Introduction
Gastric cancer is the fourth most common cancer and
remains the second most common cause of cancer-related mortality in
the world although its incidence and mortality has declined over
the last half-century (1,2). Surgery is the most common treatment
for gastric cancer. However, gastric cancer is difficult to cure
unless it is found in an early stage (before it has begun to
spread). Unfortunately, most cancer patients present with
metastasis or advanced disease when the diagnosis is made. As a
result, surgical interventions are currently curative in less than
40% of cases (3), and in cases of
metastasis, may only be palliative. Generally, these patients
should receive systemic chemotherapy apart from surgical resection.
Nevertheless, gastric cancer has not been particularly sensitive to
these drugs, and chemotherapy, if used, has usually served to
merely reduce the size of the tumor, relieve symptoms of the
disease and increase survival time.
Metastatic spread of tumor cells often occurs via
the lymphatic vessels (4).
Tumor-associated lymphatic vessels, also referred to as tumor
lymphangiogenesis, act as a conduit by which disseminating tumor
cell access regional lymph nodes and form metastases (5–7). Thus,
tumor lymphangiogenesis plays an important role in promoting tumor
metastasis. Since lymph node metastasis is an early event in the
metastatic process (8), use of
chemopreventive agents to block lymphangiogenesis pathways seems to
be an attractive anticancer treatment strategy.
Rosiglitazone (ROSI), a synthetic peroxisome
proliferator-activated receptor γ (PPARγ) agonist, is a member of
the thiazolidinedione (TZD) agents. These PPARγ ligands were
clinically used as antidiabetic drugs which could attenuate the
insulin resistance associated with obesity, hypertension, and
impaired glucose tolerance in humans (9). Recent studies suggested that ROSI have
significant anticancer effects on various human malignant tumor
cells in vitro, including gastric, breast, colorectal,
bladder, adrenocortical and pancreatic cancer types (10–15).
In vivo, ROSI also exhibited antitumor activities in the
prevention of lung, breast, prostate, and adrenocortical cancer
(16–19). These studies show that ROSI exerts
its anti anticancer effect by inducing cancer cell growth arrest,
differentiation and apoptosis, or by inhibiting neovascularization
of cancer and suppressing tumor migration.
However, the roles of ROSI in tumor
lymphangiogenesis and lymphatic metastasis were unclear. Since
nearly 85% of tumors were accompanied by lymph node metastasis in
gastric cancer patients present with advanced stage (20), in the present study, we investigated
for the first time the effect of ROSI on the growth and
lymphangiogenesis of human gastric cancer transplanted in nude
mice, so as to explore whether ROSI is a potential
anti-lymphangiogenic agent for gastric cancer therapeutic
strategy.
Materials and methods
Cells and cell cultures
Human gastric cancer cell line, SGC-7901, was
obtained from the Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China). SGC-7901 cells were cultured in
RPMI-1640 medium (Gibco-BRL, Carlsbad, CA, USA) containing 10%
fetal bovine serum (FBS) (Hangzhou Sijiqing Biological Engineering
Materials Co., Ltd, China) and 1% antibiotics (100 U/ml penicillin
G, 100 U/ml streptomycin sulfate). Cells were maintained in
logarithmic growth in a humidified atmosphere of 5% CO2
at 37°C. Adherent tumor cells were harvested from subconfluent
cultures by a brief exposure to trypsin [containing 0.25% phosphate
buffered saline (PBS) and 0.05% ethylenediaminetetraacetic acid
(EDTA)], trypsinization was stopped with medium containing 10%
serum, and the cells were washed once and resuspended in serum-free
medium. Trypan blue staining was used to assess cell viability, and
only single-cell suspensions of >95% viability were used for
injections.
Animal experiment procedure
Thirty-two BALB/C male nude mice weighing 16–18 g at
6–8 weeks of age were purchased from Beijing Laboratory Animal
Research Center (Beijing, China). All procedures involving mice
were approved by the University of South China Animal Management
Committee and Beijing City Animal Management Committee. Mice were
kept under specific pathogen-free conditions in accordance with the
NIH guidelines for the care and use of laboratory animals, fed a
standard rodent chow and maintained in a temperature-controlled
(23–25°C) facility with a strict 12 h light/dark cycle and given
free access to food and water. Each mouse was inoculated with a
subcutaneous injection of SGC-7901 cells (2×106 in 0.2
ml PBS) on the right side of the back. One day after inoculation of
tumor cells, the mice were randomly divided into 4 groups of 8 mice
and each group received a different dose of ROSI (Cayman Chemical
Company, Ann Arbor, MI, USA), i.e., group A (control group)
received normal saline, group B received ROSI 50 mg/kg, group C
received ROSI 75 mg/kg and group D received ROSI 100 mg/kg. Each
group was treated with the same volume (0.2 ml) by oral gavage once
every 2 days for 42 days. After 42 days, the mice were sacrificed
and the gastric cancer tissues were stored at −70°C for further
analysis.
Tumor growth assessment
To assess the tumor growth, tumor size was measured
with a sliding caliper for a maximal diameter (a) and a minimal
diameter (b) at 2, 7, 12, 17, 22, 27, 32, 37 and 42 days after
tumor cell inoculation, and then the volume (V) and the tumor
growth inhibition rate (IR) were calculated using the following
formula: V = a × b2/2; IR (%) = (1 − V of ROSI treatment
group/V of control group) × 100%.
Hematoxylin and eosin (H&E) staining
for histomorphological examination
Tumor tissues were fixed in 10% neutral buffered
(formalin) for 24 h and then embedded in paraffin. The
paraffin-embedded tissue blocks were cut into 4 μm sections. After
baking at 65°C for 60 min, the sections were de-paraffinized and
hydrated gradually, and examined after routine H&E staining.
The histomorphology of tumor cells was observed by light microscopy
at a magnification of ×400.
Immunohistochemistry staining for
lymphatic vessel density (LVD)
In order to evaluate tumor lymphangiogenesis, we
performed immunohistochemistry staining using D2–40 antibody for
LVD. The LVD was defined as the average number of D2–40 positive
vessels. In brief, formalin-fixed, paraffin-embedded tissue blocks
were cut into 4 μm sections and baked at 65°C for 60 min. The
sections were de-paraffinized and hydrated gradually, then heated
in a microwave oven for 15 min to retrieve antigens. Endogenous
peroxidase was blocked with 3% hydrogen peroxide methanol for 15
min at room temperature. After washing with PBS (0.01 M, pH 7.4)
for 3×5 min, the tumor sections were incubated with normal bovine
serum for 30 min at room temperature to eliminate nonspecific
staining. Mouse anti-human D2–40 antibody (1:200 dilution; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) was incubated with
the sections overnight at 4°C. After washing with 0.01 M PBS for
3×5 min, tissue sections were treated with goat anti-mouse IgG
secondary antibody (Dako, Denmark) for 1 h at room temperature.
Finally, tissue sections were immersed in 3-amino-9-ethyl carbazole
(DAB) (Sigma-Aldrich) for 5–10 min at room temperature, washed with
water, and then counterstained with 10% Mayer’s hematoxylin.
Negative and positive controls were used simultaneously to ensure
the specificity and reliability of the staining process. The
negative controls were performed by substituting PBS for the
primary antibody, and a positive section supplied by the
manufacturer of the staining kit was registered as a positive
control. Sections were observed under a microscope after being
mounted.
All sections were coded and evaluated by 2
experienced pathologists. All the stained vessels with brown by
D2–40 immunostaining were observed as typically positive lymphatic
vessels in thin-walled and tube-like structures exhibiting a
distinct inner cavity and devoid of red blood cells. LVDs
(D2–40-positive vessels) were calculated as described by Ohno et
al(21). Briefly, sections were
first scanned at a low magnification (x40) to identify lymphatic
vessel hot spots. Areas of the greatest vessel density were then
examined and counted under a higher magnification (x200), and the
average value of the 5 measurements was used for data analysis.
RT-PCR assay for vascular endothelial
growth factor (VEGF)-C and VEGF receptor-3 (VEGFR-3) mRNA
expressions
Tumor tissues were homogenized in a TissueLyzer
homogenizer (Qiagen, Hilden, Germany) with 5-mm steal beads. Total
RNA was extracted using TRIzol reagent (Omega, USA) according to
the manufacturer’s instructions. Reverse transcription was
performed using 1.0 μg total RNA and the reverse transcription
system (Sangon Biotech Co., Ltd., Shanghai, China). PCR
amplification was produced by a Biometra TGradient Thermoblock
(Biometra, Göttingen, Germany) in a reaction mixture of 25 μl
reaction volumes containing amplification primers. A 1 μl volume of
cDNA was used in each amplification reaction. The sequences of
specific primers were: VEGF-C mRNA forward, 5′-GCTT
CTTGTCTCTGGCGTGTTC-3′ and reverse, 5′-AAACTGAT TGTGACTGGTTTGG-3′;
VEGFR-3 mRNA forward, 5′-AGA TGCAGCCGGGCGCTGCGCT-3′ and reverse,
5′-TAGGCT GTCCCCGGTGTCAATC-3′; β-actin mRNA forward, 5′-TC
GTGCGTGACATCAAAGAG-3′ and reverse, 5′-TGGACAG TGAGGCCAAGATG-3′.
Amplification was performed as follows: 94°C, 5 min; 94°C, 15 sec;
68°C, 3 min and 68°C, 3 min for 35 cycles. The PCR products were
583 bp for VEGF-C; 143 bp for VEGFR-3 and 429 bp for β-actin. These
PCR products were electrophoresed on a 1.5% agarose gel and
visualized by ethidium bromide staining. Densitometries of the
bands were scanned and quantified by the LabWork software (LabWork™
version 3.00; Upland, CA, USA). Data were normalized against those
of the corresponding β-actin bands. Results were expressed as fold
increase over β-actin.
Western blotting for VEGF-C and VEGFR-3
protein expressions
The cell proteins were extracted according to NE-PER
Nuclear and Cytoplasmic Extraction Reagents kit (Pierce
Biotechnology, Inc., Rockford, IL, USA). The protein concentration
of each sample was determined by the Bradford assay. Twenty
micrograms of proteins of different groups were separated in 10%
SDS-PAGE, and transferred onto PVDF membrane (Amersham Pharmacia
Biotech). The membrane was blocked with 5.0 % nonfat milk in PBS at
4°C for 1 h and then incubated with rabbit anti-human VEGF-C
antibody (1:40 dilution), rabbit anti-human VEGFR-3 antibody (1:100
dilution) (both from Santa Cruz Biotechnology, Inc.) or mouse
anti-human β-actin (1:200 dilution; Xiaxin, China) overnight at
4°C. After washing 3 times with 0.1% Tween-20 in Tris-saline, the
membranes were incubated with goat anti-mouse or rabbit IgG
conjugated with horseradish peroxidase at 1:1,000 in PBS for 1 h at
room temperature. Blots were processed using an ECL kit (Santa Cruz
Biotechnology, Inc.) and exposed to X-ray film. Densitometries of
the bands were scanned and quantified by the LabWork software
(LabWork ™ version 3.00). Data were normalized against those of the
corresponding β-actin bands. Results were expressed as fold
increase over β-actin.
Statistical analyses
All experiments were repeated at least 6 times. SPSS
13.0 was used for the statistical analyses. Data are presented as
the means ± SD. Comparisons were made using a one-way ANOVA or
paired t-test. p<0.05 was considered to indicate statistically
significant differences.
Results
Effects of ROSI on the growth of human
gastric cancer transplanted in nude mice
Fig. 1 shows the
effects of ROSI on the growth of human gastric cancer transplanted
in nude mice in a dose-dependent manner. Before transplantation of
human gastric cancer into nude mice, the weight of nude mice had no
significant difference among each group (p>0.05). Following
transplantation, the volume of the tumor in nude mice increased
gradually with time. However, compared with the volumes of tumor in
the group treated with normal saline (group A), those in other
groups treated with different doses of ROSI were gradually
attenuated with the increasing dose of ROSI every day. After 42
days of transplantation, the tumor volume in the groups treated
with different ROSI doses were all significantly smaller than those
in the group treated with normal saline. Correspondingly, the tumor
growth inhibition rate gradually increased with the increasing dose
of ROSI every day and reached the maximum at day 42. These results
demonstrated that ROSI inhibits the tumor growth of human gastric
cancer in a time- and dose-dependent manner.
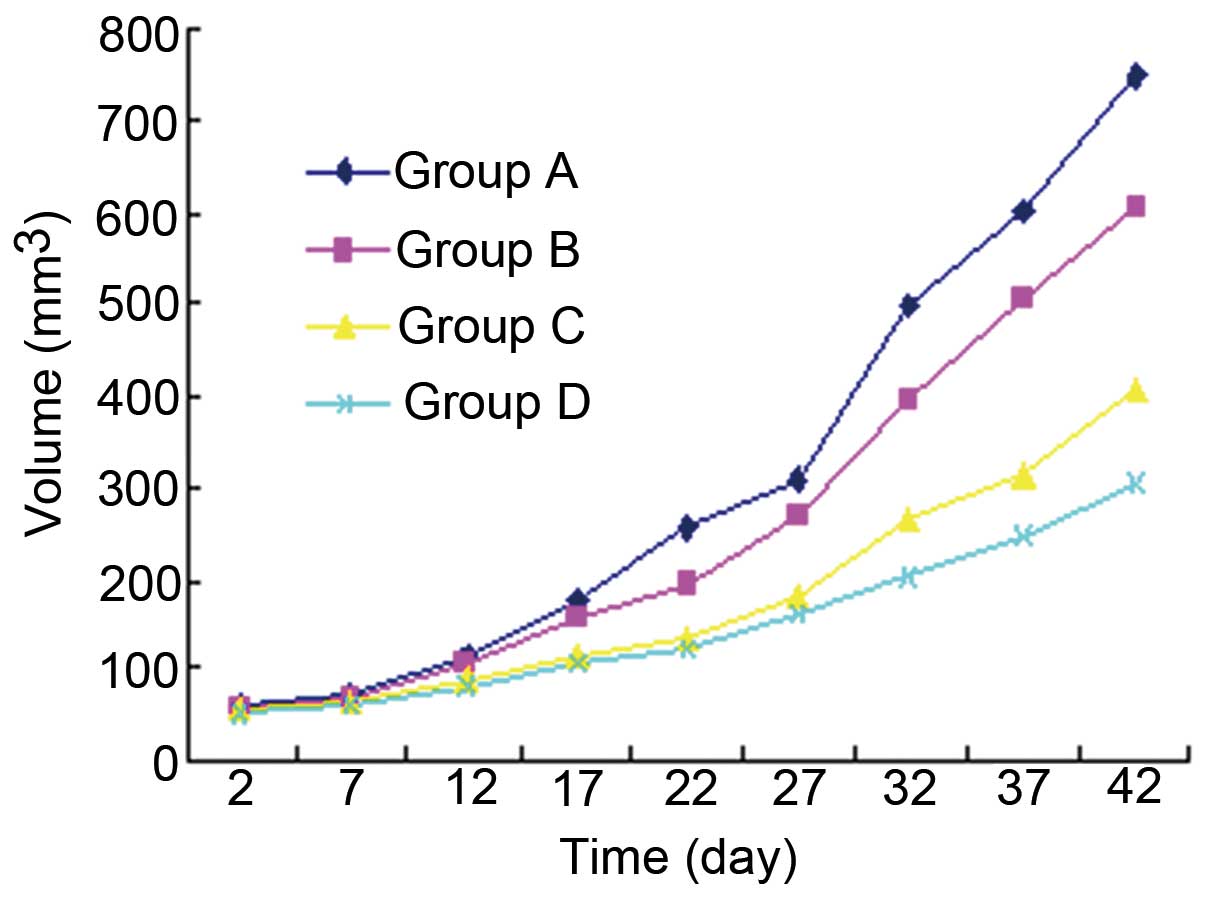 | Figure 1The inhibitory effects of
rosiglitazone (ROSI) on the growth of human gastric cancer
transplanted in nude mice. One day after inoculation of tumor
cells, mice were randomly divided into 4 groups and each group was
treated with a 0.2 ml different agent by oral gavage once every 2
days for 42 days. Tumor volume was measured at 2, 7, 12, 17, 22,
27, 32, 37 and 42 days after tumor cell inoculation. Group A
(control group), mice were treated with normal saline; group B,
mice were treated with ROSI 50 mg/kg; group G, mice were treated
with ROSI 75 mg/kg; group D, mice were treated with ROSI 100
mg/kg. |
The tumor volume changes and tumor growth inhibition
rate at day 42 are shown in Table
I.
 | Table ITumor volume and tumor growth
inhibition rate at day 42 (n=8). |
Table I
Tumor volume and tumor growth
inhibition rate at day 42 (n=8).
| Group | Tumor volume
(mm3) | Inhibition rate
(%) |
|---|
| A (normal saline,
0.2 ml/2 days) | 745.46±10.64 | - |
| B (50 mg/kg ROSI,
0.2 ml/2 days) |
606.65±10.64a,b | 18.62±0.34b |
| C (75 mg/kg ROSI,
0.2 ml/2 days) |
406.30±10.86a,c | 45.49±0.67c |
| D (100 mg/kg ROSI,
0.2 ml/2 days) |
304.59±11.92a | 59.14±1.02 |
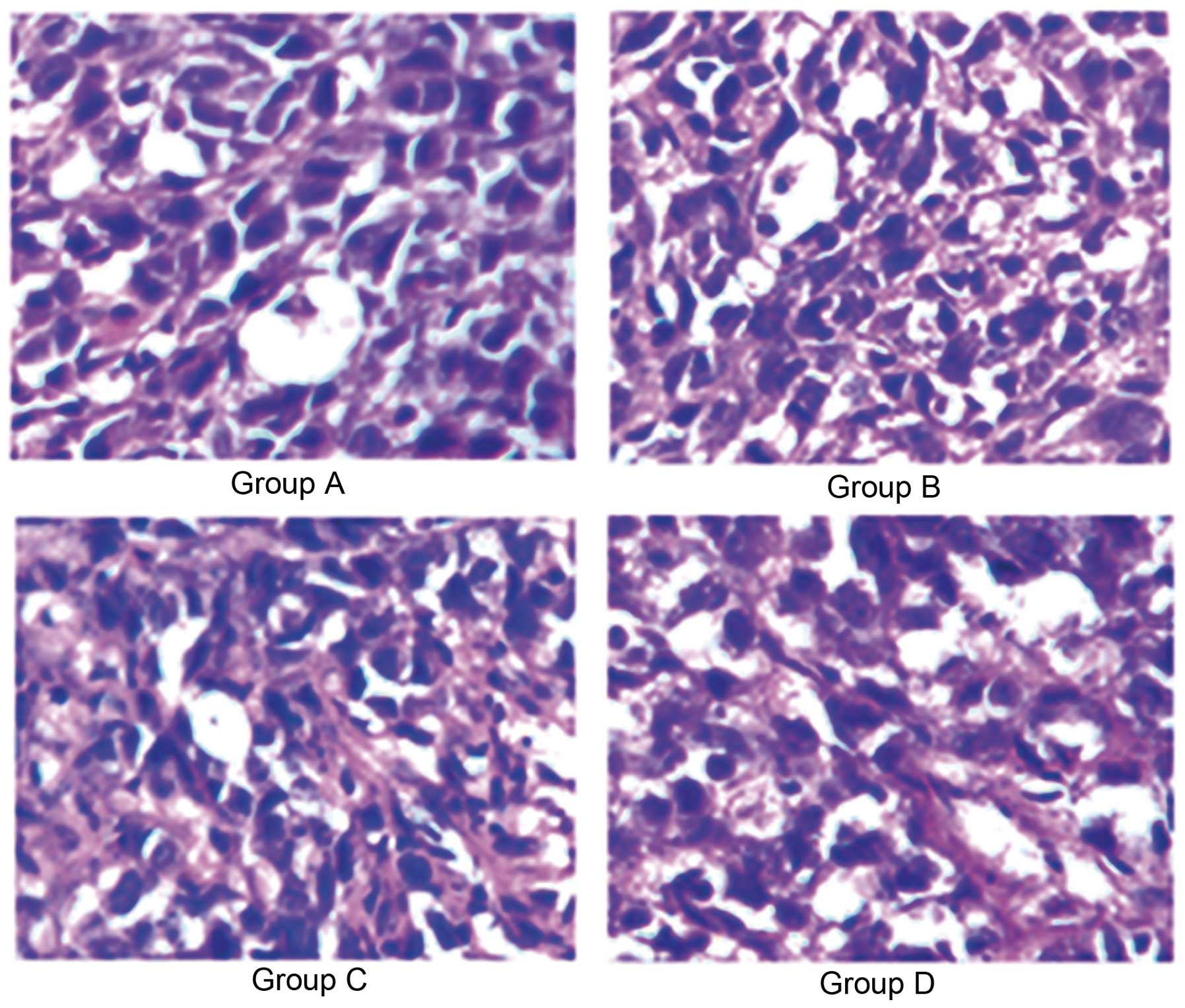
Effects of ROSI on the histomorphology of
gastric cancer cells
To observe the histomorphological changes of gastric
cancer cells, H&E staining was performed on the tumor tissue
prepared from nude mice after 42 days of transplantation with human
gastric cancer. The morphology of tumor cells was characterized by
both nuclear and cytoplasmic alteration. Nuclear features included
the presence of peculiar enlargement with hyperchromatism,
irregularity of outline, and chromatin clumping or smudging.
Cytoplasmic alterations included abundant cytoplasm, vacuolation,
or foam cell formation.
Our findings showed that the number of typical tumor
cells in the group treated with normal saline (group A) was more
than that in other groups treated with different ROSI doses.
Fig. 2 shows the representative
microscopic view of the H&E staining for tumor cells in nude
mice treated with normal saline and different ROSI dose at a
magnification of ×400, which illustrated that ROSI inhibited tumor
cell formation in a dose-dependent manner.
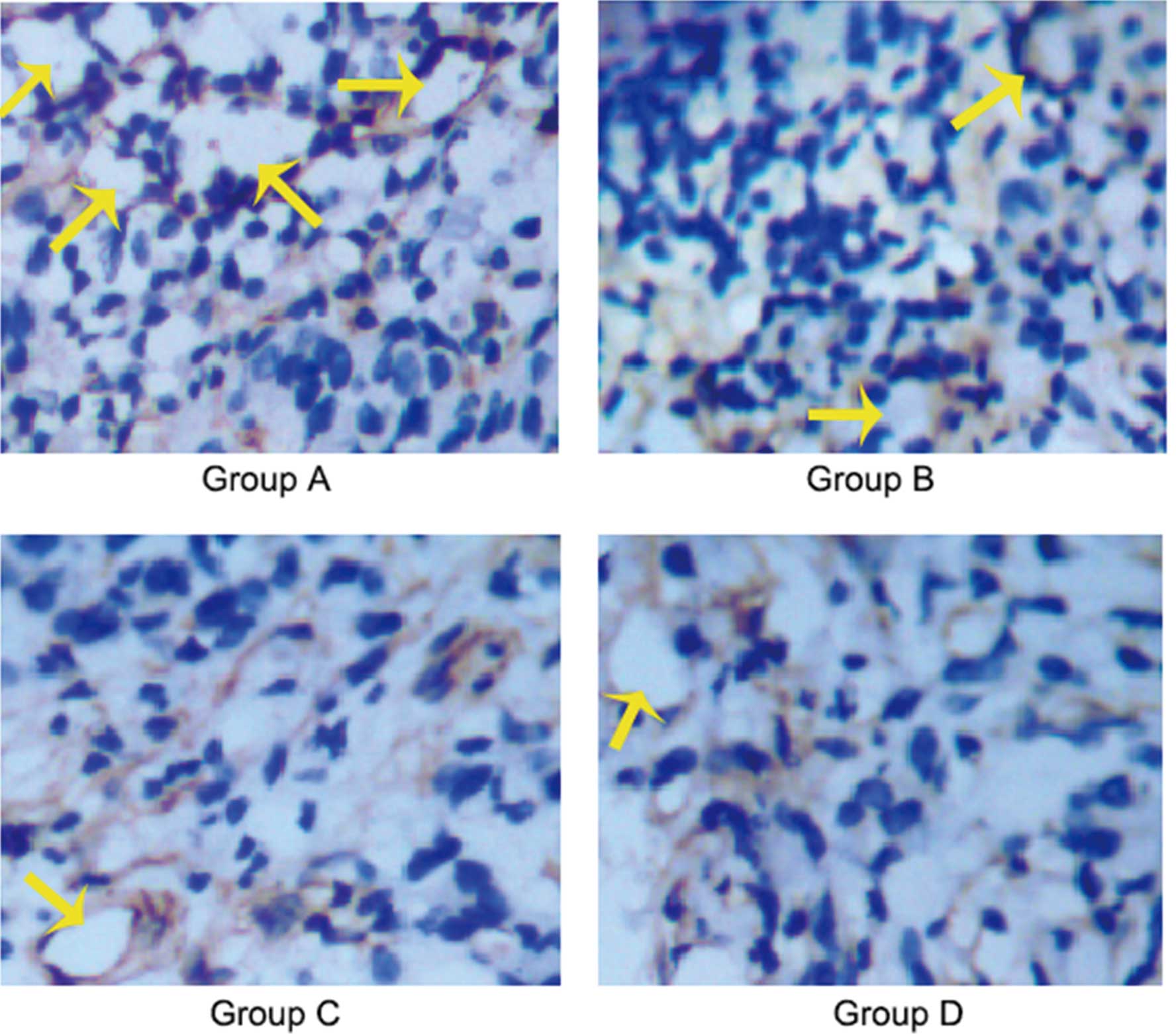
Effects of ROSI on the lymphangiogenesis
of gastric cancer
Immunohistochemical staining assays demonstrated
that there was a significantly different expression of D2–40
between control group and treatment groups, with the highest level
of expression for D2–40 being found in the control group with
normal saline (group A) and the lowest in the treatment group with
ROSI at 100 mg/kg/2 days (group D). The typically D2–40-positive
vessels (lymphatic vessels) in the control group were evidently
more than those in the treatment group. These outcomes indicated
that ROSI inhibited the lymphangiogenesis of human gastric cancer
transplanted in nude mice in a dose-dependent manner. The
representative microscopic view of the immunohistochemical staining
for LVD using D2–40 antibody in nude mice treated with normal
saline and different ROSI dose at a magnification of ×400 is
illustrated in Fig. 3, and the
average value of D2–40-positive vessels in 5 hot spot areas at ×200
magnifications (LVD) is summarized in Table II.
| Table IIThe average value of D2–40-positive
vessels in 5 hot spot areas at ×200 magnification (LVD) (n=8). |
Table II
The average value of D2–40-positive
vessels in 5 hot spot areas at ×200 magnification (LVD) (n=8).
| Group | LVD |
|---|
| A (normal saline,
0.2 ml/2 days) | 9.62±1.19 |
| B (50 mg/kg ROSI,
0.2 ml/2 days) | 6.62±0.80a,b |
| C (75 mg/kg ROSI,
0.2 ml/2 days) | 4.75±0.67a,c |
| D (100 mg/kg ROSI,
0.2 ml/2 days) | 2.25±0.45a |
Effects of ROSI on the VEGF-C and VEGFR-3
protein expressions of gastric cancer
Western blot analysis (Fig. 4A and B) showed that the expressions
of VEGF-C proteins were clearly inhibited in nude mice treated with
ROSI when compared to that in nude mice treated with normal saline
after 42 days of transplantation. Moreover, with the increasing
doses of ROSI, the inhibitory effects of ROSI on VEGF-C proteins
were more significant.
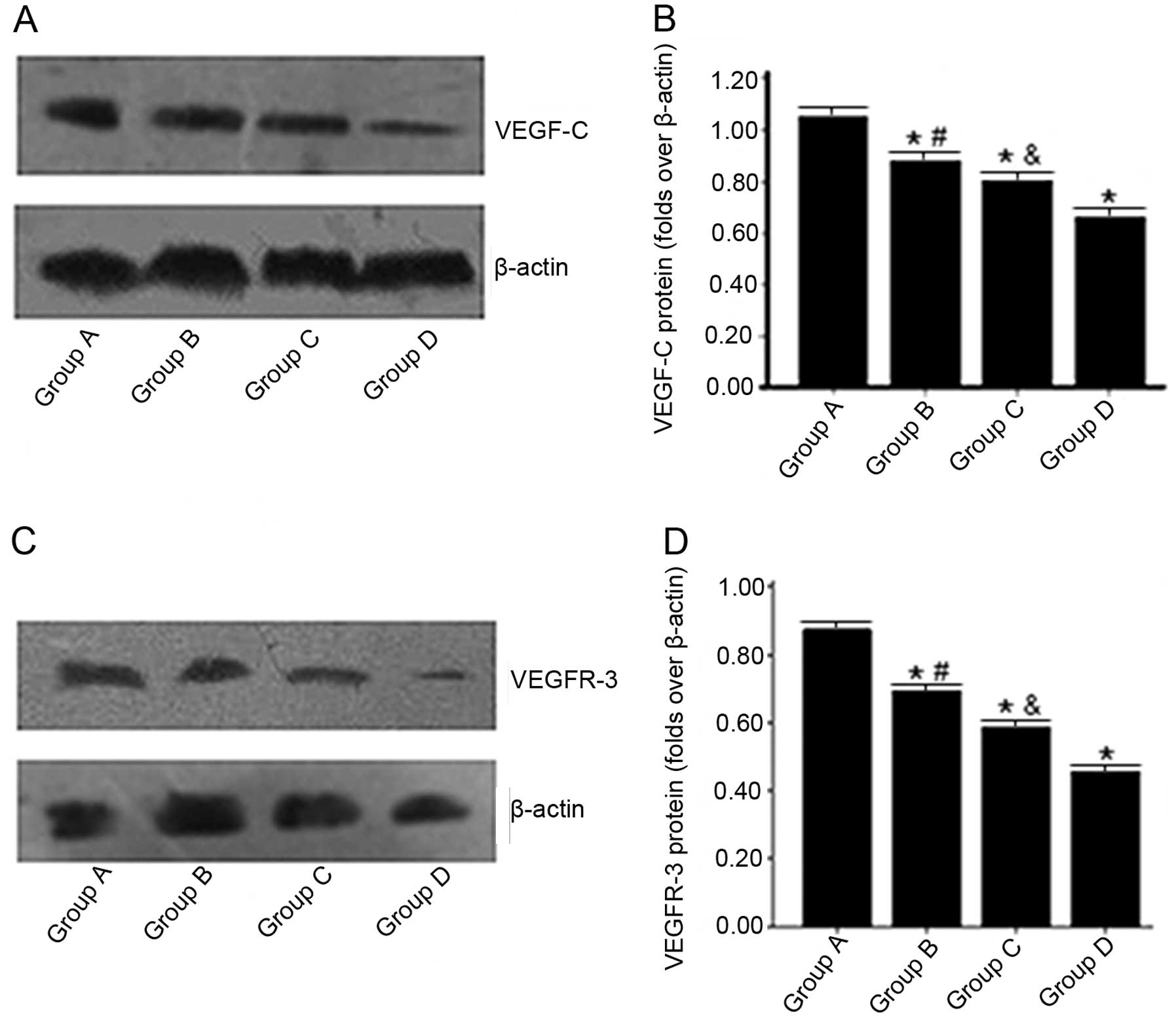 | Figure 4Effects of rosiglitazone (ROSI) on the
VEGF-C and VEGFR-3 protein expressions of gastric cancer. One day
after inoculation of tumor cells, mice were randomly divided into 4
groups and respectively treated with normal saline (group A), ROSI
50 mg/kg/2 days (group B), ROSI 75 mg/kg/2 days (group C) or ROSI
100 mg/kg/2 days (group D) for 42 days. After 42 days, the VEGF-C
and VEGFR-3 proteins of gastric cancer tissues in nude mice were
determined by western blot assay. (A) Western blot assays for
VEGF-C protein expressions in every group. Blots were processed
using an ECL kit and exposed to X-ray film. (B) Densitometric
analyses of the VEGF-C protein expressions. Results are expressed
as folds increase over β-actin. The bar represents the means ± SD
(n=8). *p<0.0001, vs. group A;
#p<0.0001, vs. group C and D and
&p<0.0001, vs. group D. (C) Western blot assays
for VEGFR-3 protein expressions in every group. Blots were
processed using an ECL kit and exposed to X-ray film. (D)
Densitometric analyses of the VEGFR-3 protein expressions. Results
are expressed as folds increase over β-actin. The bar represents
the means ± SD (n=8). *p<0.0001, vs. group A;
#p<0.0001, vs. group C and D and
&p<0.0001, vs. group D. VEGF-C, vascular
endothelial growth factor C; VEGFR-3, VEGF receptor-3. |
Similarly, Fig. 4C and
D showed that the expression levels of VEGFR-3 protein were
also gradually decreased with the ROSI dose.
Effects of ROSI on the VEGF-C and VEGFR-3
mRNA expressions of gastric cancer
We further performed RT-PCR analysis for VEGF-C and
VEGFR-3 mRNA expression of human gastric cancer transplantation in
nude mice.
Consistent with VEGF-C protein expression, Fig. 5A and B showed that the expression
levels of VEGF-C mRNA in nude mice treated with ROSI were
significantly lower than in nude mice treated with normal saline
after 42 days of transplantation. Furthermore, with the increase of
ROSI doses, the decrease of VEGF-C mRNA levels was more
significant.
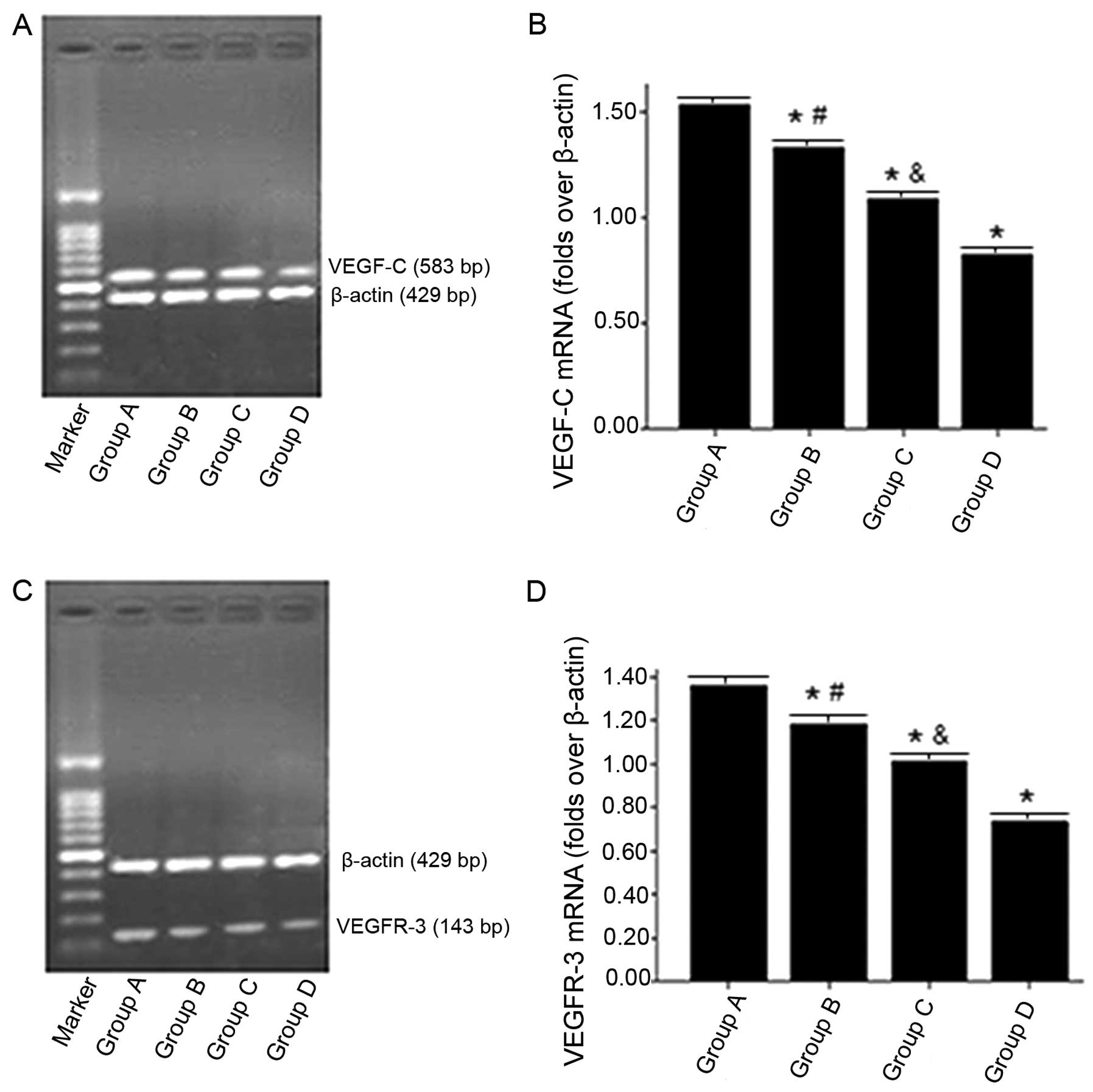 | Figure 5Effects of rosiglitazone (ROSI) on the
VEGF-C and VEGFR-3 mRNA expressions of gastric cancer. One day
after inoculation of tumor cells, mice were randomly divided into 4
groups and respectively treated with normal saline (group A), ROSI
50 mg/kg/2 days (group B), ROSI 75 mg/kg/2 days (group C) or ROSI
100 mg/kg/2 days (group D) for 42 days. After 42 days, the VEGF-C
and VEGFR-3 mRNA of gastric cancer tissues in nude mice were
determined by RT-PCR. (A) RT-PCR for VEGF-C mRNA expressions in
every group. PCR products (583 bp for VEGF-C and 429 bp for
β-actin) in a 1.5 % agarose gel were stained with ethidium
bromide.(B) Densitometric analyses of the VEGF-C mRNA expressions.
Results are expressed as folds increase over β-actin. The bar
represents the means ± SD (n=8). *p<0.0001, vs. group
A; #p<0.0001, vs. group C and D and
&p<0.0001, vs. group D. (C) RT-PCR for VEGFR-3
mRNA expressions in every group. PCR products (143 bp for VEGFR-3
and 429 bp for β-actin) in a 1.5 % agarose gel were stained with
ethidium bromide. (D) Densitometric analyses of the VEGFR-3 mRNA
expressions. Results are expressed as folds increase over β-actin.
The bar represents the means ± SD (n=8). *p<0.0001,
vs. group A; #p<0.0001, vs. group C and D and
&p<0.0001, vs. group D. VEGF-C, vascular
endothelial growth factor C; VEGFR-3, VEGF receptor-3. |
Fig. 5C and D show
that the expression levels of VEGFR-3 mRNA, also consistent with
the expression of VEGFR-3 protein, were gradually reduced with the
ROSI dose.
Discussion
In the present study, we investigated the effects of
ROSI on the growth and lymphangiogenesis of human gastric cancer
transplanted in nude mice. Our study demonstrated that nude mice
treated with different doses of ROSI had significantly decreased
tumor growth, clear histomorphological changes and less
lymphangiogenesis when compared with those treated with normal
saline. Furthermore, VEGF-C and VEGFR-3 expressions in nude mice
treated with different ROSI dose were significantly lower than in
nude mice treated with normal saline. These results indicated that
ROSI had inhibitory effects on the growth and lymphangiogenesis of
human gastric cancer, and these effects may be mediated through
modulation of VEGF-C and VEGFR-3 expression.
PPARs are ligand-activated transcription factors
that belong to the nuclear hormone receptor superfamily (22). They control several cellular and
metabolic processes. Three subtypes, PPARα, PPARβ/δ and PPARγ, have
been identified. Of these, PPARγ is considered a key regulator of
adipocyte differentiation (23–25),
and plays an important role in the induction of apoptosis,
inhibition of tumor cell growth and differentiation, inhibition of
tumor associated angiogenesis and regulation of immune system
(26,27). Perturbed PPARγ signaling has been
implicated in the formation of various solid tumors, such as
breast, stomach, colorectal and thyroid carcinomas (28–32).
Therefore, PPARγ and its ligands have been extensively evaluated as
potential molecular targets for anticancer drug development in the
past several years.
TZDs, including ROSI, troglitazone and pioglitazone,
are synthetic PPARγ ligands. Previous studies have demonstrated
that TZDs could inhibit gastric cancer cell line growth (10,33).
In addition, Lu et al(34)
found that PPARγ (+/−) mice were more susceptible to
N-methyl-N-nitrosourea-induced gastric cancer than wild-type (+/+)
mice, and troglitazone significantly reduced the incidence of
gastric cancer in PPARγ (+/+) mice but not in PPARγ (+/−) mice.
These results suggested that through PPARγ-dependent mechanism,
TZDs inhibited gastric cancer cell growth. In the present study, we
established a model of gastric cancer by subcutaneously inoculating
human gastric cancer cell line SGC-7901 into nude mice and found
that all the ROSI treatment groups had smaller tumor volume and
higher tumor growth inhibitory rate when compared with the control
group every day. Moreover, the number of typical tumor cells in the
control group was more than that in ROSI treatment groups.
Markedly, the inhibitory effect of ROSI on the growth was more
significant in the higher ROSI doses. These findings were
consistent with previous studies (10,33),
confirming that ROSI was able to inhibit the growth of human
gastric cancer.
In gastric cancer, both angiogenesis and
lymphangiogenesis are important events related to tumor growth and
progression (35–37). In fact, gastric cancer mainly
metastasizes via lymphatic vessels (20). Recent evidence suggests that tumor
lymphangiogenesis promotes lymphatic metastasis (6,8,38). The
induction of lymphangiogenesis in the sentinel lymph node started
even before tumor cells had arrived (6,38).
Thus, tumor lymphangiogenesis is an important early event of cancer
metastasis. However, less focus has been on the role of PPARγ or
ROSI on lymphangiogenesis although studies have suggested that ROSI
may have significant anticancer activities in various human
malignant tumors in vitro and in vivo(10–19).
In order to evaluate the effects of ROSI on the lymphangiogenesis
of gastric cancer, we performed immunohistochemistry assay for LVD
using D2–40 (podoplanin) antibody. D2–40 is a specific and
sensitive marker of lymphatic endothelial cells and is useful in
detecting lymphatic invasion by malignant tumors (39,40).
Our data showed that there was a significantly different expression
of D2–40 between the control group and treatment groups. The number
of LVD in the control group was significantly greater than that in
other ROSI treatment groups. This demonstrated that ROSI could
inhibit the lymphangiogenesis of human gastric cancer transplanted
in nude mice.
Lymphangiogenesis is driven primarily by the VEGF-C
and VEGF-D, members of the VEGF family of angiogenic factors. These
growth factors specifically activate their cognate receptor
tyrosine kinase VEGFR-3 located on lymphatic endothelial cells to
induce lymphatic capillary proliferation and growth (41,42).
Recent studies have demonstrated that VEGF-C/VEGFR-3 axis is
actively involved in regulating the migratory and invasive
activities of cancer cells (43,44).
Of note, the high expression of VEGF-C was found in early gastric
cancer patients with lymph node micrometastasis (45). To gain further insight into whether
ROSI would affect the expressions of these molecules, we determined
the expression changes of VEGF-C and VEGFR-3 mRNA and their
proteins in nude mice transplanted with human gastric cancer. Our
data demonstrated that the expression levels of VEGF-C protein and
mRNA in transplanted gastric tumor tissue were gradually decreased
with the ROSI dose. At the same time, the expression levels of
VEGFR-3 protein and mRNA were also gradually decreased with the
ROSI dose. These results were similar to the study reported by Liu
et al(46), who revealed
that norcantharidin suppressed lymphangiogenesis in human lymphatic
endothelial cells by simultaneously blocking VEGF-C and
VEGF-D/VEGFR-3. Collectively, our findings indicated that through
simultaneously inhibiting VEGF-C and VEGFR-3 expressions, ROSI
blocked the lymphangiogenic signaling and then prevented
lymphangiogenesis.
In conclusion, the present study provided evidence
that by suppressing VEGF-C and VEGFR-3 expressions, ROSI inhibits
the lymphangiogenesis and then inhibits the growth of human gastric
cancer transplanted in nude mice, which suggests that ROSI may be
an attractive anticancer agent for gastric cancer. However, further
studies to clarify the mechanisms by which ROSI downregulates
VEGF-C and VEGF-3 expressions in human gastric cancer are
required.
Acknowledgements
We thank Ai-guo Tang and Hui Xie (The Second XiangYa
Hospital of Central South University, Changsha, Hunan 410011,
China) for reading the manuscript and providing valuable
comments.
References
|
1
|
Brenner H, Rothenbacher D and Arndt V:
Epidemiology of stomach cancer. Methods Mol Biol. 472:467–477.
2009. View Article : Google Scholar
|
|
2
|
Jemal A, Center MM, DeSantis C and Ward
EM: Global patterns of cancer incidence and mortality rates and
trends. Cancer Epidemiol Biomarkers Prev. 19:1893–1907. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Macdonald JS, Smalley SR, Benedetti J, et
al: Chemoradiotherapy after surgery compared with surgery alone for
adenocarcinoma of the stomach or gastroesophageal junction. N Engl
J Med. 345:725–730. 2001. View Article : Google Scholar
|
|
4
|
Achen MG and Stacker SA: Molecular control
of lymphatic metastasis. Ann NY Acad Sci. 1131:225–234. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alitalo K, Tammela T and Petrova TV:
Lymphangiogenesis in development and human disease. Nature.
438:946–953. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mumprecht V and Detmar M:
Lymphangiogenesis and cancer metastasis. J Cell Mol Med.
13:1405–1416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McCarter MD, Clarke JH and Harken AH:
Lymphangiogenesis is pivotal to the trials of a successful cancer
metastasis. Surgery. 135:121–124. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matsumoto M, Roufail S, Inder R, et al:
Signaling for lymphangiogenesis via VEGFR-3 is required for the
early events of metastasis. Clin Exp Metastasis. Apr 17–2013.(Epub
ahead of print).
|
|
9
|
Saltiel AR and Olefsky JM:
Thiazolidinediones in the treatment of insulin resistance and type
II diabetes. Diabetes. 45:1661–1669. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He Q, Pang R, Song X, et al: Rosiglitazone
suppresses the growth and invasiveness of SGC-7901 gastric cancer
cells and angiogenesis in vitro via PPARγ dependent and independent
mechanisms. PPAR Res. 2008:6498082008.PubMed/NCBI
|
|
11
|
Zhang L, Hu JF, Li GQ, Xiao X and Su Q:
Rosiglitazone reverses mitomycin C resistance in human gastric
cancer cells. Am J Med Sci. 343:382–387. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Talbert DR, Allred CD, Zaytseva YY and
Kilgore MW: Transactivation of ERα by Rosiglitazone induces
proliferation in breast cancer cells. Breast Cancer Res Treat.
108:23–33. 2008.
|
|
13
|
Miao R, Xu T, Liu L, et al: Rosiglitazone
and retinoic acid inhibit proliferation and induce apoptosis in the
HCT-15 human colorectal cancer cell line. Exp Ther Med. 2:413–417.
2011.PubMed/NCBI
|
|
14
|
Cerquetti L, Sampaoli C, Amendola D, et
al: Rosiglitazone induces autophagy in H295R and cell cycle
deregulation in SW13 adrenocortical cancer cells. Exp Cell Res.
317:1397–1410. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chiu SJ, Hsaio CH, Tseng HH, et al:
Rosiglitazone enhances the radiosensitivity of p53-mutant HT-29
human colorectal cancer cells. Biochem Biophys Res Commun.
394:774–779. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lyon CM, Klinge DM, Do KC, et al:
Rosiglitazone prevents the progression of preinvasive lung cancer
in a murine model. Carcinogenesis. 30:2095–2099. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yee LD, Williams N, Wen P, et al: Pilot
study of rosiglitazone therapy in women with breast cancer: effects
of short-term therapy on tumor tissue and serum markers. Clin
Cancer Res. 13:246–252. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Esteva FJ, Moulder SL, Gonzalez-Angulo AM,
et al: Phase I trial of exemestane in combination with metformin
and rosiglitazone in nondiabetic obese postmenopausal women with
hormone receptor-positive metastatic breast cancer. Cancer
Chemother Pharmacol. 71:63–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luconi M, Mangoni M, Gelmini S, et al:
Rosiglitazone impairs proliferation of human adrenocortical cancer:
preclinical study in a xenograft mouse model. Endocr Relat Cancer.
17:169–177. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dicken BJ, Bigam DL, Cass C, Mackey JR,
Joy AA and Hamilton SM: Gastric adenocarcinoma: review and
considerations for future directions. Ann Surg. 241:27–39.
2005.PubMed/NCBI
|
|
21
|
Ohno M, Nakamura T, Kunimoto Y, Nishimura
K, Chung-Kang C and Kuroda Y: Lymphagenesis correlates with
expression of vascular endothelial growth factor-C in colorectal
cancer. Oncol Rep. 10:939–943. 2003.PubMed/NCBI
|
|
22
|
Mangelsdorf DJ, Thummel C, Beato M, et al:
The nuclear receptor superfamily: the second decade. Cell.
83:835–839. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Siersbaek R, Nielsen R and Mandrup S:
PPARγ in adipocyte differentiation and metabolism - novel insights
from genome-wide studies. FEBS Lett. 584:3242–3249. 2010.
|
|
24
|
Nedergaard J, Petrovic N, Lindgren EM,
Jacobsson A and Cannon B: PPARγ in the control of brown adipocyte
differentiation. Biochim Biophys Acta. 1740:293–304. 2005.
|
|
25
|
Fernyhough ME, Okine E, Hausman G, Vierck
JL and Dodson MV: PPARγ and GLUT-4 expression as developmental
regulators/markers for preadipocyte differentiation into an
adipocyte. Domest Anim Endocrinol. 33:367–378. 2007.
|
|
26
|
Elrod HA and Sun SY: PPARγ and apoptosis
in cancer. PPAR Res. 2008:7041652008.
|
|
27
|
Kim S, Lee JJ and Heo DS: PPARγ ligands
induce growth inhibition and apoptosis through p63 and p73 in human
ovarian cancer cells. Biochem Biophys Res Commun. 406:389–395.
2011.
|
|
28
|
Zhou J, Zhang W, Liang B, et al: PPARγ
activation induces autophagy in breast cancer cells. Int J Biochem
Cell Biol. 41:2334–2342. 2009.
|
|
29
|
Zaytseva YY, Wallis NK, Southard RC and
Kilgore MW: The PPARγ antagonist T0070907 suppresses breast cancer
cell proliferation and motility via both PPARγ-dependent and
-independent mechanisms. Anticancer Res. 31:813–823. 2011.
|
|
30
|
Kitamura S, Miyazaki Y, Hiraoka S, et al:
PPARγ inhibits the expression of c-MET in human gastric
cancer cells through the suppression of Ets. Biochem Biophys
Res Commun. 265:453–456. 1999.
|
|
31
|
Takahashi H, Hosono K, Uchiyama T, et al:
PPARγ ligand as a promising candidate for colorectal cancer
chemoprevention: a pilot study. PPAR Res. 2010:2578352010.
View Article : Google Scholar
|
|
32
|
Wood WM, Sharma V, Bauerle KT, et al:
PPARγ promotes growth and invasion of thyroid cancer cells. PPAR
Res. 2011:1717652011. View Article : Google Scholar
|
|
33
|
Sato H, Ishihara S, Kawashima K, et al:
Expression of peroxisome proliferator-activated receptor (PPAR)γ in
gastric cancer and inhibitory effects of PPARγ agonists. Br J
Cancer. 83:1394–1400. 2000.
|
|
34
|
Lu J, Imamura K, Nomura S, et al:
Chemopreventive effect of peroxisome proliferator-activated
receptor γ on gastric carcinogenesis in mice. Cancer Res.
65:4769–4774. 2005.
|
|
35
|
Chen J, Zhi Y, Chang X, Zhang S and Dai D:
Expression of ADAMTS1 and its correlation with angiogenesis in
primary gastric cancer and lymph node metastasis. Dig Dis Sci.
58:405–413. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu H, Xin Y, Xu C and Xiao Y: Capecitabine
combined with (−)-epigallocatechin-3-gallate inhibits angiogenesis
and tumor growth in nude mice with gastric cancer xenografts. Exp
Ther Med. 3:650–654. 2012.
|
|
37
|
Fidler IJ: The biology of cancer
metastasis. Semin Cancer Biol. 21:712011. View Article : Google Scholar
|
|
38
|
Al-Rawi MA and Jiang WG: Lymphangiogenesis
and cancer metastasis. Front Biosci. 16:723–739. 2011. View Article : Google Scholar
|
|
39
|
Raica M, Cimpean AM and Ribatti D: The
role of podoplanin in tumor progression and metastasis. Anticancer
Res. 28:2997–3006. 2008.PubMed/NCBI
|
|
40
|
Yu JW, Wu JG, Tajima Y, et al: Study on
lymph node metastasis correlated to lymphangiogenesis, lymphatic
vessel invasion, and lymph node micrometastasis in gastric cancer.
J Surg Res. 168:188–196. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Makinen T, Veikkola T, Mustjoki S, et al:
Isolated lymphatic endothelial cells transduce growth, survival and
migratory signals via the VEGF-C/D receptor VEGFR-3. EMBO J.
20:4762–4773. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Van Trappen PO, Steele D, Lowe DG, et al:
Expression of vascular endothelial growth factor (VEGF)-C and
VEGF-D, and their receptor VEGFR-3, during different stages of
cervical carcinogenesis. J Pathol. 201:544–554. 2003.PubMed/NCBI
|
|
43
|
Su JL, Chen PS, Chien MH, et al: Further
evidence for expression and function of the VEGF-C/VEGFR-3 axis in
cancer cells. Cancer Cell. 13:557–560. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Takizawa H, Kondo K, Fujino H, et al: The
balance of VEGF-C and VEGFR-3 mRNA is a predictor of lymph node
metastasis in non-small cell lung cancer. Br J Cancer. 95:75–79.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Arigami T, Natsugoe S, Uenosono Y, et al:
Vascular endothelial growth factor-C and -D expression correlates
with lymph node micrometastasis in pN0 early gastric cancer. J Surg
Oncol. 99:148–153. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu ZY, Qiu HO, Yuan XJ, et al:
Suppression of lymphangiogenesis in human lymphatic endothelial
cells by simultaneously blocking VEGF-C and VEGF-D/VEGFR-3 with
norcantharidin. Int J Oncol. 41:1762–1772. 2012.PubMed/NCBI
|