Introduction
Oral squamous cell carcinoma (OSCC) accounts for 5%
of all cancers in men and 2% in women (1). The clinical features of OSCC are quite
variable. Some cases show a high aggressive phenotype including
perineural invasion and/or the tendency to metastasize to the lymph
nodes resulting in clinical differences in cancer management. It is
believed that cumulative aberrations in cancer-critical genes
contribute to OSCC carcinogenesis, yet the specific genes that play
a pivotal role in cancer invasion, metastasis or clinical
diverseness remain unidentified. Our research group is focusing on
cancer-specific tumor-suppressor genes (TSGs). To date, we have
investigated the chromosomal loci that harbor frequent allelic loss
in OSCC through loss of heterozygosity (LOH) analysis.
Identification of a candidate TSG is based on the concept that
frequently and specifically deleted alleles in cancer tissue may
conceal important TSGs and we narrowed down a candidate TSG
target.
The Dickkopf (Dkk) family includes candidate TSGs
that we detected in LOH analysis (2,3). Among
the Dkk family members, Dkk-1, -2 and -4 antagonize Wnt ligands and
function as negative regulators of oncogenic Wnt signaling.
Although the Wnt inhibitory function of Dkk-3 is still elusive, the
prevention of nuclear localization (4,5) and
its decreased expression in cancers may signify its possible TSG
function. Downregulation of Dkk mRNA has been reported in a wide
range of malignancies, and studies have focused on Dkk-3 as a
candidate therapeutic target (6–9).
Contradictorily, our previous data suggested an
alternative function of Dkk-3 in OSCC. Patients with LOH in the
Dkk-3 locus exhibited a prolonged overall survival and a low
incident of nodal metastasis (2).
The Dkk-3 protein was dominantly expressed in SCC tissue and cell
lines, and patients without Dkk-3 protein expression did not
undergo metastasis but exhibited prolonged survival (10). Following the various Dkk-3
expression patterns in normal, dysplastic and SCC tissue, Dkk-3
protein expression was found to be increased accompanied by cancer
progression (11). All of these
findings strongly suggest the oncogenic function of Dkk-3.
Due to the possible oncogenic function of Dkk-3, in
the present study we investigated the mRNA expression of Dkk-3 in
OSCC cell lines, and observed the effects of Dkk-3 knockdown by RNA
interference on cell proliferation, migration and invasion.
Materials and methods
Cell lines
A total of 8 cell lines; HSC2, HSC3, HSC4 and Ca9-22
(OSCC), Kato III and AZ521 (gastric adenocarcinoma) and CW-2 and
Colo-320 (colorectal adenocarcinoma) were used in the study. Since
Dkk-3 mRNA was not detected in gastric and colorectal cancer cells
due to CpG methylation, they were used as negative controls
(12). All cell lines were
purchased from Riken BRC through the National BioResource Project
of the MEXT, Japan. Cell lines were maintained in Dulbecco’s
modified Eagle’s medium (DMEM; Life Technologies, Grand Island, NY,
USA) (HCS2, HSC3, HSC4, Ca9-22 and AZ521) or RPMI-1640
(Sigma-Aldrich, St. Louis, MO, USA) (Kato III, CW-2 and Colo-320)
supplemented with 10% fetal bovine serum (FBS, BioWest, Nuaillé,
France) and 100 U/ml and 0.25 μg/ml of
penicillin/streptomycin/amphotericin B (Life Technologies) in a
CO2 incubator with an atmosphere of 95% air plus 5%
CO2 at 37°C.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from the cell culture
according to the acid guanidinium thiocyanate-phenol/chloroform
method using TRIzol® (Invitrogen, Carlsbad, CA, USA).
The final total RNA was dissolved in diethylpyrocarbonate-treated
water, and the absorbance at 260 and 280 nm was measured in a
spectrophotometer (DU-640; Beckman Instruments, Fullerton, CA,
USA). Complementary DNA (cDNA) was synthesized using the
PrimeScript® II 1st Strand cDNA Synthesis kit (Takara,
Shiga, Japan). To synthesize cDNA, 1.0 μl of oligo dT primer and
1.0 μl of dNTP mixture were added to 5 μg of total RNA and then
incubated in a water bath at 65°C for 10 min. Then, 5X
PrimeScript® II buffer, 20 U of RNase inhibitor and 200
U of RTase were added and incubated at 42°C for 2 h.
Quantitative RT-PCR was performed by adding 50 mM of
MgCl2 and 2.5 μl of each specific primer to 2 μl of the
template cDNA reaction mixture to obtain a final concentration of 5
μM primer. Subsequently, 1.25 U of Platinum® Taq DNA
polymerase (Invitrogen) was added, and the final volume was
adjusted to 25 μl. PCR for Dkk-3 was carried out using the
following primers (12): 5′-CTG GGA
GCT AGA GCC TGA TG-3′ (forward) and 5′-TCA TAC TCA TCG GGG ACC
TC-3′ (reverse) for 35 cycles at 94°C for 30 sec, 58°C for 30 sec
and 72°C for 30 sec. For normalization, PCR for human GAPDH was
performed using the Human GAPDH Primer Set kit (Maxim Biotech Inc.,
South San Francisco, CA, USA).
PCR products were confirmed by electrophoresis on a
1.5% agarose gel, and stained with ethidium bromide. The target
amplification products were compared under conditions where PCR
amplification had not reached saturation. The band density was
quantitatively analyzed using a densitometer and the ImageJ program
(http://rsb.info.nih.gov/ij/).
Small interference RNA (siRNA)
Small interference RNA (siRNA) for Dkk-3 and the
negative control non-targeting siRNA were purchased from Thermo
Scientific Dharmacon (Waltham, MA, USA). The targeting sequence of
Dkk-3 siRNA #1 (J-018352-11-0005, ON-TARGETplus siRNA, human Dkk-3)
and Dkk-3 siRNA #2 (J-018352-12-0005, ON-TARGETplus siRNA, human
Dkk-3) were AUCCAUGUGCACCGAGAAA and CCAGAGAGGUCCCCGAUGA,
respectively. As the negative control (siRNA NC), D-001810-01-15,
ON-TARGETplus non-targeting siRNA #1 was used. The siRNA was
suspended in 100 μM of 1X siRNA buffer and diluted with 5X siRNA
buffer (Thermo Scientific Dharmacon) in order to prepare 20 μM
siRNA solutions.
siRNA transfection was performed using
Lipofectamine® RNAiMAX (Invitrogen) according to the
manufacturer’s instructions. Briefly, Ca9-22 or HSC4 cells were
plated at 30–50% confluence in 100-mm cell culture dishes in DMEM
without antibiotics. The following day, 600 pmol of siRNA in 1 ml
Opti-MEM® I reduced serum medium (Invitrogen) and 35 μl
of Lipofectamine® RNAiMAX in 1 ml Opti-MEM®
were mixed. After incubation for 20 min, the mixture was added to
the cultured cells. The cells were maintained in an incubator for 2
days. The knockdown of Dkk-3 mRNA expression was confirmed by
RT-PCR. In order to confirm the effect of the knockdown gene, the
data were collected 2 days and 1 week following siRNA
transfection.
Cell proliferation assay
In order to assess the effect of Dkk-3 knockdown on
cell proliferation, an MTT assay was performed using
TACS® Cell Proliferation Assays (Trevigen, St.
Gaithersburg, MD, USA). Ca9-22 or HSC4, cells with/without siRNA
transfection, were suspended (1.0×103 cells) in 100 μl
of medium supplemented with 10% FBS, plated in 96-well microplates
and cultured for 24 h. MTT (10 μl) was added and incubation was
carried out to form formazan crystals. After 4 h of incubation, 100
μl of each detergent agent was added and the absorbance was
measured at 570 nm. Data were acquired on day 1, 3 and 5.
Migration assay
The cell migration assay was performed using an
Ibidi Culture-insert (Ibidi GmbH, Munich, Germany) following the
manufacturer’s protocol. Briefly, Ca9-22 or HSC4 cells were
cultured, and siRNA transfection was performed. Then, the cells
were harvested using Trypsin/EDTA and resuspended in DMEM with 10%
FBS (~7.0×105 cells/ml). The cell suspension (70 μl) was
transferred to the well of the Culture-insert on a 35-mm dish and
then removed using sterilized tweezers after 24 h of incubation in
a 5% CO2 incubator. Cell migration was observed over
time by adding 2 ml DMEM with 10% FBS. The increase in the area
after 12 h was measured using ImageJ.
Matrigel invasion assay
The BD BioCoat™ Growth Factor Reduced Matrigel™
Invasion Chamber System (Becton-Dickinson, Franklin Lakes, NJ, USA)
was used for the invasion assay according to the manufacturer’s
instructions and based on a previous report (13). Briefly, siRNA transfection of the
Ca9-22 or HSC4 cells was performed as previously described. The
cells were harvested using Trypsin/EDTA and resuspended in
serum-free DMEM (~2.5×105 cells/0.5 ml/well), and seeded
into a 24-well BD Matrigel™ insert and control insert (8-μm pore
size). Inserts were placed in Falcon™ Companion Plates containing
DMEM with 10% FBS and incubated for 24 h. After cell invasion, the
cells were removed from the top chamber using cotton swabs. In
order to count the number of cells invading or migrating to the
other side of the membrane, the membranes were fixed and stained
with Diff-Quik Stain™ (Lab Aids Pty Ltd., North Narrabeen,
Australia) and mounted on a glass slide. The slide was scanned with
Virtual Slide System VS110 (Olympus, Tokyo, Japan), and cells were
counted using the ImageJ program. Cell invasion was compared with
the invasion index and calculated according to the manufacturer’s
instructions.
Western blotting
In order to confirm a decrease in Dkk-3 and how long
it was maintained, western blotting was performed. The data were
acquired in the Ca9-22 cells following 1 week with or without siRNA
transfection. The cell extracts were boiled for 5 min in sodium
dodecyl sulfate (SDS) gel-loading buffer (0.1 M Tris-HCl, pH 6.8,
20% glycerol, 2.5% SDS, 0.05% bromophenol blue and 5%
β-mercaptoethanol). An equal amount of each protein sample was then
loaded and separated onto 12.5% SDS-polyacrylamide gels and blotted
onto polyvinylidene difluoride (PVDF) membranes. After blocking the
non-specific binding by soaking the PVDF membranes in 5% skim milk,
proteins were detected using anti-Dkk-3 (Sigma-Aldrich),
β-catenin-phosphoS37 (Abcam, Cambridge, MA, USA),
non-phospho(active) β-catenin (Ser33/37/Thr41) (Cell Signaling
Technology, Danvers, MA, USA), RhoA, Cdc42 and Rac1 (Abcam).
Anti-tubulin (Sigma-Aldrich) was used for normalization. Proteins
were visualized using the ECL Prime Western Blotting Detection
System (GE Healthcare Life Sciences, Pittsburgh, PA, USA).
Statistical analysis
Significant differences between the control and the
siRNA groups were determined by the two-tailed multiple Student’s
t-test with Bonferoni correction following the Dunnett’s test. All
computations were performed using PASW Statistics 18 (SPSS Inc.,
Chicago, IL, USA). A P-value <0.05 was considered to indicate a
statistically significant result.
Results
Dkk-3 mRNA expression
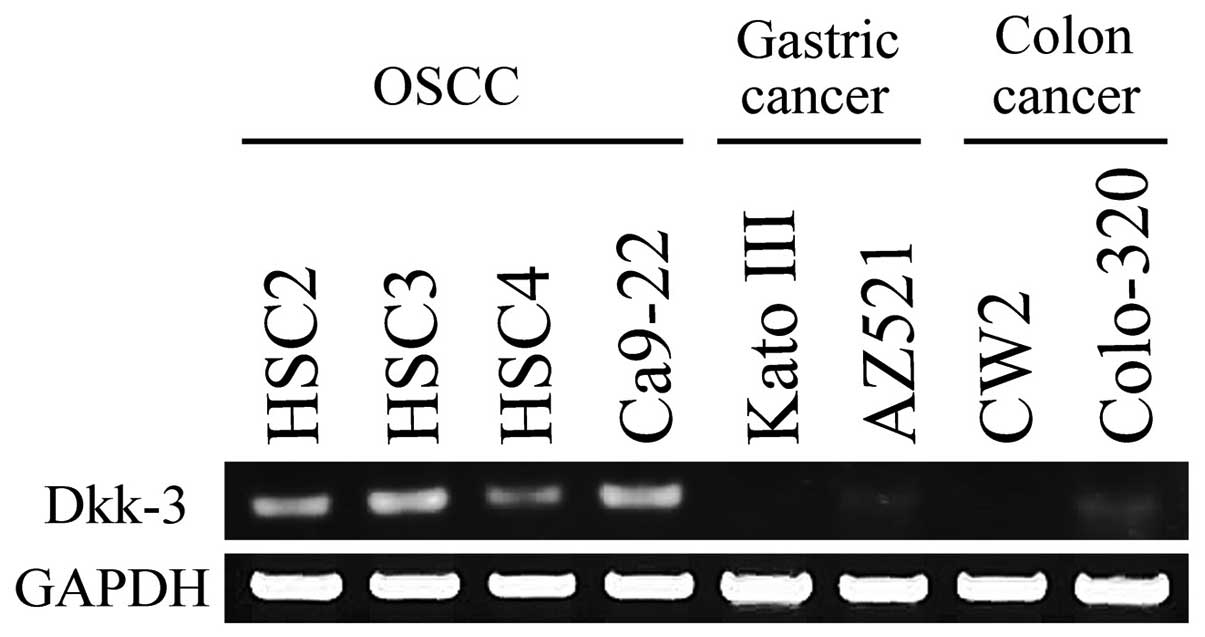
RT-PCR demonstrated Dkk-3 mRNA expression in all
OSCC cell lines. However, expression in gastric cancer cell lines
or colon cancer cell lines were lost or ignorable (Fig. 1). Among the OSCC cell lines, Dkk-3
mRNA expression was comparatively high in the Ca9-22 cells and that
of HSC4 was comparatively low. In the following experiments, Ca9-22
and HSC4 cells were used in order to investigate the effects of
Dkk-3 knockdown by siRNA on cell proliferation, migration and cell
invasion.
Knockdown of Dkk-3 mRNA expression by
siRNA
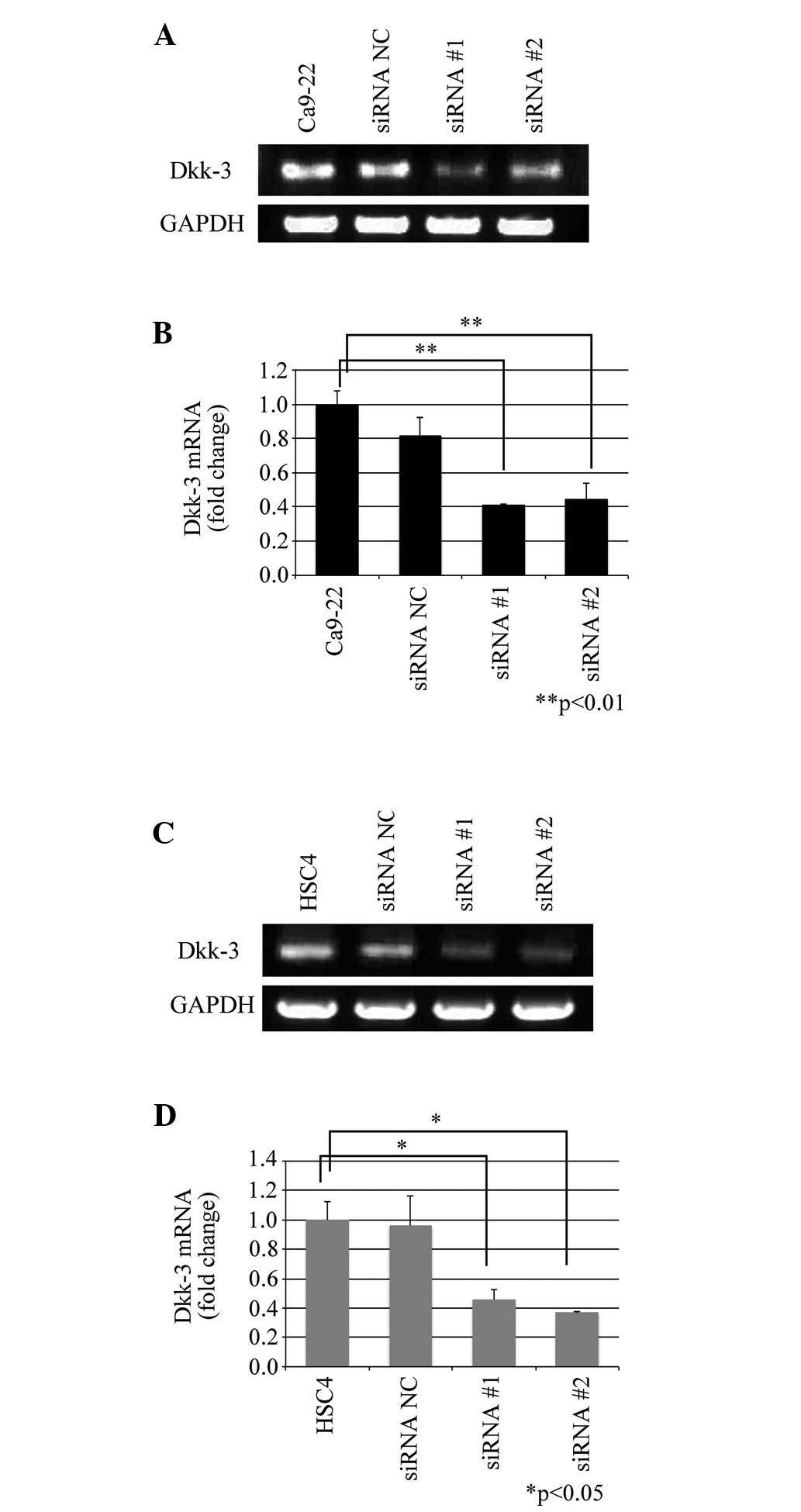
Knockdown of Dkk-3 mRNA expression was performed by
RNA interference. Both of the siRNAs for Dkk-3 (siRNA #1 and siRNA
#2) decreased Dkk-3 expression in the Ca9-22 and HSC4 cells whereas
control siRNA (siRNA NC) did not significantly impact the RNA
expression (Fig. 2A and C). Dkk-3
expressions in Ca9-22 cells was attenuated by 18.9% (siRNA NC,
P=0.175), 59.1% (siRNA #1, P=0.04) and 55.5% (siRNA #2, P=0.005),
respectively (Fig. 2B). The
attenuation in Dkk-3 expression in HSC4 cells was 3.8% (siRNA NC,
P=0.979), 54.2% (siRNA #1, P=0.028) and 64.3% (siRNA #2, P=0.016),
respectively (Fig. 2D).
MTT assay
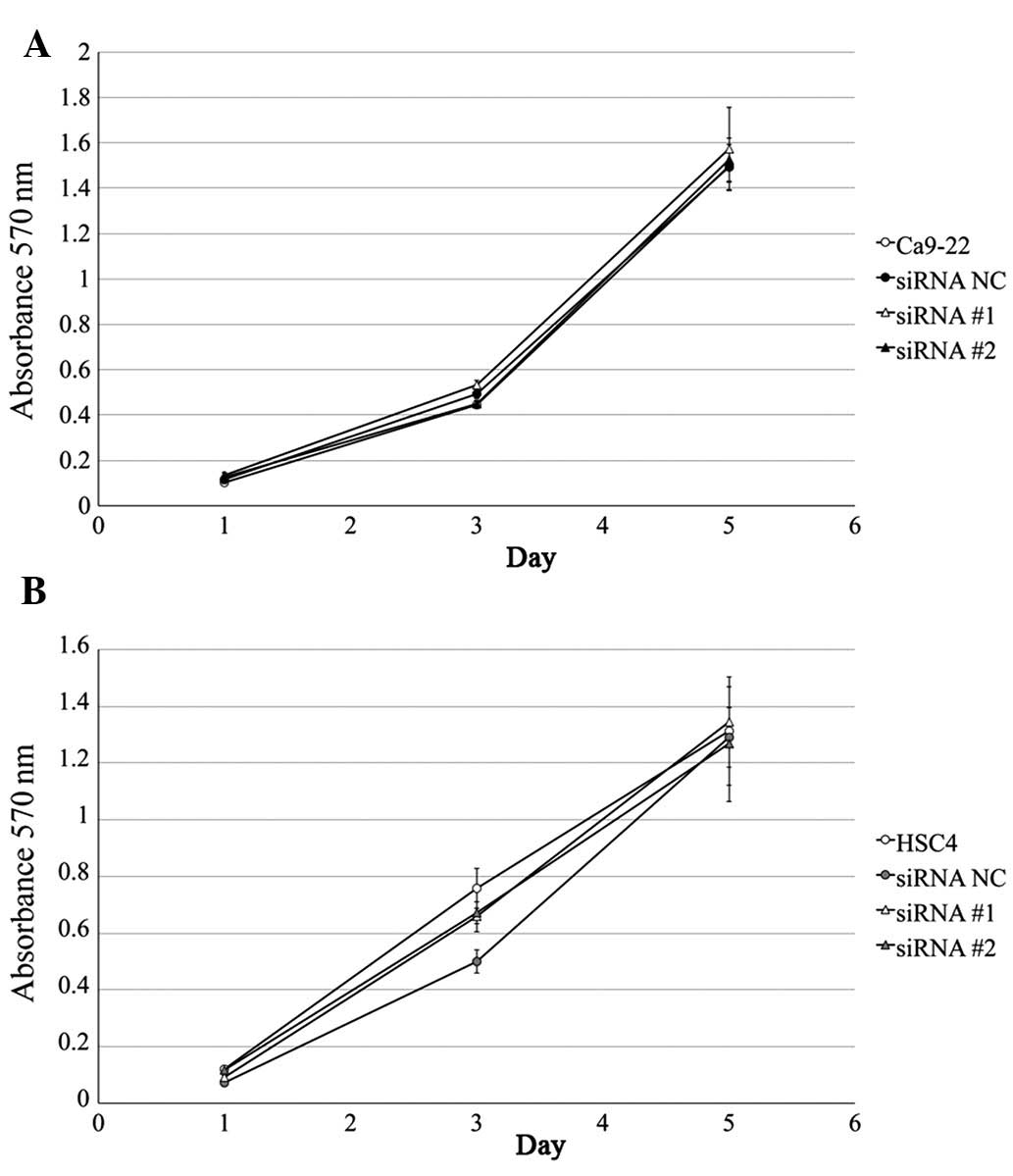
To determine the effect of Dkk-3 knockdown on cell
proliferation in the OSCC cell lines, an MTT cell proliferation
assay was carried out. Cell proliferation was assessed 1, 3 and 5
days following siRNA transfection. No significant differences among
the group were obtained in the Ca9-22 cell line (Fig. 3A). Although transfection with siRNA
NC tended to show decreased proliferation in HSC4 cells, no
significant difference was obtained (Fig. 3B).
Migration assay
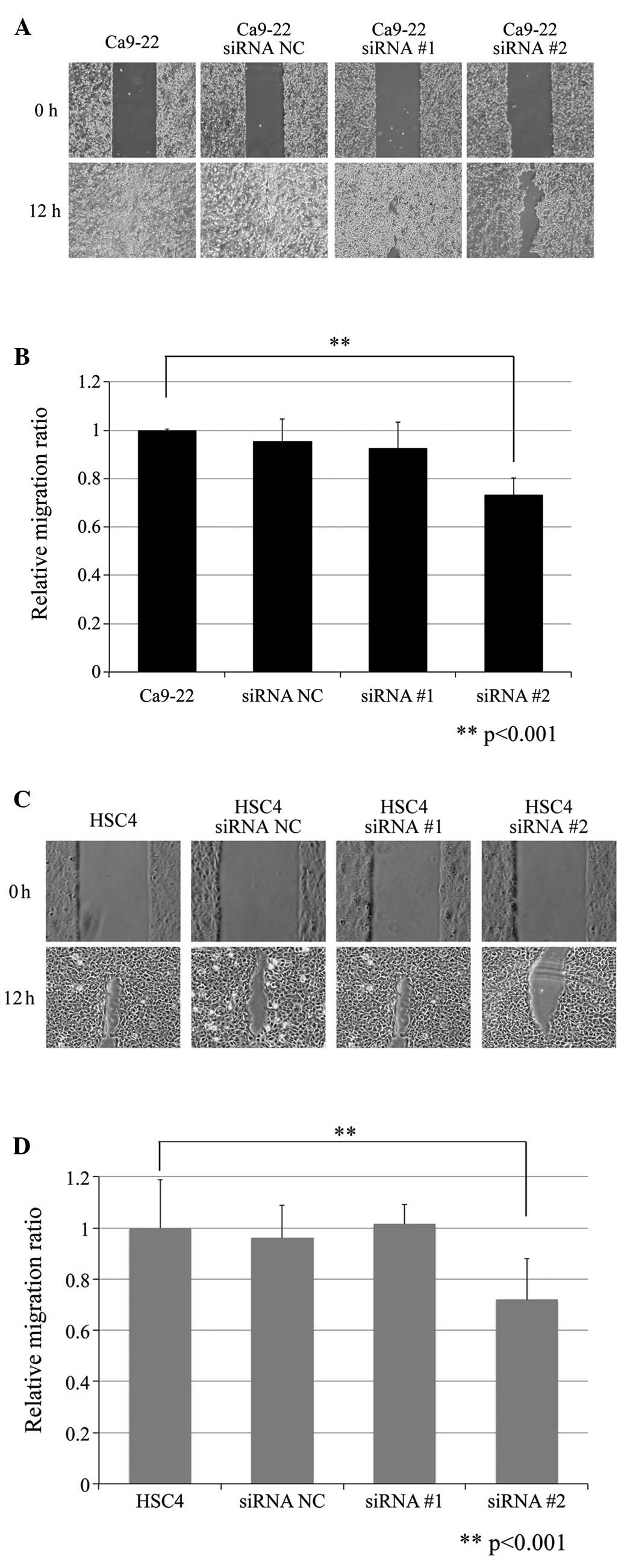
The effect of Dkk-3 knockdown on cell migration is
shown in Fig. 4. Changes in cell
migration were measured as the area recovered after the 12-h
migration and was expressed as the relative change in area.
Notably, Dkk-3 knockdown significantly decreased cell migration
both in the Ca9-22 and HSC4 cells (Fig.
4A and C). Regarding the Ca9-22 cells, the mean area recovery
ratio was 99.84±0.42 (control), 95.34±8.28 (siRNA NC), 92.42±9.83
(siRNA #1) and 73.03±6.56% (siRNA #2), respectively (Fig. 4B). The mean area recovery ratio in
HSC4 cells was 87.76±16.50 (control), 84.23±11.42 (siRNA NC),
89.15±6.82 (siRNA #1) and 63.05±14.06% (siRNA #2), respectively
(Fig. 4D). Statistical analysis
revealed that Dkk-3 knockdown by siRNA #2 significantly decreased
cell migration both in the Ca9-22 and HCS4 cells (P<0.001 and
P=0.033, respectively).
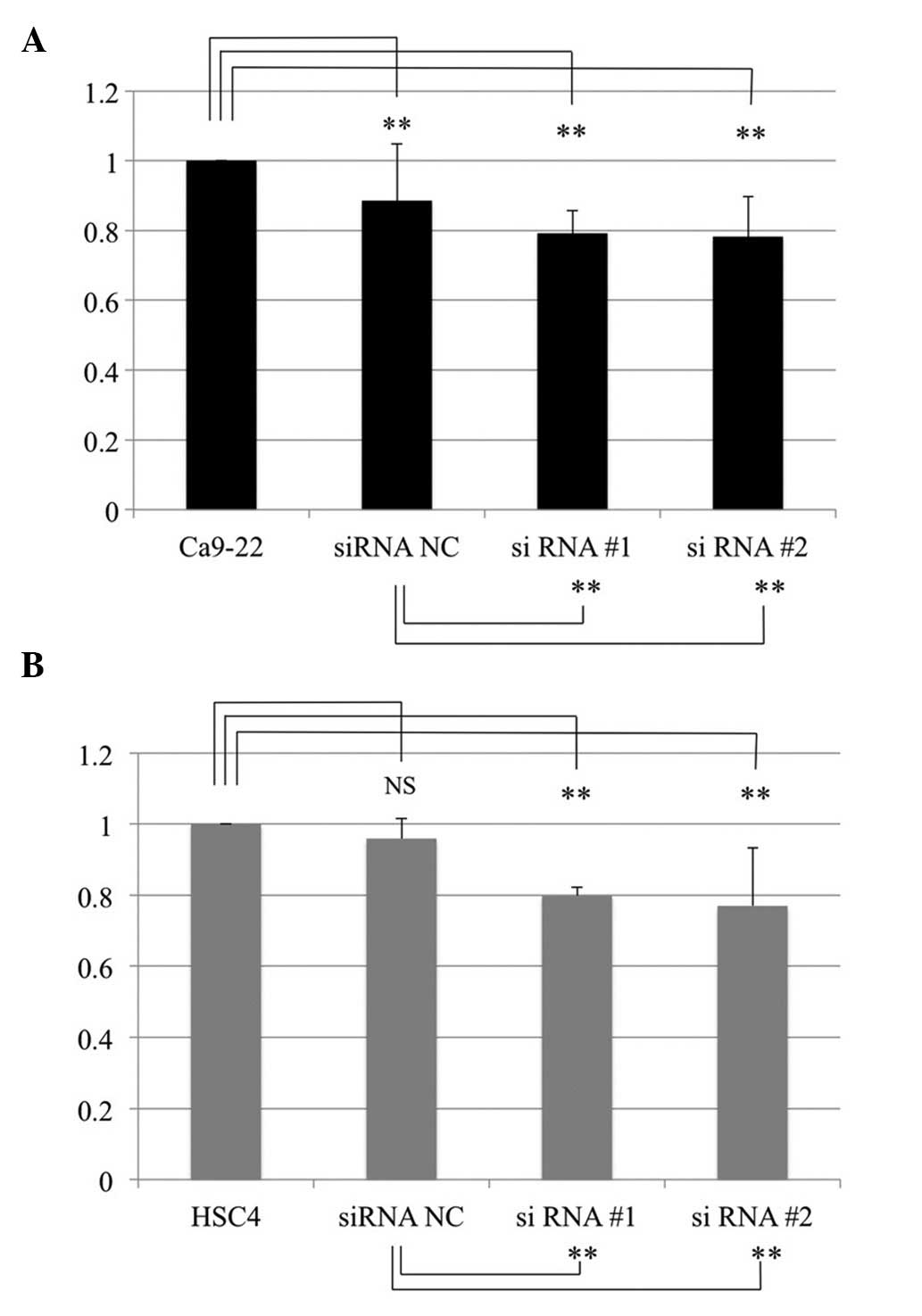
Matrigel invasion assay
In order to investigate the effect of Dkk-3
knockdown on the invasive ability, a Matrigel invasion assay was
performed. Invasive ability was assessed according to the
manufacturer’s protocol. Initially, the percentage of invasion was
calculated by dividing the number of cells that invaded through the
Matrigel insert by the number of cells that migrated through the
control insert. The relative invasion index was then calculated by
dividing the percentage of invasion of the siRNA group with that of
the control cells (Fig. 5). The
relative invasion indices for the Ca9-22 cells were 0.885±0.162
(siRNA NC, P=0.004), 0.793±0.064 (siRNA #1, P<0.001) and
0.783±0.114 (siRNA #2, P<0.001), respectively. Relative indices
for Ca9-22 NC cells were 0.872±0.084 (siRNA #1, P<0.001) and
0.896±0.042 (siRNA #2, P<0.001), respectively.
The relative invasion indices for HSC4 cells were
0.958±0.057 (siRNA NC, P=0.259), 0.798±0.023 (siRNA #1, P<0.001)
and 0.770±0.163 (siRNA #2, P<0.001), respectively. The relative
invasion indices for the HSC4 NC cells were 0.841±0.048 (siRNA #1,
P<0.001) and 0.816±0.137 (siRNA #2, P<0.001), respectively.
The relative invasion indices indicated that siRNA significantly
decreased cell invasion.
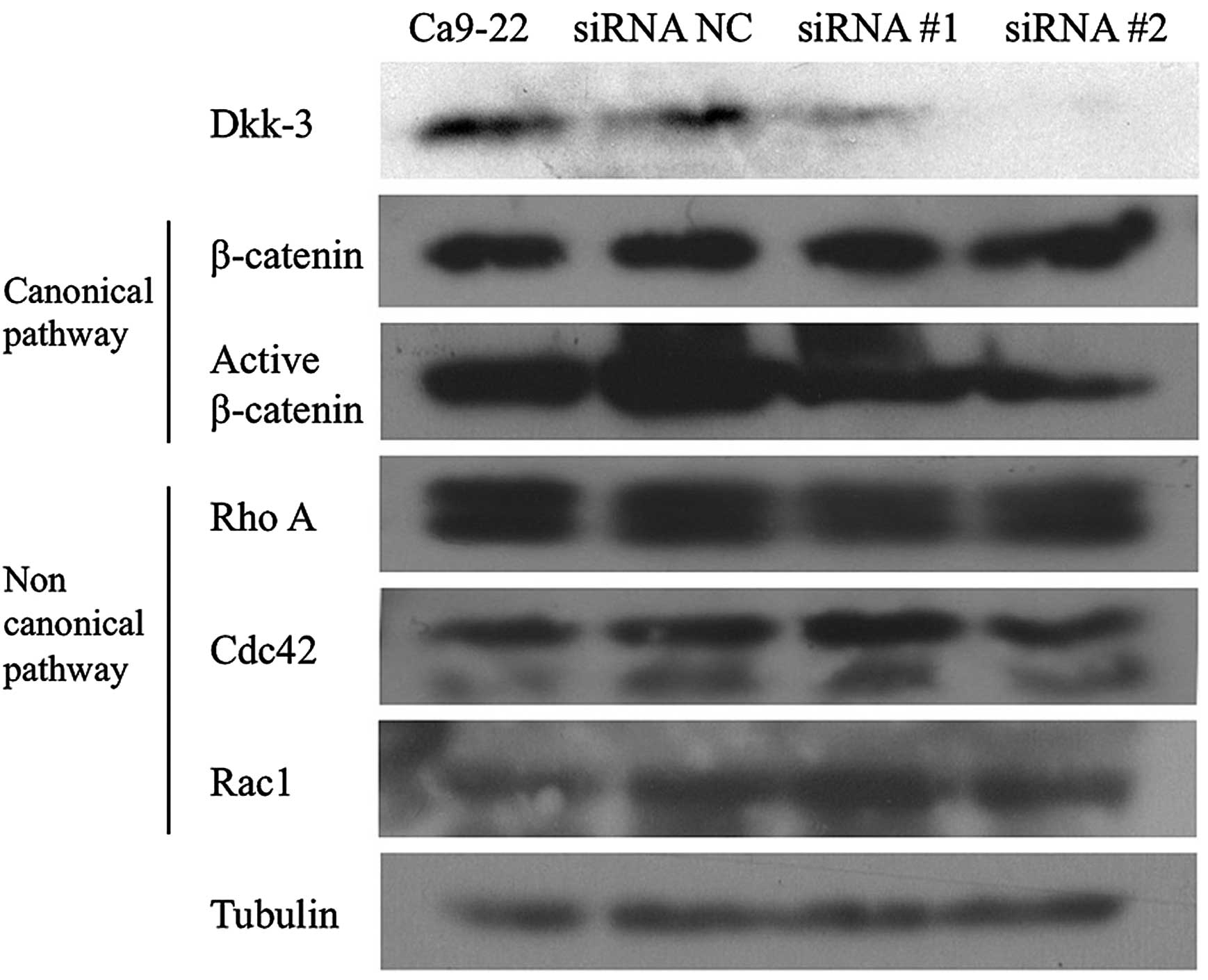
Western blotting
Western blotting revealed that knockdown of Dkk-3
mRNA resulted in protein reduction, even 1 week after siRNA
transfection (Fig. 6). The
expression profile of the proteins that are associated with the Wnt
canonical (β-catenin and active β-catenin) or the non-canonical
pathway (RhoA, cdc42 and Rac1) were determined. Dkk-3 knockdown did
not affect the expression of any protein in either the Wnt
canonical (Wnt/β-catenin) pathway or non-canonical pathway.
Discussion
OSCC is thought to arise as a result of cumulative
genetic aberrations of TSGs or signals that control cell
proliferation, migration and invasion. Wnt signaling and its
regulators are one of the important pathways often dysregulated in
cancer. The Dkk family is one of the Wnt regulators that
antagonizes Wnt ligands together with the Wnt inhibitory factor
(WIF) and secreted frizzled related proteins (sFRPs). Although the
Wnt inhibitory activity of Dkk-3 is still questionable, its
downregulation by CpG methylation was reported in various cancer
types including gastrointestinal tumors (14), hepatocellular carcinoma (15), breast cancer (16), ovarian cancer (17), cervical cancer (18) and glioma (19). Hence, various studies have regarded
Dkk-3 as a putative Wnt signaling inhibitor. In oral cancer,
Pannone et al(20) reported
that Wnt inhibitors including WIF-1, sFRPs and Dkk-3 are
epigenetically inactivated. In this regard, Dkk-3 seems to be a TSG
in which its downregulation is very common regardless of tissue
origin.
Nevertheless, our previous studies showed that the
Dkk family is a novel and independent candidate tumor suppressor in
OSCC by genome-wide LOH analysis (2,3). An
allele in the Dkk-3 locus is frequently deleted in HNSCC suggesting
that Dkk-3 functions as a TSG. However, survival analysis revealed
that Dkk-3 LOH (+) patients exhibited an increased disease-free
survival and significantly prolonged overall survival. LOH analysis
can simply detect allelic loss in particular loci or genes
including true TSGs and genes that have been deleted due to
carcinogenesis. Furthermore, Dkk-3 protein expression was found to
be increased in dysplasia and further in SCC. In addition, Dkk-3
(−) patients exhibited a significantly prolonged disease-free
survival and absence of metastasis (10,11).
Taken together, we hypothesized that Dkk-3 may not only be a TSG
but may also be an oncogene. In order to verify this hypothesis, we
performed functional analyses of Dkk-3 by RNA interference and
determined its effect on the Wnt canonical and non-canonical
pathways.
In the present research, Dkk-3 was detected only in
the OSCC-derived cell lines but not in the gastrointestinal or
colorectal adenocarcinoma-derived cells. Transfection of siRNA in
OSCC cells lines decreased Dkk-3 expression, which did not affect
cell proliferation but did decreased cellular migration. The
present study strongly supports our hypothesis that Dkk-3 possesses
potential oncogenic function. Furthermore, this may explain our
previous data that patients lacking Dkk-3 protein expression showed
significantly longer disease-free and metastasis-free survival.
Moreover, because OSCC cell lines express Dkk-3 mRNA, the loss of
Dkk-3 mRNA expression is not a part of the carcinogenic process in
oral epithelium.
The mechanism by which Dkk-3 exerts oncogenic
function remains unknown. We hypothesized the Wnt canonical and
non-canonical pathways as probable targets. Firstly, the canonical
Wnt pathway (β-catenin pathway) is a major signaling pathway that
participates in carcinogenesis. The present data revealed that both
β-catenin and active β-catenin were equally observed regardless of
Dkk-3 protein expression status, implying that Dkk-3 may not
function as regulator of Wnt/β-catenin signaling in OSCC and that
aberrant β-catenin expression is independent of Dkk-3
expression.
β-catenin acts not only as a transcriptional
activator that mediates Wnt signaling but also as a structural
protein in cell adhesion junctions (21,22).
An in vitro study using an OSCC cell line demonstrated that
aberrant β-catenin accumulation in the cytoplasm induces
TCF/Lef-mediated transcriptional activity, increased matrix
metalloproteinase (MMP)-7 expression and induced epithelial
mesenchymal transition (EMT), resulting in higher
invasion/migration capacities (23). The function of nuclear β-catenin
(active β-catenin) in the canonical pathway is also controversial.
Tenbaum et al(24) reported
that β-catenin provides resistance to AKT inhibitors and that
apoptosis-related molecule FOXO3a changes its function promoting
metastasis when it co-localizes with β-catenin in colon cancer
cells. Therefore, it is likely that Dkk-3 accumulated in cytoplasm
may act differently from secreted Dkk-3 and that the oncogenic
function of Dkk-3 is closely related to aberrantly accumulated
β-catenin.
Secondly, recent reports have indicated that the
non-canonical Wnt pathway may play an important role in cancer
invasion. Particularly, the Rho family of small GTPases acting
downstream of Wnt receptor function in cell migration (25). The Wnt/Rho kinase pathway is related
to stress fiber formation, and the Wnt/Cdc42/Rac1 pathway is
associated in lamellipodia formation in cell migration. The Rho
family of GTPases, RhoA and Rho family members Cdc42 were reported
to regulate E-cadherin-dependent cellular adhesion and may interact
with β-catenin (23). The present
data showed that Dkk-3 knockdown affected neither the Wnt/Rho
kinase pathway nor Wnt/Cdc42/Rac1 expression, indicating no evident
linkage between Dkk-3 and the Wnt non-canonical pathway.
On the other hand, Dkk-3 seems to possess various
functions. Dkk-3, otherwise known as REIC (reduced expression in
cancer), implies a tumor-suppressive function (26). Dkk-3 is now the clinical target of
gene therapy in pancreatic (7),
prostate (27,28), gastric scirrhous (8), breast (29) and testicular cancer (30). Yet, these reports conclude that the
tumor inhibitory function of Dkk-3 is caused by apoptosis induction
due to endoplasmic reticulum stress or JNK phosphorylation and not
by its primary tumor suppressor function. The possibility that
Dkk-3 mutation may exhibit a dominant-negative function warrants
discussion. To date, there is no report on Dkk-3 genetic mutations
in malignancies, although many reports suggest epigenetic changes
in Dkk-3.
Although the present results further support our
previous data that patients with Dkk-3 expression demonstrated a
shorter disease-free and metastasis-free survival, the results
provide another point of interest in the functional role of Dkk-3
in OSCC. In conclusion, increased invasion and migration by Dkk-3
knockdown may be driven by a mechanism other than Wnt/β-catenin
signaling, yet Dkk-3 may be a clinical target for prevention of
OSCC invasion.
Acknowledgements
This study was partially supported by a Grant-in-Aid
for Scientific Research from the Ministry of Education, Culture,
Sports, Science and Technology, Japan to N.K. (22791766), H.T.
(22791977), R.T. (24592766) and H.N. (24659891).
References
|
1
|
Johnson N, Franceschi S, Ferlay J, Ramadas
K, Schmid S, MacDonald DG, Bouquet JE and Slootweg PJ: Squamous
cell carcinoma. WHO Classification of Tumours. Pathology and
Genetics of Head and Neck Tumours. Barnes L, Eveson JW, Reichart P
and Sideransky D: IARC Press; Lyon: pp. 168–175. 2005
|
|
2
|
Katase N, Gunduz M, Beder L, Gunduz E,
Lefeuvre M, Hatipoglu OF, Borkosky SS, Tamamura R, Tominaga S,
Yamanaka N, Shimizu K, Nagai N and Nagatsuka H: Deletion at
Dickkopf (dkk)-3 locus (11p15.2) is related with lower lymph node
metastasis and better prognosis in head and neck squamous cell
carcinomas. Oncol Res. 17:273–282. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Katase N, Gunduz M, Beder LB, Gunduz E, Al
Sheikh Ali M, Tamamura R, Yaykasli KO, Yamanaka N, Shimizu K and
Nagatsuka H: Frequent allelic loss of Dkk-1 locus (10q11.2) is
related with low distant metastasis and better prognosis in head
and neck squamous cell carcinomas. Cancer Invest. 28:103–110. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Veeck J and Dahl E: Targeting the Wnt
pathway in cancer: the emerging role of Dickkopf-3. Biochim Biophys
Acta. 1825:18–28. 2012.PubMed/NCBI
|
|
5
|
Hoang BH, Kubo T, Healey JH, Yang R,
Nathan SS, Kolb EA, Mazza B, Meyers PA and Gorlick R: Dickkopf 3
inhibits invasion and motility of Saos-2 osteosarcoma cells by
modulating the Wnt-beta-catenin pathway. Cancer Res. 64:2734–2739.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gu YM, Ma YH, Zhao WG and Chen J:
Dickkopf3 overexpression inhibits pancreatic cancer cell growth in
vitro. World J Gastroenterol. 17:3810–3817. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Abarzua F, Sakaguchi M, Takaishi M, Nasu
Y, Kurose K, Ebara S, Miyazaki M, Namba M, Kumon H and Huh NH:
Adenovirus-mediated overexpression of REIC/Dkk-3 selectively
induces apoptosis in human prostate cancer cells through activation
of c-Jun-NH2-kinase. Cancer Res. 65:9617–9622. 2005. View Article : Google Scholar
|
|
8
|
Than SS, Kataoka K, Sakaguchi M, Murata H,
Abarzua F, Taketa C, Du G, Yashiro M, Yanagihara K, Nasu Y, Kumon H
and Huh NH: Intraperitoneal administration of an adenovirus vector
carrying REIC/Dkk-3 suppresses peritoneal dissemination of
scirrhous gastric carcinoma. Oncol Rep. 25:989–995. 2011.
|
|
9
|
Sakaguchi M, Kataoka K, Abarzua F,
Tanimoto R, Watanabe M, Murata H, Than SS, Kurose K, Kashiwakura Y,
Ochiai K, Nasu Y, Kumon H and Huh NH: Overexpression of REIC/Dkk-3
in normal fibroblasts suppresses tumor growth via induction of
interleukin-7. J Biol Chem. 284:14236–14244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Katase N, Lefeuvre M, Gunduz M, Gunduz E,
Beder LB, Grenman R, Fujii M, Tamamura R, Tsujigiwa H and Nagatsuka
H: Absence of Dickkopf (Dkk)-3 protein expression is correlated
with longer disease-free survival and lower incidence of metastasis
in head and neck squamous cell carcinoma. Oncol Lett. 3:273–280.
2012.PubMed/NCBI
|
|
11
|
Fujii M, Katase N, Lefeuvre M, Gunduz M,
Buery RR, Tamamura R, Tsujigiwa H and Nagatsuka H: Dickkopf (Dkk)-3
and β-catenin expressions increased in the transition from normal
oral mucosal to oral squamous cell carcinoma. J Mol Histol.
42:499–504. 2011.
|
|
12
|
Maehata T, Taniguchi H, Yamamoto H, Nosho
K, Adachi Y, Miyamoto N, Miyamoto C, Akutsu N, Yamaoka S and Itoh
F: Transcriptional silencing of Dickkopf gene family by CpG island
hypermethylation in human gastrointestinal cancer. World J
Gastroenterol. 14:2702–2714. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hughes L, Malone C, Chumsri S, Burger AM
and McDonnell S: Characterisation of breast cancer cell lines and
establishment of a novel isogenic subclone to study migration,
invasion and tumourigenicity. Clin Exp Metastasis. 25:549–557.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sato H, Suzuki H, Toyota M, Nojima M,
Maruyama R, Sasaki S, Takagi H, Sogabe Y, Sasaki Y, Idogawa M,
Sonoda T, Mori M, Imai K, Tokino T and Shinomura Y: Frequent
epigenetic inactivation of DICKKOPF family genes in human
gastrointestinal tumors. Carcinogenesis. 28:2459–2566. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ding Z, Qian YB, Zhu LX and Xiong QR:
Promoter methylation and mRNA expression of DKK-3 and WIF-1 in
hepatocellular carcinoma. World J Gastroenterol. 15:2595–2601.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Veeck J, Wild PJ, Fuchs T, Schüffler PJ,
Hartmann A, Knüchel R and Dahl E: Prognostic relevance of
Wnt-inhibitory factor-1 (WIF1) and Dickkopf-3 (DKK3) promoter
methylation in human breast cancer. BMC Cancer. 9:2172009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
You A, Fokas E, Wang LF, He H, Kleb B,
Niederacher D, Engenhart-Cabillic R and An HX: Expression of the
Wnt antagonist DKK3 is frequently suppressed in sporadic epithelial
ovarian cancer. J Cancer Res Clin Oncol. 137:621–627. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
van der Meide WF, Snellenberg S, Meijer
CJ, Baalbergen A, Helmerhorst TJ, van der Sluis WB, Snijders PJ and
Steenbergen RD: Promoter methylation analysis of WNT/β-catenin
signaling pathway regulators to detect adenocarcinoma or its
precursor lesion of the cervix. Gynecol Oncol. 123:116–122.
2011.
|
|
19
|
Götze S, Wolter M, Reifenberger G, Müller
O and Sievers S: Frequent promoter hypermethylation of Wnt pathway
inhibitor genes in malignant astrocytic gliomas. Int J Cancer.
126:2584–2593. 2010.PubMed/NCBI
|
|
20
|
Pannone G, Bufo P, Santoro A, Franco R,
Aquino G, Longo F, Botti G, Serpico R, Cafarelli B, Abbruzzese A,
Caraglia M, Papagerakis S and Lo Muzio L: WNT pathway in oral
cancer: epigenetic inactivation of WNT-inhibitors. Oncol Rep.
24:1035–1041. 2010.PubMed/NCBI
|
|
21
|
Laxmidevi LB, Angadi PV, Pillai RK and
Chandreshekar C: Aberrant β-catenin expression in the histologic
differentiation of oral squamous cell carcinoma and verrucous
carcinoma: an immunohistochemical study. J Oral Sci. 52:633–640.
2010.
|
|
22
|
Yu Z, Weinberger PM, Provost E, Haffty BG,
Sasaki C, Joe J, Camp RL, Rimm DL and Psyrri A: beta-Catenin
functions mainly as an adhesion molecule in patients with squamous
cell cancer of the head and neck. Clin Cancer Res. 11:2471–2477.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Iwai S, Yonekawa A, Harada C, Hamada M,
Katagiri W, Nakazawa M and Yura Y: Involvement of the Wnt-β-catenin
pathway in invasion and migration of oral squamous carcinoma cells.
Int J Oncol. 37:1095–1103. 2010.
|
|
24
|
Tenbaum SP, Ordóñez-Morán P, Puig I,
Chicote I, Arqués O, Landolfi S, Fernández Y, Herance JR, Gispert
JD, Mendizabal L, Aguilar S, Ramón y Cajal S, Schwartz S Jr,
Vivancos A, Espín E, Rojas S, Baselga J, Tabernero J, Muñoz A and
Palmer HG: β-catenin confers resistance to PI3K and AKT inhibitors
and subverts FOXO3a to promote metastasis in colon cancer. Nat Med.
18:892–901. 2012.
|
|
25
|
Ishida-Takagishi M, Enomoto A, Asai N,
Ushida K, Watanabe T, Hashimoto T, Kato T, Weng L, Matsumoto S,
Asai M, Murakumo Y, Kaibuchi K, Kikuchi A and Takahashi M: The
Dishevelled-associating protein Daple controls the non-canonical
Wnt/Rac pathway and cell motility. Nat Commun. 3:8592012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tsuji T, Miyazaki M, Sakaguchi M, Inoue Y
and Namba M: A REIC gene shows down-regulation in human
immortalized cells and human tumor-derived cell lines. Biochem
Biophys Res Commun. 268:20–24. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kawauchi K, Watanabe M, Kaku H, Huang P,
Sasaki K, Sakaguchi M, Ochiai K, Huh NH, Nasu Y and Kumon H:
Preclinical safety and efficacy of in situ REIC/Dkk-3 gene therapy
for prostate cancer. Acta Med Okayama. 66:7–16. 2012.PubMed/NCBI
|
|
28
|
Edamura K, Nasu Y, Takaishi M, Kobayashi
T, Abarzua F, Sakaguchi M, Kashiwakura Y, Ebara S, Saika T,
Watanabe M, Huh NH and Kumon H: Adenovirus-mediated REIC/Dkk-3 gene
transfer inhibits tumor growth and metastasis in an orthotopic
prostate cancer model. Cancer Gene Ther. 14:765–772. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kawasaki K, Watanabe M, Sakaguchi M,
Ogasawara Y, Ochiai K, Nasu Y, Doihara H, Kashiwakura Y, Huh NH,
Kumon H and Date H: REIC/Dkk-3 overexpression downregulates
P-glycoprotein in multidrug-resistant CF7/ADR cells and induces
apoptosis in breast cancer. Cancer Gene Ther. 16:65–72. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tanimoto R, Abarzua F, Sakaguchi M,
Takaishi M, Nasu Y, Kumon H and Nuh NH: REIC/Dkk-3 as a potential
gene therapeutic agent against human testicular cancer. Int J Mol
Med. 19:363–368. 2007.PubMed/NCBI
|