
μl of MTT solution was added to each well. After another 4 h of
culture, the supernatant was discarded and 150 μl of DMSO was added
to each well while shaking at room temperature for 10 min, so that
the crystals could be completely dissolved. The assessment
procedure was repeated 3 times and the average value was determined
from each well as the absorbance value at 490 nm on a microplate
reader.
Cell migration and invasion assays
In this assay, an 8-μm pore-sized polycarbonate
membrane Transwell chamber (24-well plate) purchased from Corning
was used. After incubation for 1 h at 37°C in a basolateral chamber
containing 600 μl RPMI-1640 medium with 20% FCS, the different
gastric cancer cells were washed with PBS for 3 times. After
trypsin digestion, the cells were resuspended into RPMI-1640 medium
with 10% FBS. Subsequently, a 100-μl cell suspension
(1×106/ml cells) was added to each well of the apical
chamber and incubated in 5% CO2 at 37°C. The cells which
were unable to pass through the microporous membrane were wiped
from the apical chamber with cotton swabs. Following a further wash
in PBS and fixation with 4% formalin, cells on the surface of the
microporous membrane were counted randomly in five ×200 microscopic
fields using Giemsa staining. This procedure was repeated 3 times
for each sample, and the cell migratory ability was determined
based on the number of migrating cells (based on the average value
in the assay). In the cell invasion assay, as distinguished from
the cell migration assay, the extracellular matrix Matrigel (BD
Pharmingen, San Diego, CA, USA) was additionally used to simulate
the in vivo extracellular matrix environment. Matrigel at 50
μl/cm2 was added to the surface of the polycarbonate
membrane in the Transwell chamber, and then placed under a fume
hood either at 37°C or at room temperature for 30 min until the
Matrigel gelled. The procedure was then performed identical to the
migration assay. The cell invasive ability was determined based on
the number of invading cells.
Establishment of the tumor xenograft
model
Twenty mice were randomly divided into 4 groups
(n=5), inoculated with the respective cells: wild-type (WT) group,
transfection targeted interference vector pSUPER-ATX group,
negative control group pSUPER-mock and blank plasmid group
pSUPER-control. The mice were subcutaneously inoculated in the
right hind lateral leg with the cells (2×107/ml) in
logarithmic growth period. Then every 3 days after tumorigenesis in
the nude mice, the measurements of the tumor diameter (a) and short
diameter (b) with a vernier caliper were used to calculate the
tumor volume according to the formula (a × b2/2), and
the tumor growth curves were drawn on the basis of the average
values of the tumor volume from each group. In the 8th week when
the mice were sacrificed, the tumors were extracted in order to
measure the final volume and weight by using a photoelectric
balance. The tumor growth inhibition rate was determined using the
volume and weight. The tumor tissue of each group was stored in
liquid nitrogen. Part of the tissue was fixed with 10% formaldehyde
solution in order to be used for H&E staining,
immunohistochemistry and western blot experiments.
Statistical methods
Data are presented as the means ± SD. Analysis of
the significance of differences between 2 groups was performed by
the two tailed Student’s t-test and one-way ANOVA analysis.
P<0.05 was considered to indicate a statistically significant
result.
Results
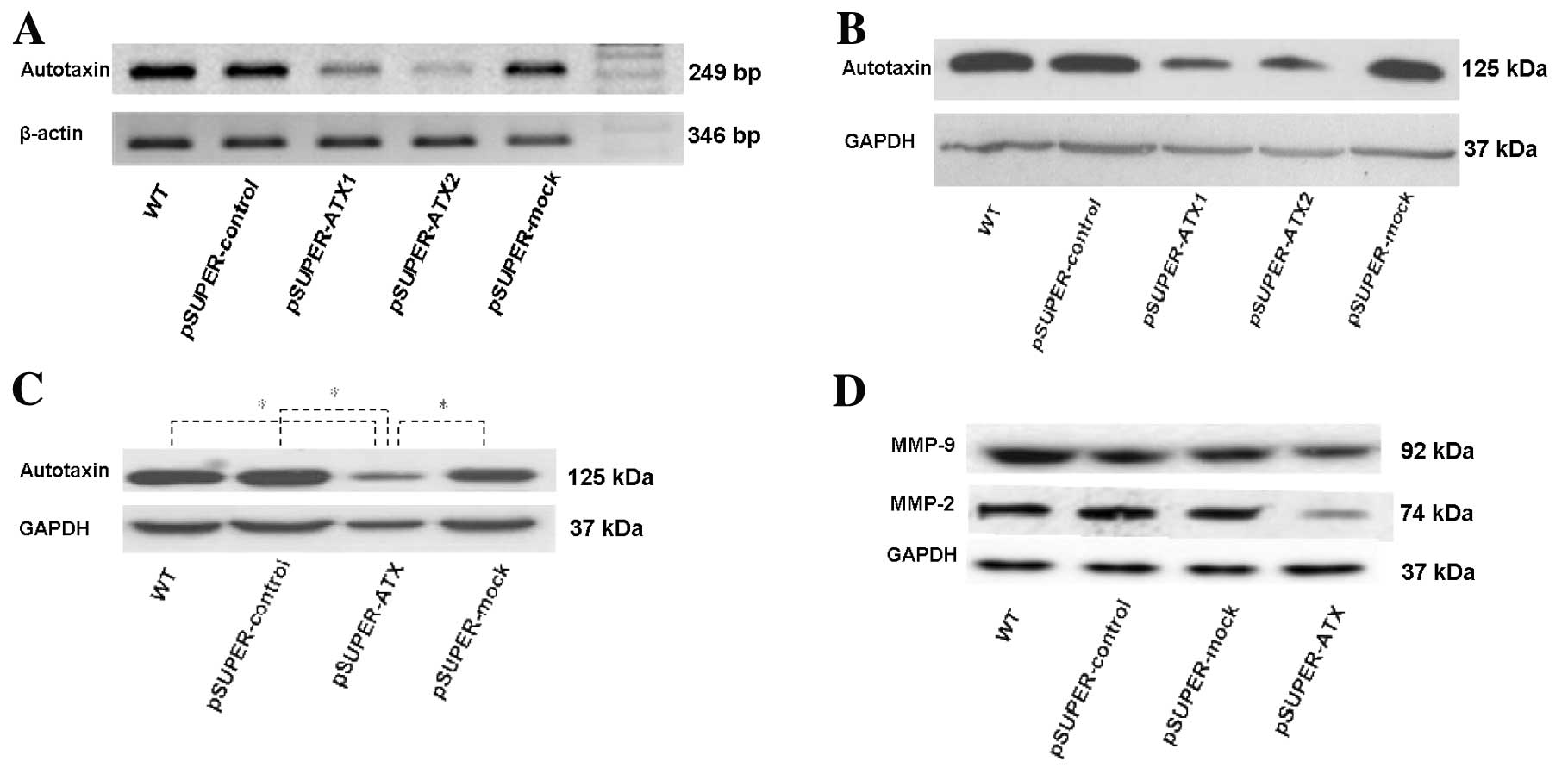
Differential ATX expression in AGS WT and
transfected cells
RT-PCR was used to detect changes in ATX mRNA
expression and showed that ATX mRNA expression in the transfected
cells in the two cloned cell groups pSUPER-ATX1 and pSUPER-ATX2
decreased 79.34±9.95 and 95.42±1.93%, respectively, in comparison
with WT cells and expression levels were was also markedly
inhibited by shRNA (t=7.247, P<0.01; t=14.391, P<0.01;
Fig. 1A), while there was no
significant difference in ATX mRNA expression among the WT,
pSUPER-control and pSUPER-mock groups.
Western blot analysis was used to detect changes in
ATX protein expression and showed that the ATX protein expression
of the transfected cells in the two cloned cell groups pSUPER-ATX1
and pSUPER-ATX2 decreased 78.45±5.45 and 85.42±3.56%, respectively,
in comparison with the WT cells. ATX protein expression levels were
also markedly inhibited by shRNA (t=8.457, P<0.01; t=11.936,
P<0.01; Fig. 1B), while there
was no significant difference in ATX protein expression between the
WT and negative control groups. pSUPER-ATX2 was found to be
superior to pSUPER-ATX1 for inhibiting the endogenous ATX
expression. Therefore, the pSUPER-ATX2 cell clone was selected for
further in vitro and in vivo investigations.
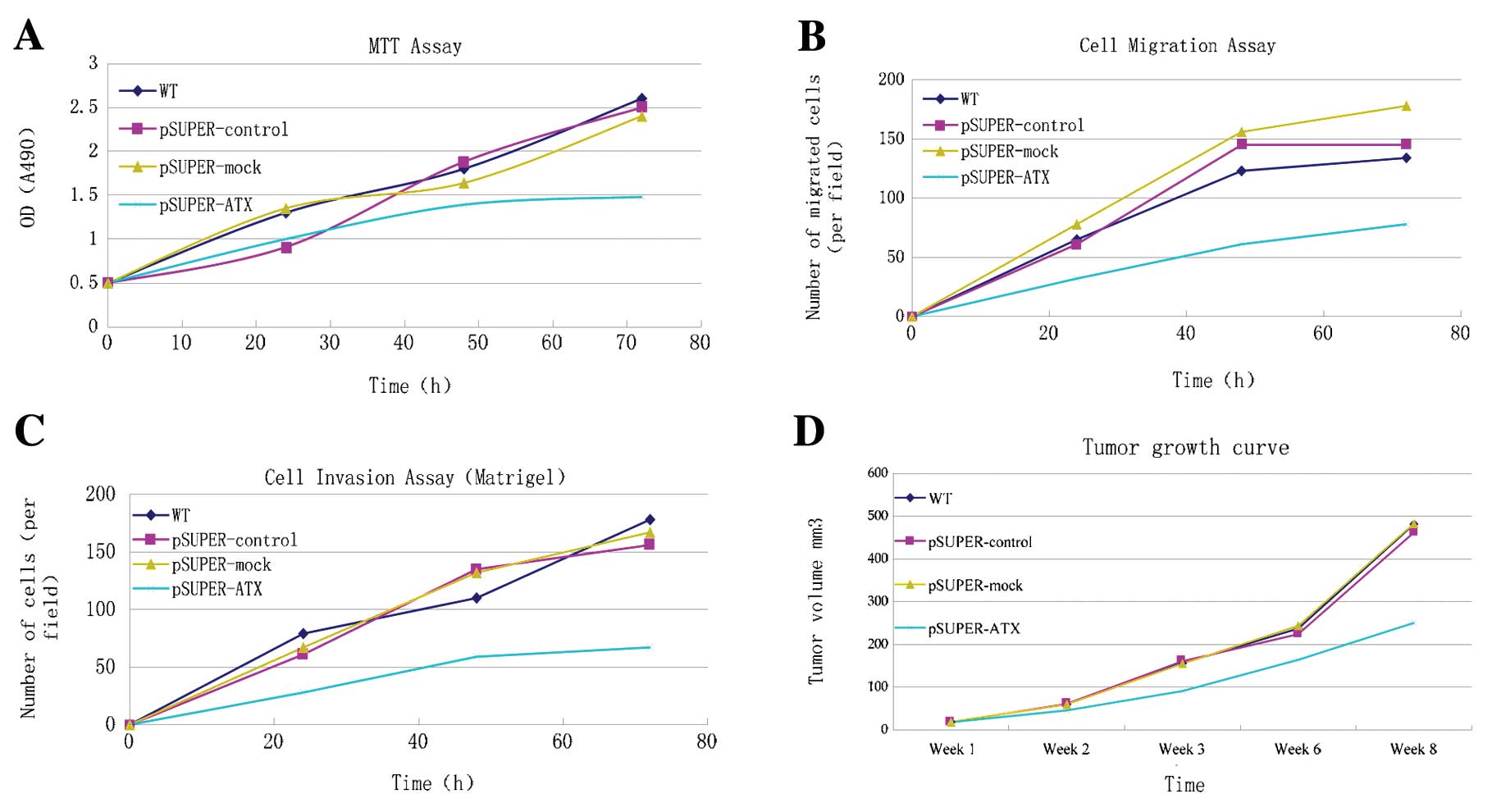
Cell proliferation
MTT assay was performed to detect cell
proliferation. As shown in Fig. 2A,
the cell proliferative ability in the pSUPER-ATX group was
constantly reduced 50 h after transfection, compared with the WT,
pSUPER-control and pSUPER-mock groups. The number of living cells
varied among the different groups which was propotional to the
absorbance value. pSUPER-ATX had an inhibitory effect on AGS cell
proliferation despite the statistical insignificance.
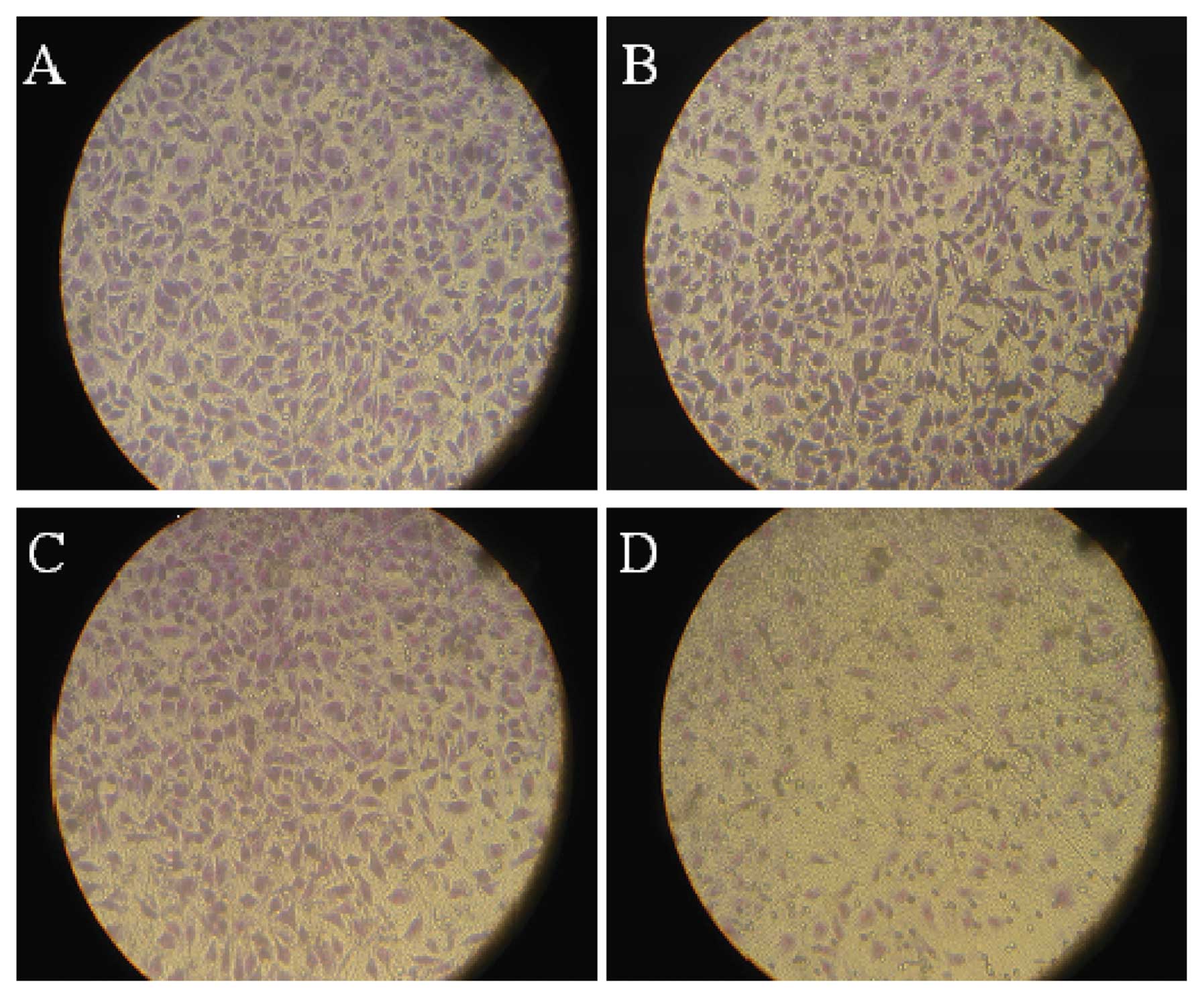
Cell migration and invasion
Cell migratory and invasive abilities were detected
using Transwell and Matrigel assays (Fig. 3). From 20 h after transfection,
migrating and invading cells in the pSUPER-ATX group decreased when
compared with the WT, pSUPER-control and pSUPER-mock groups. From
48 to 72 h after transfection, the decrease in migrating and
invading cells in the pSUPER-ATX group was significant but not in
the WT, pSUPER-control and pSUPER-mock groups. As shown in Fig. 2B and C, the number of migrating
cells decreased to 67.63±12.03% (t=15.487, P<0.01) and invading
cells to 68.02±15.63% (t=9.417, P<0.01). This indicated that
pSUPER-ATX had an inhibitory effect on both migration and
invasion.
Western blot analysis
ATX protein expression in AGS cells was detected by
western blot analysis. The internal control GAPDH was described as
a homogeneous and positively expressed band, compared with the
heterogeneously expressed band of ATX. ATX showed higher expression
in the WT, pSUPER-control, pSUPER-mock, control and transfection
groups, whereas ATX expression in the recombinant plasmid
transfection group was decreased. Their differences were
significant (P<0.05) (Fig. 1C).
In comparion with the other groups, the endogenous protein
expression of MMP-9, and particullarly, MMP-2 in the pSUPER-ATX
group was noticeably reduced. The difference was significant
(P<0.05) (Fig. 1D).
Growth of xenograft tumors in nude
mice
Within an average of 8 days, a 3- to 4-mm diameter
tumor developed at the subcutaneous injection sites of the right
hind lateral leg of the nude mice with a 100% tumor formation rate
(Fig. 4). Continuous observations
during an 8-week period were carried out and a tumor growth curve
was plotted (Fig. 2D). From the
16th day, the tumor volume between the pSUPER-ATX and the WT,
pSUPER-control and pSUPER-mock groups showed a statistically
significant difference (P<0.05); by the 8th week, a tumor size
of 477.1±8.4, 461.4±8.5, 483.6±5.2 and 248.2±6.9 mm3
was, respectively, recorded in the WT, pSUPER-control, pSUPER-mock
and pSUPER-ATX groups (Table I).
The tumor size of the pSUPER-ATX group was much smaller than the
tumor sizes in the WT, pSUPER-control and pSUPER-mock groups
(P<0.01), and no significant difference was detected between the
negative and blank group (P>0.05). Nude mice in each group had a
normal diet, good mental state, and exhibited no toxicity
throughout the feeding process with inhibition of AGS proliferation
by targeting shRNA. Until the sacrifice of nude mice in the 8th
week, the average weight of the transplanted tumor in each group
was 2.21±0.21, 2.54±0.17, 2.36±0.15 and 0.97±0.23 g. The inhibitory
rates of tumor volume and weight in the interference pSUPER-ATX
group were 52.1 and 52.3%, respectively (Table II).
 | Table IComparison of the xenograft tumor
volume (mm3) of each mouse group at different time
periods. |
Table I
Comparison of the xenograft tumor
volume (mm3) of each mouse group at different time
periods.
| Group | Week 1 | Week 2 | Week 3 | Week 6 | Week 8 |
|---|
| WT | 18.1±0.3 | 59.3±0.2 | 155.1±4.1 | 238.2±6.3 | 477.1±8.4 |
| pSUPER-control | 18.2±0.5 | 60.2±1.1 | 159.1±4.3 | 221.3±5.1 | 461.4±8.5 |
| pSUPER-mock | 18.6±0.2 | 58.9±0.9 | 154.2±2.4 | 241.9±5.8 | 483.6±5.2 |
| pSUPER-ATX | 17.9±0.2 | 45.2±0.8 | 90.7±3.5a | 163.5±6.2a | 248.2±6.9a |
| Table IIParameters of the tumors at the 8th
week following sacrifice of the nude mice. |
Table II
Parameters of the tumors at the 8th
week following sacrifice of the nude mice.
| Group | BWC/g | Tumor weight (g) | Inhibition rate
(%) |
|---|
| WT | 3.5 | 2.54±0.17 | 0.0 |
| pSUPER-control | 2.9 | 2.21±0.21 | 13.0 |
| pSUPER-mock | 3.1 | 2.36±0.15 | 7.0 |
| pSUPER-ATX | 2.8 | 0.97±0.23 | 61.8 |
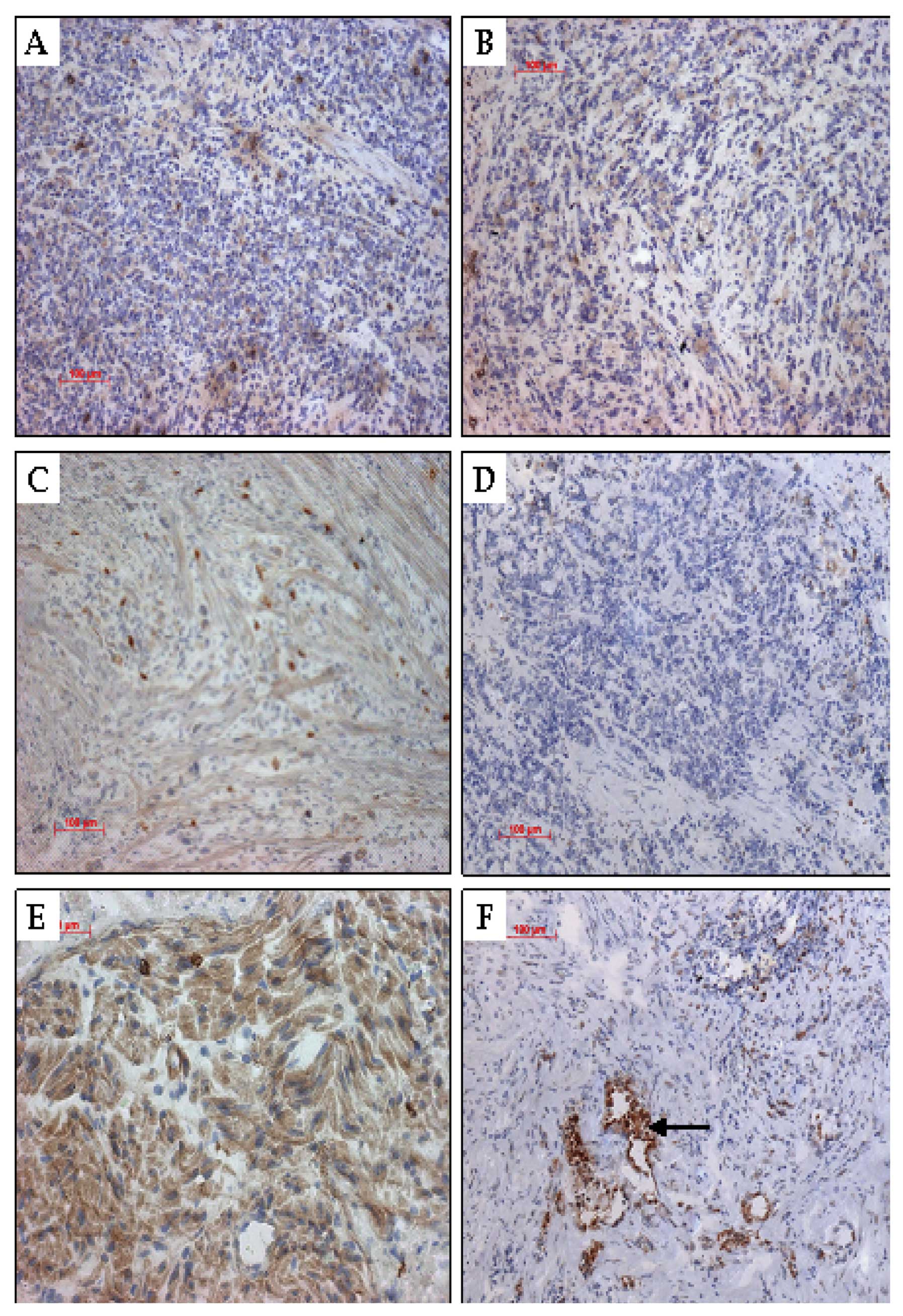
Immunohistochemical analysis of the
expression of ATX protein in the xenograft tumors
H&E staining was used to demonstrate the
inhibition of expression of the ATX protein by targeting shRNA. We
observed malformed, large, deeply stained and eccentric nuclei in
the transplanted tumor cells. Several cancer cells had multiple
nuclei. There was no abnormal change observed in the hearts,
livers, kidneys and spleens of the nude mice. A large number of
brown-like cytoplasmic particles in the immunohistochemical slices
of the blank and negative control groups was noted (Fig. 5).
Discussion
Invasion and metastasis of gastric cancer involve a
complicated and continuous process involving multiple genetic
regulations and factor interaction (15,16). A
number of different factors exert control of this invasion and
metastasis process either by means of mono-regulation at different
levels or mutual regulation (15).
It involves structural and functional gene abnormalities associated
with tumor invasion and metastasis. Dynamic cell movement is
required in the process of transition from in situ cancer to
invasive cancer, where cancer cells need to penetrate the basilar
membrane and matrix gap (14). Euer
et al used GeneChip technology to show that ATX was
correlated with the regulation of high-level expression of more
than 40 genes, which are intimately related to the highly
metastatic characteristic of cells (17). The result of an in vitro test
suggested that non-endogenous ATX-transfected ras-transformed
NIH3T3 cells showed more enhanced invasive and metastatic potential
(18). An in vitro
cytological study confirmed that the ATX system was capable of
mediating cell tumor migration (19). Furthermore an in vivo test
directly identified ATX as an angiogenic factor, since
ATX-transfected cells showed markedly enhanced angiogenic potential
(20). Noh et al firstly
used RNA interference to block ATX expression of breast cancer
MDA435 cells and employed GeneChip technology to perform a spectral
scan comparison of the parental cells and the gene expression of
transfected cells. They found the most upregulated or downregulated
cells had an intimate correlation with cellular metabolism,
cytoskeleton fabric, transcriptional regulation and cell signal
transduction. Therefore, ATX is thought to be a key regulator in
phosphatidic acid metabolism and signal transduction. ATX also
plays a role as a new molecular target for gene therapy against
breast cancer (21).
In previous research, our group found that gastric
cancer tissue samples in patients with distant and regional lymph
node metastases showed extremely high ATX expression, compared with
samples from patients without distant and lymph node metastases.
The result indicated the ATX expression positively correlates with
the metastatic potential of human gastric cancer cells. ATX in
normal gastric tissue may contribute to regulation of cell growth,
the exact mechanism of which, however, still needs to be further
investigated. In the present study, RNA interference technology was
employed to clone ATX via XhoI/BgII into
pSuper-neo-GFP with the H1 promoter, which was then transfected
into the human gastric cancer AGS cell line characterized by low
ATX expression and passaged in a stable process. The result from
the western blot analysis indicated the AGS cells transduced with
target shRNA were able to inhibit endogenous ATX mRNA and protein
expression. After transfection, the migratory and invasive
abilities of AGS cells with low ATX expression were significantly
higher than these abilities in the wild-type cells. This indicates
that AXT signal transduction pathways play an important role in the
process of ATX-mediated cell invasion and migration. Interference
with these pathways was thought effective in prevention of gastric
cancer metastasis, which also allowed us to conclude that ATX could
be a new molecular target for gastric cancer treatment. It has been
speculated by our research group that ATX-mediated gastric cancer
cell invasion and migration depends on RhoGTPase activation. The
RhoGTPase family is a group of important proteins involved in a
number of cell signaling transduction pathways, of which the
members are key regulatory molecules that link membrane surface
receptors to the organization of actin cytoskeleton. Changes in
extracellular signal-induced actin cytoskeleton organization also
cause a series of biological reactions, such as cell morphogenesis,
chemotaxis and axon orientation, (22–24).
Reymond et al comfirmed the Rho, Rac, Cdc42 and Ras promoted
the regulation of signal transduction during regulation of the
cytoskeleton, cell movement, cell proliferation and apoptosis
(25). The results provide the
basis for in vivo testing which is necessary for further
research on the molecular mechanism of ATX in promoting gastric
cancer invasion and metastasis. Our study indicates that the shRNA
plasmid targeting autotoxin effectively downregulates the protein
expression in vivo and inhibits the growth of AGS cells.
Furthermore, by downregulating indirectly the protein levels of
MMP-2 and MMP-9 of the MMP family, shRNA targeting ATX may reduce
the invasive ability of gastric cancer cells.
The decomposition of the main components in basement
membrane type IV and V collagen is carried out by MMP-2 and MMP-9.
The basement membrane acts as a physiological barrier which needs
to be overcome in the process of tumor infiltration and metastasis
(26). Effective degradation of the
extracellular matrix promotes tumor infiltration and metastasis.
What is more, MMP-2 and MMP-9 are hypothesized to play a key role
in the process of endothelial cell migration during angiogenesis.
Additionally, synthesis and release of growth factors VEGF, bFGF
and TGF-β are promoted by MMP-2 and MMP-9 (27,28).
Knockdown of ATX expression results in a decrease in MMP-2 and
MMP-9 expression in gastric cancer, which can inhibit cancer
infiltration and metastasis (29).
Our study suggests that the ATX, Rho and MMP families are
implicated to have pathways in gastric cancer signal transduction.
Low expression of ATX, MMP2 and MMP9 contributes to the inhibition
of cancer cell proliferation and metastasis, further affecting
patient prognosis. The detailed signaling pathways and their
mechanisms require further investigation. Our results provide new
evidence concerning the molecular mechanisms of gastric cancer
metastasis and may aid in developing clinical applications for gene
therapy.
Acknowledgements
This study was funded by the Zhejiang Provincial
Natural Science Foundation of China. We thank Mrs. Kathrin Hammje
and Mr. Yuping Liu for their excellent technical and experimental
assistance. We also deeply thank Ms. Ying Li for her helpful
corrections and suggestions.
References
|
1
|
Stracke ML, Krutzsch HC, Unsworth EJ, et
al: Identification, purification, and partial sequence analysis of
autotaxin, a novel motility-stimulating protein. J Biol Chem.
267:2524–2529. 1992.PubMed/NCBI
|
|
2
|
Umezu-Goto M, Kishi Y, Taira A, et al:
Autotaxin has lysophospholipase D activity leading to tumor cell
growth and motility by lysophosphatidic acid production. J Cell
Biol. 158:227–233. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tokumura A, Majima E, Kariya Y, et al:
Identification of human plasma lysophospholipase D, a
lysophosphatidic acid-producing enzyme, as autotaxin, a
multifunctional phosphodiesterase. J Biol Chem. 277:39436–39442.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kehlen A, Englert N, Seifert A, Klonisch
T, Dralle H, Langner J and Hoang-Vu C: Expression, regulation and
function of autotaxin in thyroid carcinomas. Int J Cancer.
109:833–838. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Saunders LP, Ouellette A, Bandle R, et al:
Identification of small-molecule inhibitors of autotaxin that
inhibit melanoma cell migration and invasion. Mol Cancer Ther.
7:3352–3362. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Debies MT and Welch DR: Genetic basis of
human breast cancer metastasis. J Mammary Gland Biol Neoplasia.
6:441–451. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang SY, Lee J, Park CG, et al: Expression
of autotaxin (NPP-2) is closely linked to invasiveness of breast
cancer cells. Clin Exp Metastasis. 19:603–608. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Panupinthu N, Lee HY and Mills GB:
Lysophosphatidic acid production and action: critical new players
in breast cancer initiation and progression. Br J Cancer.
102:941–946. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang Y, Mou Lj, Liu N, et al: Autotaxin
expression in non-small cell lung cancer. Am J Respir Cell Mol
Biol. 21:216–222. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Stassar MJ, Devitt G, Brosius M, et al:
Identification of human renal cell carcinoma associated genes by
suppression subtractive hybridization. Br J Cancer. 85:1372–1382.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Deissler H, Blass-Kampmann S, Bruyneel E,
et al: Neural cell surface differentiation antigen gp130(RB13-6)
induces fibroblasts and glioma cells to express astroglial proteins
and invasive properties. FASEB J. 13:657–666. 1999.
|
|
12
|
Zhang G, Zhao Z, Xu S, et al: Expression
of autotoxin mRNA in human hepatocellular carcinoma. Chin Med J
(Engl). 112:330–332. 1999.PubMed/NCBI
|
|
13
|
Chen Y, Wei X, Guo C, et al: Runx3
suppresses gastric cancer metastasis through inactivation of MMP9
by upregulation of TIMP-1. Int J Cancer. 129:1586–1598. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gong M, Meng L, Jiang B, et al: p37 from
mycoplasma hyorhinis promotes cancer cell invasiveness and
metastasis through activation of MMP-2 and followed by
phosphorylation of EGFR. Mol Cancer Ther. 7:530–537. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schwartz GK: Invasion and metastases in
gastric cancer: in vitro and in vivo models with clinical
correlations. Semin Oncol. 23:316–324. 1996.PubMed/NCBI
|
|
16
|
Yasui W, Oue N, Aung PP, Matsumura S,
Shutoh M and Nakayama H: Molecular-pathological prognostic factors
of gastric cancer: a review. Gastric Cancer. 8:86–94. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Euer N, Schwirzke M, Evtimova V, et al:
Identification of genes associated with metastasis of mammary
carcinoma in metastatic versus non-metastatic cell lines.
Anticancer Res. 22:733–740. 2002.PubMed/NCBI
|
|
18
|
Nam SW, Clair T, Campo CK, et al:
Autotaxin (ATX), a potent tumor mitogen, augments invasive and
metastatic potential of ras-transformed cells. Oncogene.
19:241–247. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kishi Y, Okudaira S, Tanaka M, et al:
Autotaxin is overexpressed in glioblastoma multiforme and
contributes to cell motility of glioblastoma by converting
lysophosphatidylcholine to lysophosphatidic acid. J Biol Chem.
281:17492–17500. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tanaka M, Okudaira S, Kishi Y, et al:
Autotaxin stabilizes blood vessels and is required for embryonic
vasculature by producing lysophosphatidic acid. J Biol Chem.
281:25822–25830. 2006. View Article : Google Scholar
|
|
21
|
Noh JH, Ryu SY, Eun JW, et al:
Identification of large-scale molecular changes of Autotaxin(ENPP2)
knock-down by small interfering RNA in breast cancer cells. Mol
Cell Biochem. 288:91–106. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li X and Lim B: RhoGTPases and their role
in cancer. Oncol Res. 13:323–331. 2003.
|
|
23
|
Bell CH, Aricescu AR, Jones EY and Siebold
C: A dual binding mode for RhoGTPases in plexin signalling. PLoS
Biol. 9:e10011342011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lazer G and Katzav S: Guanine nucleotide
exchange factors for RhoGTPases: good therapeutic targets for
cancer therapy? Cell Signal. 23:969–979. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Reymond N, Riou P and Ridley AJ: Rho
GTPases and cancer cell transendothelial migration. Methods Mol
Biol. 827:123–142. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wiercinska E, Naber HP, Pardali E, van der
Pluijm G, van Dam H and ten Dijke P: The TGF-β/Smad pathway induces
breast cancer cell invasion through the up-regulation of matrix
metalloproteinase 2 and 9 in a spheroid invasion model system.
Breast Cancer Res Treat. 128:657–666. 2011.
|
|
27
|
Kim ES, Kim MS and Moon A: Transforming
growth factor (TGF)-beta in conjunction with H-ras activation
promotes malignant progression of MCF10A breast epithelial cells.
Cytokine. 29:84–91. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chuang MJ, Sun KH, Tang SJ, et al:
Tumor-derived tumor necrosis factor-alpha promotes progression and
epithelial-mesenchymal transition in renal cell carcinoma cells.
Cancer Sci. 99:905–913. 2008. View Article : Google Scholar
|
|
29
|
Jiang WG, Raz A, Douglas-Jones A and
Mansel RE: Expression of autocrine motility factor (AMF) and its
receptor, AMFR, in human breast cancer. J Histochem Cytochem.
54:231–241. 2006. View Article : Google Scholar : PubMed/NCBI
|