Introduction
Epithelial ovarian cancer is a major cause of
mortality among women with gynecological malignancies due to its
late symptomatology (1); it is
often associated with distant metastases and has a poor prognosis
(2). Chemotherapy is essential for
the management of ovarian cancer, but the efficacy of this
treatment is limited by chemoresistance. Paclitaxel is a first-line
chemotherapeutic agent that is effective in epithelial ovarian
cancer by stabilizing microtubules, inducing cell cycle arrest in
the G2-M phase, and activating pro-apoptotic signaling (2,3). In
spite of the comparatively high sensitivity of epithelial ovarian
cancer to paclitaxel, the prognosis of advanced or recurrent cases
remains poor since mortality is mostly the result of metastasis
that is refractory to conventional chemotherapy (2).
The epithelial-mesenchymal transition (EMT) in
cancer cells has been shown to promote metastasis and
chemoresistance (4). During the
acquisition of EMT characteristics, the hallmark of EMT is loss of
the epithelial molecule E-cadherin and gain of mesenchymal markers
including N-cadherin and vimentin, leading to remodeling of the
cytoskeleton and enhancement of cancer cell migration and invasion
(5). An EMT phenotype has been
reported in gemcitabine-resistant pancreatic cancer cells (6), gefitinib-resistant non-small cell lung
cancer and oxaliplatin-resistant colorectal cancer cells (7), paclitaxel-resistant ovarian cancer
cells (2) and tamoxifen-resistant
breast cancer cells (8). Blockade
of the EMT pathways is critical for preventing cancer cell
migration and invasion as well as for restoring drug sensitivity
(4). Although the EMT in cancer
cells integrates multiple signaling pathways, such as TGFβ, NF-κB,
Wnt and Hedgehog (9–12), the details in A2780 human ovarian
cancer cells remain to be identified.
We are the first to report that phosphatidylinositol
3-kinase (PI3K) inhibition reverses paclitaxel resistance-induced
EMT in A2780 human ovarian cancer cells, indicating that the PI3K
pathway is a promising therapeutic target against ovarian cancer
with paclitaxel chemotherapy.
Materials and methods
Cells and reagents
A2780/WT and A2780/PTX cell lines were obtained from
KeyGen Biotech Co. Ltd. (Nanjing, China) and maintained in
RPMI-1640 supplemented with 10% fetal bovine serum (FBS) and
penicillin-streptomycin (100 U/ml penicillin and 100 μg/ml
streptomycin) at 37°C in a humidified atmosphere of 5%
CO2. Paclitaxel was from Qilu Pharmaceutical Co, Ltd.
(Jinan, China). LY294002 was from Selleckchem (Burlington, NC,
USA). The primers were synthesized by Sangon Biotechnology
(Shanghai, China).
Reverse transcription (RT)-PCR
Total RNA was extracted using the TRIzol protocol
and cDNA was synthesized from the mRNA using the SuperScript
First-Strand Synthesis System (Invitrogen Life Technologies,
Carlsbad, CA, USA) for RT-PCR. PCR was performed for 35 cycles of
95°C for 30 sec, 58°C for 30 sec and 72°C for 45 sec. The data were
analyzed with ImageJ software (NIH, Bethesda, MD, USA). β-actin was
used as endogenous control. The primer sequences are available upon
request.
Migration assay
Cell migration was studied by the modified Boyden
chamber method in 24-well plates, each containing 6.5-mm Transwell
chambers with 8-mm pores (BD Biosciences, Franklin Lakes, NJ, USA).
Cells (1×106 cells/ml) were harvested, washed twice with
PBS and responded in serum-free RPMI-1640. The cell suspension (100
μl) was seeded into the upper chamber of each Transwell unit. The
bottom wells contained RPMI-1640 supplemented with 10% FBS to
create a chemotactic gradient. During certain treatments, PI3K
inhibitor LY294002 (10 μM) was added. After 24 h of stimulation,
the cells that had not migrated through the filter in the Transwell
inserts (on the upper surface) were removed with a cotton swab.
Cells that had migrated to the lower surface of the filter were
stained with crystal violet and counted under a microscope. Images
were captured by a video camera (Nikon Coolpix 54, Japan) mounted
on the microscope (Leica CME, Japan). Cells were counted in four
random fields per insert.
Wound-healing assay
Cell motility was assessed by measuring the ability
of the cells to migrate into a wound made in a confluent monolayer.
Briefly, wild-type A2780/WT and A2780/PTX cells were plated at
2×105 cells/well in 6-well plates. After overnight
growth and attachment, both media were replaced with serum-free
media for another 24 h. The monolayers were scratched using a
sterile pipette tip; floating cells were washed off with PBS. The
cells were incubated in 2% FBS medium with or without LY294002 (10
μM), The ability of cells to migrate into the wound gap was
observed and recorded photographically. Wound closure was analyzed
as the ratio of the remaining wound area relative to the initial
wound area with ImageJ software (NIH). Experiments were repeated at
least 3 times.
In vitro cell proliferation assay
Serial dilutions of cells in culture medium were
prepared, and 100 μl of the suspensions containing
1×104, 5×103 and 1×103 cells were
each added into 6-wells of 96-well plates. Cells were incubated for
48 h, then 10 μl MTT (5 mg/ml) (Sigma-Aldrich, St. Louis, MO, USA)
was added to form formazan crystals, which were dissolved in 150 μl
dimethyl sulfoxide (DMSO). Absorbance was measured at 490 nm using
a plate reader (Thermo Labsystems Multiskan MK3; Thermo Scientific,
Rockford, IL, USA). The values from triplicate readings were
averaged.
Paclitaxel chemosensitivity assay
The cells were seeded at 8,000 cells/well
(triplicates) in 96-well plates. After 24 h, they were treated with
paclitaxel (1.5–1,500 μg/ml in 2-fold serial dilutions) for 48 h.
After the treatment, MTS (20 μl; Promega Corporation, Madison, WI,
USA) was added to each well and the cells were incubated at 37°C
with 5% CO2 for 4 h. Absorbance at 590 nm was then
measured with a microplate reader (Thermo Synergie HT, USA).
Immunofluorescence staining
The cells were fixed for 30 min in 4%
paraformaldehyde, permeabilized with 0.1% Triton X-100 for 10 min
at room temperature, respectively. Cells were washed 3 times in PBS
after each treatment. They were then blocked with 2% BSA in PBS for
30 min at room temperature, incubated with primary antibody for
β-tubulin (20 μg/ml) diluted in 2% BSA/PBS overnight at 4°C, and
then incubated with fluorescent secondary antibody for 1 h at room
temperature. Images were acquired with a confocal laser scanning
microscope (LSM 710; Carl Zeiss, Jena, Germany). If required, cells
were pretreated with LY294002 (10 μM, 24 h).
Statistical analysis
Results are presented as the means ± SD. Statistical
differences were determined by Student’s t-test. A value of
p<0.05 was considered to indicate a statistically significant
difference.
Results
Acquisition of paclitaxel resistance in
A2780/PTX cells induces morphological and molecular changes
consistent with EMT signaling
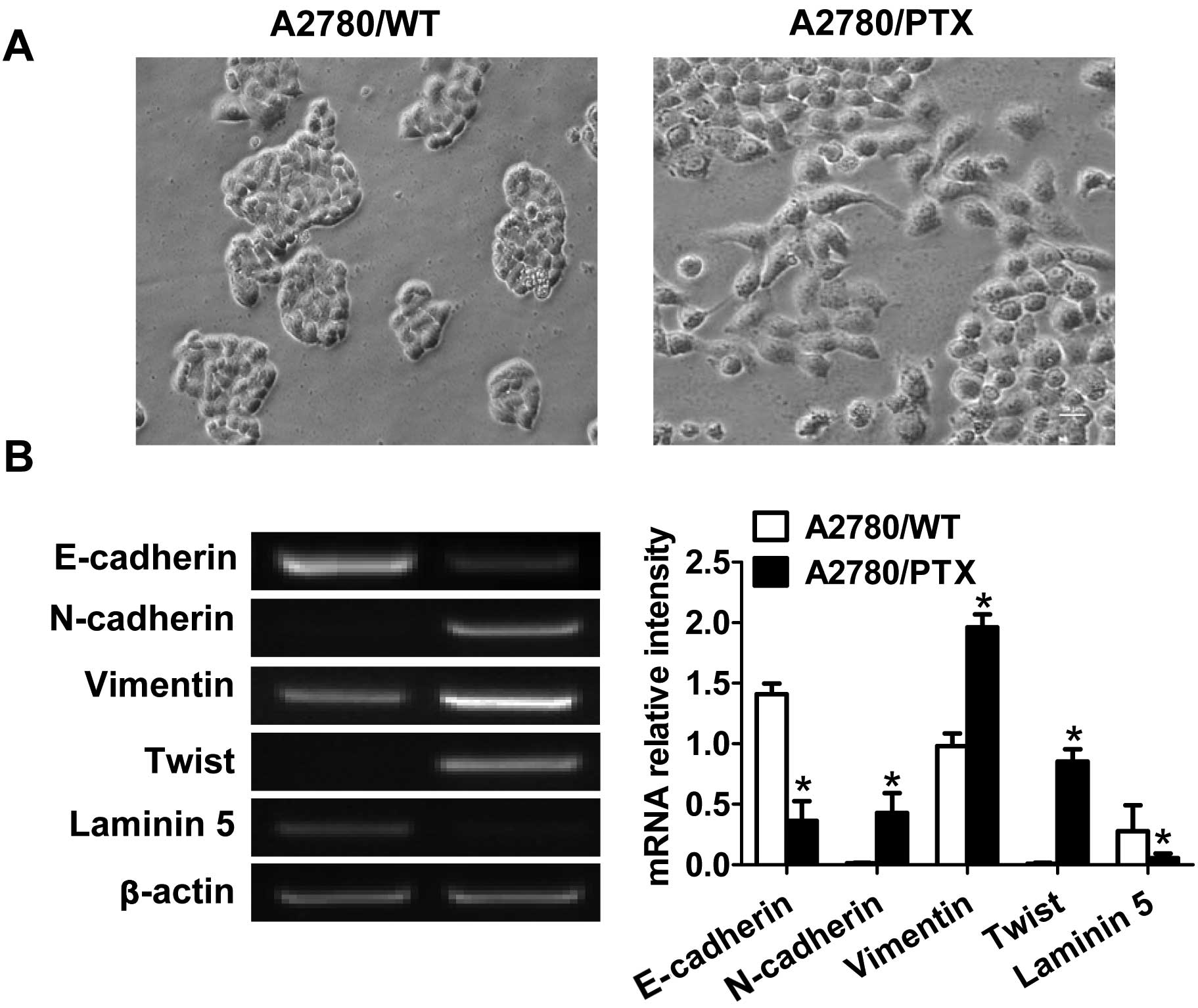
We first examined the morphological characteristics
of the cell lines during exponential growth. A2780/WT cells formed
cohesive clusters with round cellular morphology in vitro
(Fig. 1A), consistent with an
epithelial phenotype. By contrast, A2780/PTX cells, grown in the
presence of 100 nM PTX, displayed a spindle-like morphology and
formed dyscohesive sheets (Fig.
1A).
To determine whether the acquisition of paclitaxel
resistance induced specific molecular changes consistent with EMT,
we investigated EMT-related biomarkers. Compared with the wild-type
cells, the A2780/PTX cells showed significant reduction in
E-cadherin and laminin-5 expression and upregulation of N-cadherin,
vimentin and Twist (Fig. 1B).
A2780/PTX cells display increased
potential for proliferation and migration
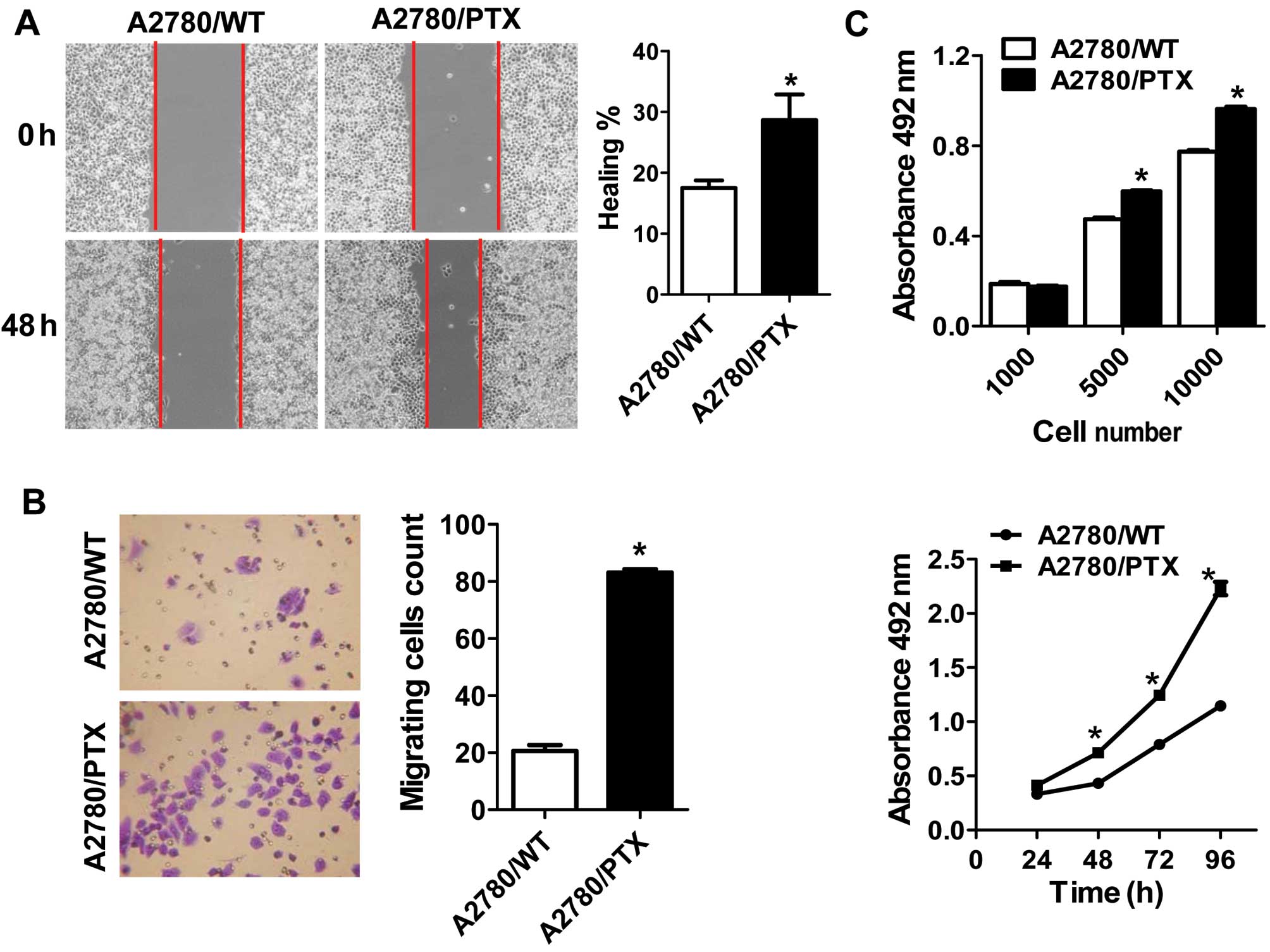
We performed wound healing and Transwell migration
assays to compare the migratory potential between the 2 cell lines.
The capacity for wound healing in A2780/PTX cells was significantly
enhanced (Fig. 2A), and the
relative numbers of migrating and invasive A2780/PTX cells were
significantly superior to wild-type cells (Fig. 2B).
We further investigated the proliferative potential
of A2780/WT and A2780/PTX cells at various times after plating and
with various cell numbers at 48 h by assessing cell viability using
a modified MTT assay. The MTT assay with various numbers showed a
decrease in the growth of A2780/WT cells compared with A2780/PTX
cells. In addition, similar results were found in the proliferative
potential at various times (Fig.
2C).
PI3K signaling pathway is important for
the induction of cell migration and proliferation
Recently, activation of the PI3K/AKT axis has been
shown to be a central feature of EMT (13–16).
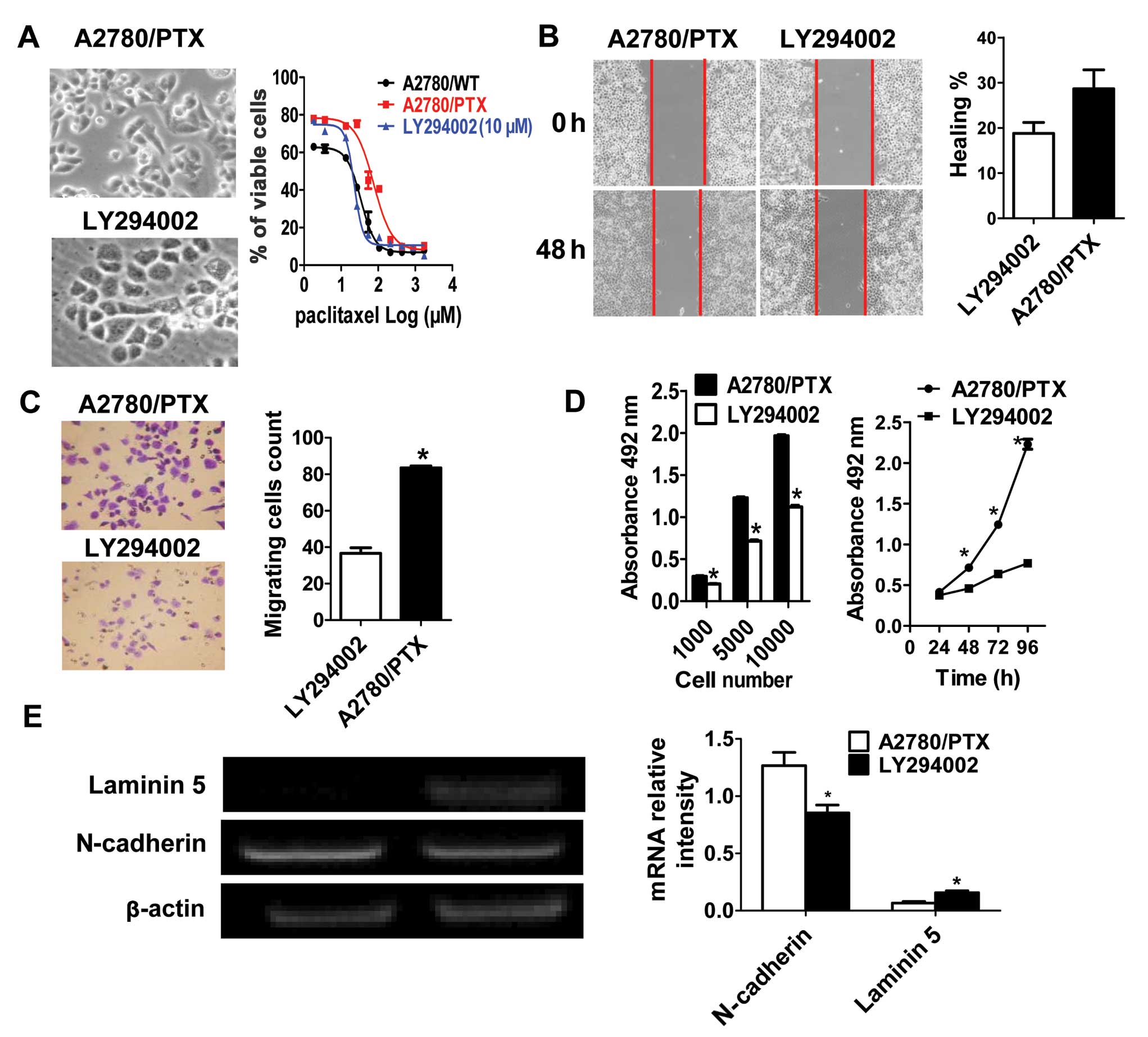
To assess the involvement of PI3K signaling, the PI3K inhibitor
LY294002 (10 μM) was applied to A2780/PTX cells, and they exhibited
well-organized cell-cell association, appeared cohesive, and
exhibited classical epithelial morphology (Fig. 3A). The expression of epithelial
marker laminin 5 was significantly enhanced, while the levels of
mesenchymal marker N-cadherin decreased (Fig. 3E). Inhibition of PI3K significantly
decreased migration and proliferation in A2780/PTX cells (Fig. 3B–D). These results showed that
inhibition of PI3K signaling in A2780/PTX cells led to reversal of
the EMT, indicating the involvement of the PI3K signaling pathway
in the chemoresistance process.
Inhibition of the PI3K activity increases
sensitivity of A2780/PTX cells to paclitaxel
We next explored whether inhibiting PI3K reverses
the resistance to paclitaxel in A2780/PTX cells. MTT assays showed
that A2780/PTX cells treated with LY294002 restored the sensitivity
to paclitaxel-induced cell death (Fig.
3A). The IC50 was reduced to half of that in
untreated A2780/PTX cells.
Inhibition of the PI3K activity alters
the cytoskeleton of A2780/PTX cells
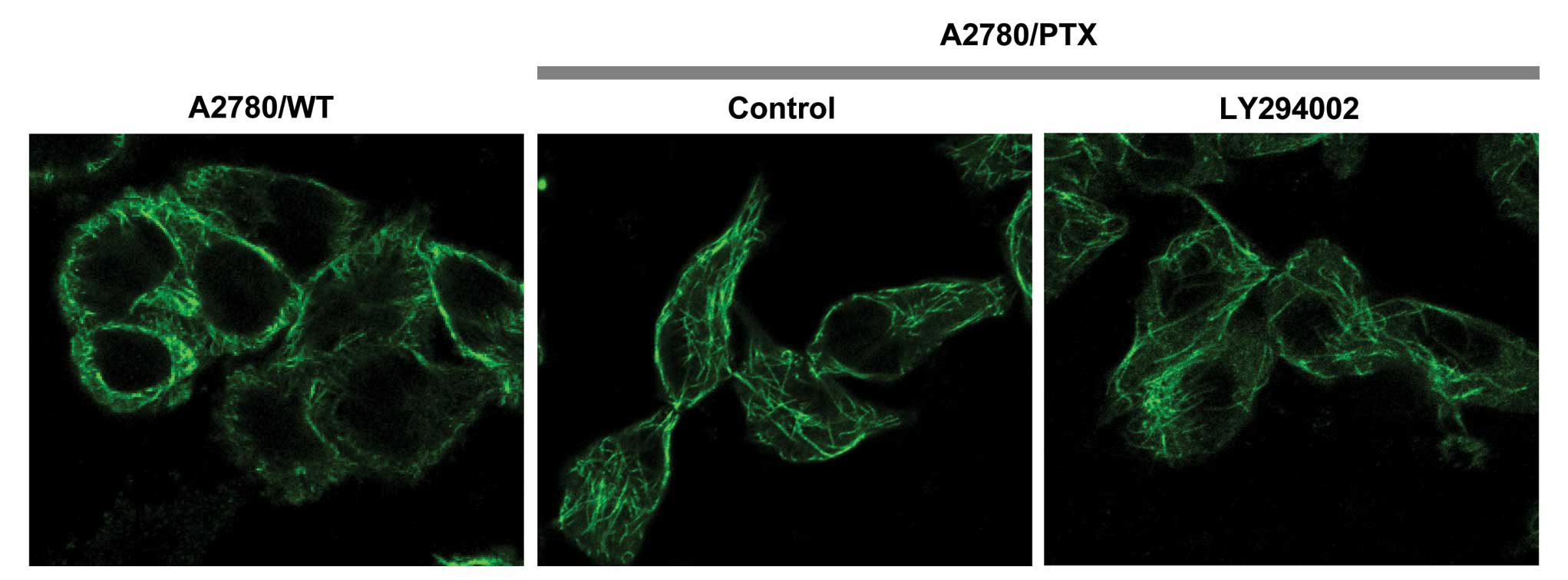
The anticancer drug paclitaxel binds to β-tubulin in
assembled microtubules and causes cytoskeleton changes in cells
(17,18). We further investigated organization
of the cytoskeleton in A2780/PTX cells. A2780/PTX cells had
spindle-like morphology or filopodia formation, in which brightly
stained longitudinal β-tubulin bundles were detected, whereas cells
in the wild-type and LY294002-treated groups displayed polygonal
cell shapes (Fig. 4).
Discussion
Chemoresistance remains the major therapeutic
barrier in epithelial ovarian cancer (4,19). A
thorough understanding of the mechanisms is essential for improving
therapeutic agents. Our study used wild-type (A2780/WT) and
paclitaxel-resistant human epithelial ovarian cancer cells
(A2780/PTX) to investigate the molecular mechanism of paclitaxel
resistance and associated cellular behaviors.
In the present study, we demonstrated that A2780/PTX
cells underwent EMT processes. This was confirmed as follows: (i)
morphological change from cobblestone-like to spindle-shaped cells;
(ii) increased potential for proliferation and migration; and (iii)
changes in molecular markers (significant reduction in E-cadherin
and laminin-5 expression and upregulation of N-cadherin and
vimentin). In agreement with our results, some reports have shown
that the EMT is induced by other chemotherapeutic agents in
different cancer cell lines (20–22).
Moreover, several studies have demonstrated that the EMT plays an
important role in functional malignancy behaviors including
facilitating the transformation, initiation, progression and
metastasis of human cancer (23,24).
We further examined the proliferation in these 2 cell types. As
expected, our findings showed that A2780/PTX cells displayed
enhanced multiplication in the MTT assay. Overall, we obtained
evidence that paclitaxel resistance and facilitated malignant
potential are linked with the EMT.
Since the PI3K/AKT axis is frequently activated in
human cancer (15), we focused on
the EMT with particular emphasis on the PI3K/AKT pathways involved
in the regulation of malignancy. To determine whether the PI3K
pathway was involved in the process, the PI3K inhibitor LY294002
was applied to A2780/PTX cells, resulting in inhibition of
fibroblastic morphology, proliferation and motility. Our evidence
seems to be associated with other research in gefitinib-resistant
cells from a head and neck squamous cell carcinoma cell line
(22). Furthermore, we found that
the inhibition of PI3K activity attenuated the paclitaxel
resistance and led to loss of cell polarity in terms of β-tubulin
distribution suggesting that there should also be defects in cell
migration and invasion. Although further studies are required to
elucidate the chemoresistance mechanisms in PI3K-mediated EMT, the
use of PI3K inhibitors during chemotherapy could be a useful
approach to improve its efficacy.
In conclusion, we have shown that the development of
paclitaxel resistance in epithelial ovarian cancer cells is
accompanied by inducible EMT-like changes with increased potential
to metastasize. Targeting with the PI3K inhibitor LY294002 may
sensitize cancer cells to chemotherapeutics and reduce motility.
The clinical significance of the PI3K signal in acquired resistance
to paclitaxel merits further investigation.
Acknowledgements
This study was supported by the Program for New
Century Excellent Talents in University of The Ministry of
Education of China (to X.M.); Fundamental Research Funds for the
Central Universities JUSRP51311A (to X.M.); China National Natural
Science Foundation grants 81100185 (to X.M.), 81130057 (to J.J.),
81201600 (to D.H.) and 81101667 (to R.Z.); Jiangsu Province
National Natural Science Foundation grant BK2010161 (to D.H.); and
a ‘Strategic Priority Research Program’ grant from the Chinese
Academy of Sciences XDA01040000 (to J.J.).
References
|
1
|
Vergara D, Merlot B, Lucot JP, et al:
Epithelial-mesenchymal transition in ovarian cancer. Cancer Lett.
291:59–66. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kajiyama H, Shibata K, Terauchi M, et al:
Chemoresistance to paclitaxel induces epithelial-mesenchymal
transition and enhances metastatic potential for epithelial ovarian
carcinoma cells. Int J Oncol. 31:277–283. 2007.
|
|
3
|
Jia L, Zhang S, Ye Y, et al: Paclitaxel
inhibits ovarian tumor growth by inducing epithelial cancer cells
to benign fibroblast-like cells. Cancer Lett. 326:176–182. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rosano L, Cianfrocca R, Spinella F, et al:
Acquisition of chemoresistance and EMT phenotype is linked with
activation of the endothelin A receptor pathway in ovarian
carcinoma cells. Clin Cancer Res. 17:2350–2360. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shah AN, Summy JM, Zhang J, Park SI,
Parikh NU and Gallick GE: Development and characterization of
gemcitabine-resistant pancreatic tumor cells. Ann Surg Oncol.
14:3629–3637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rho JK, Choi YJ, Lee JK, et al: Epithelial
to mesenchymal transition derived from repeated exposure to
gefitinib determines the sensitivity to EGFR inhibitors in A549, a
non-small cell lung cancer cell line. Lung Cancer. 63:219–226.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hiscox S, Jiang WG, Obermeier K, et al:
Tamoxifen resistance in MCF7 cells promotes EMT-like behaviour and
involves modulation of β-catenin phosphorylation. Int J Cancer.
118:290–301. 2006.PubMed/NCBI
|
|
9
|
Zhou Q, Zeng R, Xu C, et al: Erbin
inhibits TGF-β1-induced EMT in renal tubular epithelial cells
through an ERK-dependent pathway. J Mol Med (Berl). 90:563–574.
2012.
|
|
10
|
Conti B, Minutolo A, Arciello M and
Balsano C: Are Hedgehog and Wnt/β-catenin pathways involved in
hepatitis C virus-mediated EMT? J Hepatol. 58:636–637. 2012.
|
|
11
|
Radisky DC and Bissell MJ: NF-κB links
oestrogen receptor signalling and EMT. Nat Cell Biol. 9:361–363.
2007.
|
|
12
|
Xu X, Zhou Y, Xie C, et al: Genome-wide
screening reveals an EMT molecular network mediated by Sonic
Hedgehog-Gli1 signaling in pancreatic cancer cells. PLoS One.
7:e431192012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim M, Chuong CM and Roy-Burman P: PI3K,
Erk signaling in BMP7-induced epithelial-mesenchymal transition
(EMT) of PC-3 prostate cancer cells in 2- and 3-dimensional
cultures. Horm Cancer. 2:298–309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Y, Maitah MY, Ahmad A, Kong D, Bao B
and Sarkar FH: Targeting the Hedgehog signaling pathway for cancer
therapy. Expert Opin Ther Targets. 16:49–66. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Larue L and Bellacosa A:
Epithelial-mesenchymal transition in development and cancer: role
of phosphatidylinositol 3′ kinase/AKT pathways. Oncogene.
24:7443–7454. 2005.
|
|
16
|
Hennessy BT, Smith DL, Ram PT, Lu Y and
Mills GB: Exploiting the PI3K/AKT pathway for cancer drug
discovery. Nat Rev Drug Discov. 4:988–1004. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang Y, Alcaraz AA and Snyder JP: The
tubulin-bound conformation of paclitaxel: T-taxol vs “PTX-NY”. J
Nat Prod. 72:422–429. 2009.PubMed/NCBI
|
|
18
|
Lee KM, Cao D, Itami A, et al: Class III
β-tubulin, a marker of resistance to paclitaxel, is overexpressed
in pancreatic ductal adenocarcinoma and intraepithelial neoplasia.
Histopathology. 51:539–546. 2007.
|
|
19
|
Poulain L, Lincet H, Duigou F, et al:
Acquisition of chemoresistance in a human ovarian carcinoma cell is
linked to a defect in cell cycle control. Int J Cancer. 78:454–463.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Işeri OD, Kars MD, Arpaci F, Atalay C, Pak
I and Gündüz U: Drug resistant MCF-7 cells exhibit
epithelial-mesenchymal transition gene expression pattern. Biomed
Pharmacother. 65:40–45. 2011.PubMed/NCBI
|
|
21
|
Haslehurst AM, Koti M, Dharsee M, et al:
EMT transcription factors snail and slug directly contribute to
cisplatin resistance in ovarian cancer. BMC Cancer. 12:912012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maseki S, Ijichi K, Tanaka H, et al:
Acquisition of EMT phenotype in the gefitinib-resistant cells of a
head and neck squamous cell carcinoma cell line through
Akt/GSK-3β/snail signalling pathway. Br J Cancer.
106:1196–1204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ahmed N, Abubaker K, Findlay J and Quinn
M: Epithelial mesenchymal transition and cancer stem cell-like
phenotypes facilitate chemoresistance in recurrent ovarian cancer.
Curr Cancer Drug Targets. 10:268–278. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brabletz T, Jung A, Spaderna S, Hlubek F
and Kirchner T: Opinion: migrating cancer stem cells - an
integrated concept of malignant tumour progression. Nat Rev Cancer.
5:744–749. 2005. View
Article : Google Scholar : PubMed/NCBI
|