Introduction
Breast cancer is the most frequently diagnosed
cancer and the leading cause of cancer-related mortality in females
(1). With the development of
gene-expression profiling, researchers have found that breast
cancer can be divided into different molecular subtypes with
distinct clinical features and varying responses to therapeutic
regimens (2–4). Patients with luminal A-type tumors may
gain little benefit from chemotherapy. For those patients, hormone
therapy may be sufficient for systemic treatment (5). However, chemotherapy remains the
standard systemic treatment for other types of cancer, particularly
human epidermal growth factor receptor 2 (HER-2)-positive tumors
and basal-like tumors. Paclitaxel-containing drug regimens are the
standard chemotherapy schemes for the treatment of breast cancer.
Relative to anthracycline, paclitaxel significantly improves the
disease-free survival and overall survival of patients; however,
paclitaxel has a greater effect when used in conjunction with
anthracycline (6–9).
The use of oncolytic viruses is a relatively new
strategy in cancer therapy. Oncolytic herpes simplex virus (oHSV)
vectors are inherently cytotoxic to and specific for tumor cells
(10). The vector G47Δ was
constructed as a third-generation replication-competent HSV-1
vector from HSV-1 laboratory strain F. The ICP47 gene and both
copies of the γ34.5 neurovirulence gene are deleted in G47Δ, and
the ribonucleotide reductase (RR) gene is inactivated by insertion
of the E. coli LacZ gene (11). Due to these gene
deletions/mutations, the replication ability of the virus is
attenuated and, therefore, the safety and tumor selectivity are
increased. The use of G47Δ has been shown to be a safe and
effective therapeutic approach for various types of cancer
(11–14).
Several preclinical studies have demonstrated that
when oHSVs are used in combination with various cytotoxic agents,
increased treatment efficacy for various malignancies is observed
(15–18). GADD34 is a DNA repair enzyme that is
homologous with the HSV-1 γ34.5 gene. Due to safety considerations,
γ34.5 was deleted in the construction of G47Δ. This deletion
attenuates the replication ability of HSV. Previous studies have
demonstrated that chemotherapy agents that upregulate GADD34 in
cancer cells in response to DNA damage enhance the antitumor
effects of oHSV by facilitating viral proliferation (15–18).
However, no published studies have investigated the mechanistic
interactions between G47Δ and paclitaxel in the treatment of breast
cancer.
Our previous preclinical studies showed that G47Δ
can effectively treat primary breast tumors and brain and lung
metastases (14,19,20).
In the present study, we found that G47Δ and paclitaxel, when
administered together, demonstrated a synergistic effect against
the breast cancer cell lines MCF-7 and MDA-MB-468 in vitro.
However, the use of paclitaxel did not enhance or impair viral
proliferation. G47Δ replicated and spread among cancer cells,
causing cancer cell lysis and enhancing paclitaxel activity. G47Δ
facilitated the induction of mitotic arrest and apoptosis by
paclitaxel. Finally, in vivo efficacy studies demonstrated
that G47Δ and paclitaxel combination therapies are a safe and
effective regimen for the treatment of breast cancer. When using
paclitaxel in combination with G47Δ, the dose of paclitaxel could
be reduced at least 5-fold while maintaining levels of tumor
reduction similar to those achieved with the administration of
paclitaxel alone. To the best of our knowledge, this is the first
study to demonstrate that the combination of G47Δ and paclitaxel
has synergistic effects in the treatment of breast cancer.
Materials and methods
Cells and viruses
MCF-7 and MDA-MB-468 cells (all obtained from Dr
Xiao-Ming Xie, Sunsen University Cancer Center, China) were grown
in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, Carlsbad,
CA, USA) supplemented with 10% heat-inactivated fetal calf serum
(Invitrogen) and 4.5 g/l glucose. Vero cells (African green monkey
kidney cells; American Type Culture Collection, Manassas, VA, USA)
were grown in DMEM supplemented with 10% heat-inactivated fetal
calf serum (Invitrogen). Cells were cultured at 37°C and 5%
CO2. G47Δ was obtained from Samuel D. Rabkin (Molecular
Neurosurgery Laboratory, Massachusetts General Hospital, Harvard
Medical School, Boston, MA, USA) and it was constructed as
previously described (11).
Cell susceptibility assays and
Chou-Talalay analysis
MCF-7 and MDA-MB-468 cells were seeded at 2,000
cells/well into 96-well plates. Following overnight incubation,
paclitaxel (Sigma, St. Louis, MO, USA) or G47Δ was added at the
indicated concentration. To select the appropriate dose ranges, we
used serial dilutions to test cell susceptibility. After 4 days of
incubation, cytotoxicity assays were performed using a Cell
Counting Kit-8 (Dojindo, Japan) according to the manufacturer’s
instructions. Median effect doses (ED50) were calculated
for the drug and G47Δ for each cell line.
For combination studies, G47Δ and paclitaxel were
added to cells at a fixed dose ratio, and Cell Counting Kit-8
assays were performed after 4 days of incubation. To analyze the
combination of G47Δ and paclitaxel, Chou-Talalay combination
indices (CI) were calculated using CompuSyn software (Combo Syn,
Inc., Paramus, NJ, USA). Fixed ratios of G47Δ and paclitaxel and
mutually exclusive equations were used to determine the CIs. A CI
between 0.9 and 1.1 is considered additive, whereas CI<0.9 and
CI>1.1 indicate synergism and antagonism, respectively.
Viral proliferation
To determine whether paclitaxel enhances G47Δ
proliferation, we tested the viral titers with or without the
addition of paclitaxel. MCF-7 and MDA-MB-468 cells were seeded into
12-well plates at 2×104 cells/well overnight and then
treated with G47Δ at an MOI of 0.5 (MCF-7) or 1.5 (MDA-MB-468) and
with paclitaxel at 1 nmol/l (MCF-7) or 3 nmol/l (MDA-MB-468). The
cells and supernatants were collected for 4 days. After three
freeze-thaw cycles, the titers of infectious virus were determined
using a plaque assay with Vero cells. Next, we calculated the viral
pfu per viable cancer cell. The cells were treated with G47Δ and a
range of paclitaxel doses for 2 days. The viable cells were
counted, and the viral titers were calculated.
Flow cytometric analysis of the cell
cycle
MCF-7 and MDA-MB-468 cells were seeded into 10-cm
dishes at 8×105 cells/plate and treated with either the
mock treatment, G47Δ, paclitaxel or the combination of G47Δ and
paclitaxel. At the indicated time points, the adherent and detached
cells were collected and fixed in 70% ethanol at 4° overnight. The
cells were then washed twice in phosphate-buffered saline (PBS)
containing bovine serum albumin (0.5%) and treated in 1 ml of PBS
containing 0.1% Triton-X, RNase A (100 μg/ml; Sigma) and propidium
iodide (PI) (50 μg/ml; Sigma) at room temperature for 30 min. The
cells were then immediately analyzed by flow cytometry in a BD
FACSCalibur. The resulting data were analyzed with ModFit LT v3.2
(Verity Software House, Topsham, ME, USA).
Apoptosis assay
MCF-7 and MDA-MB-468 cells were seeded into 6-cm
dishes at 4×105 cells/plate and treated with either the
mock treatment, G47Δ, paclitaxel or the combination of G47Δ and
paclitaxel. Following 48 h of incubation, the adherent and detached
cells were collected and washed twice with PBS. The cells were then
counted, adjusted to a density of 1×106/ml, and
double-stained with fluorescein isothiocyanate-conjugated Annexin-V
and PI using an Annexin-V-FITC Apoptosis Detection Kit according to
the manufacturer’s instructions (Nanjing KeyGen Biotech., Co.,
Ltd., Nanjing, China). The stained cells were then immediately
analyzed using flow cytometry with a BD FACSCalibur. The resulting
data were analyzed using FlowJo v8.5.3 software (Tree Star,
Ashland, OR, USA).
In vivo treatment studies
First, 5×106 MDA-MB-468 cells were
implanted into the left flank of 6-week-old female BALB/c nude mice
(Vital River Laboratory Animal Technology Co., Ltd., Beijing,
China). When the maximum diameters of the tumors reached ~5 mm, the
mice were randomized into 5 groups (n=7 per group): a G47Δ
treatment group treated with intratumor (i.t.) injections of G47Δ
virus (2×105 pfu) on Days 0 and 3, two paclitaxel
treatment groups treated with intraperitoneal (i.p.) injections of
paclitaxel (3 or 15 mg/kg) on Days 0 and 7 of a 7-day cycle for 2
weeks, a combination treatment group treated with G47Δ
(2×105 pfu) and paclitaxel (3 mg/kg), and a mock
treatment group. The tumor volume was calculated using the formula
width (mm)2 × length (mm) × 0.5. The body weight and
motor activity of each animal were monitored as indicators of
general health and toxicity.
Statistical analysis
The Student’s t-test (two-tailed) was used to
analyze the significance of differences between the treatment
groups. The test was implemented using the SPSS version 13.0
software. A P-value of <0.05 was considered to indicate a
statistically significant difference.
Results
G47Δ exhibits a synergistic effect in
conjunction with paclitaxel in the killing of breast cancer cells
in vitro
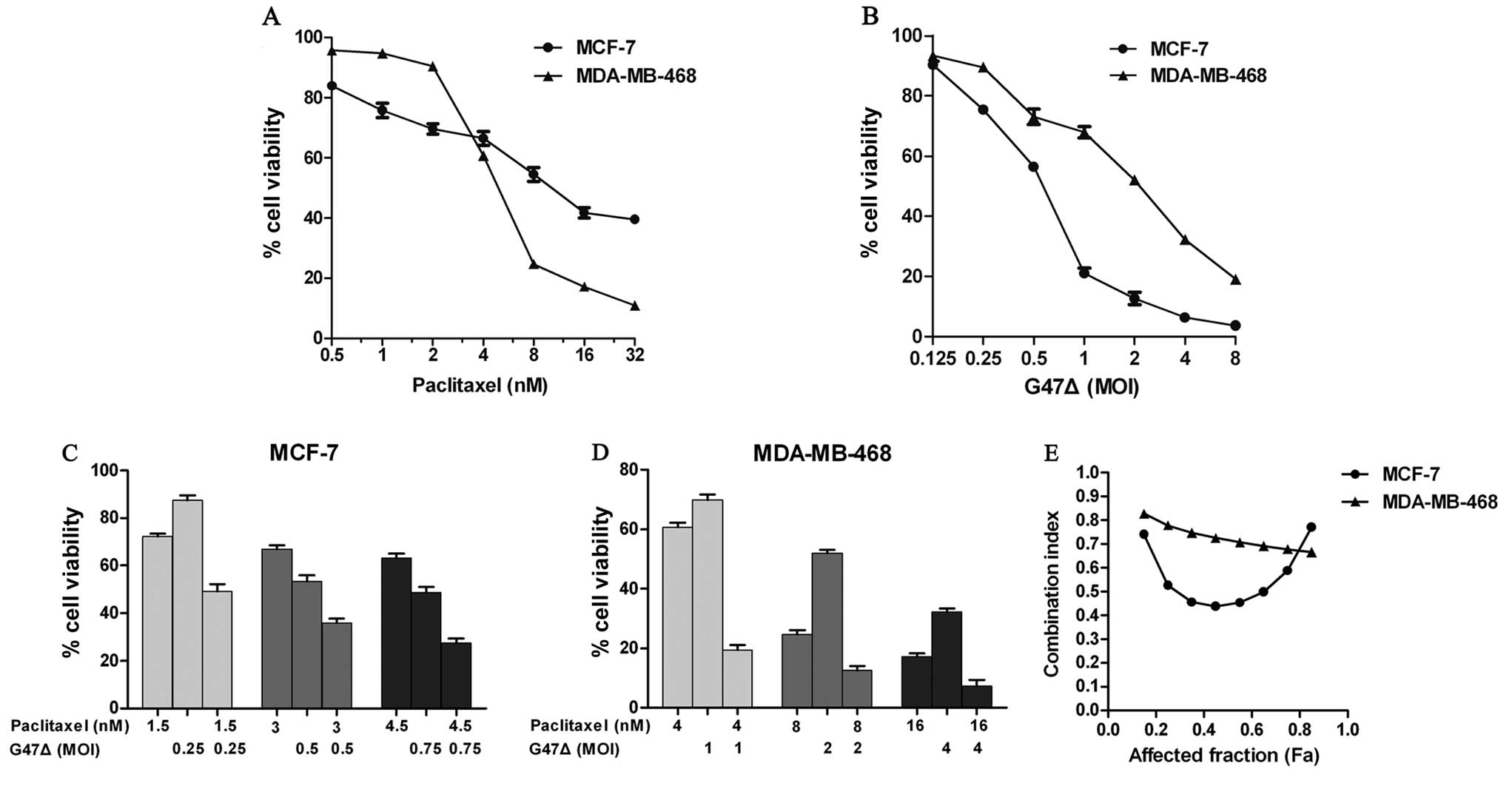
First, we performed cytotoxicity assays of
paclitaxel and G47Δ with MCF-7 and MDA-MB-468 cells. Both the MCF-7
and MDA-MB-468 cells exhibited dose-dependent cytotoxicity
following exposure to paclitaxel or G47Δ (Fig. 1). MDA-MB-468 cells were more
sensitive than MCF-7 cells to paclitaxel at higher concentrations
(4–32 nmol/l), although MDA-MB-468 cells were less sensitive at
lower concentrations (0.5–2 nmol/l). MCF-7 cells were more
sensitive than MDA-MB-468 cells to G47Δ cytotoxicity. The
ED50, i.e., the dose causing 50% cytotoxicity, was
calculated for each agent. The ED50 values of G47Δ were
an MOI of 0.70 (MCF-7) and an MOI of 1.93 (MDA-MB-468). The
ED50 values of paclitaxel were 11.54 nm/l (MCF-7) and
5.90 nm/l (MDA-MB-468).
Next, we performed combination cytotoxicity assays
at a fixed concentration ratio based on the above results. CIs were
calculated to determine whether G47Δ and paclitaxel used in
combination exhibit a synergetic breast cancer cell-killing effect
in vitro. As defined, a CI between 0.9 and 1.1 is considered
additive, whereas CI<0.9 and CI>1.1 indicate synergism and
antagonism, respectively. The combination of G47Δ and paclitaxel
produced a synergistic effect against both MCF-7 and MDA-MB-468
cells (Fig. 1). The Chou-Talalay
CIs ranged from 0.44 to 0.77 for MCF-7 cells and from 0.68 to 0.83
for MDA-MB-468 cells. The most significant synergistic effect was
achieved at moderate and high concentrations for MCF-7 cells and
MDA-MB-468 cells, respectively. Overall, these results demonstrate
that G47Δ and paclitaxel exhibit a synergistic anti-MCF-7 and
anti-MDA-MB-468 effect in vitro when used in
combination.
Paclitaxel does not affect the
replication of G47Δ
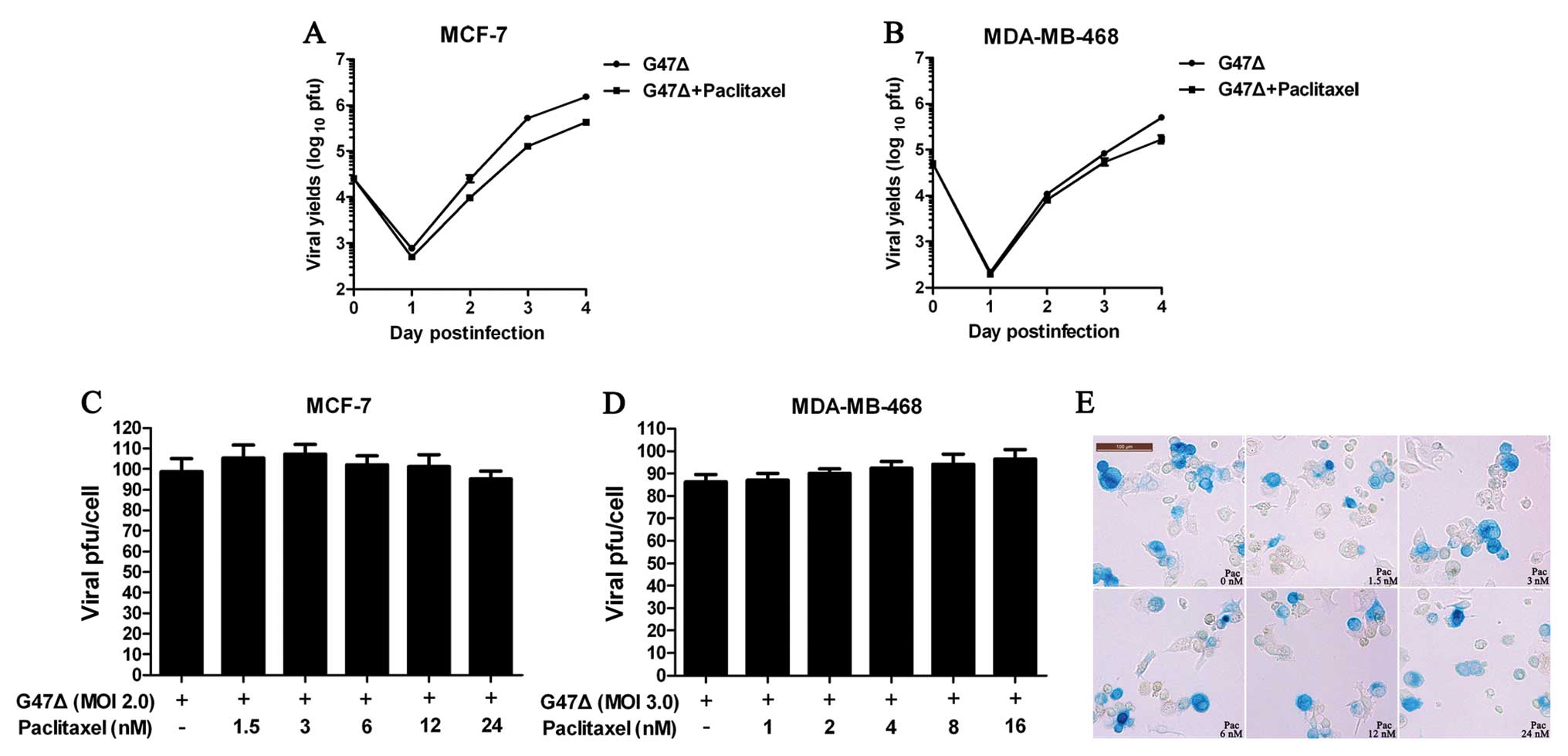
To determine whether paclitaxel affects the
replication of G47Δ in vitro, we calculated viral titers
using plaque assays after infecting the cells with G47Δ and
exposing the cells to paclitaxel or vehicle. The viral titers were
tested daily. We found that paclitaxel did not significantly affect
the replication of G47Δ (Fig. 2A and
B). To extend these results, we next tested viral proliferation
using different doses of paclitaxel and calculated the viral titer
per viable cell. Similar to our previous results, the viral titers
per viable cell did not significantly change across the different
paclitaxel doses tested against MCF-7 or MDA-MB-468 cells (Fig. 2C and D). Finally, X-gal staining of
MCF-7 cells infected with G47Δ and exposed to different doses of
paclitaxel confirmed that paclitaxel did not significantly affect
the spread of G47Δ (Fig. 2E).
G47Δ facilitates the induction of mitotic
arrest by paclitaxel
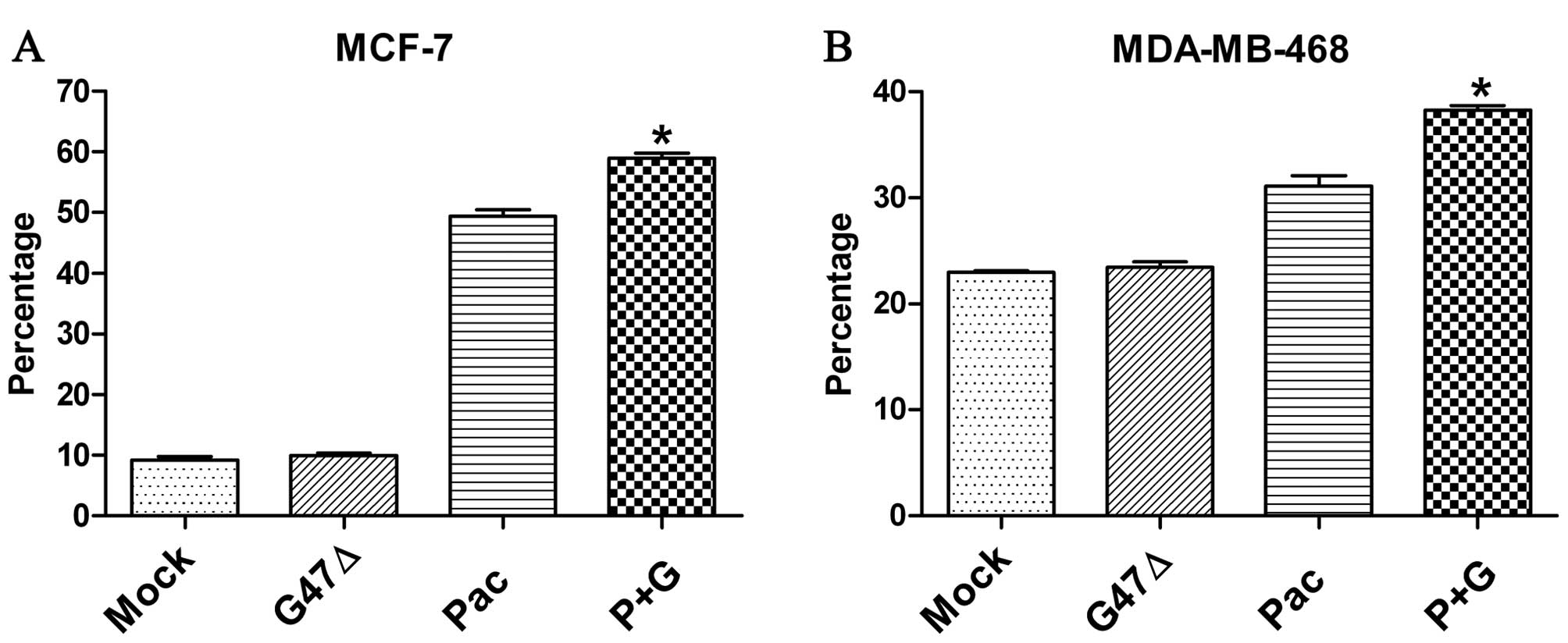
Subsequently, we determined whether G47Δ affects the
antitumoral activity of paclitaxel. Paclitaxel exerts its cytotoxic
effect by interacting with β-tubulin, stabilizing the structure of
microtubules and preventing the depolymerization of microtubules.
This microtubule stabilization leads to cell cycle arrest in the
G2/M phase and ultimately leads to cell death by apoptosis. We
found that G47Δ alone did not significantly influence the cell
cycle (Fig. 3). However, relative
to treatment with paclitaxel alone, treatment with G47Δ plus
paclitaxel significantly promoted cell cycle arrest in the G2/M
phase after a 24-h exposure for both MCF-7 and MDA-MB-468 cells
(Fig. 3). For MCF-7 cells, the
percentage of G2/M phase-arrested cells for the paclitaxel alone
group and the combination group were 49.35±1.01% and 58.96±0.83%
(P<0.001), respectively. For MDA-MB-468 cells, the percentages
were 31.11±0.96% and 38.29±0.43% (P<0.001), respectively.
G47Δ enhances the ability of paclitaxel
to induce apoptosis in breast cancer cells
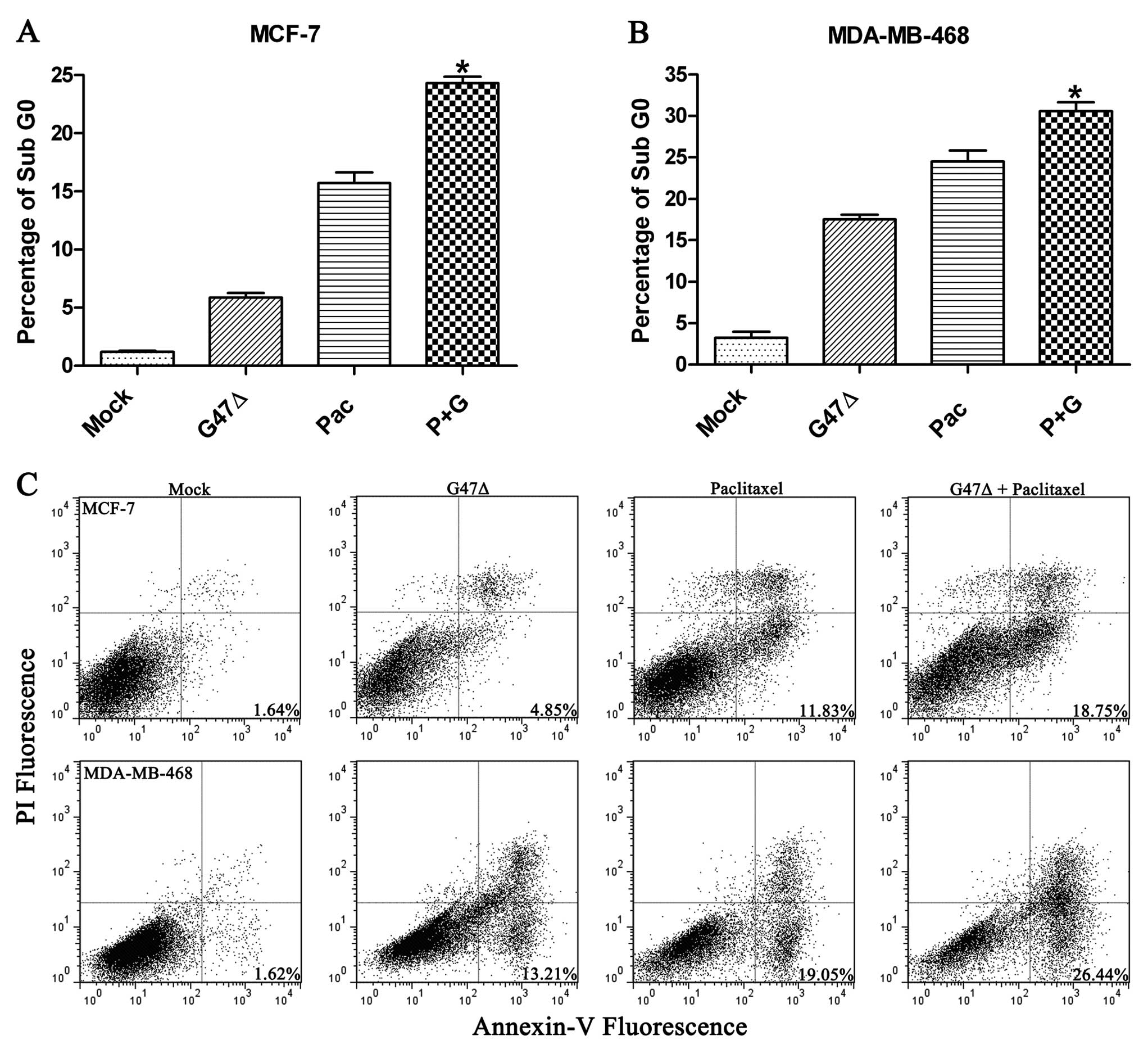
We used two methods to assess the level of
apoptosis. First, we calculated the percentage of sub-G0 cells
after 72 h of exposure to the mock treatment, paclitaxel, G47Δ or a
combination of paclitaxel and G47Δ. Relative to the mock treatment
(1.17±0.12%), paclitaxel alone (15.70±0.95%), G47Δ alone
(5.37±0.385%), and the combination of paclitaxel and G47Δ
(24.29±0.60%) significantly increased the level of apoptosis in
MCF-7 cells (P<0.05 for all comparisons) (Fig. 4A). Similar results were observed
with MDA-MB-468 cells (Fig. 4B).
Annexin-V and PI staining also confirmed that paclitaxel and G47Δ
in combination produced a significant increase in apoptosis
(Fig. 4C).
G47Δ and paclitaxel exhibit synergistic
effects in vivo
Finally, we tested whether G47Δ and paclitaxel could
exhibit a synergistic effect in vivo. We chose MDA-MB-468
cells to form tumors in BALB/C mice. After flank tumors developed,
the mice were divided into 5 treatment groups. One group was
treated with a mock treatment, another group received i.t. G47Δ
(2×105 pfu) injections on Days 0 and 3, two other groups
received i.p. paclitaxel injections at different doses (3 or 15
mg/kg) twice a week for 2 weeks, and one group received a
combination of G47Δ (2×105 pfu) + paclitaxel (3 mg/kg).
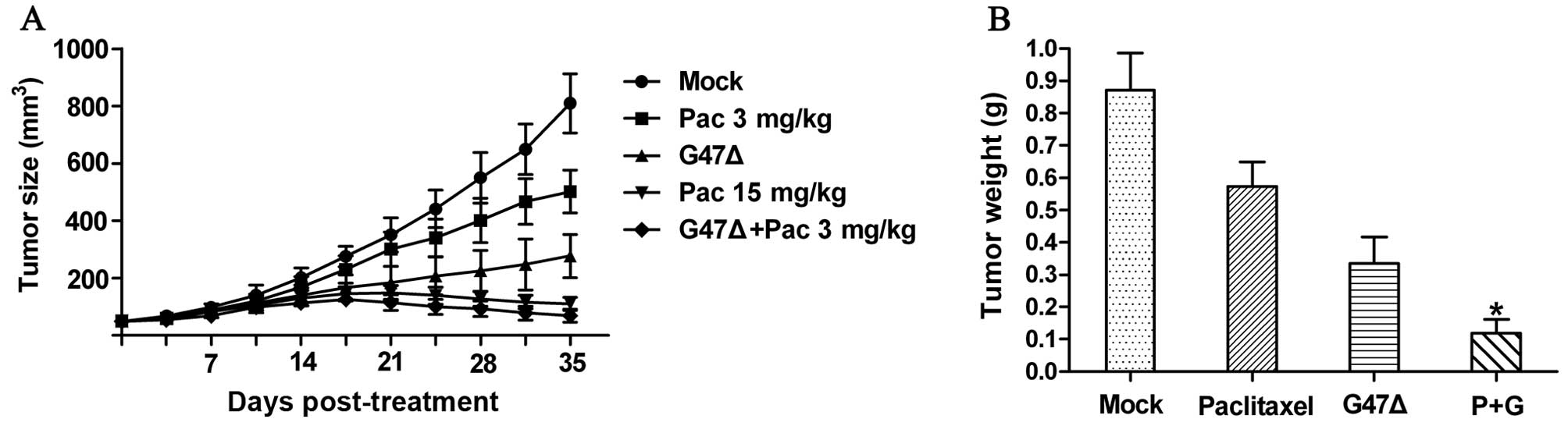
We found that both G47Δ and paclitaxel could inhibit tumor growth
in vivo. Compared with the mock treatment group, both the
G47Δ and paclitaxel (3 mg/kg) alone groups presented a significant
reduction in the mean tumor volume by Day 35. In addition, we
observed that the treatment of mice with a combination of G47Δ and
paclitaxel resulted in a synergistic effect in vivo and a
significant reduction in the mean tumor volume (68.71±22.65
mm3) compared to treatment with G47Δ alone (277.57±75.35
mm3), paclitaxel alone (501.86±74.79 mm3), or
the mock treatment (809.14±102.58 mm3) by Day 35
(P<0.05 for all three comparisons) (Fig. 5A). Of note, when paclitaxel was
combined with G47Δ, the dose of paclitaxel could be reduced at
least 5-fold while maintaining levels of tumor reduction similar to
those attained with the administration of paclitaxel alone at 15
mg/kg. We also found that the combination treatment produced a
significant reduction in tumor weight compared with the other
treatments (Fig. 5B). Furthermore,
the combined treatment caused no additional signs of toxicity, and
the body weight of the mice was similar among all groups (mean body
weight, 22.5 g). In addition, the body weight did not change
significantly over the course of the study.
Discussion
Breast carcinoma is a relatively low-malignancy type
of tumor. Patients who suffer from early-stage breast cancer have
positive clinical outcomes. However, approximately 30% of patients
with early-stage disease eventually develop recurrent or metastatic
lesions (21). At present,
metastatic cancer remains an incurable disease. Therefore, new
therapies are urgently required. Our group demonstrated previously
that G47Δ effectively targets primary breast tumors and brain and
lung metastases (14,19,20).
To increase safety and tumor selectivity, some genes
were deleted from or mutated in the viral vector, including the RR
gene and the γ34.5 neurovirulence gene. These changes attenuated
the replication ability of HSV, resulting in oHSV being relatively
selective for dividing tumor cells as there are cellular homologs
that can compensate for the missing genes, such as RR and GADD34
for γ34.5. Some preclinical studies have reported that oHSVs, when
combined with chemotherapeutic agents that upregulate the
expression or activities of RR and/or GADD34 in tumor cells,
exhibit synergistic effects against various tumors (15–18).
Petrowsky et al(15)
reported that fluorodeoxyuridine, which causes nucleotide pool
imbalances and DNA damage-induced upregulation of RR and GADD34 in
colorectal cancer cell lines, promotes enhanced viral replication
and tumor cell death.
In the present study, we demonstrated, using
Chou-Talalay assays, that G47Δ and paclitaxel in combination
exhibited a synergistic effect against MCF-7 and MDA-MB-468 breast
cancer cells. We initially hypothesized that paclitaxel might
enhance viral replication. However, both viral titer experiments
and X-gal staining indicated that paclitaxel did not significantly
influence viral replication or spread. These results indicated that
the mechanism mediating the synergistic cytotoxicity between G47Δ
and paclitaxel does not involve the enhancement of viral
replication by paclitaxel.
To further investigate the interaction between G47Δ
and paclitaxel, we determined whether G47Δ enhanced the antitumor
activity of paclitaxel. Paclitaxel stabilizes the structure of
microtubules, leading to cell cycle arrest in the G2/M phase and
ultimately inducing cell death by apoptosis. First, we found that
although G47Δ alone did not significantly impact the cell cycle of
either MCF-7 or MDA-MB-468 cells, G47Δ and paclitaxel, when
administered in combination, resulted in significantly greater
numbers of tumor cells arrested in the G2/M phase, compared to
treatment with either paclitaxel or G47Δ alone. Then, we found that
both G47Δ and paclitaxel could cause tumor cell apoptosis, but G47Δ
alone only produced a small percentage of apoptotic cells. However,
G47Δ significantly enhanced the ability of paclitaxel to induce
apoptosis, relative to paclitaxel alone. Elliott and O’Hare
(22) found that the HSV-1 tegument
protein VP22 can reorganize microtubules into thick bundles that
became highly resistant to microtubule-depolymerizing agents,
suggesting that VP22 might have the capacity to stabilize the
microtubule network. Our data indicate that G47Δ administered by
itself at a low dose did not significantly influence the cell cycle
of MCF-7 or MDA-MD-468 cells, but VP22 might enhance the ability of
paclitaxel to stabilize the structure of microtubules, eventually
leading to G2/M phase arrest and cell apoptosis.
Although paclitaxel is used widely to treat a
variety of tumors, the toxic effect of the drug is a key factor in
restricting its broader clinical use. Hematopoietic toxicity and
cumulative peripheral neuropathy limit the long-term use of
paclitaxel, which always leads to dose reduction and the delay of
paclitaxel chemotherapy, particularly for patients with recurrent
or metastatic disease, most of whom have been heavily treated for
the primary disease. Kim et al(23) found that when breast cancer patients
received adriamycin, cyclophosphamide and paclitaxel as adjuvant
chemotherapy for their primary tumors, the drug dose was reduced
for 17.1% of patients, and 14.3% patients delayed treatment due to
the toxicity of the chemotherapy agents. Loibl et
al(24) observed that patients
receiving a lower relative total dose had a shorter overall
survival. In the present study, we demonstrated that G47Δ and
paclitaxel combination therapy can reduce the required paclitaxel
dose by at least 5-fold while maintaining levels of tumor reduction
similar to those achieved with the administration of paclitaxel
alone. In addition, the combination therapy caused no additional
signs of toxicity. These data may have important clinical
implications. For some patients, especially patients who have ever
been heavily treated for recurrent or metastatic disease, G47Δ and
paclitaxel combined therapy may be an effective and safe
therapeutic regimen.
In summary, we report that a third-generation
replication-competent HSV-1 vector G47Δ and paclitaxel, when given
in combination, had a synergistic anti-breast cancer effect both
in vitro and in vivo. Paclitaxel did not
significantly influence the replication and spread of G47Δ, but
G47Δ may enhance the antitumor activity of paclitaxel through
mitosis arrest and apoptosis. G47Δ and paclitaxel combined therapy
appears to be an effective and safe therapeutic regimen for the
treatment of breast cancer, and, thus, the presented data may have
critical clinical implications for the treatment of breast cancer.
To the best of our knowledge, this is the first report
demonstrating a synergetic effect of a combination of G47Δ and
paclitaxel against breast cancer cells.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 30672410) and the Natural
Science Foundation of Guangdong Province, China (grant no.
06104599).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Perou CM, Sørlie T, Eisen MB, et al:
Molecular portraits of human breast tumours. Nature. 406:747–752.
2000. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sørlie T, Perou CM, Tibshirani R, et al:
Gene expression patterns of breast carcinomas distinguish tumor
subclasses with clinical implications. Proc Natl Acad Sci USA.
98:10869–10874. 2001.PubMed/NCBI
|
|
4
|
Sotiriou C, Neo SY, McShane LM, et al:
Breast cancer classification and prognosis based on gene expression
profiles from a population-based study. Proc Natl Acad Sci USA.
100:10393–10398. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lim E and Winer EP: Adjuvant chemotherapy
in luminal breast cancers. Breast. 20(Suppl 3): S128–S131. 2011.
View Article : Google Scholar
|
|
6
|
Paridaens R, Biganzoli L, Bruning P, et
al: Paclitaxel versus doxorubicin as first-line single-agent
chemotherapy for metastatic breast cancer: a European Organization
for Research and Treatment of Cancer Randomized Study with
cross-over. J Clin Oncol. 18:724–733. 2000.PubMed/NCBI
|
|
7
|
Sledge GW, Neuberg D, Bernardo P, Ingle
JN, Martino S, Rowinsky EK and Wood WC: Phase III trial of
doxorubicin, paclitaxel, and the combination of doxorubicin and
paclitaxel as front-line chemotherapy for metastatic breast cancer:
an intergroup trial (E1193). J Clin Oncol. 21:588–592. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ghersi D, Wilcken N and Simes RJ: A
systematic review of taxane-containing regimens for metastatic
breast cancer. Br J Cancer. 93:293–301. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Henderson IC, Berry DA, Demetri GD, et al:
Improved outcomes from adding sequential paclitaxel but not from
escalating doxorubicin dose in an adjuvant chemotherapy regimen for
patients with node-positive primary breast cancer. J Clin Oncol.
21:976–983. 2003. View Article : Google Scholar
|
|
10
|
Varghese S and Rabkin SD: Oncolytic herpes
simplex virus vectors for cancer virotherapy. Cancer Gene Ther.
9:967–978. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Todo T, Martuza RL, Rabkin SD and Johnson
PA: Oncolytic herpes simplex virus vector with enhanced MHC class I
presentation and tumor cell killing. Proc Natl Acad Sci USA.
98:6396–6401. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fukuhara H, Martuza RL, Rabkin SD, Ito Y
and Todo T: Oncolytic herpes simplex virus vector g47delta in
combination with androgen ablation for the treatment of human
prostate adenocarcinoma. Clin Cancer Res. 11:7886–7890. 2005.
View Article : Google Scholar
|
|
13
|
Wang JN, Hu P, Zeng MS and Liu RB:
Anti-tumor effect of oncolytic herpes simplex virus G47delta on
human nasopharyngeal carcinoma. Chin J Cancer. 30:831–841. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu R, Varghese S and Rabkin SD: Oncolytic
herpes simplex virus vector therapy of breast cancer in C3(1)/SV40
T-antigen transgenic mice. Cancer Res. 65:1532–1540. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Petrowsky H, Roberts GD, Kooby DA, et al:
Functional interaction between fluorodeoxyuridine-induced cellular
alterations and replication of a ribonucleotide reductase-negative
herpes simplex virus. J Virol. 75:7050–7058. 2001. View Article : Google Scholar
|
|
16
|
Eisenberg DP, Adusumilli PS, Hendershott
KJ, et al: 5-fluorouracil and gemcitabine potentiate the efficacy
of oncolytic herpes viral gene therapy in the treatment of
pancreatic cancer. J Gastrointest Surg. 9:1068–1077. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bennett JJ, Adusumilli P, Petrowsky H, et
al: Up-regulation of GADD34 mediates the synergistic anticancer
activity of mitomycin C and a gamma134.5 deleted oncolytic herpes
virus (G207). FASEB J. 18:1001–1003. 2004.PubMed/NCBI
|
|
18
|
Aghi M, Rabkin S and Martuza RL: Effect of
chemotherapy-induced DNA repair on oncolytic herpes simplex viral
replication. J Natl Cancer Inst. 98:38–50. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu R, Martuza RL and Rabkin SD:
Intracarotid delivery of oncolytic HSV vector G47Delta to
metastatic breast cancer in the brain. Gene Ther. 12:647–654. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang J, Hu P, Zeng M, Rabkin SD and Liu R:
Oncolytic herpes simplex virus treatment of metastatic breast
cancer. Int J Oncol. 40:757–763. 2012.PubMed/NCBI
|
|
21
|
Gonzalez-Angulo AM, Morales-Vasquez F and
Hortobagyi GN: Overview of resistance to systemic therapy in
patients with breast cancer. Adv Exp Med Biol. 608:1–22. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Elliott G and O’Hare P: Herpes simplex
virus type 1 tegument protein VP22 induces the stabilization and
hyperacetylation of microtubules. J Virol. 72:6448–6455.
1998.PubMed/NCBI
|
|
23
|
Kim WY, Woo SU, Seo JH, Son GS, Lee JB and
Bae JW: Toxicities, dose reduction and delay of docetaxel and
paclitaxel chemotherapy in breast cancer without distant
metastases. J Cancer Res Ther. 7:412–415. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Loibl S, Skacel T, Nekljudova V, et al:
Evaluating the impact of Relative Total Dose Intensity (RTDI) on
patients’ short and long-term outcome in taxane- and
anthracycline-based chemotherapy of metastatic breast cancer- a
pooled analysis. BMC Cancer. 12:121–131. 2011.PubMed/NCBI
|