Introduction
The prognosis for malignant melanoma is generally
poor, and effective treatment relies on early diagnosis and
surgical excision with wide margins (1). Understanding the mechanisms that
control the proliferation of malignant melanoma cells is crucial
for developing new diagnostic methods and therapies.
Cyclic AMP (cAMP) is an intracellular second
messenger. A balance between the synthesis and degradation of cAMP,
catalyzed by adenylate cyclase and phosphodiesterase (PDE),
respectively, regulates its intracellular concentration and thus
its various physiological functions (2). The PDE superfamily of enzymes is
divided into 11 families (PDE1-PDE11), and 21 genes have been
identified. Approximately half of the PDE families contain more
than one gene, and most of the individual genes can produce
multiple mRNAs and protein sequences through alternative
transcriptional start sites or alternative splicing. These isoforms
show different distributions within a cell, which serve to
compartmentalize specific cAMP and cyclic GMP (cGMP) signaling
pathways to modulate their responses to different stimuli (2,3).
In mammals, some PDEs selectively hydrolyze cAMP or
cGMP, while others hydrolyze both cAMP and cGMP. PDE2A hydrolyzes
both substrates, and its binding to cGMP enhances the hydrolysis of
cAMP (2,4). The PDE2A monomer is a protein of
approximately 103-kDa that contains GAF-A, GAF-B and catalytic
domains. PDE2A forms a homodimer, likely through the GAF-A domain.
The GAF-B domain binds to cGMP, while the catalytic domain is
responsible for enzymatic activity (4–6). The
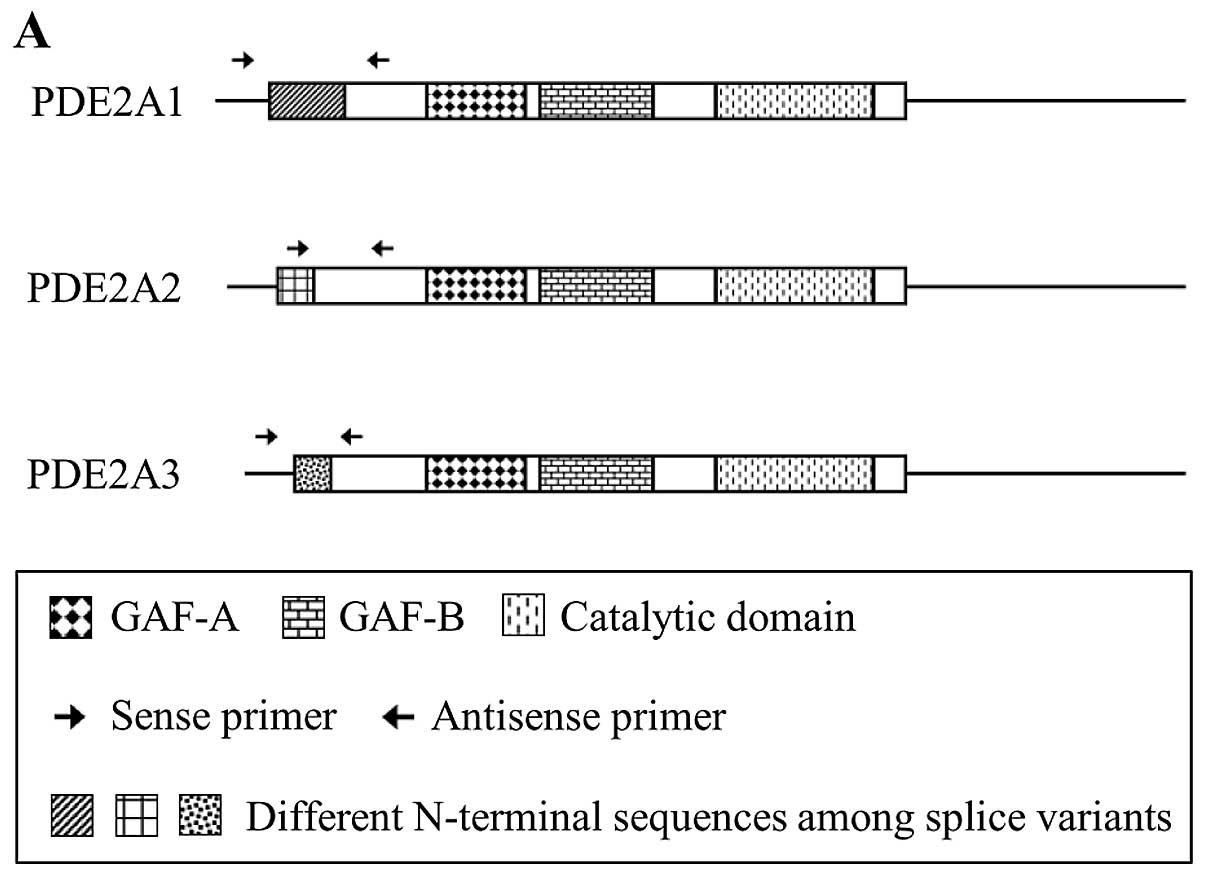
three splicing variants (PDE2A1, PDE2A2 and PDE2A3) of PDE2A each
have a distinct N-terminal structure. These differences in the
N-terminal structure appear to be responsible for the distinct
subcellular localization of these variants, which determines their
interactions with selective binding partners (4). Bovine PDE2A1 is soluble, while rat
PDE2A2 and human PDE2A3 are membrane-bound enzymes (2,4).
We previously reported that PDE1, PDE3 and PDE5 are
expressed in human malignant melanoma cells (7–9).
Understanding the role of PDE2, which remains largely unknown,
could be of value for developing new treatment strategies for
malignant melanoma. Herein, we examined the expression of splicing
variants and mutants of PDE2A in human oral malignant melanoma PMP
cells. We also investigated the effect of PDE2 inhibition on cell
motility, cytotoxicity, DNA synthesis, apoptosis and cell cycle
progression.
Materials and methods
Materials
Cell culture media (RPMI-1640) and fetal bovine
serum (FBS) were purchased from Invitrogen (Carlsbad, CA, USA).
Phosphate-buffered saline (PBS, lacking Ca2+ and
Mg2+) and erythro-9-(2-hydroxy-3-nonyl)-adenine (EHNA)
were obtained from Sigma (St. Louis, MO, USA).
3-(4,5-Dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
(MTS) was obtained from Promega (Madison, WI, USA). The GFX PCR DNA
and Gel Band Purification kit and DYEnamic ET Terminator Cycle
Sequencing kit were purchased from GE Healthcare (Little Chalfont,
UK). Diff-Quik stain™ was from Sysmex Co. (Kobe, Japan). Human
brain total RNA was from Agilent Technologies (Santa Clara, CA,
USA).
Cell culture
Human malignant melanoma PMP cells were obtained
from a 65-year-old patient with primary palatal malignant melanoma
as previously described (10).
Cells were maintained in RPMI-1640 medium supplemented with 5% FBS,
at 37°C in a humidified 5% CO2 atmosphere.
Reverse transcription-PCR (PDE2A splicing
variants)
PMP cells were seeded at 1×106 cells/25
cm2 flask. After 3 days, total RNA was isolated with the
QuickGene RNA Cultured Cell Kit S (Fuji Photo Film Co., Tokyo,
Japan). First-strand cDNA was synthesized using total RNA with the
High Capacity RNA-to-cDNA kit (Applied Biosystems, Foster City, CA,
USA). PCR was performed with primer pairs specific for PDE2 splice
variants (Table I, Fig. 1A). PCR amplification was carried out
in a total volume of 50 μl containing PCR buffer (with 1.5 mM
MgCl2), 200 μM dNTPs, 2.5 units HotStarTaq™ DNA
polymerase (Qiagen, Hilden, Germany) and 0.5 μM sense and antisense
primers. HotStarTaq DNA polymerase was activated by incubation of
the reactions at 95°C for 15 min. For PDE2A1 and PDE2A2, this
activation step was followed by 35 cycles of amplification (94°C
for 1 min, 60°C for 1 min and 72°C for 1 min) and 72°C for 10 min.
For PDE2A3, this activation step was followed by 38 cycles of
amplification (94°C for 1 min, 60°C for 1 min and 72°C for 1 min)
and 72°C for 10 min. Products were subjected to electrophoresis on
2.5% agarose gels and visualized by SYBR® Green nucleic
acid gel staining (Invitrogen).
 | Table IPrimers used to amplify the PDE2A
splice variants. |
Table I
Primers used to amplify the PDE2A
splice variants.
| Sequence |
|---|
| PDE2A1 |
| Sense primer |
5′-GATTGGGGTGTTGAGTCCAG-3′ |
| Antisense
primer |
5′-TCGATGACAGAGCCCAGACT-3′ |
| PDE2A2 |
| Sense primer |
5′-CACATCCTCATCGCTGTTGT-3′ |
| Antisense
primer |
5′-AGATGATAGCCTCCCGGACT-3′ |
| PDE2A3 |
| Sense primer |
5′-TGGAAGTGCAGAGCTTAGCC-3′ |
| Antisense
primer |
5′-GTCGATGACAGAGCCCAGAC-3′ |
Western blotting
PMP cells were suspended in NuPAGE LDS sample buffer
(Invitrogen). Gel electrophoresis [NuPAGE 4–12% Bis-Tris gel
(Invitrogen)] was performed in an XCell SureLock Mini-Cell
(Invitrogen). Following electrophoresis, proteins were transferred
to iBlot Transfer stack PVDF membranes using iBlot (Invitrogen),
and the membranes were blocked by incubation with PBST (0.1%
Tween-20 in PBS) supplemented with 2% ECL Advance Blocking Agent
(GE Healthcare) for 1 h. The blots were incubated with primary
rabbit polyclonal antibody (PD2A-101AP; FabGennix, Frisco, TX, USA)
overnight at 4°C and rinsed five times with PBST. Rinsed blots were
incubated with horseradish peroxidase-conjugated donkey anti-rabbit
IgG (GE Healthcare) for 1 h and rinsed with PBST. Immunoreactivity
was detected by chemiluminescence using ECL Advance Western
Blotting detection kits (GE Healthcare). The protein content was
determined using BCA protein assay kits (Pierce, Rockford, IL,
USA).
Sequencing of PDE2A2
PMP cells were seeded at 1×106 cells/25
cm2 flask. After 3 days, total RNA was isolated with
QuickGene RNA Cultured Cell Kit S. First-strand cDNA was
synthesized using total RNA with SuperScript® II Reverse
Transcriptase (Invitrogen). PCR was performed with specific primer
pairs for PDE2A2 (Table II and
Fig. 2A). PCR amplification was
performed in a total volume of 50 μl containing PCR buffer for
KOD-Plus™, 1.0 mM MgSO4, 200 μM dNTPs, 1 unit KOD-Plus
(Toyobo, Osaka, Japan), and 0.3 μM sense and antisense primers. KOD
DNA polymerase was activated by incubating the reactions at 96°C
for 30 min followed by 30 cycles of amplification (94°C for 30 sec
and 68°C for 3 min) and 68°C for 5 min. Products were subjected to
electrophoresis on 1% agarose gels and visualized by SYBR Green
nucleic acid gel staining. The PCR product generated by the PDE2A2
primers was purified using an Illustra™ GFX™ PCR DNA Gel Band
Purification kit and verified by DNA sequencing.
| Table IIPrimers used to amplify the PDE2A2
sequence. |
Table II
Primers used to amplify the PDE2A2
sequence.
| Sequence |
|---|
| PDE2A2 |
| Sense primer
(Fw1) |
5′-GGACCAGGCGAAGCTGTCGC-3′ |
| Antisense primer
(Rv1) |
5′-CTGAGGCCCAGGAAGGTAGTAC-3′ |
In vitro migration assay
PMP cells (4×104 cells) in RPMI-1640
medium containing 0.1% FBS were transferred to 8-μm pore inserts
(BD Biosciences), which were then placed in companion wells
containing RPMI-1640 medium supplemented with 10% FBS as a
chemoattractant. Following a 12-h incubation, the inserts were
removed, and the non-migrating cells on the upper surface were
harvested using a cotton swab. Cells on the lower surface of the
membrane were fixed and stained with Diff-Quik and counted under a
microscope.
Cell growth assay
The cells were plated at a density of 400 cells/well
in 96-well plates, allowed to adhere for 24 h, and then cultured in
the absence or presence of different concentrations of EHNA for 3
or 5 days. MTS assays were performed using CellTiter 96®
Aqueous One Solution Cell Proliferation Assay (Promega), and the
number of viable cells was counted.
Trypan blue exclusion test
PMP cells (3.3×104) were plated in 25
cm2 flasks and cultured for 24 h. Then, the cells were
treated with EHNA for 3 days (50 and 100 μM) or 5 days (10, 20, 50,
and 100 μM). Trypan blue stain 0.4% (Invitrogen) was added to the
cell suspension, and both live and dead cells were counted under
the microscope.
BrdU cell proliferation assay
DNA synthesis inhibition was detected by a BrdU cell
proliferation assay using the Amersham Cell Proliferation Biotrak
ELISA System, version 2 (GE Healthcare). Cells were plated at 2000
cells/well in 96-well plates. After 24 h of culture, the cells were
treated with EHNA (1, 10, 20, 50 and 100 μM) for 24 h. Cells were
treated with solution A [BrdU labeling reagent plus RPMI-1640
(1:1000, v/v)] and incubated at 37°C for 2 h. After aspirating
solution A, the cells were treated with fixative for 30 min,
followed by solution B [blocking reagent plus redistilled water
(1:100, v/v)]for 30 min. After aspirating solution B, the cells
were treated with solution C [peroxidase-labeled anti-BrdU stock
solution plus antibody dilution solution (1:100, v/v)] for 90 min.
The cells were then washed three times with solution D [wash buffer
concentrate plus redistilled water (1:10, v/v)]. The cells were
treated with substrate solution for 5 min in the dark. After adding
1 M sulfuric acid to the cells, the optical density was measured in
a microplate reader at 450 nm.
Terminal deoxynucleotidyl
transferase-mediated dUTP nick end-labeling assay
Apoptosis-related DNA fragmentation was detected
using a terminal deoxynucleotidyl transferase-mediated dUTP nick
end-labeling (TUNEL) assay using an Apop-Tag Plus Peroxidase in
situ Apoptosis Detection kit (Millipore, Billerica, MA, USA).
Cells were seeded in Lab-Tek chamber slides (Nalge Nunc
International, Rochester, NY, USA) at a density of 530 cells per
well. After culturing for 24 h, the cells were treated with EHNA
(50 and 100 μM) for 24 h. The cells were then fixed in 1%
paraformaldehyde for 10 min. The fixed cells were preserved in
precooled ethanol plus acetic acid (2:1, v/v) for 5 min.
Equilibration buffer was added, and the cells were incubated at
37°C for 1 h in a terminal deoxynucleotidyl transferase (TdT)
enzyme solution containing
deoxyuridine-5′-triphosphate-digoxigenin. After the reaction was
stopped with a pre-warmed stop/wash buffer, the cells were
incubated with an anti-digoxigenin antibody fragment carrying a
conjugated peroxidase in a humidified chamber for 30 min at room
temperature. Peroxidase activity was detected using
3,3′-diaminobenzidine as a substrate. Methyl green was applied for
20 min at room temperature for counterstaining. Total and apoptotic
cell numbers were counted in 10 different fields in each well under
a microscope, and the average apoptotic cell number was expressed
as the percentage of the total cell number to denote the apoptotic
index.
Cell cycle analysis by flow
cytometry
Cell cycle progression was analyzed using a
CycleTEST™ Plus DNA Reagent kit (BD Biosciences). Cells were plated
at 3.2×104 cells in 25 cm2 flasks and
cultured for 24 h. Then, cells were treated with EHNA (50 and 100
μM) for 5 days. A cell suspension was made from the 25
cm2 flasks. The cell suspension was centrifuged, the
supernatant was aspirated, the buffer solution was added to the
cells, and the cells were gently vortexed. After performing the
same procedure twice, cells were counted and transferred to 15-ml
plastic tubes (5×105 cells/tube). The cells were
centrifuged, the supernatant was aspirated, and solution A was
added to the cells and incubated for 10 min, after which solution B
was added to the cells and incubated for 10 min. Solution C was
then added to the cells, incubated for 10 min on ice in the dark,
and the cells were analyzed by flow cytometry.
RT-PCR (CDKs and cyclins)
PMP cells (4×104) were plated in 25
cm2 flasks and cultured for 24 h. Then, cells were
treated with EHNA (50 and 100 μM) for 5 days. The total RNA of the
cells was isolated using a QuickGene RNA Cultured Cell Kit S.
First-strand cDNA was synthesized using total RNA with a High
Capacity RNA-to-cDNA kit. PCR was performed with primer pairs
specific for GAPDH, cyclins and CDKs (Table III). PCR amplification was carried
out in a total volume of 50 μl containing PCR buffer (with 1.5 mM
MgCl2), 200 μM dNTPs, 2.5 units HotStarTaq DNA
polymerase (Qiagen) and 0.5 μM sense and antisense primers.
HotStarTaq DNA polymerase was activated by incubation of the
reactions at 95°C for 15 min. For GAPDH, this activation step was
followed by 18 cycles of amplification (94°C for 1 min, 59°C for 1
min and 72°C for 1 min) with a final extension step at 72°C for 10
min. For cyclin A, this activation step was followed by 31 cycles
of amplification (94°C for 1 min, 60°C for 1 min and 72°C for 1
min) and a final extension step at 72°C for 10 min. For cyclin B1,
this activation step was followed by 24 cycles of amplification
(94°C for 1 min, 58°C for 1 min and 72°C for 1 min) and a final
extension step at 72°C for 10 min. For cyclin D1, this activation
step was followed by 25 cycles of amplification (94°C for 1 min,
68°C for 1 min and 72°C for 1 min) and a final extension step at
72°C for 10 min. For cyclin E, this activation step was followed by
34 cycles of amplification (94°C for 1 min, 68°C for 1 min and 72°C
for 1 min) and a final extension step at 72°C for 10 min. For CDK1,
this activation step was followed by 25 cycles of amplification
(94°C for 1 min, 52.5°C for 1 min and 72°C for 1 min) and a final
extension step at 72°C for 10 min. For CDK2, this activation step
was followed by 23 cycles of amplification (94°C for 1 min, 60°C
for 1 min and 72°C for 1 min) and a final extension step at 72°C
for 10 min. For CDK4, this activation step was followed by 23
cycles of amplification (94°C for 1 min, 63°C for 1 min and 72°C
for 1 min) and a final extension step at 72°C for 10 min. PCR
products were subjected to electrophoresis on 2.5% agarose gels and
visualized by SYBR Green nucleic acid gel staining.
| Table IIIPrimers used to amplify GAPDH, CDKs
and cyclins. |
Table III
Primers used to amplify GAPDH, CDKs
and cyclins.
| Sequence |
|---|
| GAPDH |
| Sense primer |
5′-ACAGTCAGCCGCATCTTCTT-3′ |
| Antisense
primer |
5′-TGAGCTTGACAAAGTGGTCG-3′ |
| CDK1 |
| Sense primer |
5′-GATTCTATCCCTCCTGGTC-3′ |
| Antisense
primer |
5′-TAGGCTTCCTGGTTTCC-3′ |
| CDK2 |
| Sense primer |
5′-GCTTTCTGCCATTCTCATCG-3′ |
| Antisense
primer |
5′-GTCCCCAGAGTCCGAAAGAT-3′ |
| CDK4 |
| Sense primer |
5′-ACGGGTGTAAGTGCCATCTG-3′ |
| Antisense
primer |
5′-TGGTGTCGGTGCCTATGGGA-3′ |
| Cyclin A |
| Sense primer |
5′-TCCAAGAGGACCAGGAGAATATCA-3′ |
| Antisense
primer |
5′-TCCTCATGGTAGTCTGGTACTTCA-3′ |
| Cyclin B1 |
| Sense primer |
5′-AAGAGCTTTAAACTTTGGTCTGGG-3′ |
| Antisense
primer |
5′-CTTTGTAAGTCCTTGATTTACCATG-3′ |
| Cyclin D1 |
| Sense primer |
5′-TGGATGCTGGAGGTCTGCGAGGAA-3′ |
| Antisense
primer |
5′-GGCTTCGATCTGCTCCTGGCAGGC-3′ |
| Cyclin E |
| Sense primer |
5′-AGTTCTCGGCTCGCTCCAGGAAGA-3′ |
| Antisense
primer |
5′-TCTTGTGTCGCCATAATCCGGTCA-3′ |
Statistical analysis
All of the experiments were performed in triplicate.
Differences in multiple group comparisons were analyzed using the
Tukey-Kramer multiple comparisons test. Significance was defined as
calculated P-values of <0.01.
Results
Expression of PDE2A splice variants in
PMP cells
We first examined the expression of PDE2A splice
variants (PDE2A1, PDE2A2 and PDE2A3) in PMP cells by RT-PCR using
three specific primer pairs (Table
I and Fig. 1A). PDE2A1 and
PDE2A3 amplicons were not detectable, suggesting that these two
splice variants were expressed at extremely low levels, or not
expressed at all. Primers specific for PDE2A2 yielded a doublet by
PCR (Fig. 1B), which was shown by
sequence analysis to include a lower band derived from PDE2A2
(GenBank NM_001143839.3) and an upper band corresponding to the
non-coding transcript variant 4 of PDE2A (GenBank NR_026572.2) with
an additional 62 bp compared with PDE2A2 (data not shown). Western
blot analysis showed a single band (~105 kDa) corresponding to
PDE2A2 (Fig. 1C).
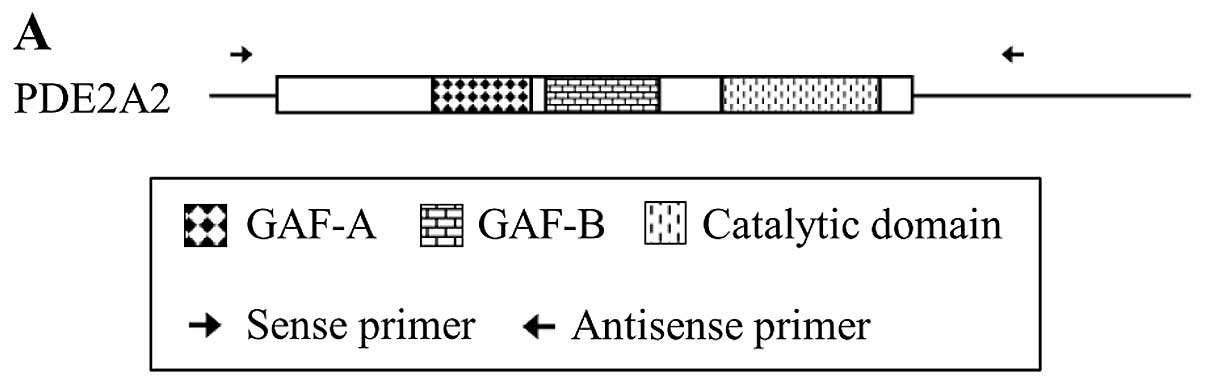
Sequencing of PDE2A2 in PMP cells
We next examined the mutations in PDE2A2 expressed
in PMP cells. RT-PCR using a different set of primers (Table II and Fig. 2A) yielded a 3031-bp band (Fig. 2B). Sequencing of this RT-PCR product
revealed point mutations at nucleotide positions 734 (C to T), 1800
(T to C), 2268 (C to T) and 2718 (C to T). The missense mutation at
position 734 resulted in the substitution of threonine with
isoleucine at amino acid 214 (Fig.
2C). The other three were silent mutations.
Effect of EHNA on the migration of PMP
cells
Since cell motility plays an important role in
metastasis, we examined whether PDE2A2 is involved in cell
motility. Cells treated with 100 μM EHNA migrated in a manner
similar to that of control cells (Fig.
3), indicating that inhibition of PDE2A2 did not affect the
motility of PMP cells.
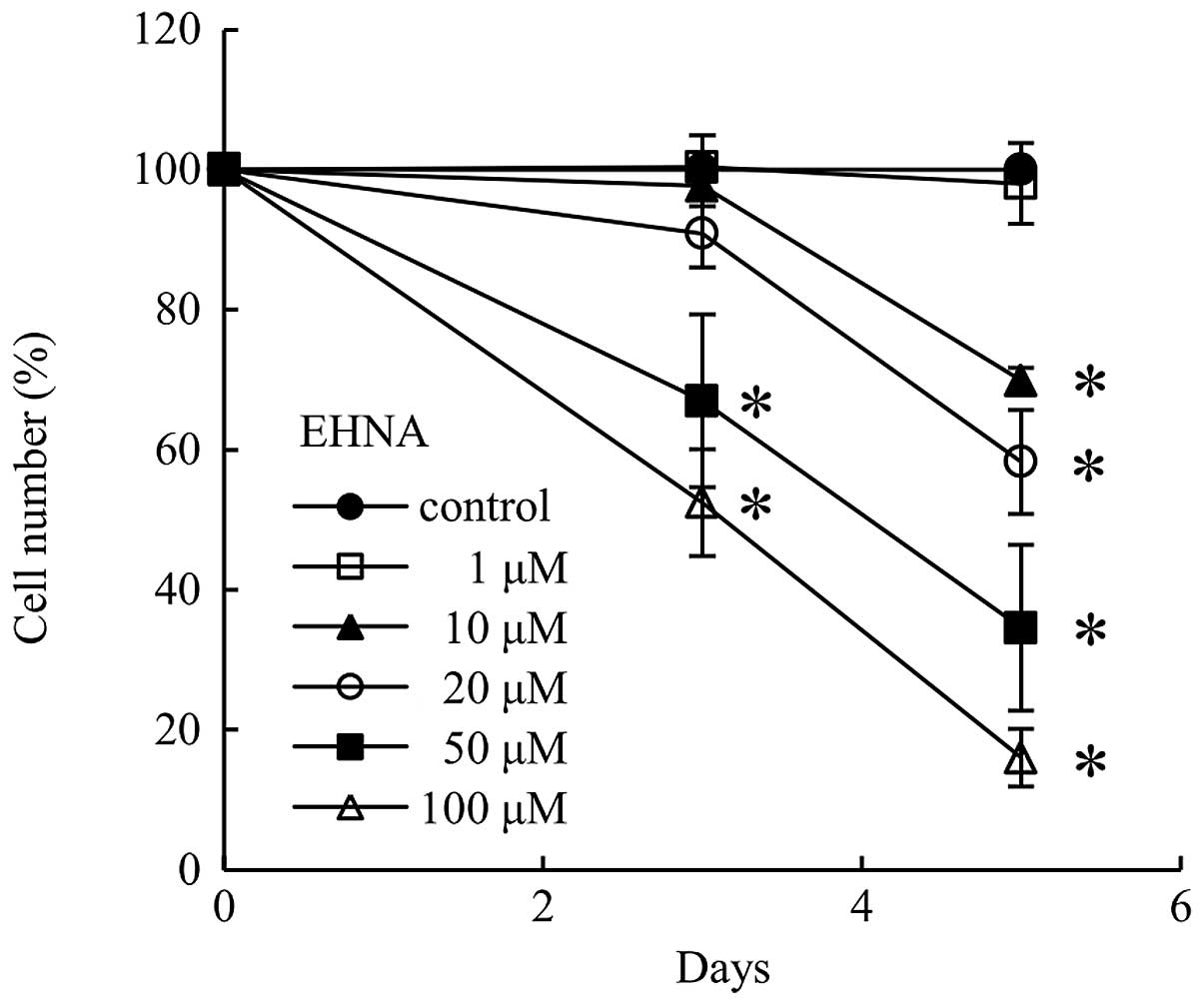
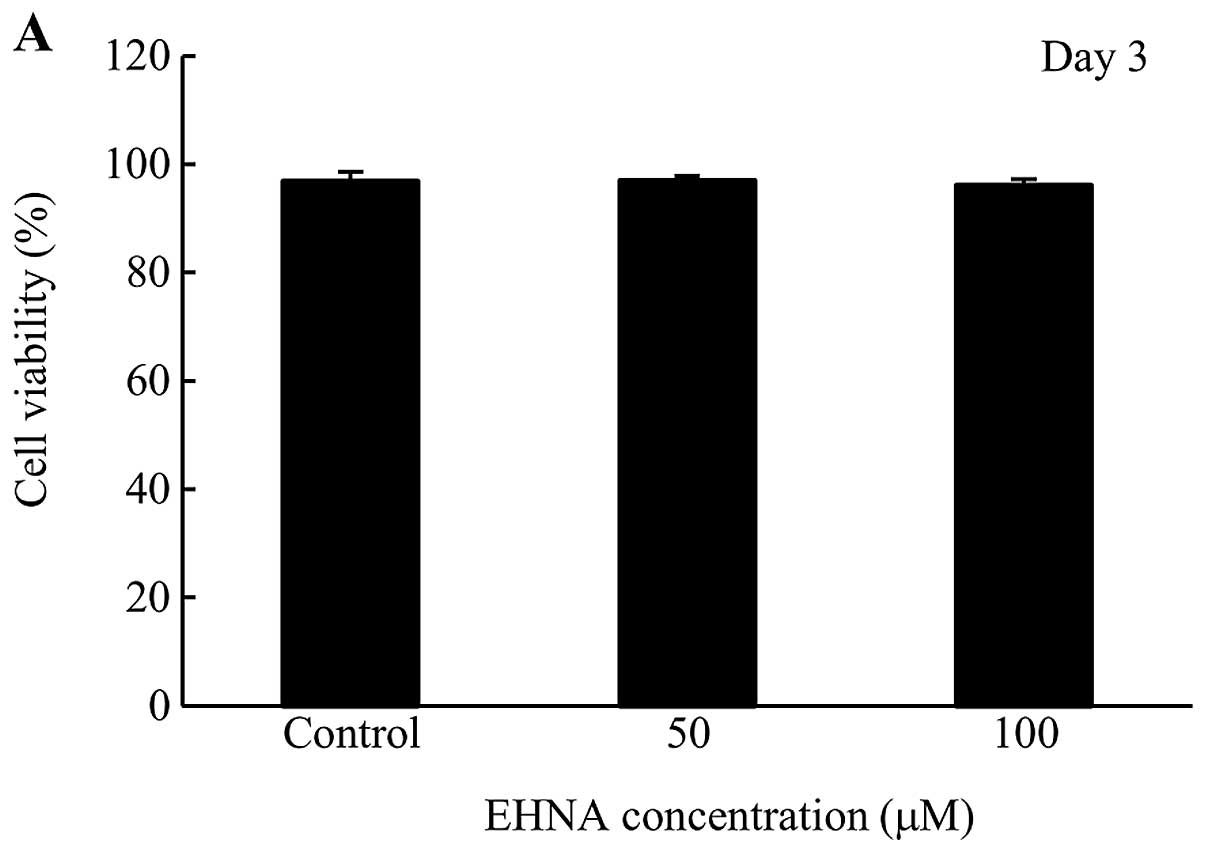
Cytotoxicity of EHNA in PMP cells
Since EHNA-treated PMP cells grew at a slower rate
than control cells (Fig. 4), we
tested the cytotoxicity of EHNA using a trypan blue exclusion
assay. No cytotoxicity was observed at day 3 and 5 following the
addition of EHNA to PMP cells (Fig.
5).
Effect of EHNA on DNA synthesis in PMP
cells
To examine the involvement of PDE2A in DNA
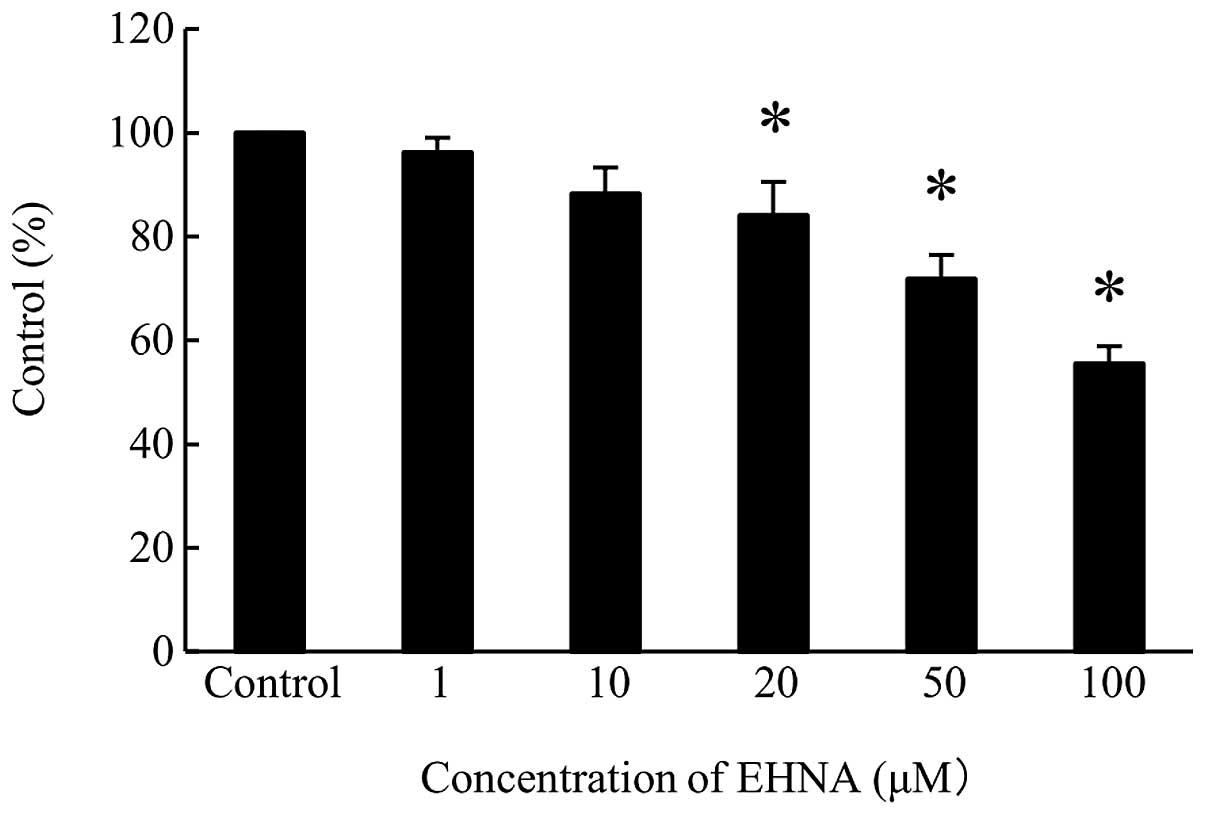
synthesis, PMP cells were treated with 20, 50 and 100 μM EHNA,
which decreased DNA synthesis levels to 15, 25 and 40%,
respectively, of the control cell level (Fig. 6).
Effect of EHNA on the apoptosis of PMP
cells
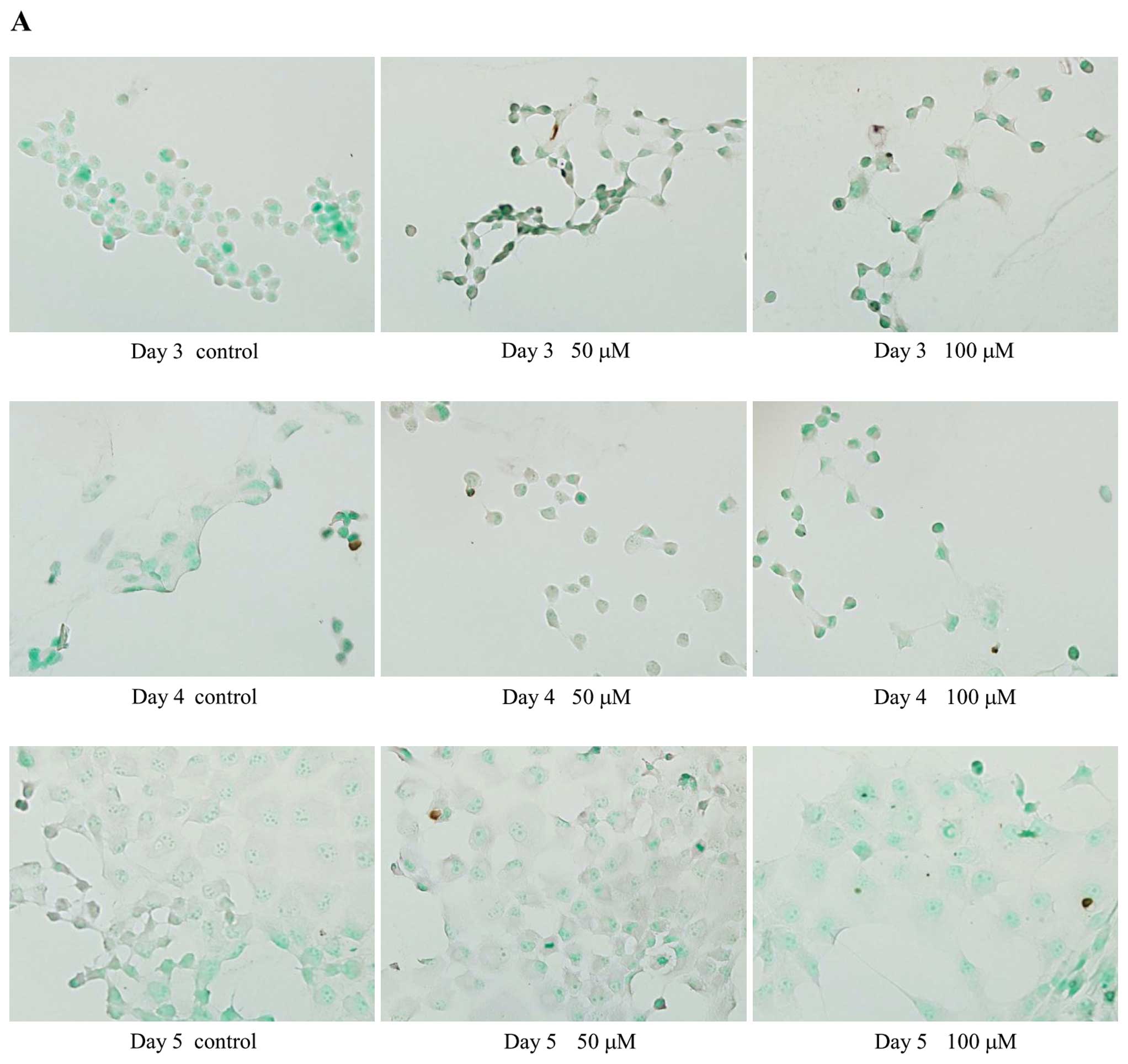
The TUNEL assay was used to examine the effect of
PDE2A inhibition on apoptosis. TUNEL-positive cells were rarely
observed in EHNA-treated or control cells (Fig. 7), suggesting that PDE2A is not
involved in apoptosis.
Effect of EHNA on cell cycle progression
in PMP cells
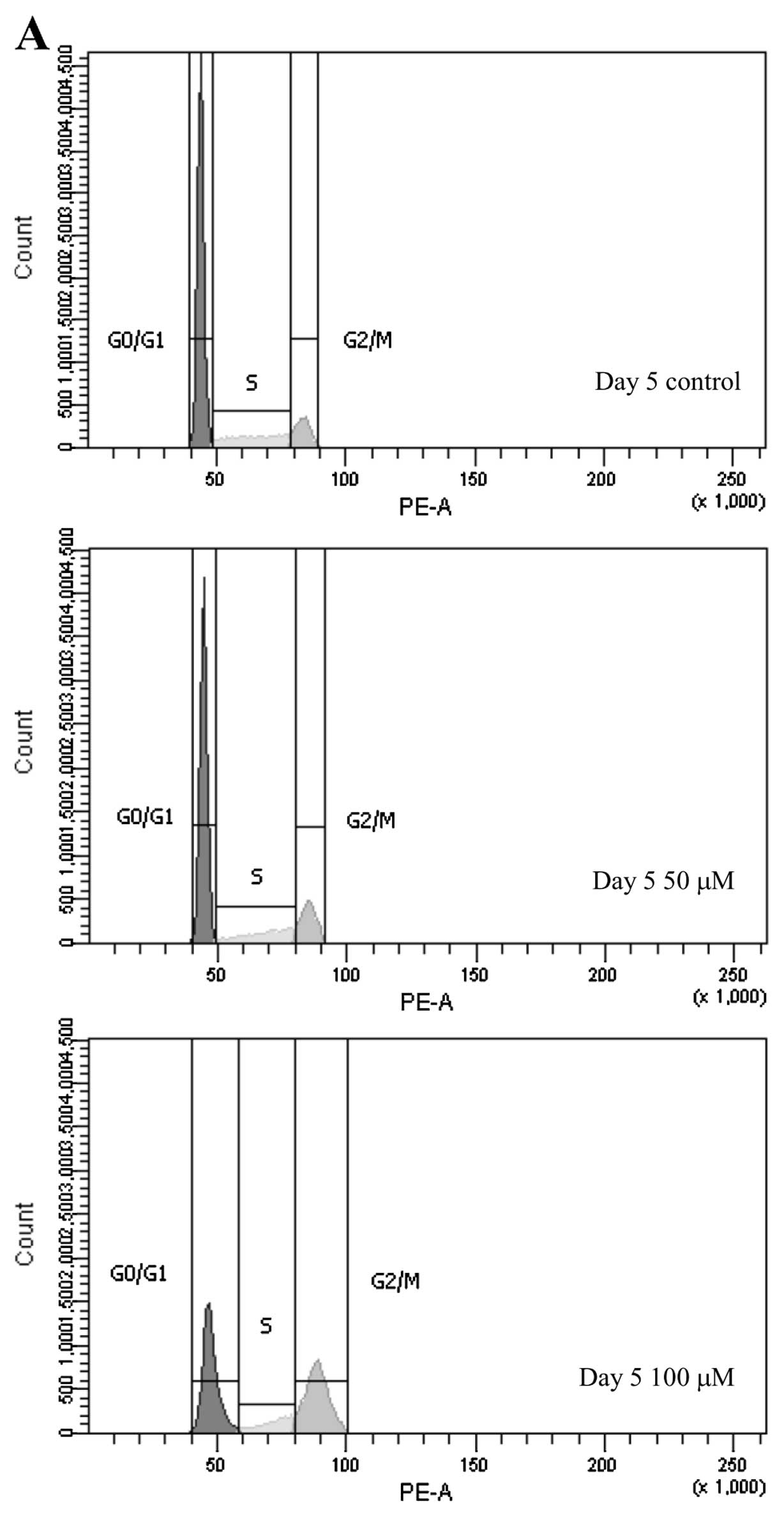
Since DNA synthesis was suppressed in PMP cells
treated with EHNA, we investigated the effect of PDE2 on cell cycle
progression. EHNA treatment for 5 days decreased the proportion of
PMP cells in the G0/G1 phase, and increased
the number of cells in the G2/M phase (Fig. 8).
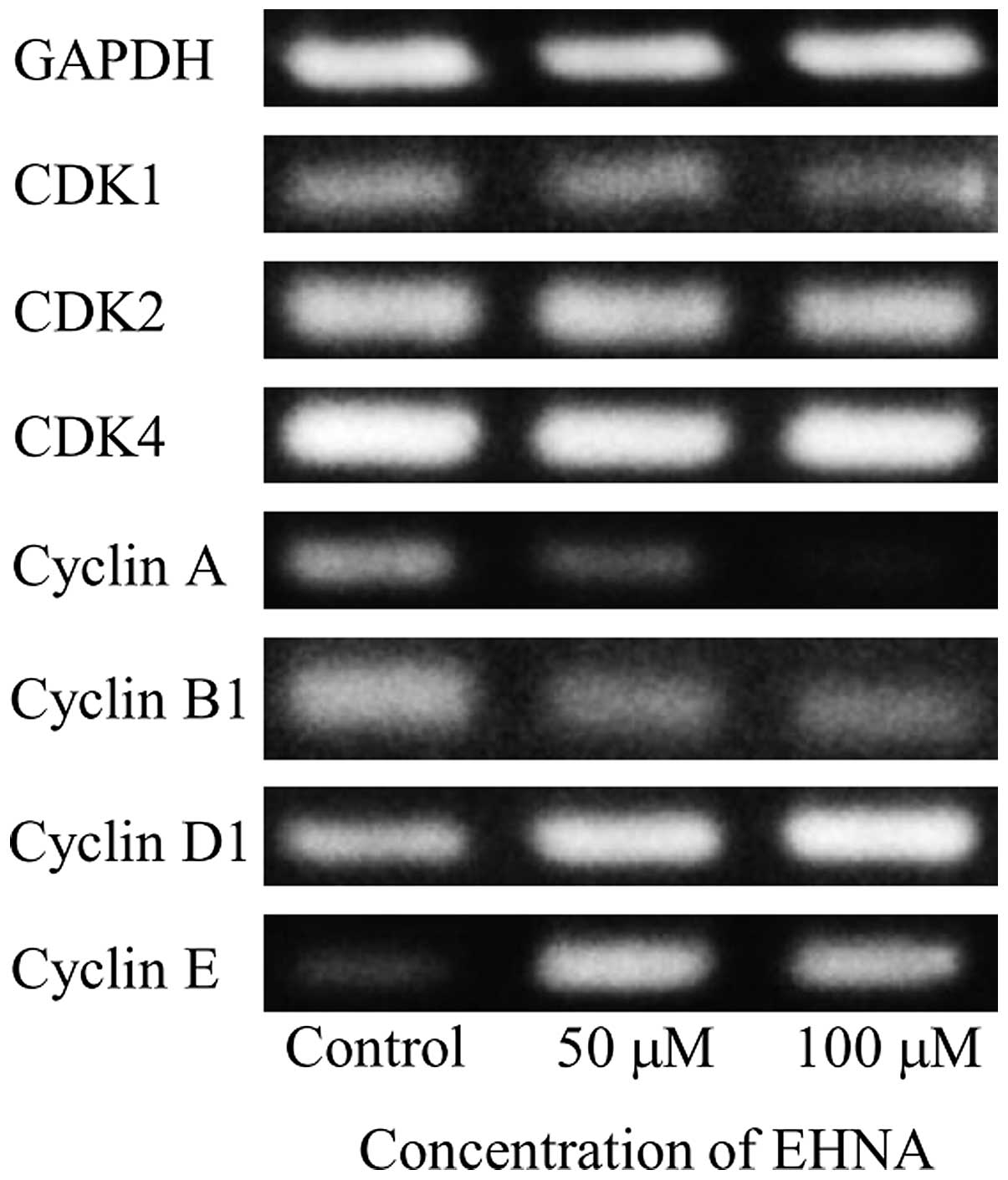
Effect of EHNA on the expression of CDKs
and cyclins in PMP cells
The effect of EHNA treatment on the expression of
CDKs and cyclins was examined by RT-PCR using eight primer sets
(Table III), which showed
downregulation of cyclin A and upregulation of cyclin E in PMP
cells treated with EHNA for 5 days (Fig. 9).
Discussion
Malignant melanoma is associated with a poor
prognosis, and there are currently no effective strategies for
treating the disease in the advanced stages (1). Thus, the development of new diagnostic
methods and treatments is necessary to improve prognosis. None of
the conventional antitumor agents, even those used as adjunctive
therapies to surgical treatment, improve the survival rates of
patients with malignant melanoma, and they are not effective for
controlling the disease after it metastasizes to other sites
(1). Understanding the mechanisms
underlying the growth, invasion and metastasis of malignant
melanoma may lead to the identification of molecular targets for
new diagnostic and therapeutic procedures.
PDEs hydrolyze cAMP and cGMP, regulating
intracellular signal transduction (2). The 11 PDE families contain several
variants that show distinct tissue distribution profiles and
subcellular localizations (11).
PDEs are considered potential drug targets and have been
intensively studied using pharmacological approaches (5). Highly selective inhibitors of several
PDE families have already been approved for clinical use, or are
being evaluated in clinical trials (2). The PDE3 inhibitor milrinone is used to
treat acute heart failure. Another PDE3 inhibitor, cilostazol
(Pletal®), is used to treat ischemic symptoms caused by
peripheral arterial occlusive disease and intermittent
claudication, and to prevent stroke recurrence (2,6,12). The
PDE4 inhibitors cilomilast (Ariflo®) and roflumilast
(Daxas®) are being evaluated clinically for the
treatment of chronic obstructive pulmonary disease (2,3,6).
Furthermore, the PDE5 inhibitors, sildenafil (Viagra™), vardenafil
(Levitra™) and tadalafil (Cialis™), are used for treating erectile
dysfunction (2,5,6).
The clinical applications of PDE inhibitors
described above were based on the roles of PDEs in normal cells.
Meanwhile, PDEs also play pivotal roles in malignant tumor cells.
PDE4 is involved in the regulation of apoptosis in chronic
lymphocytic leukemia (13). In
2004, we reported that PDE1 is a target molecule of
differentiation-inducing factor-1, which inhibits cell
proliferation and induces cell differentiation in Dictyostelium
discoideum, oocytes, mammalian cells and malignant tumor cells
(14,15). These findings suggest that PDEs are
strongly involved in the differentiation and growth of malignant
tumor cells, as well as normal cells, and that PDE signaling may
serve as a novel therapeutic target in malignant tumors.
Previously, we examined the expression of PDE1, PDE3
and PDE5 in malignant melanoma cells and demonstrated the
involvement of these isoforms in regulating cell proliferation
(7–9). In this study, we examined the role of
PDE2 in malignant melanoma and demonstrated that only one splice
variant, PDE2A2, is expressed in PMP cells. Furthermore, a specific
inhibitor of PDE2, EHNA, inhibited the growth of PMP cells in a
dose-dependent manner.
PDE2 activity and PDE2A mRNA expression in normal
and malignant tumor cells have been reported. PDE2 activity was
detected in the cytosolic fraction of normal human umbilical vein
endothelial cells (HUVECs), which was isolated by high-performance
liquid chromatography (16). PDE2
activity was also detected in malignant tumor cells including mouse
malignant melanoma B16 and human glioblastoma A172 cells (17,18).
PDE2A mRNA expression was confirmed in the human brain (19), heart (19), vascular endothelial cells (20), and human MG-63 osteosarcoma cells
(21), but has not been reported in
malignant melanoma cells. PDE2A is present at high levels in the
heart, liver, adrenal cortex, brain and epithelial cells (22), although the tissue distribution of
individual variants has not yet been reported (4), and differences in kinetic behavior
remain unclear. On the other hand, differential subcellular
localization of these variants, which may be caused by differences
in the N-terminal regions, has been documented (4). PDE2A2 and PDE2A3 are associated with
the membrane (4), and mouse PDE2A2
is present in mitochondria (23).
In the present study, the expression of PDE2A2 and a non-coding RNA
(GenBank NR_026572.2), of which the RT-PCR product was 62 bp longer
than that of PDE2A2, was detected in PMP cells. This was confirmed
by western blot analysis of PMP cell extracts, which showed an
approximately 105 kDa band corresponding to the PDE2A2 protein. The
size of this protein is comparable to that (102 kDa) of a protein
isolated from HUVECs (24). In
addition, we observed that inhibition of PDE2A2 by EHNA affected
the proliferation of PMP cells.
Although no abnormalities in the PDE2A gene have
been reported in humans, we found four point mutations, one of
which (nucleotide position 734) resulted in the substitution of
threonine with isoleucine at amino acid 214 in the N-terminal
domain (amino acids, 1–233) of PDE2A (GenBank NP_001137311.1),
which is adjacent to the GAF-A domain (amino acids, 234–380).
Whether this mutation is responsible for the effect of PDE2A2 on
cell proliferation is unclear. However, given the influence of the
N-terminal regions on the subcellular localization of PDE2A2
variants (4), it is possible that a
mutation in the N-terminus altered the subcellular distribution of
PDE2A2 in PMP cells. Consequently, intracellular signaling pathways
involving PDE2A2, and their effects on proliferation, may be
different in normal and PMP cells.
Cell motility is an important factor in the
mechanism of metastasis. Although the PDE2 inhibitor, EHNA,
suppressed vascular endothelial growth factor (VEGF)-induced HUVEC
migration (16), the effect of EHNA
treatment on PMP cell migration was negligible, suggesting that
PDE2A2 is not involved in the motility of malignant melanoma cells.
Further studies will reveal whether these discrepancies can be
explained by the changes in subcellular localization of PDE2A2
caused by the mutation in its N-terminal region.
In the present study, EHNA inhibited the
proliferation of PMP cells in a dose-dependent manner. The
cytotoxic effect of EHNA was assessed using the trypan blue
exclusion test. The BrdU cell proliferation assay results showed
suppression of DNA synthesis in EHNA-treated PMP cells, and this
was thought to be associated with the inhibition of cell growth.
Since the PDE2/5 inhibitor, exisulind, triggers apoptosis in human
colon carcinoma SW480 cells (25),
we investigated a possible similar effect of EHNA on PMP cells.
However, the results of TUNEL assays were similar in untreated and
EHNA-treated cells, indicating that PDE2A2 is not involved in
apoptotic pathways in PMP cells.
Our findings that EHNA suppressed DNA synthesis, but
did not induce apoptosis, in PMP cells suggests a possible effect
of the PDE2 inhibitor on cell cycle progression. In HUVECs, EHNA
altered EGF-induced cell cycle progression and reduced the
proportion of cells in the G2/M phase by 19% (16). The present flow cytometric results
showed that EHNA treatment increased the proportion of PMP cells in
the G2/M phase, indicating cell cycle arrest at either
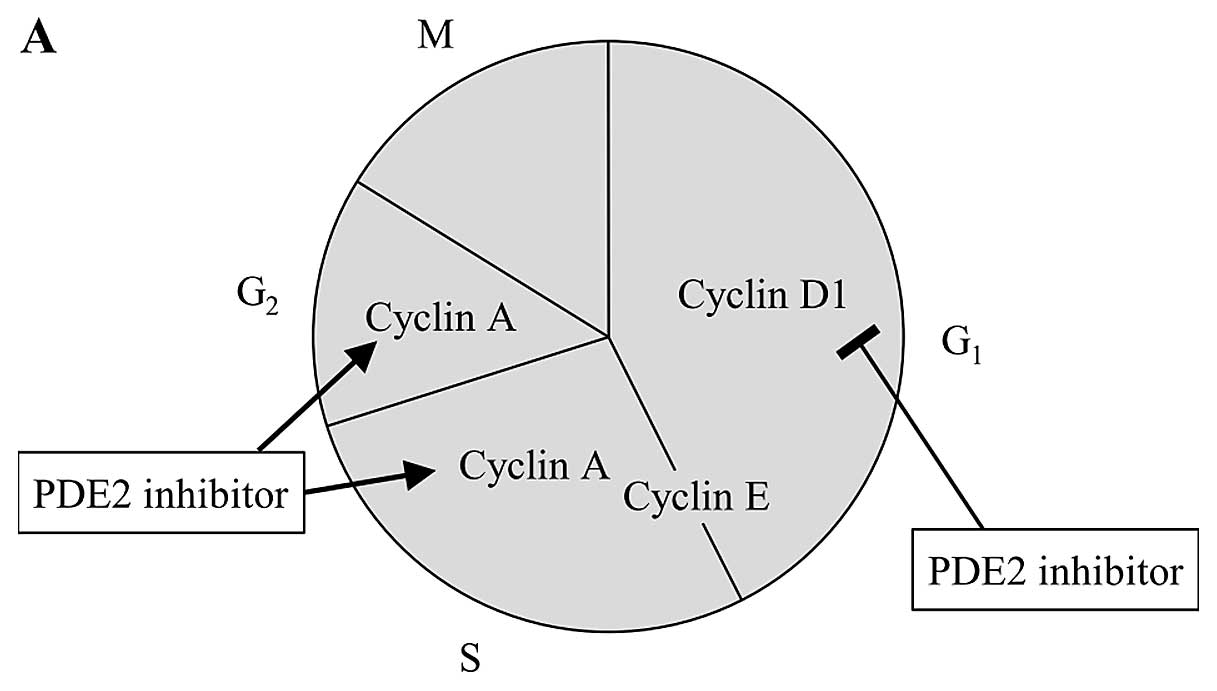
the G2 or M phase. Given the downregulation of cyclin A
and the upregulation of cyclin E by EHNA in PMP cells (Fig. 9), it is possible that the formation
of the cyclin A-CDK1 complex was inhibited, causing
G2-checkpoint arrest and suppression of spindle
formation in EHNA-treated PMP cells. Contrary to these changes in
PMP cells, upregulation of cyclin A and downregulation of cyclin D1
were previously reported in HUVECs (26). The different effects of EHNA on the
cell cycle progression in PMP cells and HUVECs are illustrated in
Fig. 10. However, the reasons for
these differences are unclear and further studies are necessary. In
general, cells are most radiosensitive in the G2/M phase
(27). In addition, the glutamate
release inhibitor riluzole inhibits metabotropic glutamate receptor
1 (GRM1), blocking the MAPK pathway, which is a crucial signaling
pathway controlling the pathogenesis of melanoma. Examination of
the effects of riluzole in melanoma cells showed an increase in the
proportion of cells in the G2/M phase and enhanced
ionizing radiation-induced cytotoxicity (28). Cyclooxygenase-2 (COX-2) and
phosphatidylinositol 3-kinase (PI3K)-AKT signaling are implicated
in the radioresistance of melanoma cells, and their respective
inhibitors, NS-398 and LY294002, increased the percentage of
G2/M-arrested cells and decreased clonogenic survival
after γ-irradiation of melanoma cells (29). Thus, it is possible that the
radiosensitivity of melanoma cells is high in the G2/M
phase, and if so, a PDE2 inhibitor in combination with radiotherapy
may improve the antitumor effect.
In conclusion, this study showed that PDE2A2, which
carries an amino acid mutation, is expressed in PMP cells and may
downregulate cyclin A, inducing G2/M arrest. Thus, the
development of drugs targeting PDE2A2 may be useful for the
treatment of malignant melanoma.
Abbreviations:
|
BrdU
|
5-bromo-2′-deoxyuridine
|
|
cAMP
|
adenosine 3′,5′-cyclic
monophosphate
|
|
cDNA
|
complementary deoxyribonucleic
acid
|
|
cGMP
|
guanosine 3′,5′-cyclic
monophosphate
|
|
DNA
|
deoxyribonucleic acid
|
|
EHNA
|
erythro-9-(2-hydroxy-3-nonyl)-adenine
|
|
FBS
|
fetal bovine serum
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
MTS
|
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
|
|
PBS
|
phosphate-buffered saline
|
|
PDE
|
phosphodiesterase
|
|
RNA
|
ribonucleic acid
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
|
SEM
|
standard error of the mean
|
|
TUNEL
|
terminal deoxynucleotidyl
transferase-mediated dUTP nick end-labeling
|
|
VEGF
|
vascular endothelial growth factor
|
References
|
1
|
Thompson JF, Scolyer RA and Kefford RF:
Cutaneous melanoma. Lancet. 365:687–701. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Beavo JA, Houslay MD and Francis SH:
Cyclic nucleotide phosphodiesterase superfamily. Cyclic Nucleotide
Phosphodiesterases in Health and Disease. Beavo JA, Francis SH and
Houslay MD: CRC Press; Boca Raton: pp. 3–17. 2007
|
|
3
|
Houslay MD: Underpinning compartmentalised
cAMP signalling through targeted cAMP breakdown. Trends Biochem
Sci. 35:91–100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Martinez SE: PDE2 structure and functions.
Cyclic Nucleotide Phosphodiesterases in Health and Disease. Beavo
JA, Francis SH and Houslay MD: CRC Press; Boca Raton: pp. 55–77.
2007
|
|
5
|
Conti M and Beavo J: Biochemistry and
physiology of cyclic nucleotide phosphodiesterases: essential
components in cyclic nucleotide signaling. Annu Rev Biochem.
76:481–511. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lugnier C: Cyclic nucleotide
phosphodiesterase (PDE) superfamily: a new target for the
development of specific therapeutic agents. Pharmacol Ther.
109:366–398. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shimizu K, Murata T, Watanabe Y, Sato C,
Morita H and Tagawa T: Characterization of phosphodiesterase 1 in
human malignant melanoma cell lines. Anticancer Res. 29:1119–1122.
2009.PubMed/NCBI
|
|
8
|
Murata T, Shimizu K, Narita M, Manganiello
VC and Tagawa T: Characterization of phosphodiesterase 3 in human
malignant melanoma cell line. Anticancer Res. 22:3171–3174.
2002.PubMed/NCBI
|
|
9
|
Murata T, Shimizu K, Watanabe Y, Morita H,
Sekida M and Tagawa T: Expression and role of phosphodiesterase 5
in human malignant melanoma cell line. Anticancer Res. 30:355–358.
2010.PubMed/NCBI
|
|
10
|
Kamei T, Inui M, Nakamura S, Okumura K,
Goto A and Tagawa T: Interferon-γ and anti-Fas antibody-induced
apoptosis in human melanoma cell lines and its relationship to
bcl-2 cleavage and bak expression. Melanoma Res. 13:153–159.
2003.
|
|
11
|
Omori K and Kotera J: Overview of PDEs and
their regulation. Circ Res. 100:309–327. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kambayashi J, Shakur Y and Liu Y: Bench to
bedside: multiple actions of the PDE3 inhibitor cilostazol. Cyclic
Nucleotide Phosphodiesterases in Health and Disease. Beavo JA,
Francis SH and Houslay MD: CRC Press; Boca Raton: pp. 627–648.
2007
|
|
13
|
Kim DH and Lerner A: Type 4 cyclic
adenosine monophosphate phosphodiesterase as a therapeutic target
in chronic lymphocytic leukemia. Blood. 92:2484–2494.
1998.PubMed/NCBI
|
|
14
|
Morris HR, Taylor GW, Masento MS, Jermyn
KA and Kay RR: Chemical structure of the morphogen differentiation
inducing factor from Dictyostelium discoideum. Nature.
328:811–814. 1987. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shimizu K, Murata T, Tagawa T, Takahashi
K, Ishizawa R, Abe Y, Hosaka K and Kubohara Y: Calmodulin-dependent
cyclic nucleotide phosphodiesterase (PDE1) is a pharmacological
target of differentiation-inducing factor-1, an antitumor agent
isolated from Dictyostelium. Cancer Res. 64:2568–2571. 2004.
View Article : Google Scholar
|
|
16
|
Favot L, Keravis T, Holl V, Le Bec A and
Lugnier C: VEGF-induced HUVEC migration and proliferation are
decreased by PDE2 and PDE4 inhibitors. Thromb Haemost. 90:334–343.
2003.PubMed/NCBI
|
|
17
|
Drees M, Zimmermann R and Eisenbrand G:
3′,5′-Cyclic nucleotide phosphodiesterase in tumor cells as
potential target for tumor growth inhibition. Cancer Res.
53:3058–3061. 1993.
|
|
18
|
Dunkern TR and Hatzelmann A:
Characterization of inhibitors of phosphodiesterase 1C on a human
cellular system. FEBS J. 274:4812–4824. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rosman GJ, Martins TJ, Sonnenburg WK,
Beavo JA, Ferguson K and Loughney K: Isolation and characterization
of human cDNAs encoding a cGMP-stimulated 3′,5′-cyclic nucleotide
phosphodiesterase. Gene. 191:89–95. 1997.PubMed/NCBI
|
|
20
|
Seybold J, Thomas D, Witzenrath M, Boral
S, Hocke AC, Bürger A, Hatzelmann A, Tenor H, Schudt C, Krüll M,
Schütte H, Hippenstiel S and Suttorp N: Tumor necrosis
factor-α-dependent expression of phosphodiesterase 2: role in
endothelial hyperpermeability. Blood. 105:3569–3576. 2005.
|
|
21
|
Ahlström M, Pekkinen M, Huttunen M and
Lamberg-Allardt C: Dexamathasone down-regulates
cAMP-phosphodiesterase in human osteosarcoma cells. Biochem
Pharmacol. 69:267–275. 2005.PubMed/NCBI
|
|
22
|
Gupta R, Kumar G and Kumar RS: An update
on cyclic nucleotide phosphodiesterase (PDE) inhibitors:
phosphodiesterases and drug selectivity. Methods Find Exp Clin
Pharmacol. 27:101–118. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Acin-Perez R, Russwurm M, Günnewig K,
Gertz M, Zoidl G, Ramos L, Buck J, Levin LR, Rassow J, Manfredi G
and Steegborn C: A phosphodiesterase 2A isoform localized to
mitochondria regulates respiration. J Biol Chem. 286:30423–30432.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Netherton SJ and Maurice DH: Vascular
endothelial cell cyclic nucleotide phosphodiesterases and regulated
cell migration: implications in angiogenesis. Mol Pharmacol.
67:263–272. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Thompson WJ, Piazza GA, Li H, Liu L,
Fetter J, Zhu B, Sperl G, Ahnen D and Pamukcu R: Exisulind
induction of apoptosis involves guanosin 3′,5′-cyclic monophosphate
phosphodiesterase inhibition, protein kinase G activation, and
attenuated β-catenin. Cancer Res. 60:3338–3342. 2000.PubMed/NCBI
|
|
26
|
Favot L, Keravis T and Lugnier C:
Modulation of VEGF-induced endothelial cell cycle protein
expression through cyclic AMP hydrolysis by PDE2 and PDE4. Thromb
Haemost. 92:634–645. 2004.PubMed/NCBI
|
|
27
|
Pawlik TM and Keyomarsi K: Role of cell
cycle in mediating sensitivity to radiotherapy. Int J Radiat Oncol
Biol Phys. 59:928–942. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khan AJ, Wall B, Ahlawat S, Green C,
Schiff D, Mehnert JM, Goydos JS, Chen S and Haffty BG: Riluzole
enhances ionizing radiation-induced cytotoxicity in human melanoma
cells that ectopically express metabotropic glutamate receptor 1 in
vitro and in vivo. Clin Cancer Res. 17:1807–1814. 2011. View Article : Google Scholar
|
|
29
|
Johnson GE, Ivanov VN and Hei TK:
Radiosensitization of melanoma cells through combined inhibition of
protein regulations of cell survival. Apoptosis. 13:790–802. 2008.
View Article : Google Scholar : PubMed/NCBI
|