Introduction
We recently described in vivo effects of
high-dose lipotropes that reduce mammary tumor growth of female rat
offspring by suppressing histone deacetylase 1 (HDAC1) gene
expression (1). In an attempt to
further expand our knowledge on the effects of high-dose
lipotropes, we conducted a series of in vitro experiments
using MCF-7 human breast cancer cells in which we combined
lipotropes with chemotherapeutic drugs such as the HDAC inhibitor
suberoylanilide hydroxamic acid (SAHA) and the anticancer drug
doxorubicin (DOX).
Nutritional studies as well as epidemiological
studies have shown that dietary manipulation of methyl donors and
cofactors (either deficiency or supplementation) can lead to
permanent alterations in patterns of gene expression (2). Mammalian methyl metabolism is
dependent on dietary nutrients which influence DNA and histone
methylation (3). Previous studies
revealed that reduced dietary intake or low tissue/plasma levels of
methyl nutrients are associated with higher risk for developing
cancer, while increased methyl nutrient intake reduces the risk of
neural tube defects and the incidence of low birth weight (4,5).
Lipotropes are methyl group-containing essential
nutrients (methionine, choline, folate and vitamin B12)
which can affect genomic DNA methylation, a process correlated with
controlling the expression of genes involved in cell growth,
apoptosis, and metabolism (6–8).
Lipotropes play key roles in one-carbon metabolism, a process that
maintains the imprinting status of genes and provides methyl groups
for all biological methylation pathways (6–8).
Methionine and choline are major methyl donors, while folate and
vitamin B12 are critical cofactors for methyl metabolism
(7). Methyl donors and cofactors
are also involved in the synthesis of nucleotides and phospholipids
as well as amino acid metabolism and signal transduction (3,8).
Apoptosis is critical for the pathogenesis of cancer
and its regulation is often impaired in cancer cells (9). Caspase-3 is a critical regulator
involved in the execution phase of apoptosis and some cancer cells
evade apoptosis by suppressing caspase-3 (10). The p53 tumor suppressor gene plays a
key role in inhibiting cancer cell growth and murine double minute
oncogene 2 (MDM2) binds specifically to the p53 protein and
negatively regulates its transcriptional activity and stability
through multiple mechanisms in cancer cells (11,12).
The interruption of p53 activity by MDM2 is one of the main
mechanisms by which cancer cells evade apoptosis (12).
Increasing the susceptibility of tumor cells to
apoptosis is one of the main strategies of cancer treatment
(9). HDAC inhibitors are emerging
chemotherapeutic drugs and have been characterized as potential
inducers of growth arrest and apoptosis of cancer cells in
vitro and in vivo(13).
Drug resistance of cancer cells to a variety of chemotherapeutic
agents is one of the major obstacles for cancer treatment (14). Defects in apoptosis signaling
contribute to the development of drug resistance in cancer
chemotherapy (15). Apoptosis plays
an essential role in drug resistance and modulating apoptosis in
tumor cells is a principal target in breast cancer treatment
(15).
Although there is a growing body of literature
investigating the intake of individual nutrients and breast cancer
risk, less is known about the interplay among these methyl
nutrients and their implications for breast cancer risk reduction.
Herein, we investigated the relationship between apoptosis and the
synergistic effects of lipotropes with anticancer drugs in MCF-7
human breast cancer cells.
Materials and methods
Cell line and cell culture
The estrogen receptor (ER) positive MCF-7 (HTB-22,
adenocarcinoma) human breast cancer cell line was purchased from
the American Type Culture Collection (ATCC, Manassas, VA, USA). The
cells were grown in Dulbecco’s modified Eagle’s medium (Gibco
Invitrogen, Carlsbad, CA, USA) supplemented with 10% (v/v)
heat-inactivated fetal bovine serum (Gibco Invitrogen) and 1% (v/v)
antibiotic-antimycotic (Gibco Invitrogen) solution as recommended
by the supplier. Cells were maintained as monolayer cultures in 25-
and 75-cm2 tissue culture flasks (BD Biosciences,
Franklin Lakes, NJ, USA) at 37°C in a 5% CO2-humidified
atmosphere during experimentation. Cells were passaged using 0.5%
trypsin-EDTA (Gibco Invitrogen) at 80–90% confluence. The lipotrope
stock solutions were prepared by dissolving L-methionine, choline
chloride, and folic acid (Table I)
(Sigma-Aldrich, St. Louis, MO, USA) in culture media and filtered
aseptically through 0.45 μm pore size cellulose acetate membrane
filters (Nalgene, Rochester, NY, USA). The stock solutions were
pre-warmed and vigorously stirred before adding to cell cultures.
The appropriate or optimal dose of lipotropes was determined from
dose response studies (0, 5, 10 and 20 times).
 | Table IAmount of lipotropes in culture
medium. |
Table I
Amount of lipotropes in culture
medium.
| DMEM |
|---|
|
|
|---|
| Ingredients
(mg/l) | Control
(basal) | Lipotropes
(20x) |
|---|
| L-Methionine | 30.0 | 600.0 |
| Choline
chloride | 4.0 | 80.0 |
| Folic acid | 4.0 | 80.0 |
| Vitamin
B12 | 0.0 | 0.0 |
Cell proliferation assay
Colorimetric MTS
[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium]
assay (CellTiter 96® AQueous One Solution Reagent;
Promega, Madison, WI, USA) was used to measure cell proliferation.
Briefly, cells were seeded in 96-well flat-bottomed tissue culture
plates (5×103 cells/well in 100 μl), and incubated with
various concentrations of lipotropes for a duration of 0, 24, 48,
72 or 96 h in cultures. Following incubation, 10 μl of MTS solution
was added and cells were incubated at 37°C for a further 1–4 h. The
degree of cell proliferation was evaluated numerically by measuring
the absorbance at 490 nm with a SpectraMax Microplate Reader
(Molecular Devices, Sunnyvale, CA, USA). The cell proliferation was
calculated and expressed based on the following formula: {[treated
cell absorbance - initial (seeding) cell absorbance]/initial
(seeding) cell absorbance} × 100.
Caspase-3 assay
The caspase-3 activity was measured by a
colorimetric assay with the CaspACE assay system (Promega).
Briefly, cells were cultured in 6-well flat-bottomed tissue culture
plates (5×104 cells/ml) and treated with lipotropes (20
times) for 96 h. Cells were harvested and washed twice with
ice-cold PBS and then lysed in lysis buffer (Promega). The protein
concentration was quantified using a NanoDrop 2000c (Thermo Fisher
Scientific, Waltham, MA, USA). Cell lysates (25 μg/sample) were
incubated with colorimetric substrate,
N-acetyl-Asp-Glu-Val-Asp-amino-p-nitroanilide (Ac-DEVD-pNA).
Following overnight incubation at 4°C, the release of
p-nitroaniline from Ac-DEVD-pNA was measured at 405 nm using a
SpectraMax Microplate Reader (Molecular Devices).
p53 assay
The p53 activity was determined using an enzyme
immunometric assay kit (TiterZyme ELISA Kit, Assay Designs, Ann
Arbor, MI, USA). Briefly, cells were cultured in 6-well
flat-bottomed tissue culture plates (5×104 cells/ml) and
treated with lipotropes (20 times) for 96 h. Cells were harvested
and washed twice with ice-cold PBS and then resuspended in lysis
buffer (Sigma-Aldrich). The protein concentration was quantified
using a NanoDrop 2000c (Thermo Fisher Scientific). The supernatants
(100 μg/sample) were incubated on a plate pre-immobilized with p53
polyclonal antibody and then reacted with the labeled antibody. The
absorbance was measured at 450 nm using a SpectraMax Microplate
Reader.
Flow cytometric analysis
Apoptosis was determined by double-staining with
fluorescein isothiocyanate (FITC)-conjugated Annexin V and
propidium iodide (PI) (Sigma-Aldrich) as previously described
(16). Briefly, cells were cultured
in 12-well flat-bottomed tissue culture plates (5×104
cells/ml) and treated with lipotropes (20 times) for 96 h. After
harvesting, cells were washed with PBS and resuspended in assay
buffer (Sigma-Aldrich). Cells were stained with FITC-conjugated
Annexin V and PI, and then analyzed using Accuri C6 cytometer and
CFlow software (Accuri Cytometers, Ann Arbor, MI, USA).
Quantitative real-time PCR
MCF-7 cells treated with lipotropes (20 times) for
96 h were harvested and placed in RNAlater (Ambion, Austin, TX,
USA) prior to freezing, and then disrupted into small pieces. RNA
was purified by the standard method. Briefly, cells were
homogenized in TRI Reagent (Molecular Research Center, Cincinnati,
OH, USA) and total RNA was isolated using 1-bromo-3-chloropropane
phase separation reagent (Molecular Research Center). RNA was
precipitated by isopropanol and washed with 75% ethanol and then
the RNA pellet was dried and resuspended in RNase-free water. The
RNA concentration was quantified using a NanoDrop 2000c (Thermo
Fisher Scientific). A total of 1 μg RNA of each sample was
reverse-transcribed to cDNA using the QuantiTect Reverse
Transcription kit (Qiagen, Valencia, CA, USA) and a 2720 Thermal
Cycler (Applied Biosystems, Foster City, CA, USA) at 42°C for 15
min, and 95°C for 3 min, in accordance with the manufacturer’s
recommendations. Real-time RT-PCR was performed with SYBR-Green PCR
Master Mix (Applied Biosystems) using a 7500 Fast Real-Time PCR
system (Applied Biosystems) with QuantiTect Primers (Qiagen,
product reference is in brackets); tumor protein p53 (p53,
QT00060235), murine double minute oncogene (MDM2,
QT00056378), estrogen receptor 1 (ESR1, QT00044492),
ATP-binding cassette sub-family C member 1 (ABCC1,
QT00061159), and ATP-binding cassette sub-family G member 2
(ABCG2, QT00073206). The relative amounts of gene expression
were standardized and calculated by the expression of house-keeping
control gene, β-actin (ACTB, QT01680476) as an internal
standard, using the 2−ΔΔCt method.
Cell proliferation assay with anticancer
drugs
MCF-7 cells were seeded in 96-well flat-bottomed
tissue culture plates (5×104 cells/ml) and cultured
simultaneously with media containing dimethyl sulfoxide (DMSO,
Sigma-Aldrich) vehicle alone or 250 nM SAHA (Enzo Life Sciences,
Farmingdale, NY, USA) and/or lipotropes (20 times). DOX is a well
known anticancer drug with broad spectrum antitumor efficacy,
including human breast cancer (17). MCF-7/DOX (DOX-resistant MCF-7) cells
were seeded in 96-well flat-bottomed tissue culture plates
(5×104 cells/ml) and cultured simultaneously with media
containing 100 nM of DOX (Sigma-Aldrich) and/or lipotropes (20
times). The cell proliferation was measured by the MTS assay.
Statistical analysis
For the comparison of two groups with similar
variance, a paired t-test was used. Means of several groups were
compared with one-way analysis of variance (ANOVA) followed by
Tukey’s test. Statistical data analyses were performed using
Minitab Release 14.1 (Minitab Inc., State College, PA, USA).
P<0.05 was considered to indicate statistically significant
differences.
Results
Lipotropes significantly reduce MCF-7
cancer cell growth
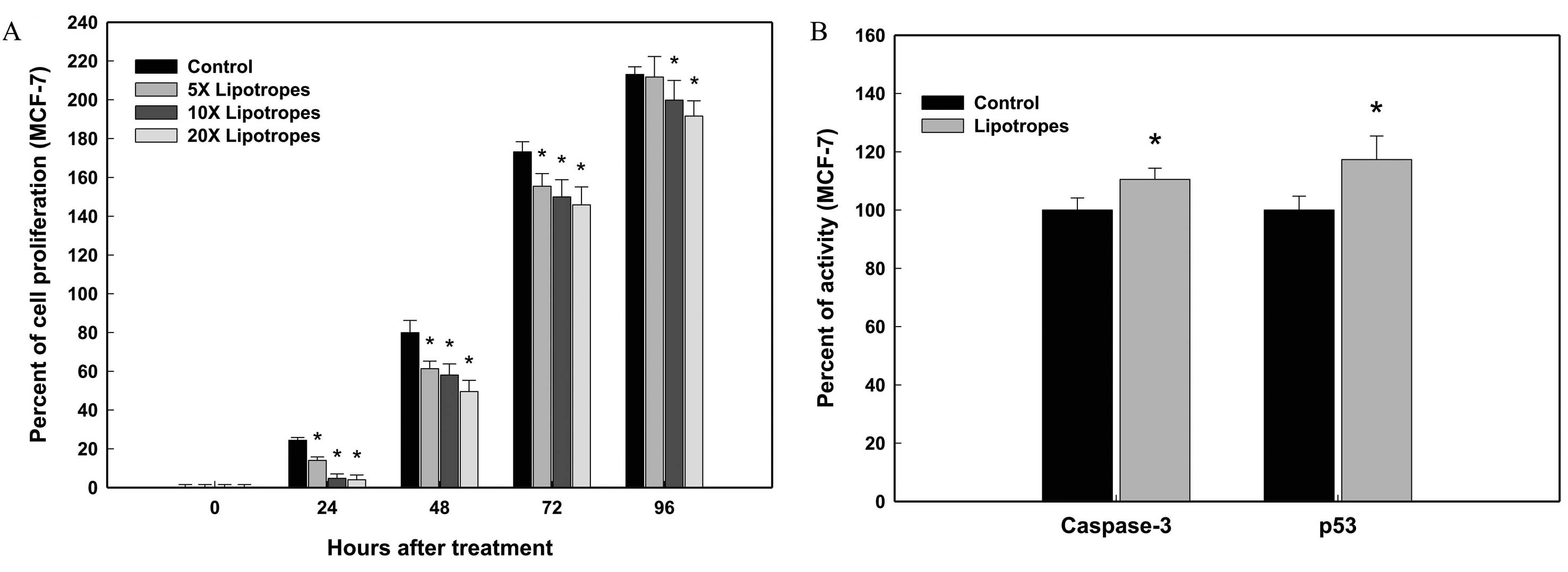
The cell proliferation of MCF-7 cells was examined
over a wide range of doses and times for dose- and time-dependent
studies. Lipotropes (20 times) lowered the cell density of MCF-7
cells at 24, 48, 76 and 96 h following treatment (Fig. 1A). At a 10 times concentration and
96 h, lipotropes showed a mild growth inhibitory effect on MCF-7
cells. As shown in Fig. 1A,
lipotropes resulted in a significant growth reduction of MCF-7
cells in dose- and time-dependent manners, which was detectable
during early and late phases of cell growth. In MCF-7 cells,
lipotropes (20 times) caused 83.6% growth reduction at 24 h after
treatment, compared to the control. The dose of lipotropes (20
times) for maximum cancer cell growth inhibition was used for
subsequent experiments.
Lipotropes significantly increase
caspase-3 and p53 activities in MCF-7 cells
In order to determine if lipotropes inhibit the
growth of MCF-7 cells by inducing cell death, caspase-3 and p53
activities were measured using colorimetric assays at 96 h after
treatment. In MCF-7 cells, lipotropes (20 times) increased both
caspase-3 and p53 activity. At 96 h after treatment, lipotropes
increased the caspase-3 activity in MCF-7 cells (10.5%; Fig. 1B). Consistent results were obtained
from the p53 assay. At 96 h, p53 activity was significantly
upregulated in MCF-7 cells (14.8%; Fig.
1B). These results suggest that lipotropes induce apoptosis in
MCF-7 cells, at least partially by modulating caspase-3 and p53
activities.
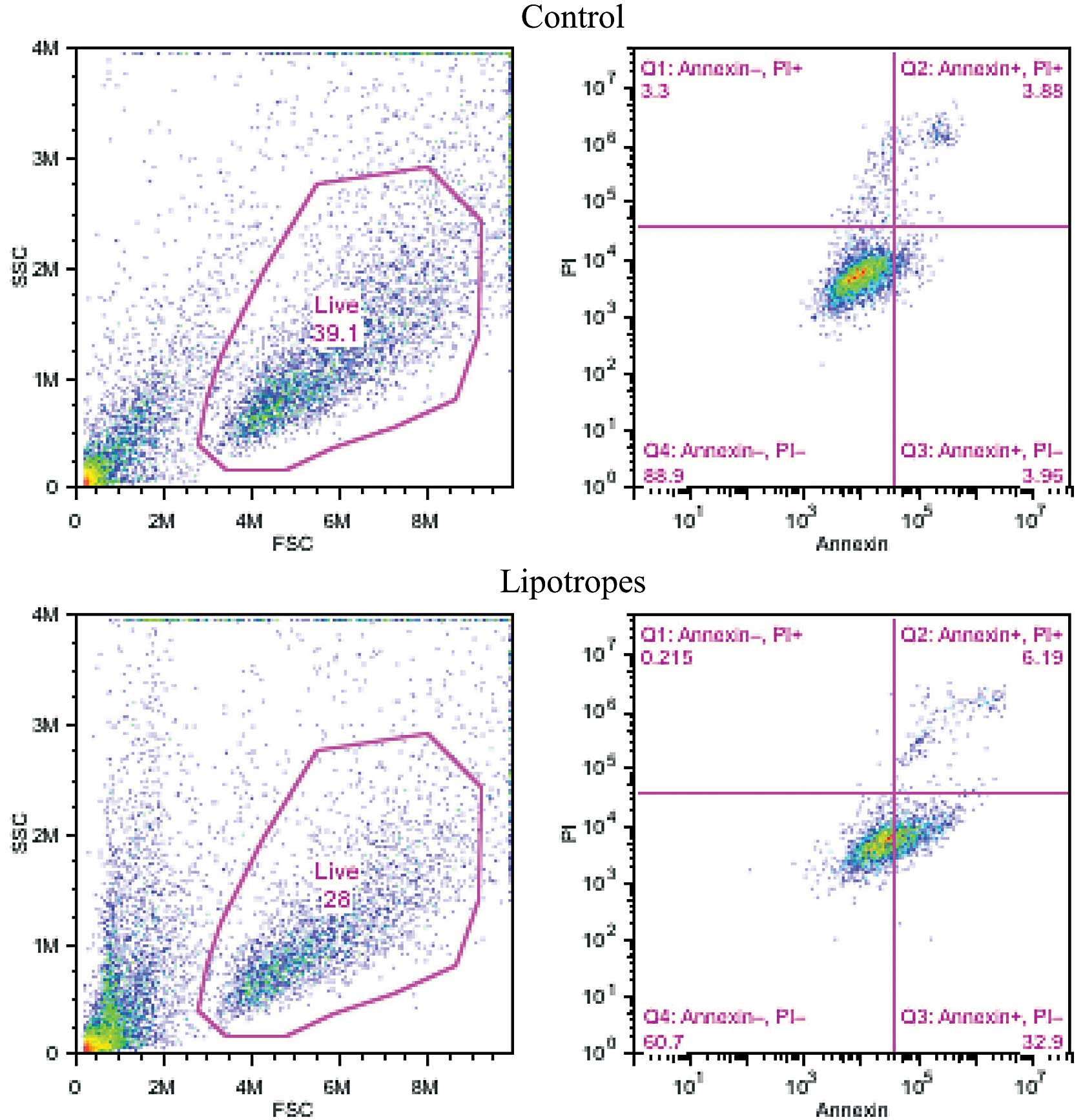
Lipotropes induce apoptosis in MCF-7
cells
Apoptosis was measured using a flow cytometric assay
with Annexin V and PI at 96 h after treatment. As shown in Fig. 2, lipotropes (20 times) increased
apoptosis (upper- and lower-right quadrants), +31.3% in MCF-7 cells
(7.8 vs. 39.1%). Moreover, lipotropes showed a decreased percentage
of live cells, −11.1 in MCF-7 cells (39.1 vs. 28.0%) as shown in
Fig. 2 (left panels). These results
indicate that lipotropes induce apoptotic cell death in the early
(lower-right quadrant) and late (upper-right quadrant) apoptosis
stage of MCF-7 cancer cells.
Expression of genes associated with
apoptosis, cancer and drug resistance in MCF-7 cells
The mRNA levels of apoptosis- and cancer-related
genes in MCF-7 cells were analyzed by quantitative real-time PCR.
As shown in Table II, lipotropes
did not affect the expression of MDM2, ESR1,
ABCC1 and ABCG2 genes in MCF-7 cells treated with 20
times lipotropes for 96 h. However, lipotropes significantly
increased p53 gene expression (41.1%) in MCF-7 cells. The
p53 data supports the results of the caspase-3, p53, and flow
cytometric assays and also indicates that lipotropes reduce cancer
cell growth by inducing apoptosis.
| Table IIExpression of apoptosis- and
cancer-related genes in MCF-7 cells. |
Table II
Expression of apoptosis- and
cancer-related genes in MCF-7 cells.
| Gene symbol | Treatment |
Fold-difference | P-value |
|---|
|
|---|
| Control | Lipotropes |
|---|
| p53a | 0.88±0.35 | 1.24±0.15 | 1.41 | 0.03d |
| MDM2a | 1.12±0.15 | 0.94±0.29 | 0.84 | 0.11 |
| ESR1b | 1.07±0.10 | 1.00±0.39 | 0.93 | 0.72 |
|
ABCC1c | 1.16±0.16 | 0.93±0.36 | 0.80 | 0.14 |
|
ABCG2c | 1.22±0.48 | 1.00±0.43 | 0.82 | 0.34 |
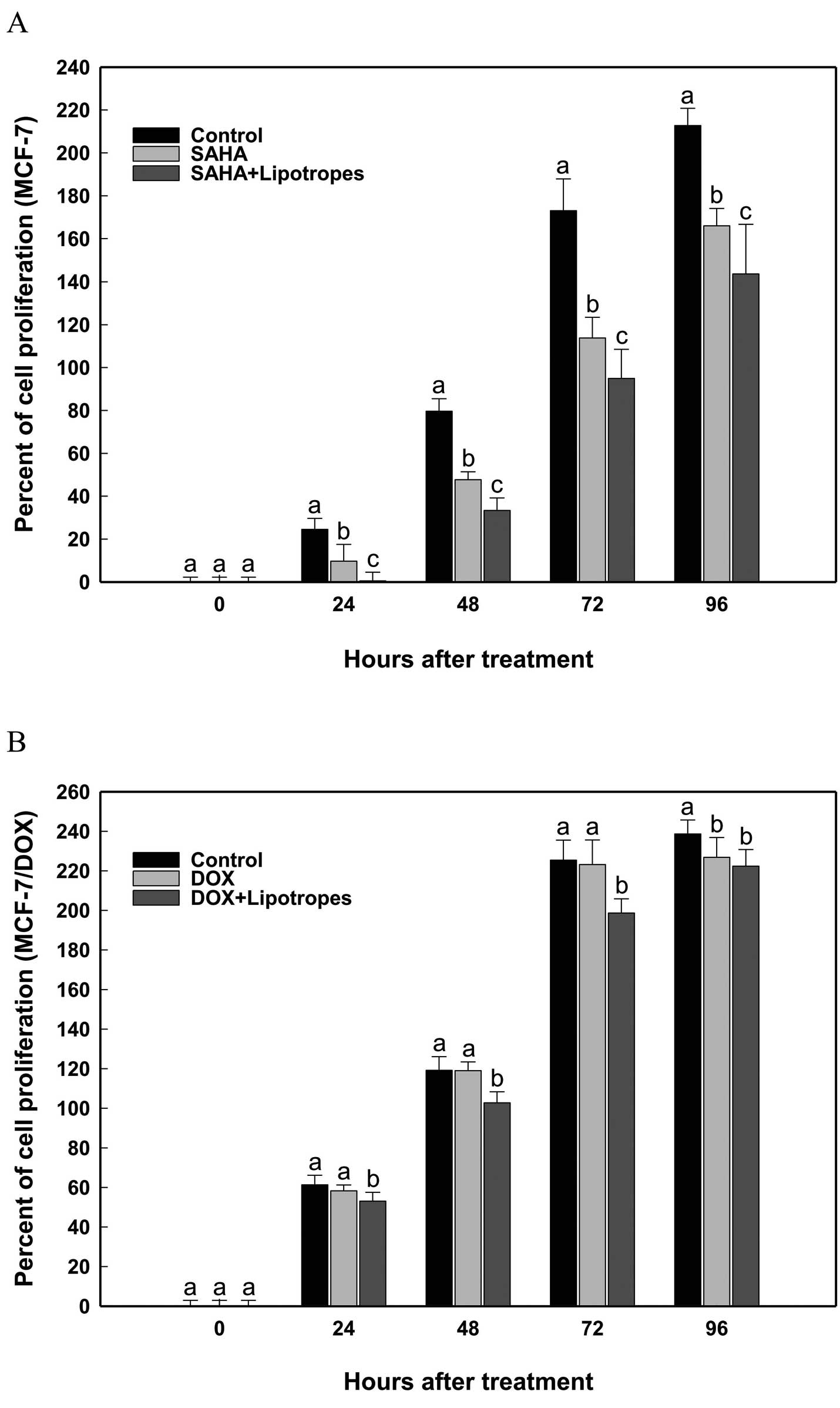
Lipotropes show additive effects with
HDAC inhibitor SAHA and anticancer drug DOX in the growth
inhibition of MCF-7 cells
Although no significant differences in the
expression of drug resistance genes (ABCC1 and ABCG2)
were found, the possible synergistic or additive effect of
lipotropes in combination with chemotherapeutic agents was
investigated. The HDAC inhibitor SAHA and anticancer drug DOX were
assessed in combination with lipotropes (20 times). As shown in
Fig. 3, lipotropes showed
significant additive effect with SAHA in MCF-7 cells (Fig. 3A) and inhibitory effect with DOX in
MCF-7/DOX cells (Fig. 3B). This
indicates that the observed in vitro effects of lipotropes
might reflect a response towards apoptosis rather than modifying
chemotherapeutic drug effects.
Discussion
Breast cancer is the most common malignancy in women
in the United States, accounting for approximately 40,000 deaths
each year (18). Nearly one out of
every eight women develops breast cancer during her lifetime
(19). The incidence of breast
cancer is associated with lifestyle, and nutrition is one of the
most important factors influencing the risk of breast cancer
(18). In January 1998, the US Food
and Drug Administration mandated the folate fortification of all
enriched cereal-grain products in the United States (20). This policy was intended to ensure
sufficient folate intake among women of childbearing age in order
to decrease the risk of neural tube defects in their babies
(20). Some case-control studies
have demonstrated that increased intake of methyl nutrients may
reduce the risk of breast cancer, while low intake of methyl
nutrients is associated with an elevated risk of breast cancer
(7,21). In mammals, methyl metabolism is
dependent on dietary methyl nutrients which influence DNA and
histone methylation of genes (8).
Lipotropes significantly inhibited MCF-7 human
breast cancer cell growth (Fig.
1A). Lipotropes also showed significantly increased caspase-3
and p53 levels in MCF-7 cells (Fig.
1B). These findings show that in MCF-7 cells treated with
lipotropes, increased activities of caspase-3 and p53 correlate
with MCF-7 cell growth inhibition, possibly due to apoptosis.
Similarly, the flow cytometric result showed increased apoptosis,
confirmed by Annexin V and PI (Fig.
2). Apoptosis has been accepted as a fundamental process in the
elimination of defective or unwanted cells in multicellular
organisms (22), and caspases are
the important components in the initiation and execution of
apoptosis, with caspase-3 being a key mediator of apoptosis
(10). In order to suppress cancer,
the p53 tumor suppressor protein induces or represses a multitude
of genes encoding proteins involved in key processes, including
cell cycle control, DNA repair, senescence, and apoptosis (23,24).
Regulating the transcriptional activity of p53 in cancer cells is
one of the main strategies to increase the susceptibility of cancer
cells to apoptosis in cancer treatment (9).
Estrogen receptor (ER) plays a role in breast cancer
development and the abundance of this receptor tends to increase in
breast cancer (25). The
development of drug resistance to a variety of chemotherapeutic
agents is one of the major obstacles for cancer treatment (14). Drug resistance often results from
the overexpression of members of the ATP-binding cassette (ABC)
transporter family, such as ATP-binding cassette sub-family C
member 1 (ABCC1) and ATP-binding cassette sub-family G member 2
(ABCG2) (26–28). Lipotropes significantly increased
p53 gene expression, but the expression of MDM2,
ESR1, ABCC1 and ABCG2 genes showed similar
levels between two groups (Table
II). Collectively, these results suggest that lipotropes reduce
MCF-7 cell growth in relation to p53-dependent apoptosis through
transcriptional and translational control.
Lipotropes enhanced the tumor-suppressive effects of
the histone deacetylase (HDAC) inhibitor suberoylanilide hydroxamic
acid (SAHA) and the anticancer drug doxorubicin (DOX) on MCF-7 cell
growth (Fig. 3). HDAC inhibitors
are emerging anticancer drugs and SAHA is a potent HDAC inhibitor,
causing growth arrest and apoptosis in several types of cancer
(13). DNA methylation correlates
with epigenetic regulation of oncogenes and tumor suppressor genes
(29). Methyl CpG binding protein 2
(MeCP2) selectively recognizes methylated CpG dinucleotides and
forms a complex with HDAC, thereby suppressing gene transcription
leading to chromatin remodeling (30). Some studies have shown that,
compared to normal cells, MeCP2 and HDAC1 gene
expression is elevated in cancer cells (31).
The present study demonstrated that lipotropes
induce apoptosis and show anti-proliferative effects in combination
with SAHA and DOX in MCF-7 cells. The findings of the present study
provide insight for understanding dietary intervention of methyl
nutrients in cancer cell growth and apoptosis, and yield useful
information in designing the in vivo and in vitro
experiments to reduce breast cancer risk.
Acknowledgements
The authors thank Dr Elena V. Batrakova (University
of Nebraska) for kindly providing the MCF-7/DOX cell line and Dr
Raushan K. Singh for technical assistance. The present study was
partially supported by the US DODCDMRP/BCRP (grant
W81XWH-09-1-0610).
Abbreviations:
|
ABCC1
|
ATP-binding cassette sub-family C
member 1
|
|
ABCG2
|
ATP-binding cassette sub-family G
member 2
|
|
ACTB
|
β-actin
|
|
CpG
|
cytosine-phosphate-guanine
dinucleotide
|
|
DOX
|
doxorubicin
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
ER
|
estrogen receptor
|
|
ESR1
|
estrogen receptor 1 gene
|
|
HDAC
|
histone deacetylase
|
|
MDM
|
murine double minute oncogene
|
|
MeCP2
|
methyl CpG binding protein 2
|
|
p53
|
tumor protein 53
|
|
RT-PCR
|
reverse transcription-polymerase chain
reaction
|
|
SAHA
|
suberoylanilide hydroxamic acid
|
References
|
1
|
Cho K, Mabasa L, Bae S, Walters MW and
Park CS: Maternal high methyl diet suppresses mammary
carcinogenesis in female rat offspring. Carcinogenesis.
33:1106–1112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zeisel SH: Epigenetic mechanisms for
nutrition determinants of later health outcomes. Am J Clin Nutr.
89:1488S–1493S. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Van den Veyver IB: Genetic effects of
methylation diets. Annu Rev Nutr. 22:255–282. 2002.PubMed/NCBI
|
|
4
|
Zhang SM, Willett WC, Selhub J, Hunter DJ,
Giovannucci EL, Holmes MD, Colditz GA and Hankinson SE: Plasma
folate, vitamin B6, vitamin B12,
homocysteine, and risk of breast cancer. J Natl Cancer Inst.
95:373–380. 2003.PubMed/NCBI
|
|
5
|
Beilby J, Ingram D, Hähnel R and Rossi E:
Reduced breast cancer risk with increasing serum folate in a
case-control study of the C677T genotype of the
methylenetetrahydrofolate reductase gene. Eur J Cancer.
40:1250–1254. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Park CS, Cho K, Bae DR, Joo NE, Kim HH,
Mabasa L and Fowler AW: Methyl-donor nutrients inhibit breast
cancer cell growth. In Vitro Cell Dev Biol Anim. 44:268–272. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Newberne PM: Lipotropic factors and
oncogenesis. Adv Exp Med Biol. 206:223–251. 1986.PubMed/NCBI
|
|
8
|
Mason JB: Biomarkers of nutrient exposure
and status in one-carbon (methyl) metabolism. J Nutr.
133:941S–947S. 2003.PubMed/NCBI
|
|
9
|
Call JA, Eckhardt SG and Camidge DR:
Targeted manipulation of apoptosis in cancer treatment. Lancet
Oncol. 9:1002–1011. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Riedl SJ and Shi Y: Molecular mechanisms
of caspase regulation during apoptosis. Nat Rev Mol Cell Biol.
5:897–907. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kussie PH, Gorina S, Marechal V, Elenbaas
B, Moreau J, Levine AJ and Pavletich NP: Structure of the MDM2
oncoprotein bound to the p53 tumor suppressor transactivation
domain. Science. 274:948–953. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eischen CM and Lozano G: p53 and MDM2:
antagonists or partners in crime? Cancer Cell. 15:161–162. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Marks PA, Richon VM, Miller T and Kelly
WK: Histone deacetylase inhibitors. Adv Cancer Res. 91:137–168.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Linn SC, Pinedo HM, van Ark-Otte J, van
der Valk P, Hoekman K, Honkoop AH, Vermorken JB and Giaccone G:
Expression of drug resistance proteins in breast cancer, in
relation to chemotherapy. Int J Cancer. 71:787–795. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eliopoulos AG, Kerr DJ, Herod J, Hodgkins
L, Krajewski S, Reed JC and Young LS: The control of apoptosis and
drug resistance in ovarian cancer: influence of p53 and Bcl-2.
Oncogene. 11:1217–1228. 1995.PubMed/NCBI
|
|
16
|
Cho K, Mabasa L, Fowler AW, Walsh DM and
Park CS: Canola oil inhibits breast cancer cell growth in cultures
and in vivo and acts synergistically with chemotherapeutic drugs.
Lipids. 45:777–784. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Singal PK and Iliskovic N:
Doxorubicin-induced cardiomyopathy. N Engl J Med. 13:900–905. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
19
|
Feuer EJ, Wun LM, Boring CC, Flanders WD,
Timmel MJ and Tong T: The lifetime risk of developing breast
cancer. J Natl Cancer Inst. 85:892–897. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Honein MA, Paulozzi LJ, Mathews TJ,
Erickson JD and Wong LY: Impact of folic acid fortification of the
US food supply on the occurrence of neural tube defects. JAMA.
285:2981–2986. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Das PM and Singal R: DNA methylation and
cancer. J Clin Oncol. 22:4632–4642. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Khan N, Adhami VM and Mukhtar H: Apoptosis
by dietary agents for prevention and treatment of cancer. Biochem
Pharmacol. 76:1333–1339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oren M: Decision making by p53: life,
death and cancer. Cell Death Differ. 10:431–442. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Meek DW: Tumour suppression by p53: a role
for the DNA damage response? Nat Rev Cancer. 9:714–723.
2009.PubMed/NCBI
|
|
25
|
Lapidus RG, Nass SJ and Davidson NE: The
loss of estrogen and progesterone receptor gene expression in human
breast cancer. J Mammary Gland Biol Neoplasia. 3:85–94. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pohl A, Devaux PF and Herrmann A: Function
of prokaryotic and eukaryotic ABC proteins in lipid transport.
Biochim Biophys Acta. 1733:29–52. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kaminski WE, Piehler A and Wenzel JJ: ABC
A-subfamily transporters: structure, function and disease. Biochim
Biophys Acta. 1762:510–524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Park S, Shimizu C, Shimoyama T, Takeda M,
Ando M, Kohno T, Katsumata N, Kang Y, Nishio K and Fujiwara Y: Gene
expression profiling of ATP-binding cassette (ABC) transporters as
a predictor of the pathologic response to neoadjuvant chemotherapy
in breast cancer patients. Breast Cancer Res Treat. 99:9–17. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Razin A: CpG methylation, chromatin
structure and gene silencing-a three-way connection. EMBO J.
17:4905–4908. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fuks F, Hurd PJ, Wolf D, Nan X, Bird AP
and Kouzarides T: The methyl-CpG-binding protein MeCP2 links DNA
methylation to histone methylation. J Biol Chem. 278:4035–4040.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Müller HM, Fiegl H, Goebel G, Hubalek MM,
Widschwendter A, Müller-Holzner E, Marth C and Widschwendter M:
MeCP2 and MBD2 expression in human neoplastic and non-neoplastic
breast tissue and its association with oestrogen receptor status.
Br J Cancer. 89:1934–1939. 2003.PubMed/NCBI
|