Introduction
Osteosarcoma is the most common type of malignant
bone tumor in children and adolescents. Clinical studies have
reported that this tumor has a poor prognosis, even with current
treatment options, such as surgical intervention and chemotherapy
(1,2). Over the last few decades, neoadjuvant
chemotherapy and surgery have been considered as successful
treatments for osteosarcoma and the limb salvage rate has
significantly increased, considerably raising the survival to
65–75%. However, approximately 30% of osteosarcoma patients develop
metastasis, particularly pulmonary metastasis, which is the leading
cause of mortality (3). Thus, it is
crucial to identify metastasis-associated molecules and to
elucidate the mechanisms involved in pulmonary metastasis from
osteosarcoma. A number of studies have shown that specific
microRNAs (miRNAs) are aberrantly expressed in malignant
hepatocellular carcinoma (HCC) cells or tissues compared to
non-malignant hepatocytes or tissues (4,5).
miRNAs are metastasis-associated molecules during the progression
and metastasis of osteosarcoma.
miRNAs are a class of small non-coding regulatory
RNA molecules (21–23 nucleotides) encoded in the genome, with
profound impacts on various cellular processes (6,7). These
small molecules mainly bind imperfectly by base pairing to the 3′
untranslated region (3′UTR) of target mRNAs and repress protein
expression through either degradation or silencing (7–9).
Evidence has shown that miRNA mutations or misexpression correlate
with various human cancers, where they were thought to function as
tumor suppressors or oncogenes by regulating the expression of
other tumor suppressors or oncogenes (8–10).
However, the mechanisms by which miRNAs mediate tumor metastasis
have only been recently investigated and have not been fully
elucidated.
miR-34 family members, consisting of miR-34a, b and
c, are direct transcription targets of the tumor suppressor protein
p53 (11). miR-34a has been
reported to induce cell apoptosis, cell cycle arrest or cell
senescence depending on the cell type analyzed (11–13).
Some targets of miR-34a have been recently identified including
cyclin-dependent kinase (CDK)4/6, cyclin D1 (CCND1), cyclin E2
(CCNE2), E2F transcription factor 3 (E2F3), Bcl-2, MYCN, c-Met,
sirtuin 1 (SIRT1) (14–17). However, its role in mediating
osteosarcoma metastasis to the lungs has only been recently
investigated and remains largely ambiguous.
The current study was carried out to investigate the
potential role of miR-34a in the invasion and metastasis of
osteosarcoma cells. We investigated the expression level and
functional pattern of miR-34a in osteosarcoma cells. This was
performed by the quantification of miR-34a in the highly metastatic
subline, F5M2, and in the F4 subline with low metastatic potential
of the paired human osteosarcoma cell line, SOSP-9607. The
transfection outcomes were subsequently assessed according to cell
viability patterns, cell migration and alterations in gene
expression by real-time PCR, as well as the protein level by
immunocytochemistry (ICC) and western blot analysis. Functional
analysis and sequence analysis were then carried out by the
transfection of miR-34a mimics or inhibitors into the highly
metastatic subline, F5M2, with a low endogenous miR-34a level.
In the present study, we demonstrated that the
miR-34a expression level in the F4 human osteosarcoma cells with a
low metastatic potential was higher than that in the highly
metastatic F5M2 cells. Subsequently, we investigated the effects of
the ectopic expression of miR-34a on F5M2 cells, and showed that
miR-34a inhibited the invasion and metastasis of osteosarcoma cells
in vitro. Sequence analysis suggested an interaction between
the 3′UTR of CD44 mRNA and miR-34a. Therefore, we hypothesized that
miR-34a can repress the expression of CD44 and analyzed its
functions in human osteosarcoma cells.
Materials and methods
Cell culture
A pair of human osteosarcoma cell lines with
different pulmonary metastatic potentials, the highly metastatic
subline, F5M2, and the F4 subline with low metastatic potential,
originating from the human osteosarcoma cell line, SOSP-9607, were
established by Dr X. Chen in our laboratory (18). The F5M2 and F4 cell sublines were
cultured in complete RPMI-1640 medium (HyClone, Logan, UT, USA)
supplemented with 10% fetal bovine serum (Sijiqing Co. Hangzhou,
China) in 5% CO2 at 37°C.
Transfection
In this study, transfection was performed with
Lipofectamine 2000 Reagent (Invitrogen, Carlsbad, CA, USA)
according to the manufacturer’s instructions. miR-34a mimics, mimic
negative controls, miR-34a inhibitor and negative controls were
purchased from RiboBio (Guangzhou, China). A low concentration of
20 nM or a high concentration of 50 nM of the mimics were used for
each transfection in the migration, invasion, apoptosis and
proliferation assays, compared with the F5M2 cells transfected with
the mimic negative controls, miR-34a inhibitor and negative
controls. The efficiency of the miR-34a transfection was measured
by quantitative real-time PCR.
Quantitative real-time PCR analysis
miR-34a, CD44 mRNA expression was measured by
real-time PCR. Total RNA was extracted using TRIzol reagent
(Invitrogen) according to the manufacturer’s instructions. For
miR-34a quantitative real-time PCR, total RNA was reverse
transcribed with a miR-sepicfic primer (RiboBio) and then
quantitative real-time PCR was performed with a miR-sepicfic primer
using the ABI PRISM 7500 real-time PCR system (Applied Biosystems,
Bedford, MA, USA). U6 was used as the normalization control.
Quantitative real-time PCR for CD44 mRNA was performed with CD44
mRNA primers (forward, 5′-CATCTACCCCA GCAACCCTA-3′; and reverse,
5′-ACTGTCTTCGTCTGGG ATGG-3′) and the relative expression level
normalized to GAPDH was calculated using the comparative Ct
method.
Transwell assay
For migration assay, we used the 6.5-mm transwell
insert (pore size, 8 μm; 24-well insert; Corning) to examine the
effect of miR-34a on the migration and invasion of the F5M2 and F4
cells. Cells were serum-free-starved overnight, and then harvested
and resuspended in migration medium (RPMI-1640 medium free with
fetal bovine serum). Cell suspension (5,000 cells) in 100 μl
migration medium was then added to the inside of the insert.
RPMI-1640 medium (600 μl) containing 20% fetal bovine serum was
added to the lower chambers outside of the insert. After the cells
were incubated for 48 h, the non-invading cells that remained on
the upper surface of the insert were removed with a cotton swab.
The invading cells on the lower surface of the insert were fixed
with 95% ethanol for 30 min and then stained with 0.1% crystal
violet for 5 min. The number of cells on the lower surface was
counted under a microscope in 6 randomized visual fields of each
insert at a magnification of ×100.
Invasion assay was carried out in a similar manner
as the migration assay but the inserts were pre-coated with 40 μl
BD Matrigel (1:3 dilution; BD Biosciences, San Jose, CA USA)
according to the manufacturer’s instructions.
Wound healing assay
Adhered cell monolayers on 6-well plates were
scratched with a 20-μl pipette tip (Eppendorf) and grown in
RPMI-1640 medium with 10% fetal bovine serum (Sijiqing Biological
Engineering Materials Co., Ltd., Hangzhou, China) with 5%
CO2 at 37°C. Wound healing capacity was monitored under
a microscope after 0, 12, 24 and 36 h.
Apoptosis and proliferation assay
For the apoptosis assay, the cells were stained with
FITC-conjugated anti-Annexin V antibody. The Annexin V-FITC
apoptosis detection kit (BD Pharmingen, San Diego, CA, USA) was
employed to analyze cell apoptosis with flow cytometry (BD
Biosciences FACSAria cell sorter).
For the proliferation assay, the proliferative
capacity of the cells was evaluated with an MTT assay, which was
performed following standard procedure in 96-well plates. Cells
were seeded at a density of 2,000 cells/well containing 100 μl of
culture medium and cultured overnight. At 24-h intervals, 20 μl of
5 mg/ml MTT (dimethyl thiazolyl diphenyl tetrazolium; Sigma) were
added to each well and then the cells were incubated for 4 h at
37°C. The medium was then removed, and 100 μl of dimethyl sulfoxide
(DMSO) were added to each well to dissolve the formazan. The
optical density (OD) was evaluated by measuring the absorbance.
Western blot analysis and ICC
The CD44 expression level was analyzed by western
blot analysis using CD44 mouse monoclonal antibody (Abcam) and
anti-β-actin mouse monoclonal antibody (Epitomics, Inc.,
Burlingame, CA, USA). ICC was performed with CD44 mouse monoclonal
antibody (Abcam) and the Envision™ Detection kit (Gene-Tech Co.,
Ltd., Shanghai, China) according to a standard method.
Luciferase reporter assay
F5M2 cells were transfected in 6-well plates with 2
μg of CD44 3′UTR (containing the binding site of miR-34a)
luciferase reporter plasmid per well, using Lipofectamine 2000
(Invitrogen). Cells in each well were also co-transfected with 100
pmol of each miR-34 mimics, negative control mimics, or miR-34a
inhibitors as indicated. Cells were collected 48 h after
transfection, and luciferase assays were performed using a
dual-luciferase reporter assay system (Promega). Luciferase
activity was normalized relative to the control activity as
previously described (19).
Statistical analysis
All statistical analyses were performed using SPSS
17.0 software. The experiments were repeated 3 times independently
and the results are presented as the means ± SD. The differences
between groups were analyzed using Student’s t-tests; P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-34a expression levels are lower in
F5M2 cells than in F4 cells
In this study, the F5M2 and F4 cell sublines were
chosen as the objectives since they originate from the same
maternal cell line, SOSP-9607 (human osteosarcoma cell line), but
display dramatically significant differences in metastatic and
invasive potential (18).
Of all potential miRNAs, we focused on miR-34a since
it is one of the most obviously altered miRNAs and is
underexpressed in mouse hepatoma cells with a high lymphatic
metastatic potential and in highly metastatic human prostate cancer
cells (20,21). However, the functional mechanism of
miR-34a in these cancers remains unclear.
In order to investigate the differential expression
of miR-34a in the osteosarcoma cell sublines with different
metastatic potential, we compared the miR-34a expression between
the F5M2 and F4 cell sublines by quantitative real-time PCR.
Consistent with the results obtained for the mouse hepatoma cells
and for the highly metastatic human prostate cancer cells,
real-time PCR revealed that the expression level of miR-34a in the
highly metastatic F5M2 cells was lower than that in the F4 cells
with low metastatic potential. This difference in expression level
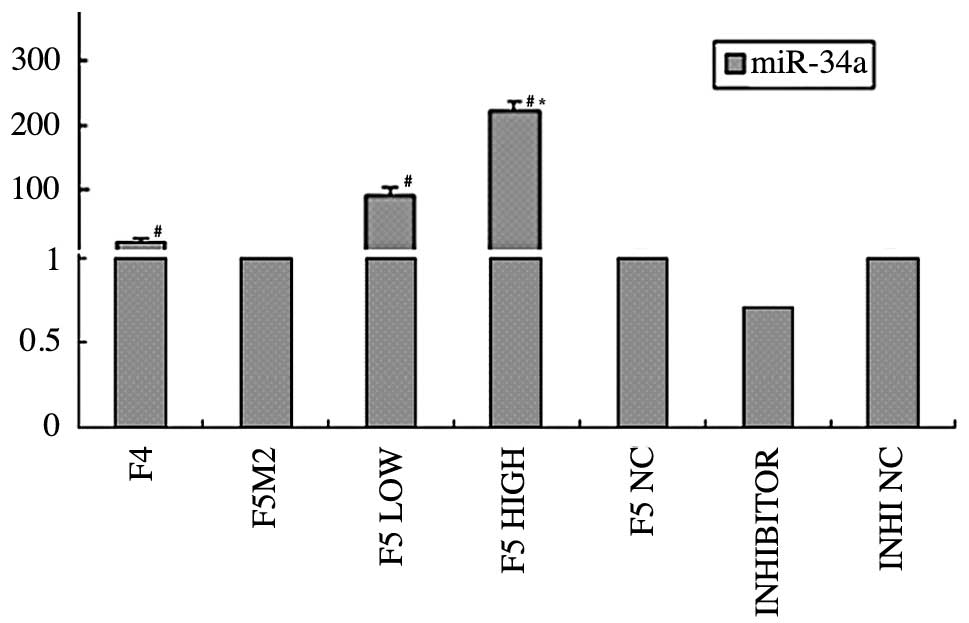
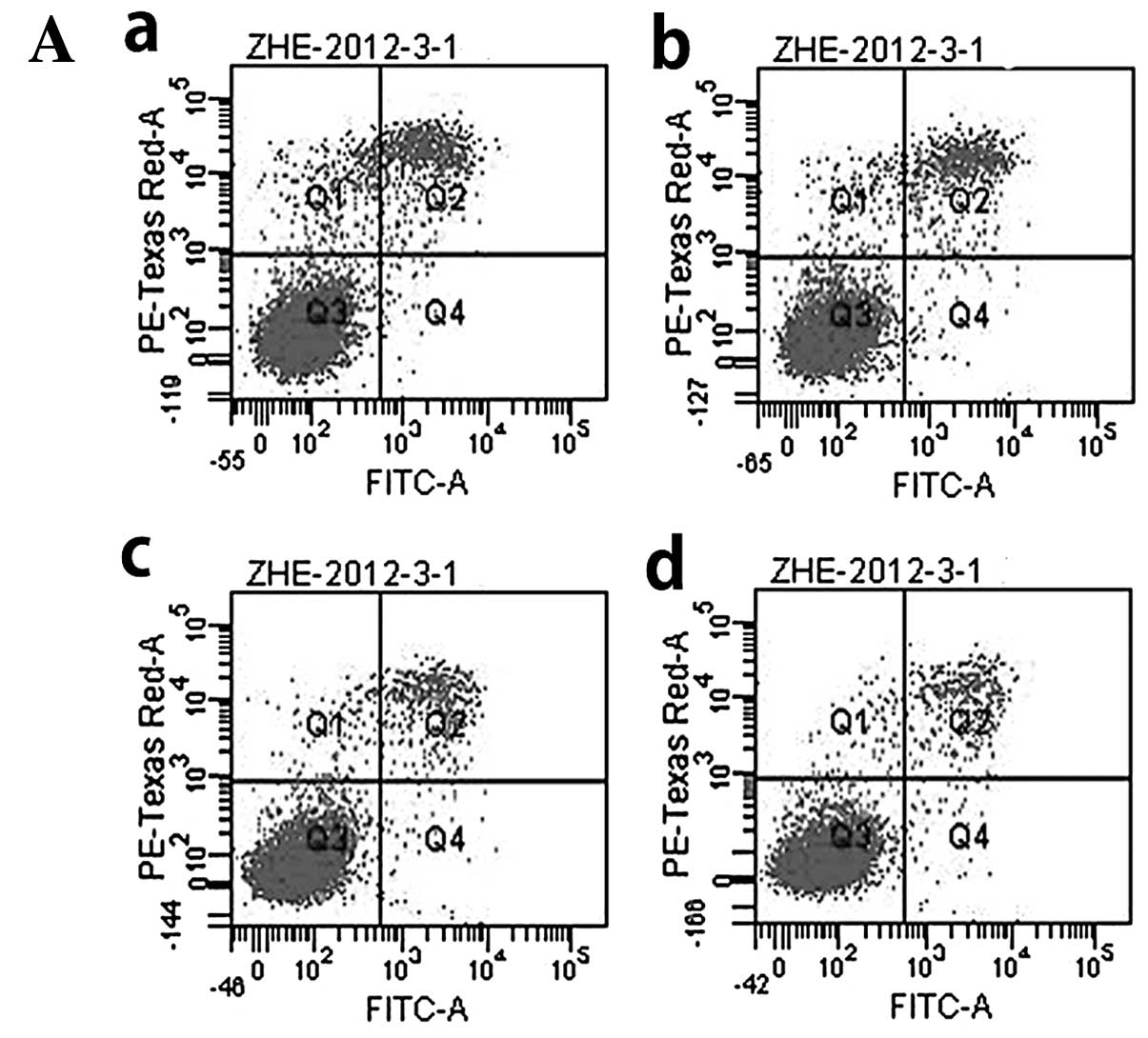
was statistically significant (P<0.05; Fig 1). These results suggest that the
downregulation of miR-34a is involved in the metastatic and
invasive potential of human osteosarcoma cells.
F5M2 cells significantly overexpress
miR-34a after transfection
F5M2 cells were transfected with the miR-34a mimics
at 2 different concentrations, 20 or 50 nM. The control groups
included F5M2 cells that were untreated or transfected with the
mimic negative controls, miR-34a inhibitor and negative controls.
To verify the efficiency of the transfection, the expression of
miR-34a was measured by quantitative real-time PCR of total RNA 48
h after transfection.
The results from real-time PCR showed that the
expression of miR-34a significantly increased in the F5M2 cells
after transfection with the miR-34a mimics, compared with the
control groups (P<0.05; Fig. 1).
The results also showed that miR-34a expression in the F5M2 cells
transfected with a high concentration of mimics (50 nM) was
significantly higher than that in the cells transfected with a low
concentration of mimics (20 nM). Thus, the transfection was
successful. The higher the concentration of the miR-34a mimics, the
higher the expression of the miR-34a in the F5M2 cells after
transfection.
CD44 mRNA is a target of miR-34a
The prediction of miR-34a putative target genes was
performed using bioinformatics algorithms based on sequence
similarity between miRNAs and mRNAs. We used TargetScan (http://www.targetscan.org/) (22) and PicTa’r (http://pictar.mdc-berlin.de/) (23,24) to
predict the targets of miR-34a. Remarkably, the programs predicted
that CD44 was one of the targets of miR-34a. Liu et al
established miR-34a as a critical negative regulator of
CD44+ PCa cells and CD44 itself as an important target
of miR-34a (21). Therefore, we
hypothesized that miR-34a may alter F5M2 cell migration and
invasion by regulating the expression of CD44. To verify this
hypothesis, we examined the expression level of CD44 in F5M2 cells
after transfection with miR-34a mimics by ICC and western blot
analysis.
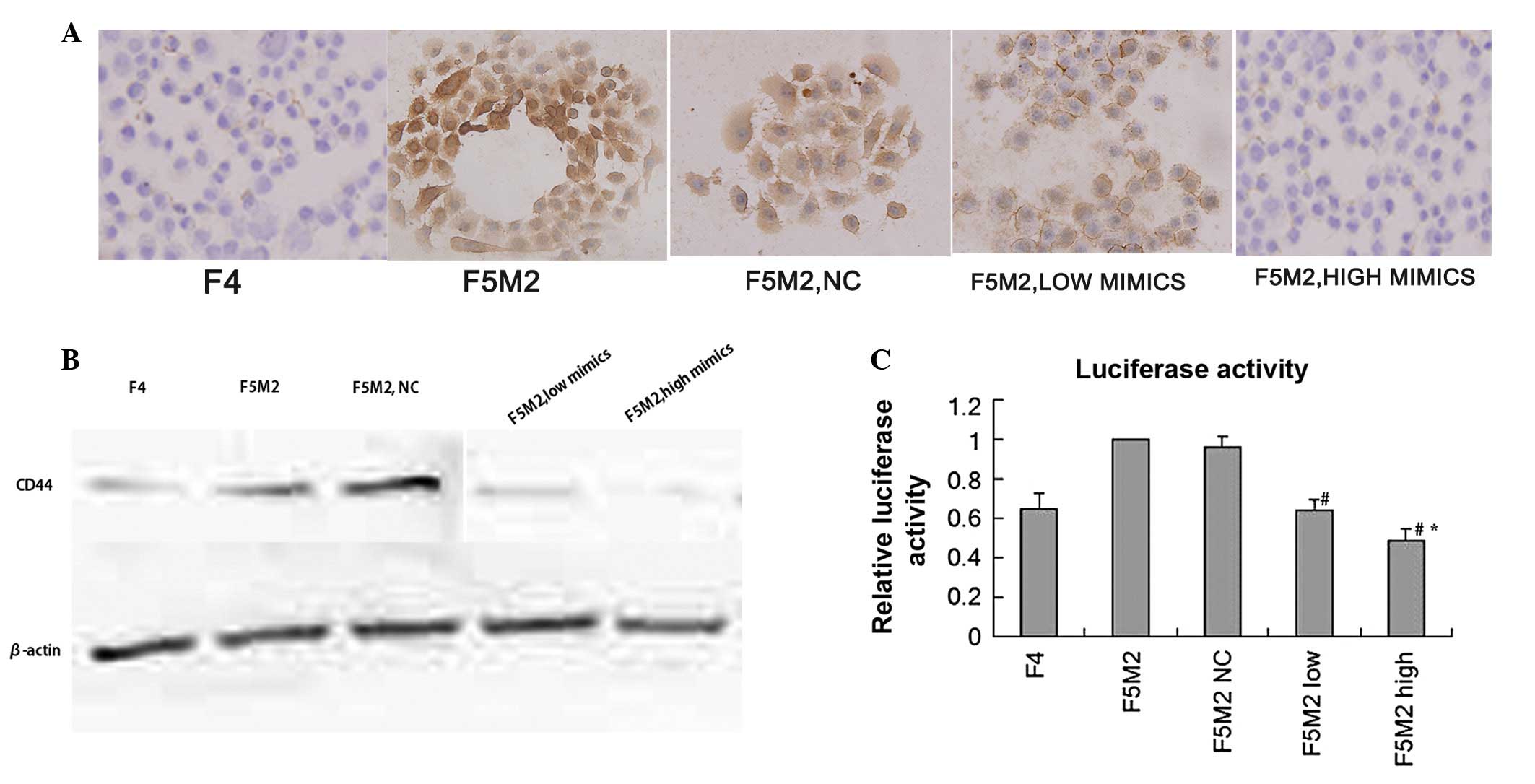
ICC analysis showed that the staining intensity of
CD44 in the F5M2 cells was stronger than that in the F4 cells; this
was significantly reduced in the F5M2 cells after transfection with
miR-34a mimics (Fig. 2A). The
results from western blot analysis were similar to those from ICC
analysis, in that the expression level of CD44 in the F5M2 cells
was significantly higher than that in the F4 cells and that the
expression of CD44 in the F5M2 cells markedly decreased after
transfection with miR-34a mimics, compared with the F5M2 cells
untreated or treated with the negative controls (Fig. 2B). Both in western blot analysis and
ICC, the expression of CD44 had the same tendency; the F5M2 cells
treated with 50 nM miR-34a mimics expressed lower levels of CD44
than the cells treated with 20 nM miR-34a mimics (P<0.05). There
was a negative correlation between the CD44 levels and the
exogenous miR-34a levels. The overexpression of miR-34a may thus
inhibit the expression of CD44. Therefore, CD44 mRNA is a possible
target of miR-34a.
To further prove that the changes in the protein
levels were a direct effect of miR-34a, we fused the 3′UTRs of CD44
to a luciferase reporter construct. The results demonstrated that
the luciferase activity of the miR-CD44 UTR construct was
significantly inhibited after the introduction of miR-34a mimics
(Fig. 2C; P<0.05). Consistent
with the results reported previously by others (21,25),
the transfection of miR-34a mimics effectively modulated the
luciferase reporter gene expression compared with the control
groups. The results of luciferase reporter assay demonstrated that
CD44 was a direct target of miR-34a in the F5M2 cells.
miR-34a significantly decreases the
migration and invasive ability of F5M2 cells
Consistent with the conclusions of the metastatic
potential of F5M2 cells presented in the study by Chen et
al(18), in our study, F5M2
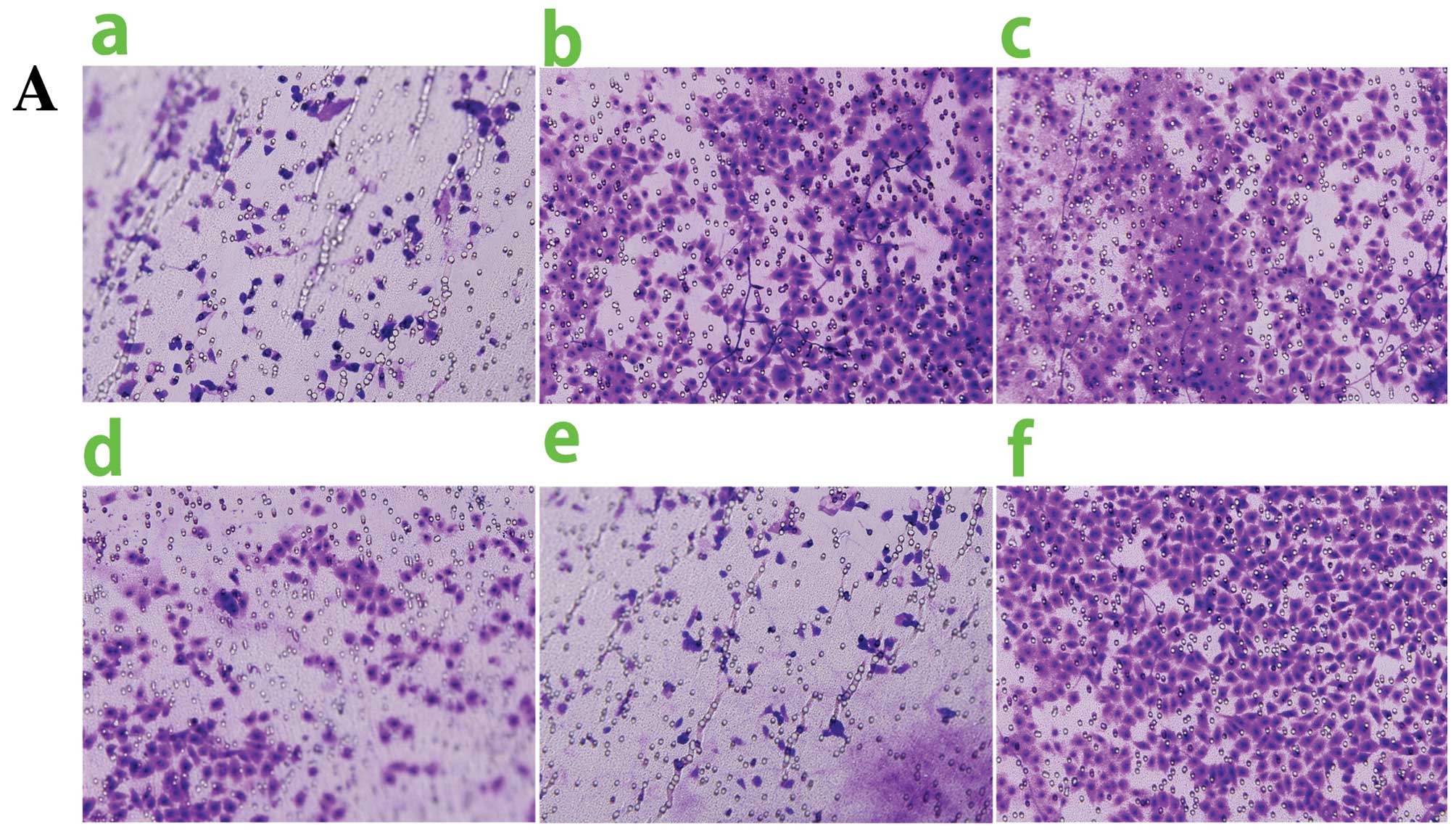
cells displayed a significantly higher migration and invasive
capability than F4 cells in vitro, as shown by transwell
assay (Fig. 3A and B).
To explore the potential role of miR-34a in
osteosarcoma cell metastasis, we examined the cell motility and
invasive capability after the ectopic expression of miR-34a in F5M2
cells, which had been verified to underexpress endogenous miR-34a
previously.
Transwell assay without Matrigel was employed to
examine the migration capability. Cells that penetrated the
membrane and reached the under side of the transwell were recorded
after incubation for 48 h. The results showed that the ectopic
overexpression of miR-34a significantly repressed the chemotaxis of
F5M2 cells, compared with the F5M2 cells that were untransfected or
transfected with the mimic controls (Fig. 3A and C; P<0.05).
Transwell inserts with a layer of Matrigel on top of
the inserts were employed to examine the invasive capability. Cells
that penetrated both Matrigel and membrane were counted after
incubation for 48 h. The results showed that ectopic miR-34a
expression in the F5M2 cells significantly inhibited their invasive
ability (Fig. 3B and D; P<0.05),
which was consistent with the results obtained for migration.
Additionally, the cell motility of F5M2 cells
transfected with a high concentration of mimics was significantly
weaker than that of F5M2 cells transfected with a low concentration
of mimics, as shown by both the migration and invasion assay. Taken
together, these results demonstrate that miR-34a inhibits the F5M2
cell migration and invasion potential in vitro.
miR-34a significantly decreases wound
healing capacity of F5M2 cells
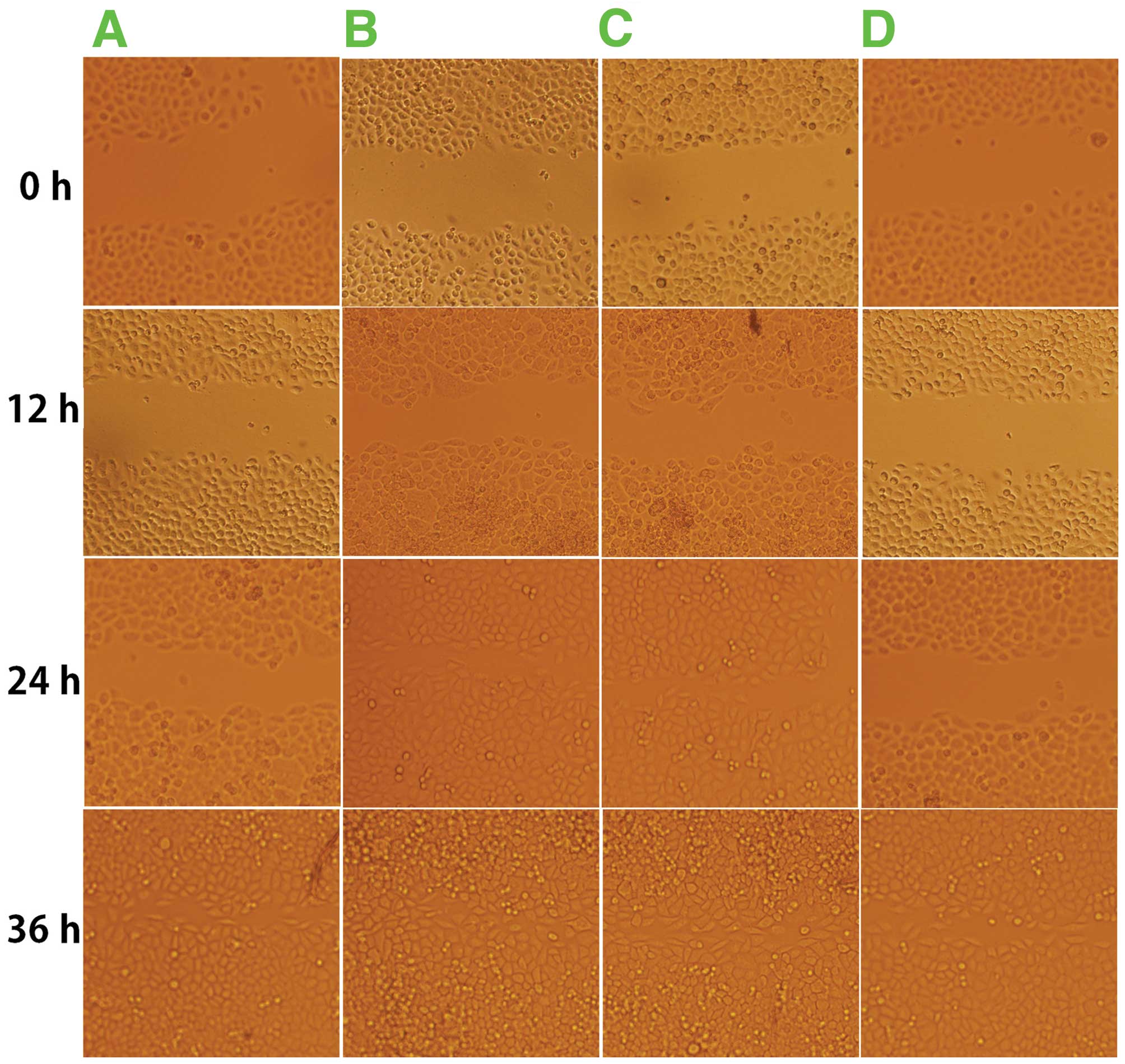
To compare the polarized migration of F5M2 and F4
cells, we employed the scratch wound cell model. The results showed
that in the F5M2 cells, the scratch wounds healed faster than those
in the F4 cells (Fig. 4;
P<0.05). This model also showed that ectogenic miR-34a
significantly decreased the wound healing capacity of F5M2 cells,
compared with those cells untreated or transfected with the
negative controls (Fig. 4 and
Table I). These results
demonstrated that the inhibitory effect of miR-34a was dependent on
the concentration of miR-34a mimics (data not shown). The higher
the concentration of the miR-34a mimics, the slower the rate at
which the F5M2 cells closed the scratch wounds after
transfection.
 | Table IWound closure rate of F4 or F5M2
transfected with miR-34a mimics, untreated or treated with the
negative controls. |
Table I
Wound closure rate of F4 or F5M2
transfected with miR-34a mimics, untreated or treated with the
negative controls.
| Time (h) |
|---|
|
|
|---|
| Cell
transfection | 0 | 12 | 24 | 36 |
|---|
| F4 | 0 | 0.42±0.08 | 0.62±0.02a | 0.90±0.13a |
| F5M2 | 0 | 0.41±0.12 | 0.83±0.16 | 0.99±0.08 |
| F5M2, NC | 0 | 0.45±0.12 | 0.85±0.06 | 0.97±0.03 |
| F5M2, miR-34a | 0 | 0.43±0.07 | 0.61±0.08a | 0.78±0.11a |
miR-34a influences apoptosis and
proliferation of F5M2 cells
The results of flow cytometry showed that there was
no statistically significant difference either between F5M2 and F4
cells, or between F5M2 cells untreated or transfected with miR-34a
mimics and negative controls (Fig. 5A
and B). Therefore, miR-34a had little effect on cell
apoptosis.
In order to investigate the effects of miR-34a on
cell proliferation, MTT assay was performed. The proliferation
curve was then depicted. The results demonstrated that the F5M2
cells transfected with the miR-34a mimics exhibited a similar
tendency in proliferation capacity between the groups. This
demonstrates that the exogenous miR-34a expression had little
effect on the proliferation of osteosarcoma cells (Fig. 5C).
Discussion
It has previously been reported that several miRNAs
are metastatic suppressors, including miR-126, miR-335, let-7c,
miR-100, miR-218, miR-375, miR-125, miR-198 and miR-142. The
reduced expression of miR-126 and miR-335 has been found in breast
cancer characterized by poor metastatic-free survival (26), while the expression of miR-let7c,
miR-100 and miR-218 has been shown to be significantly decreased in
metastatic prostate cancer compared with localized prostate cancer
(27). The ectopic expression of
miR-375 induces changes in cell morphology, and inhibits melanoma
cell invasion and wound healing, strongly suggesting the functional
role of miR-375 in cytoskeletal architecture and migration
(28). Moreover, the enforced
ectopic expression of miR-125 has been shown to impair cell
migration and invasion in a breast cancer cell line, and the
reduction of miR-125 expression enhances the migration ability of
the cells (29). miR-198 has been
shown to be downregulated in HCC and the forced ectopic expression
of miR-198 represses HCC cell migration and invasion in a
c-MET-dependent manner (30).
Another study reported that miR-142 expression was downregulated in
HCC cells and that the overexpression of miR-142 inhibited cell
migration and invasion, while the blocking of miR-142 increased
cell migration and invasion. It has also been demonstrated that
miR-142 inhibits HCC cell migration and invasion by targeting RAC1
(31).
The functional analysis of miR-34a has recently been
reported in mouse HCC cell lines and in human prostate cancer
progenitor cells. Liu et al(21) presented miR-34a as a potent
metastasis inhibitor of prostate cancer and reported that the
upregulation of miR-34a inhibited the migration of cancer cells and
extended animal survival. They demonstrated that miR-34a induced
the dysregulation of genes related to migration and invasion by
directly repressing CD44. Guo et al demonstrated that the
ectopic upregulation of miR-34a inhibited the lymphatic metastatic
potential of Hca-F cells (highly metastatic mouse hepatoma cells);
however, this was shown to be mediated via the downregulation of
cyclin D1 and CDK6 in Hepa1-6 cells (20). Some targets of miR-34a have been
recently identified, including CDK4/6, CCND1, CCNE2, E2F3, Bcl-2,
MYCN, c-Met, SIRT1, Notch1 and Jagged1 (14,32–35).
However, the role of miR-34a in tumor metastasis remains to be
fully elucidated. Taken together, these data indicate that miR-34a
is likely to have a number of targets through which it regulates
the biological functions of cancer cells, including metastasis.
In the results of previous biochemical studies, it
has been reported that the miRNA repression of mRNA expression is
dependent on specific cellular conditions (36). Studies have demonstrated that the
expression profiling of miR-34a is tissue-specific, and that it may
have divergent functions depending on the tumor tissue or cell
type. To identify the potential role of miR-34a in osteosarcoma
metastasis, we carried out this study to compare the miR-34a
expression level in F5M2 cells with that in F4 cells, which are
sublines of the osteosarcoma cell line, SOSP-9607, with high and
low metastatic potential, respectively (18). We used a variety of methods to
analyze the inhibitory role of miR-34a in the migration and
invasion of F5M2 cells. Real-time PCR demonstrated that miR-34a
expression in F5M2 cells was lower than that in F4 cells. After
transfection with miR-34a mimics, the results indicated that the
overexpression of miR-34a mainly inhibited the migration and
invasion of the transfected F5M2 cells, compared with the F5M2
cells untreated or transfected with the negative controls.
Therefore, it is reasonable to conclude that miR-34a may act as a
tumor suppressor in osteosarcoma.
The identification of cancer-specific miRNAs and
their targets is pivotal for understanding their role in
tumorigenesis and metastasis, and may be important for the
discovery of novel therapeutic targets. To identify the suppressive
mechanism of miR-34a in osteosarcoma cell metastasis, we employed
miRNA target prediction programs (TargetScan and PicTa’r) to match
the direct targets of miR-34a (23). Markedly, the programs predicted that
CD44 was one of the targets of miR-34a. CD44 which contained the
corresponding binding site of miR-34a in its 3′UTR, was regulated
by miR-34a. A previous study demonstrated that miR-34a inhibited
prostate cancer stem cells and metastasis by directly repressing
CD44 (21). The results of the
present study showed that the CD44 level inversely correlated with
the miR-34a level. The results of luciferase reporter assay
demonstrated that CD44 was a direct target of miR-34a in F5M2
cells. Of note, this study also demonstrated that CD44 levels
positively correlated with osteosarcoma cell metastasis and
invasion. Therefore, it is reasonable to conclude that alterations
in miR-34a expression may regulate cell migration and invasion by
targeting CD44.
CD44 (also known as homing cellular adhesion
molecule, Hermes antigen and PGP-1) is encoded by a single, highly
conserved gene with a length of 50–60 kb and is composed of 20
exons. Hhuman CD44 is an 85–90 kDa transmembrane glycoprotein
receptor belonging to the family of cell adhesion proteins and
involved in cell-extracellular matrix interactions as well as in
cell-cell interactions. CD44 participates in many cellular
processes including the regulation of cell survival, migration and
adhesion through the binding of its major ligand, hyaluronic acid
(37–40). Recently, it has been reported that
the aberrant overexpression of CD44 correlates with the metastatic
potential of several malignant tumors, such as prostate cancer,
breast tumors and chondrosarcoma (21,41,42).
CD44 is a molecular marker in a variety of tumors.
Chen et al reported that CD133+CD44+
stem-like cancer cells were highly enriched in HCT116 colon cancer
cells and that metastatic colon cancer cells resided exclusively in
a subpopulation of cells with a high CD44 expression (43). Zhang et al strongly suggested
that the CD44+ subpopulation of human gastric cancer
cell lines, AGS, is gastric cancer stem cells (44). It has been reported that CD44 is
involved in cancer cell adhesion, motility and invasion through the
interaction with hyaluronic acid and is necessary for the
metastasis and recurrence of breast cancer cells (45,46).
The same study also concluded that antibodies against different
epitopes on CD44 mediated distinct functional effects on breast
cancer cells. Heyse et al suggested that the overexpression
of CD44s correlated significantly with the metastatic potential of
human chondrosarcoma cells and there was a significant association
beetween high CD44 expression and poor survival in patients with
chondrosarcoma (42). They also
reported that the overexpression of CD44 was necessary for the
metastasis of osteosarcoma and that the high expression of CD44 was
also associated with the low survival rate of patients with
osteosarcoma (47).
The overexpression of CD44 may be considered a
prognostic parameter of osteosarcoma and a metastatic parameter of
chondrosarcoma (42,47), but whether miRNAs regulate CD44 and
osteosarcoma metastasis remains unclear. In this study, through
expression analysis, we showed that miR-34a was underexpressed in
highly metastatic F5M2 cells originating from the osteosarcoma cell
line, SOSP-9607.
To verify the inhibitory effect of miR-34a on CD44
expression, we conducted a trial to transfect miR-34a into F5M2
cells, which expressed higher levels of CD44 protein and had a
higher pulmonary metastatic potential than the paired F4 cells
(18). After transfection with
miR-34a mimics, CD44 expression in F5M2 cells was almost abolished,
as shown by western blot analysis and ICC. On the contrary, F5M2
cells transfected with the negative controls or miR-34a inhibitors
showed only a slight change in CD44 expression. Our study also
showed that the enforced ectopic expression of miR-34a in F5M2
cells significantly inhibited cell motility and invasion. We also
demonstrated that miR-34a had little effect on cell apoptosis and
proliferation. Taken together, these data suggest that miR-34a
expression is inversely related to the metastasis of osteosarcoma
cells. CD44 expression positively correlated with the metastasis
and invasion of osteosarcoma cells. Therefore, it can be concluded
that the enforced ectopic expression of miR-34a in F5M2 cells might
act as a significant inhibitory factor in the progression of
osteosarcoma cells by downregulating the expression of CD44. Of
note, CD44 was identified and validated as a direct and functional
target of miR-34a in inhibiting osteosarcoma cell regeneration and
metastasis.
Many human cancers contain cancer stem cells or
tumor progenitor cells, which have the ability of self-renewal,
partial recreation of the cellular heterogeneity of the parental
cells, and possess an enhanced tumor-initiating capacity, and are
thus more resistant than most general cancer cells to conventional
anticancer treatment. Due to these properties, cancer stem cells or
tumor progenitor cells have been linked to tumor progression and
metastasis (48). Chen et al
demonstrated with clonal analysis that a lineage of self-renewing
tumor-initiating cell types existed in glioblastoma (49). Either individually or in combination
with other markers, the adhesion molecule, CD44, has identified
cancer stem cells in many tumors, including cancers of the breast,
pancreas, head and neck, colon, small intestine, live, stomach,
bladder and ovaries.
The exact mechanisms by which miR-34a inhibits
osteosarcoma cell metastasis by repressing CD44 still remain
largely ambiguous; however, the enforced ectopic expression of
miR-34a inhibited the migration and invasion of the F5M2 cell
subline. Considering the widespread expression of CD44 in cancer
stem cells and the functional involvement of CD44 in mediating
cancer stem cell migration and the metastasis of many cancers
(50,51), the newly identified miR-34a
suppression of CD44 reveals a key role for miRNA-based gene
regulation. The emerging tumor-suppressive role of miR-34a in
prostate and lung tumors (21,52),
establishes a strong rationale for developing miR-34a as a novel
therapy targeting osteosarcoma cells.
During the course of our study, other reports on
miRNAs were published, which identified miR-219, miR-20a, miR-223,
miR-222, miR-195 tumor-associated miRNAs. miR-219 has been shown to
exert tumor-suppressive effects in hepatic carcinogenesis through
the negative regulation of GPC3 expression in vitro(53). miR-20a promotes the migration and
invasion of cervical cancer cells through the direct upregulation
of TNKS2, which induces the colony formation, migration and
invasion of cervical cancer cells (54). The overexpression of miR-223
significantly inhibits cell proliferation by regulating FOXO1
expression (55). These studies
further suggest that miRNAs are involved in tumor metastasis and
progression.
The findings of this study illustrate that, by
downregulating CD44 expression, miR-34a plays an inhibitory role in
the migration and invasion of osteosarcoma cells. To our knowledge,
this is the first study to demonstrate that miR-34a regulates the
metastasis and progression of osteosarcoma cells, by targeting the
expression of CD44 in F5M2 cells in vitro. The data
presented in our study may provide an novel avenue for further
analysis with an aim to develop a novel potential therapeutic
target for the treatment of highly metastatic osteosarcoma in
vivo. Further study would be required to fully elucidate the
exact roles of miR-34a and CD44 in osteosarcoma in vitro and
in vivo.
Acknowledgements
We would like to thank Yanhua Wen, Chen Xiang,
Yunyan Liu, Dianzhong Zhang and Hong Zhao for their excellent
technical assistance and helpful discussions.
References
|
1
|
Bentzen SM, Poulsen HS, Kaae S, Jensen OM,
Johansen H, Mouridsen HT, Daugaard S and Arnoldi C: Prognostic
factors in osteosarcomas, a regression analysis. Cancer.
62:194–202. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Davis AM, Bell RS and Goodwin PJ:
Prognostic factors in osteosarcoma: a critical review. J Clin
Oncol. 12:423–431. 1994.PubMed/NCBI
|
|
3
|
Mankin HJ, Hornicek FJ, Rosenberg AE,
Harmon DC and Gebhardt MC: Survival data for 648 patients with
osteosarcoma treated at one institution. Clin Orthop Relat Res.
429:286–291. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Medina PP and Slack FJ: microRNAs and
cancer: an overview. Cell Cycle. 7:2485–2492. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pillai RS, Bhattacharyya SN and
Filipowiczm W: Repression of protein synthesis by miRNAs: how many
mechanisms? Trends Cell Biol. 17:118–126. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Caldas C and Brenton JD: Sizing up miRNAs
as cancer genes. Nat Med. 11:712–714. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He L, He X, Lim LP, de Stanchina E, Xuan
Z, et al: A microRNA component of the p53 tumour suppressor
network. Nature. 447:1130–1134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Raver-Shapira N, Marciano E, Meiri E,
Spector Y, Rosenfeld N, et al: Transcriptional activation of
miR-34a contributes to p53-mediated apoptosis. Mol Cell.
26:731–743. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Welch C, Chen Y and Stallings RL:
MicroRNA-34a functions as a potential tumor suppressor by inducing
apoptosis in neuroblastoma cells. Oncogene. 26:5017–5022. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun F, Fu H, Liu Q, Tie Y, Zhu J, et al:
Downregulation of CCND1 and CDK6 by miR-34a induces cell cycle
arrest. FEBS Lett. 582:1564–1568. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei JS, Song YK, Durinck S, Chen QR, Cheuk
AT, et al: The MYCN oncogene is a direct target of miR-34a.
Oncogene. 27:5204–5213. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yamakuchi M, Ferlito M and Lowenstein CJ:
miR-34a repression of SIRT1 regulates apoptosis. Proc Natl Acad Sci
USA. 105:13421–13426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen X, Yang TT, Wang W, Sun HH, Ma BA, Li
CX, Ma Q, Yu Z and Fan QY: Establishment and characterization of
human osteosarcoma cell lines with different pulmonary metastatic
potentials. Cytotechnology. 61:37–44. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He C, Xiong J, Xu X, Lu W, Liu L, Xiao D
and Wang D: Functional elucidation of MiR-34 in osteosarcoma cells
and primary tumor samples. Biochem Biophys Res Commun. 388:35–40.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo Y, Li S, Qu J, Wang S, Dang Y, et al:
MiR-34a inhibits lymphatic metastasis potential of mouse hepatoma
cells. Mol Cell Biochem. 354:275–282. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu C, Kelnar K, Liu B, Chen X,
Calhoun-Davis T, et al: The microRNA miR-34a inhibits prostate
cancer stem cells and metastasis by directly repressing CD44. Nat
Med. 17:211–215. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lewis BP, Shih IH, Jones-Rhoades MW,
Bartel DP and Burge CB: Prediction of mammalian microRNA targets.
Cell. 115:787–798. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Krek A, Grün D, Poy MN, Wolf R, Rosenberg
L, et al: Combinatorial microRNA target predictions. Nat Genet.
37:495–500. 2005. View
Article : Google Scholar
|
|
24
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human MicroRNA targets. PLoS Biol.
2:e2642004. View Article : Google Scholar
|
|
25
|
Coltella N, Manara MC, Cerisano V,
Trusolino L, et al: Role of the MET/HGF receptor in proliferation
and invasive behavior of osteosarcoma. FASEB J. 17:1162–1164.
2003.PubMed/NCBI
|
|
26
|
Tavazoie SF, Alarcón C, Oskarsson T, Padua
D, Wang Q, et al: Endogenous human microRNAs that suppress breast
cancer metastasis. Nature. 451:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Leite KR, Sousa-Canavez JM, Reis ST,
Tomiyama AH, Camara-Lopes LH, Sañudo A, Antunes AA and Srougi M:
Change in expression of miR-let7c, miR-100, and miR-218 from high
grade localized prostate cancer to metastasis. Urol Oncol.
29:265–269. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mazar J, DeBlasio D, Govindarajan SS,
Zhang S and Perera RJ: Epigenetic regulation of microRNA-375 and
its role in melanoma development in humans. FEBS Lett.
585:2467–2476. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kumar MS, Lu J, Mercer KL, Golub TR and
Jacks T: Impaired microRNA processing enhances cellular
transformation and tumorigenesis. Nat Genet. 39:673–677. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tan S, Li R, Ding K, Lobie PE and Zhu T:
miR-198 inhibits migration and invasion of hepatocellular carcinoma
cells by targeting the HGF/c-MET pathway. FEBS Lett. 585:2229–2234.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu L, Cai C, Wang X, Liu M, Li X and Tang
H: MicroRNA-142-3p, a new regulator of RAC1, suppresses the
migration and invasion of hepatocellular carcinoma cells. FEBS
Lett. 585:1322–1330. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cozzolino AM, Pedace L, Castori M, De
Simone P, Preziosi N, et al: Analysis of the miR-34a locus in 62
patients with familial cutaneous melanoma negative for CDKN2A/CDK4
screening. Fam Cancer. 11:201–208. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yan D, Zhou X, Chen X, Hu D, Dong X, et
al: MicroRNA-34a Inhibits uveal melanoma cell proliferation and
migration through downregulation of c-Met. Invest Ophthalmol Vis
Sci. 50:1559–1565. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao T, Li J and Chen AF: MicroRNA-34a
induces endothelial progenitor cell senescence and impedes its
angiogenesis via suppressing silent information regulator 1. Am J
Physiol Endocrinol Metab. 299:E110–E116. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pang RT, Leung CO, Ye TM, Liu W, Chiu PC,
et al: MicroRNA-34a suppresses invasion through downregulation of
Notch1 and Jagged1 in cervical carcinoma and choriocarcinoma cells.
Carcinogenesis. 31:1037–1044. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Doench JG and Sharp PA: Specificity of
microRNA target selection in translational repression. Genes Dev.
18:504–511. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aruffo A, Stamenkovic I, Melnick M,
Underhill CB and Seed B: CD44 is the principal cell surface
receptor for hyaluronate. Cell. 61:1303–1313. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ponta H, Sherman L and Herrlich PA: CD44:
from adhesion molecules to signalling regulators. Nat Rev Mol Cell
Biol. 4:33–45. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Marhaba R and Zöller M: CD44 in cancer
progression: adhesion, migration and growth regulation. J Mol
Histol. 35:211–231. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gotte M and Yip GW: Heparanase,
hyaluronan, and CD44 in cancers: a breast carcinoma perspective.
Cancer Res. 66:10233–10237. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Olsson E, Honeth G, Bendahl PO, Saal LH,
Gruvberger-Saal S, Ringnér M, et al: CD44 isoforms are
heterogeneously expressed in breast cancer and correlate with tumor
subtypes and cancer stem cell markers. BMC Cancer. 11:4182011.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Heyse TJ, Malcherczyk D, Moll R,
Timmesfeld N, Wapelhorst J, et al: CD44: survival and metastasis in
chondrosarcoma. Osteoarthritis Cartilage. 18:849–856. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen KL, Pan F, Jiang H, Chen JF, Pei L,
Xie FW and Liang HJ: Highly enriched
CD133+CD44+ stem-like cells with
CD133+CD44high metastatic subset in HCT116
colon cancer cells. Clin Exp Metastasis. 28:751–763. 2011.
|
|
44
|
Zhang C, Li C, He F, Cai Y and Yang H:
Identification of CD44+CD24+ gastric cancer
stem cells. J Cancer Res Clin Oncol. 137:1679–1686. 2011.PubMed/NCBI
|
|
45
|
Takaishi S, Okumura T, Tu S, Wang SS,
Shibata W, et al: Identification of gastric cancer stem cells using
the cell surface marker CD44. Stem Cells. 27:1006–1020. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Afify A, Purnell P and Nguyen L: Role of
CD44s and CD44v6 on human breast cancer cell adhesion, migration,
and invasion. Exp Mol Pathol. 86:95–100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kim HS, Park YB, Oh JH, Jeong J, Kim CJ
and Lee SH: Expression of CD44 isoforms correlates with the
metastatic potential of osteosarcoma. Clin Orthop Relat Res.
396:184–190. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Visvader JE and Lindeman GJ: Cancer stem
cells in solid tumours: accumulating evidence and unresolved
questions. Nat Rev Cancer. 8:755–768. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chen R, Nishimura MC, Bumbaca SM, et al: A
hierarchy of self-renewing tumor-initiating cell types in
glioblastoma. Cancer Cell. 17:362–375. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jin L, Hope KJ, Zhai Q, Smadja-Joffe F and
Dick JE: Targeting of CD44 eradicates human acute myeloid leukemic
stem cells. Nat Med. 12:1167–1174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ji Q, Hao X, Zhang M, Tang W, Yang M, et
al: MicroRNA miR-34 inhibits human pancreatic cancer
tumor-initiating cells. PLoS One. 4:e68162009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wiggins JF, Ruffino L, Kelnar K, Omotola
M, Patrawala L, et al: Development of a lung cancer therapeutic
based on the tumor suppressor microRNA-34. Cancer Res.
70:5923–5930. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Huang N, Lin J, Ruan J, Su N, Qing R, Liu
F, He B, Lv C, Zheng D and Luo R: MiR-219-5p inhibits
hepatocellular carcinoma cell proliferation by targeting
glypican-3. FEBS Lett. 586:884–891. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kang HW, Wang F, Wei Q, Zhao YF, Liu M, Li
X and Tang H: miR-20a promotes migration and invasion by regulating
TNKS2 in human cervical cancer cells. FEBS Lett. 586:897–904. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wu L, Li H, Jia CY, Cheng W, Yu M, Peng M,
Zhu Y, Zhao Q, Dong YW, Shao K, Wu A and Wu XZ: MicroRNA-223
regulates FOXO1 expression and cell proliferation. FEBS Lett.
586:1038–1043. 2012. View Article : Google Scholar : PubMed/NCBI
|