Introduction
Prostate cancer is the most common cancer in males
in Europe and in the United States, and the third leading cause of
death from cancer in Europe (1),
resulting in 903,500 new cases and 258,400 deaths annually; 80% of
these men succumb to bone metastatic cancer (2). The multistep process of carcinogenesis
of prostate epithelial transformation is the transition from an
androgen-dependent non-metastatic phenotype to a more malignant
metastatic androgen-independent phenotype (3). As a consequence, nearly 30,000
individuals die of bone metastatic cancer every year in the US
alone (4). These figures
demonstrate the tragic contribution of bone metastatic cancer to
morbidity and mortality. Thus, novel therapies for the prevention
and treatment of bone metastatic cancers are imperative. Due to the
protracted natural progression of prostate cancer, clinical and
epidemiological studies alone may not provide the knowledge
necessary to design strategies for preventing, predicting and
treating metastatic disease (5).
Therefore, to gain further insight into human
prostate cancer progression and metastasis, well-defined in
vivo models that mimic the different aspects of the natural
course of disease progression are essential. However, the field of
prostate cancer research continues to be hindered by the lack of
relevant preclinical models with which to study tumorigenesis and
develop effective prevention and therapeutic interventions
(6). The most effective models are
those that can be used to mimic changes in human disease, that can
be utilized to ask questions that explain observed phenomena, and
that are predictive of therapeutic efficacy (7). Several mouse models have been
developed; these require injection or implantation of cancer cells
into various locations, such as subcutaneous tissue, intra-osseous
injection or injection into the left ventricle of the heart.
Additionally, carcinomas can be induced by either chemical or
physical agents (4,6,8–10).
Although valuable for studying the growth, physiology and
metastasis of metastatic processes in prostate cancer, these models
are inadequate to provide a full understanding of the disease
(11).
X-ray examination is the technique most commonly
used to detect bone metastases. Other techniques are also used to
characterize bone metastases in animal models, such as
microcomputed tomography, positron-emission tomography and
scintigraphy. However, cancer tends to be diagnosed at a later
stage, reducing access to suitable therapeutic facilities and drugs
(12). Therefore, more sensitive
methods are needed to detect cancer at the very early stages.
Bioluminescence imaging is an emerging method that allows
visualization of cancer of any stage, including growth of the
primary tumor, tumor cell motility and invasion, and evaluation of
the cytotoxicity of therapeutics. Bioluminescence imaging leads to
increased detection sensitivity and more accurate quantification of
metastases (13,14).
Since the model we describe here closely mimics the
natural history of human prostate cancer, it is useful for future
studies of the mechanisms and therapy of prostate tumor progression
and metastasis. In addition, animal models of metastasis have been
used in drug development (5,15).
Materials and methods
Tumor cell line
The LNCaP cell line was purchased from the Institute
of Biochemistry and Cell Biology, Shanghai Institutes for
Biological Sciences, Chinese Academy of Sciences, Shanghai, China.
Cells were routinely cultured in RPMI-1640 medium (Gibco, Grand
Island, NY, USA) supplemented with 20% F12K (Irvine Scientific,
Santa Ana, CA, USA), 100 μg/ml streptomycin, 100 U/l penicillin G
and 15% fetal bovine serum (FBS) (Gibco) in a humidified atmosphere
of 5% CO2 at 37°C. The cells were passaged weekly at a
1:3 dilution using 0.05% trypsin (Gibco).
Cell transfection
LNCaP cells were co-transfected with the
PGK-luciferase-green fluorescent protein (GFP) lentiviral vector
containing the PGK expression vector plasmid encoding luciferase
and GFP (Invitrogen, Carlsbad, CA, USA), using the LNCaP-luc
cationic liposome method. GFP expression was observed by
fluorescence microscopy after 48 h, and transfection efficiency was
monitored by flow cytometry.
Subcutaneous injection into nude
mice
The LNCaP-luc cell line was fed with fresh culture
medium 48 h before cell inoculation Cells were harvested and then
incubated in culture medium for 3 min and suspended in
phosphate-buffered saline (PBS) immediately before inoculation.
Cells (2×106) in 0.1 ml PBS mixed with 0.1 ml Matrigel
were injected subcutaneously (s.c.) into the groin of nude mice.
Tumor growth was monitored by external caliper measurements [tumor
size = (length × width × height) × 0.52].
Statement of ethics
Male BALB/c nude mice were purchased from Beijing
Weitong Lihua (Beijing, China). Experiments were carried out
according to the National Institutes of Health Guide for the Care
and Use of Laboratory Animals and were approved by the Bioethics
Committee of Sun Yat-sen University. Mice were maintained in
laminar flow boxes under sterile conditions. Sterile water and food
were provided. Mice were 4–5 weeks old when used for intracardiac
injections of tumor cells and 5–6 weeks old when injected with
tumor cells.
Generation of the LNCaP1-luc cell
line
Animals were sacrificed when subcutaneous tumors
grew to nearly 1 cm in diameter. Tumor tissues were harvested from
the subcutaneous tissue under aseptic conditions. Briefly, tumors
were minced into 1-mm3 cubes, placed on small culture
dishes, and immersed in tissue-culture medium. Tumor cells were
outgrown together with host fibroblasts within 1.5 weeks. LNCaP-luc
tumor cells, which adhered loosely to the plastic dishes, were
enriched by washing the culture dishes with PBS. The pure LNCaP
cell line (LNCaP1-luc) was obtained after 5–10 of these
subculturing steps.
CCK-8 assay
LNCaP-luc and LNCaP1-luc cells were digested and
dispensed into 96-well plates at a density of 3×103/well
(100 μl/well) for the cell viability assay. After a 42-h
incubation, 10 μl of CCK-8 was added to each well for an additional
4 h at 37°C. Plates were then shaken for 10 min, and the optical
density values at 450 nm were measured according to the
manufacturer’s instructions. All experiments were carried out in
triplicate to ascertain reproducibility. Growth curves were
generated from these data. The control group contained only
cell-culture medium.
Transwell migration assay
Transwell migration assays were carried out using
24-well cell-culture plate inserts and Transwell filters with 8-μm
pores. Cells were cultured for 24 h in serum-free RPMI-1640 medium
and then seeded (1.5×105) into the inner chamber with
300 μl serum-free RPMI-1640 and 500 μl of RPMI-1640 containing 10%
FBS in the outer chamber. Cells were allowed to migrate for 24 h at
37°C in 5% CO2. The cells were stained with hematoxylin
and washed twice in PBS. Non-migratory cells on the upper surface
of the inner chamber were removed using cotton swabs. Cells in five
random high-power microscopic fields (x100) per filter were
counted.
Western blot analysis
Total protein of the LNCaP-luc and LNCaP1-luc cells
was extracted using a cell lysis buffer containing 20 mM Tris-HCl
(pH 7.6), 1% NP-40, 3 mM EGTA, 3 mM EDTA, 0.15 mM NaCl, 1 mM
phenylmethylsulfonyl fluoride, 5 mg/ml leupeptin and 20 mg/ml
aprotinin. Homogenates were centrifuged at 12,000 rpm for 20 min at
4°C and the supernatants were retained. The protein concentration
was determined using a Bio-Rad DC Protein Assay (Bio-Rad
Laboratories, Hercules, CA, USA). Equal amounts of the protein (20
μg) were electrophoresed on 10% sodium dodecyl sulfate
polyacrylamide gels and transferred electrophoretically to
nitrocellulose membranes (Bio-Rad Laboratories). Following blocking
with 5% non-fat milk in wash buffer at room temperature for 2 h,
the membranes were incubated with the primary vimentin, E-cadherin
and GAPDH antibodies (1:1,000; Santa Cruz Biotechnology, Santa
Cruz, CA, USA) at 4°C overnight. The membranes were then incubated
for 1 h at room temperature with a horseradish
peroxidase-conjugated secondary antibody at a 1:5,000 dilution. The
membranes were investigated using Western Blotting Plus
Chemiluminescence Reagent (Thermo Fisher Scientific, Rockland, IL,
USA) and fluorography using X-ray film (Fuji, Tokyo, Japan).
Real-time reverse
transcription-polymerase chain reaction (PCR) analysis
Total RNA was extracted from LNCaP-luc and
LNCaP1-luc cells with the E.Z.N.A.® HP Total RNA kit
(Omega Bio-Tek, Norcross, GA, USA). cDNA was synthesized using the
PrimeScript® RT reagent kit (Takara Bio, Shiga, Japan).
After mixing the resulting complementary DNA template with
vimentin, E-cadherin, Snail, Slug, or GAPDH primers, respectively,
and Takara SYBR® Premix Ex Taq™, the quantitative
real-time PCR reaction was performed on a LightCycler 480 Real-Time
PCR system (Roche). Quantified expression values for individual
genes were normalized to that of GAPDH. Changes in mRNA expression
were expressed as fold-changes relative to the control. The
gene-specific primers used in this study were as follows. Primers
for Vimentin, E-cadherin, Snail and Slug were as previously
published (16). GAPDH sense was
5′-CACCCAGAAGACTGTGGATGG-3′ and GAPDH antisense was
5′-GTCTACATGGCAACTGTGAGG-3′.
Intracardiac injection
Nude mice were anesthetized with ketamine, and
LNCaP1-luc cells (0.5×106) suspended in 0.1 ml PBS were
injected into the left cardiac ventricle following a modification
of the technique first described by Arguello et al(11). Successful injection was indicated
retrospectively by pulsatile bright red blood in the 29G needle
fitted with a 1.0-ml insulin syringe. Successful injection was also
confirmed by the lack of abnormal tumor mass growth in the pleural
cavity 3–4 weeks after injection.
Bioluminescence imaging
Mice were imaged for bioluminescence weekly for 4–6
weeks after intracardiac injection using the IVIS®
Imaging system from Caliper (Alameda, CA, USA). Images were
acquired and processed using the Living Image® version
4.2 software. Anesthesia was induced with inhaled isoflurane and
maintained with 2% isoflurane mixed with oxygen/nitrogen via a nose
cone. D-luciferin (3 mg dissolved in water) was administered at 150
mg/kg in DPBS via intraperitoneal injection. Optical imaging was
carried out ~10–15 min later. One to three mice were imaged at a
time, and the tails, feet and paws were taped to prevent movement
artifacts. One-minute acquisitions were conducted at the
high-sensitivity setting of the instrument software. Background was
subtracted using an automated feature of the program.
Histology and immunostaining for the
androgen receptor (AR)
Mouse skeletons were fixed in 10% formalin for 24 h
and then decalcified in 10% nitric acid overnight; other tissues
were harvested and fixed in 10% formalin (Sigma, St. Louis, MO,
USA), and all tissues were embedded in paraffin, sectioned, and
stained with hematoxylin and eosin (H&E). The H&E-stained
sections were observed by microscopy, and images were obtained
using a SPOT Insight color camera (Nikon 80i, Tokyo, Japan).
AR (BioGenex F 39.4.1) immunostaining was performed
using the two-step EnVision immunohistochemical procedure (Dako,
Glostrup, Denmark) as previously described (17). A monoclonal antibody against AR,
which is present in prostate cancer, was used. Signals were
visualized with DAB, and the slides were counterstained with
Mayer’s hematoxylin. AR-positive tissues were used as positive
controls. PBS without the primary antibody was used as the negative
control. Tumors were considered positive when ≥10% of the
neoplastic cells were stained. The results of all specimens were
scored by two independent observers.
Statistical analysis
Statistical differences and variances were evaluated
with the t-test and variance analysis using statistical SPSS (SPSS,
Inc., Chicago, IL, USA) software. Differences were considered
significant at P<0.05.
Results
Transfection efficiency of the LNCaP
cells
After parental LNCaP cells were transfected with the
lentiviral vector (PGK-luciferase-GFP), we visualized the expressed
GFP at 24 h (the fluorescence intensity increased gradually with
increasing passages). Transfection efficiency was determined by
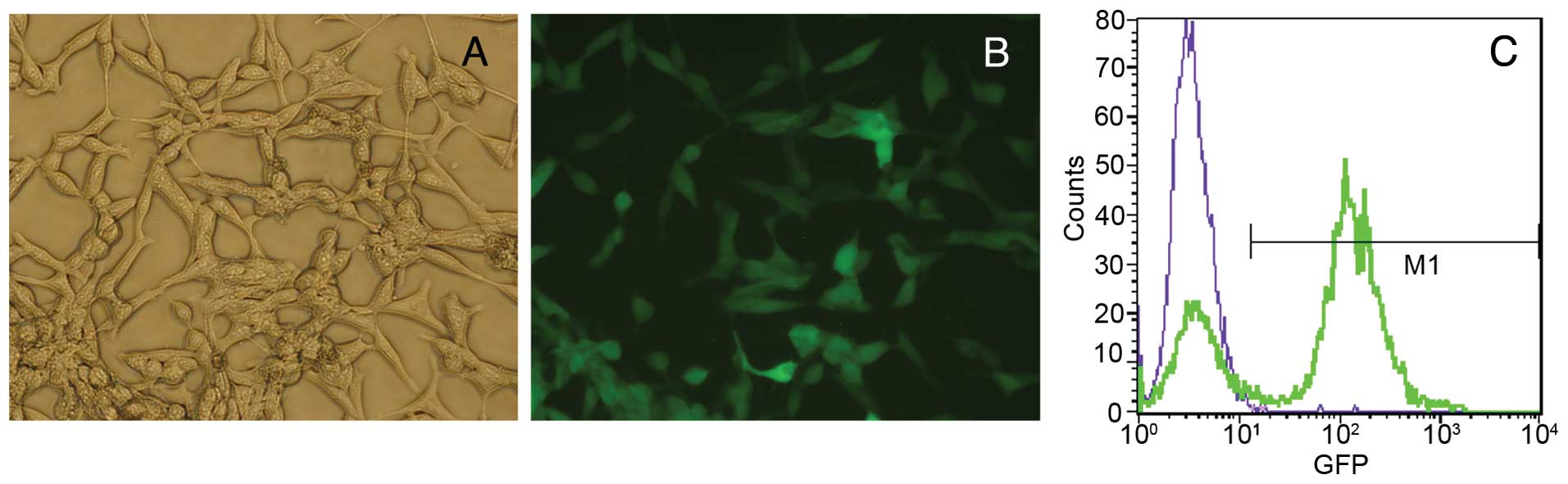
flow cytometry. Fig. 1A and B shows
the proportion of cells expressing GFP and the fluorescence
intensity. LNCaP-luc cells were polygonal in shape, and the
fluorescence intensity was unequal. Flow cytometry detected GFP
expression (Fig. 1C), and the
transfection efficiency approached 80%.
Characteristics of the LNCaP cell
subline
Incidence of tumor formation
Table I summarizes
the incidence of tumor formation in the nude mice inoculated s.c.
with 2×106 LNCaP-luc cells and those inoculated with
0.5×106 LNCaP1-luc cells into the left ventricle of the
heart. The development of primary tumors (8 of 20 for the LNCaP-luc
cells, 16 of 20 for the LNCaP1-luc cells, respectively) by s.c
injection and (2 of 20 for the LNCaP-luc cells, 9 of 20 for the
LNCaP1-luc cells, respectively) by intracardiac injection was
observed. When the LNCaP cell line was injected s.c. into nude
mice, it induced tumors only at the site of injection; no
metastases were detected. The pattern of the incidence of tumor
formation differed between mice given s.c. injections and those
that received intracardiac injection of LNCaP-luc or LNCaP1-luc
cells. Approximately 2-fold more mice injected with LNCaP1-luc
cells developed tumors compared with mice injected with LNCaP-luc
cells. Additionally, the tumor formation rate following
intracardiac injection of LNCaP1-luc cells was higher than that
after injection with LNCaP-luc cells. Thus, LNCaP1-luc cells
injected into the heart had significantly higher tumorigenicity
than did LNCaP-luc cells injected s.c.
 | Table ITumorigenicity and metastatic
behavior of the LNCaP-luc and LNCaP1-luc cells injected either
subcutaneously (s.c.) or into the left cardiac ventricle of nude
mice. |
Table I
Tumorigenicity and metastatic
behavior of the LNCaP-luc and LNCaP1-luc cells injected either
subcutaneously (s.c.) or into the left cardiac ventricle of nude
mice.
| Cell line | Mouse strain | Injection | Tumor formation
(%) | End-point
(days) |
|---|
| LNCaP-luc | Nude | s.c. | 8/20 (40) | 45 |
| | Intracardiac | 2/20 (10) | 90 |
| LNCaP1-luc | Nude | s.c. | 16/20 (80) | 45 |
| | Intracardiac | 9/20 (45) | 90 |
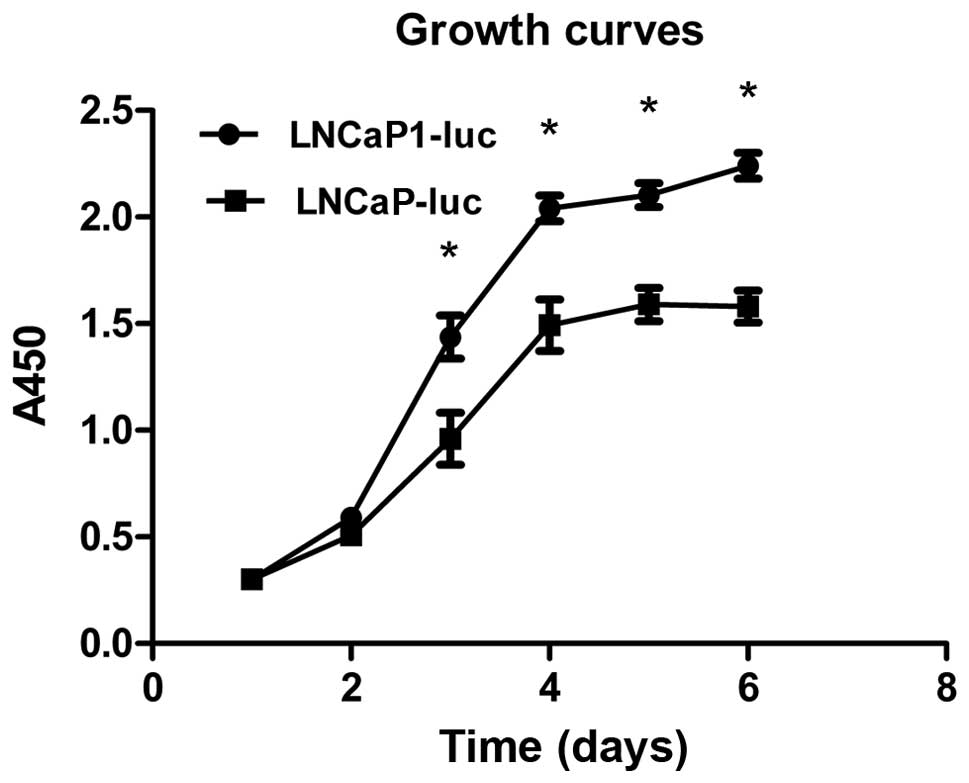
Enhanced cell viability and growth of
LNCaP1-luc cells compared to LNCaP-luc cells
Cell viability and growth kinetics were measured by
the CCK-8 assay. Growth curves of the two LNCaP cell line
populations were generated to compare the growth characteristics
after passage at various times. LNCaP1-luc cells grew more rapidly
than did the LNCaP-luc cells (Fig.
2), indicating that LNCaP1-luc cells proliferated more rapidly.
Additionally, the growth ability of the LNCaP1-luc cells increased
gradually with the number of the passage (data not shown).
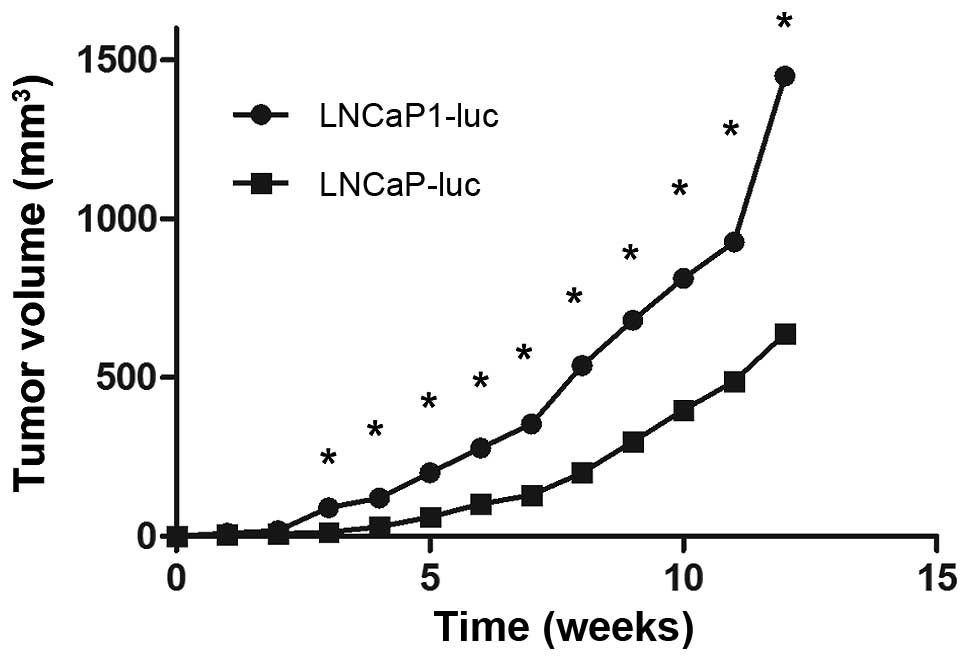
Enhanced tumorigenicity of LNCaP1-luc
cells compared to the LNCaP-luc cell line
The prostate tumor growth rates of the two
luciferase-expressing derivatives are shown in Fig. 3. External caliper measurements
indicated that LNCaP1-luc cells grew at a faster rate than did the
LNCaP-luc cells after s.c injection. After 5 weeks of growth in
nude mice, the LNCaP1-luc tumors had reached a volume more than
twice that of tumors derived from the LNCaP-luc cell line,
suggesting that the LNCaP1-luc cells had a higher
tumorigenicity.
Increased invasiveness and induced
epithelial-mesenchymal transition (EMT) in the LNCaP1-luc cell
line
We found that the LNCaP1-luc cell line exhibited
increased invasiveness when compared to the LNCaP-luc cell line;
the relative percentage of LNCaP1-luc cells that migrated through
the filters compared to LNCaP-luc cells was 276.2±26.4% as observed
by Transwell migration assay (Fig.
4A). mRNA and protein expression of the epithelial cell marker
E-cadherin was significantly lower, whereas mRNA and protein
expression of the mesenchymal cell marker vimentin was higher in
the LNCaP1-luc cells when compared to the LNCaP-luc cells (Fig. 4B and C). This may explain why the
LNCaP1-luc cell line exhibited increased invasiveness when compared
to the LNCaP-luc cell line. The Snail mRNA level was higher in the
LNCaP1-luc cell line, which is crucial for the EMT process, whereas
Slug mRNA expression did not differ between the LNCaP1-luc and the
LNCaP-luc cells.
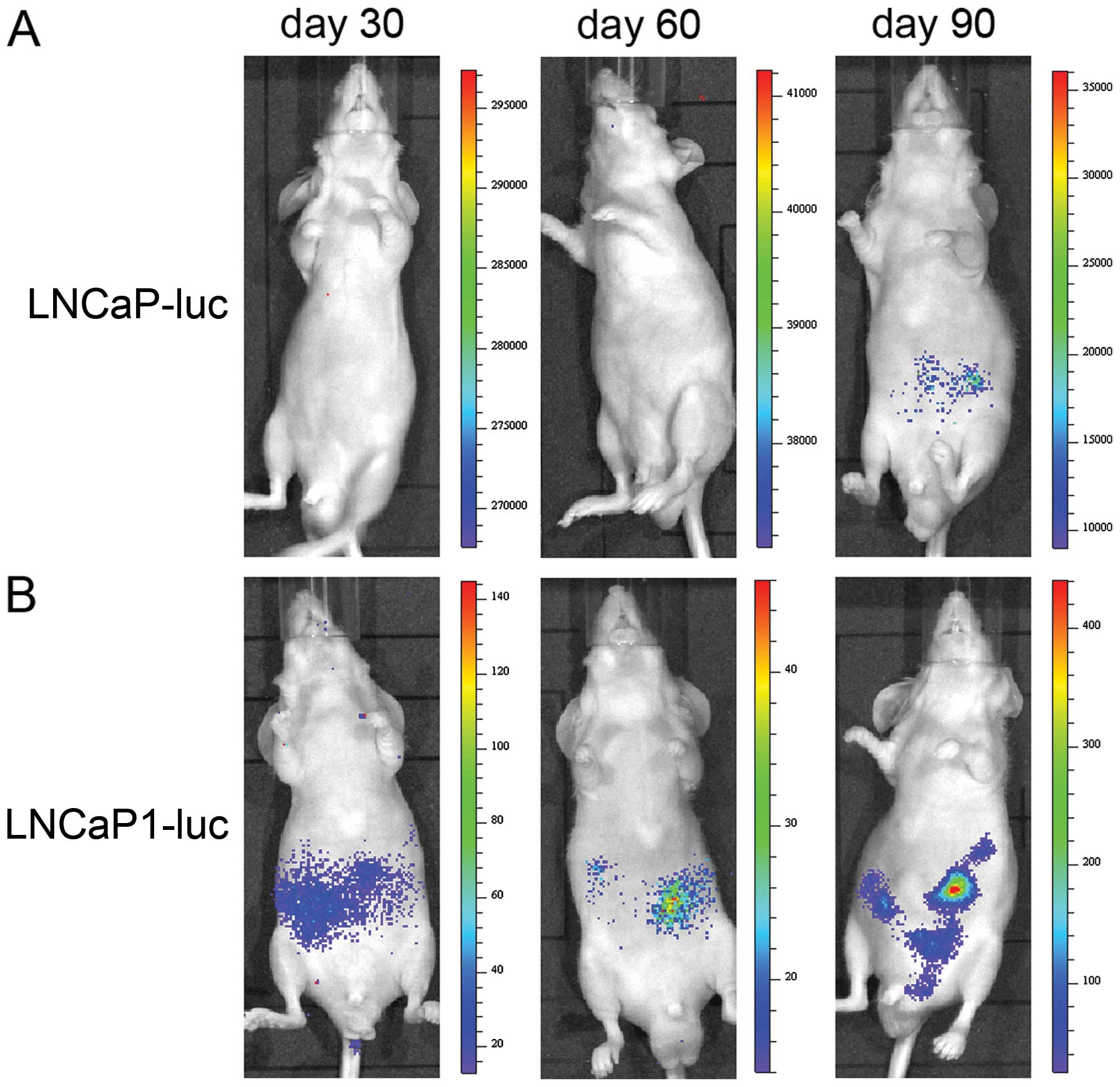
Bioluminescence imaging after
intracardiac injection
We injected LNCaP-luc or LNCaP1-luc cells into the
left cardiac ventricle of male nude mice at a rate of
0.5×106 cells/mouse. Since the LNCaP cell line was
labeled with luciferase and GFP, we were able to observe metastatic
tumors by bioluminescence imaging. The mice were imaged weekly
beginning 4 weeks after the intracardiac injection. The whole mouse
image in Fig. 5 shows the results
of the injection of LNCaP-luc and LNCaP1-luc cells. A small hot
spot of bioluminescence, which increased with time, was observed
following LNCaP-luc cell injection (Fig. 5A). Fig.
5B shows the localized and distant metastasized cancer in the
LNCaP1-luc heart-injected animal model; transformation of a
hot-spot of the bioluminescence signal in the lower abdomen was
widely distributed over time. Thus, the LNCaP1-luc cell line
induced multiple metastases after intracardiac inoculation. In
contrast, the inoculation of LNCaP-luc cells rarely induced such
metastases. This result suggests that LNCaP1-luc cells have a
higher metastatic potential than their parental LNCaP-luc
cells.
Histology and immunohistochemistry of
mouse tissues
Mouse tissues were also evaluated by histological
and immunohistochemical analyses. Mice were decalcified and
processed for conventional histological and immunohistochemistry
examination 90 days after injection. Representative histological
sections of the lung (A), liver (B), skeletal (C), kidney (D),
heart tissues (E), and lymph nodes (F) and metastases are shown in
Fig. 6. Tumor cells had a nucleus
larger (stained blue) than that in the normal cells. The lung
lobes, kidneys, and hepatic lobules were full of tumor cells.
Additionally, slight tumor growth was observed along the needle
tract in the heart (arrow). The groin and para-vertebral lymph
nodes were nearly infiltrated by tumor cells, and the normal
structure of the lymph nodes could not be seen. The prostatic
origin of the epithelial cells derived from the s.c. tumors in the
lymph nodes of male nude mice was confirmed to be positive for AR,
but the staining was scattered (Fig. 6G
and H); all other metastatic sublines showed similar
immunostaining (data not shown). The results of the
immunohistochemical analysis suggest that all sublines were human
and shared a common AR marker with the parental LNCaP cells.
Discussion
Experimental animal models of human cancers are
required to elucidate the molecular events associated with
prostatic carcinogenesis and progression. In vitro testing
provides a significant amount of information on cellular
characterization but is inadequate to provide a full understanding
of bone metastatic processes. As we know, at present few similar
animal models are available for studying the progression of human
prostate cancer from the primary tumor to metastasis. We described
the establishment of such an animal model in this study. We
generated LNCaP1-luc cells which showed multiple metastases in mice
when injected into the left cardiac muscle. Notably, the
characteristics of LNCaP1-luc cells differed from those of the
parental cells, and showed increased cell proliferation, cell
invasiveness, tumorigenicity, and metastastic potential and
underwent EMT.
Human prostate cancer is initially androgen
dependent, but it acquires the ability to grow in the absence of
androgens after androgen-ablation therapy. The AR is expressed in
the majority of primary and metastatic prostate cancers (19). LNCaP cells are an
androgen-responsive human prostate cancer cell line derived
originally from a lymph node with prostate cancer metastasis
(20). LNCaP cells express the AR,
and their growth is stimulated by androgens (21). Many studies have suggested that the
AR gene is amplified in bone metastases from an androgen-dependent
state to hormone-refractory prostate cancer and that it requires
active AR signaling during prostate cancer progression (22). Not many models have been described
for the LNCaP human prostate cancer cell line in nude mice. We
injected LNCaP1-luc cells directly into the left heart ventricle of
nude mice, which resulted in successful development of a multiple
metastatic animal model. Moreover, the AR was detected in LNCaP
cells (Fig. 6G and H) and may
provide a foundation for further study of AR-targeted anti-cancer
therapy drugs (23–26).
The LNCaP1-luc cell subline metastasized to the
lymph nodes, bones, lungs and liver with an incidence of 10–45%
(Table I). This tumor formation
rate is higher than that previously reported (2,27).
There are several possible explanations for, and potential
implications of, these findings. First, the characteristics of the
LNCaP sublines differed from those of the primary line. LNCaP1-luc
cells were obtained using the LNCaP-luc prostate carcinoma cell
line following repeated subcutaneous injections with Matrigel in
nude mice, which increased cell tumorigenicity as mentioned in
numerous studies (28,29), followed by isolation of the cells
from tumors. Subcutaneous tumor volume measured using external
calipers indicated that LNCaP1-luc cells grew more rapidly than did
LNCaP-luc cells (Fig. 2) (30). The CCK-8 assay results also showed
that the growth ability of LNCaP1-luc cells was enhanced when
compared with LNCaP-luc cells (Fig.
3). Thus, the LNCaP1-luc cells exhibited a higher tumor
formation rate than did LNCaP-luc cells. Second, E-cadherin
expression was used to monitor the epithelial phenotype, and its
loss suggests EMT (16). The EMT
triggers tumor metastasis and enhances metastatic potential
(31,32). The results of the western blot
analysis and PCR suggested that LNCaP1-luc cells may have induced
LNCaP-luc cells to undergo EMT through the Snail transcription
factor. This result is consistent with many other studies (33,34).
At the same time, the Transwell assay results illustrated that
LNCaP1-luc cells were more invasive than LNCaP-luc cells.
Therefore, LNCaP1-luc cells may have greater potential for bone
metastasis. Indeed, our results support a number of reports of the
importance of EMT in cancer metastases and the mechanisms involved
(33,35–39).
These studies demonstrated that prostate cells undergoing EMT
become more invasive and express several genes associated with
metastasis. Third, the technique most widely used for detection of
metastasis is radiological examination. However, it should be noted
that micrometastases elude radiographic detection, and cancer tends
to be diagnosed at later stages, reducing access to appropriate
therapeutic facilities and drugs and adversely affecting survival.
In this study, we used bioluminescence imaging to visualize cancer
growth, tumor cell motility, and invasion. Wetterwald et
al(14) reported that
whole-body bioluminescence reporter imaging detects microscopic
bone marrow metastases with volumes of ~0.5 mm3 when
used with luciferase-transfected cells. This sensitive technology
for early detection of tumor growth is preceded by the appearance
of a radiologically evident metastasis in ~2 weeks. Thus, applying
bioluminescence imaging allows detection of micrometastases at the
very early stage, which improves the detection rate. In summary,
the LNCaP1-luc cell line exhibited high metastatic potential which
facilitated the generation of a metastatic animal model of prostate
cancer. We obtained the following evidence for metastasis: i)
monitoring of tumor growth and multiple metastases over time in
vivo by bioluminescence imaging; and ii) histological and
immunohistochemical data.
In short, our results have various differences
compared with other previously reported results. The use of this
method yielded a cell line with greater invasive potential. Thus,
using these LNCaP1-luc cells can generate an animal model of
multiple metastatic prostate cancers.
Acknowledgements
This work was supported by Science and Technology
Planning Project of Guangdong Province (No. 2010B060500005),
Guangdong Key Natural Science Foundation (No. 8251008901000009),
and National Natural Science Foundation of China (No.
30970857).
References
|
1
|
Marech I, Vacca A, Ranieri G, Gnoni A and
Dammacco F: Novel strategies in the treatment of
castration-resistant prostate cancer (Review). Int J Oncol.
40:1313–1320. 2012.PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
3
|
Wang Q, Li W, Zhang Y, et al: Androgen
receptor regulates a distinct transcription program in
androgen-independent prostate cancer. Cell. 138:245–256. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Power CA, Pwint H, Chan J, et al: A novel
model of bone-metastatic prostate cancer in immunocompetent mice.
Prostate. 69:1613–1623. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zuluaga MF, Gabriel D and Lange N:
Enhanced prostate cancer targeting by modified protease sensitive
photosensitizer prodrugs. Mol Pharm. 9:1570–1579. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pienta KJ, Abate-Shen C, Agus DB, et al:
The current state of preclinical prostate cancer animal models.
Prostate. 68:629–639. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rosol TJ, Tannehill-Gregg SH, LeRoy BE,
Mandl S and Contag CH: Animal models of bone metastasis. Cancer.
97:748–757. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Blouin S, Basle MF and Chappard D: Rat
models of bone metastases. Clin Exp Metastasis. 22:605–614. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Singh AS, Macpherson GR, Price DK, Schimel
D and Figg WD: Evaluation of human fetal bone implants in SCID mice
as a model of prostate cancer bone metastasis. Oncol Rep.
15:519–524. 2006.PubMed/NCBI
|
|
10
|
Song H, Shahverdi K, Huso DL, et al: An
immunotolerant HER-2/neu transgenic mouse model of metastatic
breast cancer. Clin Cancer Res. 14:6116–6124. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Arguello F, Baggs RB and Frantz CN: A
murine model of experimental metastasis to bone and bone marrow.
Cancer Res. 48:6876–6881. 1988.PubMed/NCBI
|
|
12
|
Jenkins DE, Hornig YS, Oei Y, Dusich J and
Purchio T: Bioluminescent human breast cancer cell lines that
permit rapid and sensitive in vivo detection of mammary tumors and
multiple metastases in immune- deficient mice. Breast Cancer Res.
7:R444–R454. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fritz V, Louis-Plence P, Apparailly F, et
al: Micro-CT combined with bioluminescence imaging: a dynamic
approach to detect early tumor-bone interaction in a tumor
osteolysis murine model. Bone. 40:1032–1040. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wetterwald A, van der Pluijm G, Que I, et
al: Optical imaging of cancer metastasis to bone marrow: a mouse
model of minimal residual disease. Am J Pathol. 160:1143–1153.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xing N, Chen Y, Mitchell SH and Young CY:
Quercetin inhibits the expression and function of the androgen
receptor in LNCaP prostate cancer cells. Carcinogenesis.
22:409–414. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Emadi BM, Soheili ZS, Schmitz I, Sameie S
and Schulz WA: Snail regulates cell survival and inhibits cellular
senescence in human metastatic prostate cancer cell lines. Cell
Biol Toxicol. 26:553–567. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen JN, Jiang Y, Li HG, et al:
Epstein-Barr virus genome polymorphisms of Epstein-Barr
virus-associated gastric carcinoma in gastric remnant carcinoma in
Guangzhou, Southern China, an endemic area of nasopharyngeal
carcinoma. Virus Res. 160:191–199. 2011. View Article : Google Scholar
|
|
18
|
Yang M, Jiang P, Sun FX, et al: A
fluorescent orthotopic bone metastasis model of human prostate
cancer. Cancer Res. 59:781–786. 1999.PubMed/NCBI
|
|
19
|
Guo Z, Yang X, Sun F, et al: A novel
androgen receptor splice variant is up-regulated during prostate
cancer progression and promotes androgen depletion-resistant
growth. Cancer Res. 69:2305–2313. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu HC, Hsieh JT, Gleave ME, Brown NM,
Pathak S and Chung LW: Derivation of androgen-independent human
LNCaP prostatic cancer cell sublines: role of bone stromal cells.
Int J Cancer. 57:406–412. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hobisch A, Culig Z, Radmayr C, Bartsch G,
Klocker H and Hittmair A: Distant metastases from prostatic
carcinoma express androgen receptor protein. Cancer Res.
55:3068–3072. 1995.PubMed/NCBI
|
|
22
|
Brown RS, Edwards J, Dogan A, et al:
Amplification of the androgen receptor gene in bone metastases from
hormone-refractory prostate cancer. J Pathol. 198:237–244. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ullen A, Farnebo M, Thyrell L, et al:
Sorafenib induces apoptosis and autophagy in prostate cancer cells
in vitro. Int J Oncol. 37:15–20. 2010. View Article : Google Scholar
|
|
24
|
Liu Y, Wu X, Dong Z and Lu S: The
molecular mechanism of Vav3 oncogene on upregulation of androgen
receptor activity in prostate cancer cells. Int J Oncol.
36:623–633. 2010.PubMed/NCBI
|
|
25
|
Martin TA and Jiang WG: Anti-cancer agents
in medicinal chemistry (Formerly current medicinal chemistry -
Anti-cancer agents). Anticancer Agents Med Chem.
10:12010.PubMed/NCBI
|
|
26
|
Sturge J, Caley MP and Waxman J: Bone
metastasis in prostate cancer: emerging therapeutic strategies. Nat
Rev Clin Oncol. 8:357–368. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu TT, Sikes RA, Cui Q, et al:
Establishing human prostate cancer cell xenografts in bone:
induction of osteoblastic reaction by prostate-specific
antigen-producing tumors in athymic and SCID/bg mice using LNCaP
and lineage-derived metastatic sublines. Int J Cancer. 77:887–894.
1998. View Article : Google Scholar
|
|
28
|
Kleinman HK and Martin GR: Matrigel:
basement membrane matrix with biological activity. Semin Cancer
Biol. 15:378–386. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tilghman RW, Blais EM, Cowan CR, et al:
Matrix rigidity regulates cancer cell growth by modulating cellular
metabolism and protein synthesis. PLoS One. 7:e372312012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Thalmann GN, Anezinis PE, Chang SM, et al:
Androgen-independent cancer progression and bone metastasis in the
LNCaP model of human prostate cancer. Cancer Res. 54:2577–2581.
1994.PubMed/NCBI
|
|
31
|
Xu J, Wang R, Xie ZH, et al: Prostate
cancer metastasis: role of the host microenvironment in promoting
epithelial to mesenchymal transition and increased bone and adrenal
gland metastasis. Prostate. 66:1664–1673. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: at the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kogan-Sakin I, Tabach Y, Buganim Y, et al:
Mutant p53(R175H) upregulates Twist1 expression and promotes
epithelial-mesenchymal transition in immortalized prostate cells.
Cell Death Differ. 18:271–281. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kong D, Banerjee S, Ahmad A, et al:
Epithelial to mesenchymal transition is mechanistically linked with
stem cell signatures in prostate cancer cells. PLoS One.
5:e124452010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Birchmeier C, Birchmeier W and
Brand-Saberi B: Epithelial-mesenchymal transitions in cancer
progression. Acta Anat. 156:217–226. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Grille SJ, Bellacosa A, Upson J, et al:
The protein kinase Akt induces epithelial mesenchymal transition
and promotes enhanced motility and invasiveness of squamous cell
carcinoma lines. Cancer Res. 63:2172–2178. 2003.PubMed/NCBI
|
|
37
|
Guarino M, Rubino B and Ballabio G: The
role of epithelial-mesenchymal transition in cancer pathology.
Pathology. 39:305–318. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Larue L and Bellacosa A:
Epithelial-mesenchymal transition in development and cancer: role
of phosphatidylinositol 3′ kinase/AKT pathways. Oncogene.
24:7443–7454. 2005.
|
|
39
|
Wells A, Yates C and Shepard CR:
E-cadherin as an indicator of mesenchymal to epithelial reverting
transitions during the metastatic seeding of disseminated
carcinomas. Clin Exp Metastasis. 25:621–628. 2008. View Article : Google Scholar : PubMed/NCBI
|