Introduction
Photodynamic therapy (PDT) is used clinically to
treat malignant cancer (1) and is
recognized as a minimally invasive therapeutic strategy. PDT leads
to the photochemical generation of cytotoxic singlet oxygen through
light irradiation of the photosensitizer, which ultimately kills
the target cancer cells (2).
The chlorophyll-based photosensitizer, pheophorbide
a (Pa), localizes to the mitochondria and exhibits an antitumor
effect on human lung cancer, liver cancer and TPA-induced skin
tumor in in vivo models (3–5).
Furthermore, Pa-mediated PDT (Pa-PDT) leads to the depolarization
of mitochondrial membrane potential by the rapid generation of
singlet oxygen during light irradiation and inhibits tumor growth
in a number of human cancer cells, including Jurkat leukemia,
pigmented melanoma, colonic cancer and pancreatic carcinoma cells
(6–9). The inhibitory effect of Pa-PDT has
also been reported in the hepatitis B virus- and hepatitis C
virus-induced human hepatoma cell lines with multidrug resistance
(10,11). Pa-PDT has been reported to induce
the release of cytochrome c and trigger the activation of
the mitochondrial-mediated apoptosis pathway in malignant carcinoma
cells (4,10–13).
Although photosensitizer-mediated PDT exerts antitumor effects in
several types of cancer, the molecular mechanisms of PDT signaling
remain unknown.
The most common types of skin cancer are
non-melanoma skin cancers (NMSCs): basal cell carcinoma (BCC) and
squamous cell carcinoma (SCC) (14). Clinical studies have applied PDT
treatment to NMSC, such as BCC. However, PDT is not commonly used
to treat SCC due to the poor penetration of photosensitizers
through the keratotic layers generally covering these tumors
(14,15). In addition, melanoma skin cancers
are also difficult to treat with PDT as, unlike BCC, melanomas grow
aggressively, and the highly pigmented melanomas are unaffected by
treatment with photosensitizers that absorb in the visible range
(16,17).
In the present study, the therapeutic potential of
Pa-PDT was evaluated in the human non-melanoma and melanoma skin
cancer cell lines, A431 and G361. Our findings provided the first
evidence that Pa-PDT can induce autophagy and apoptosis pathways by
upregulating ERK1/2 and p38 MAPK activation, and our research
demonstrated that Pa-PDT can inhibit the growth of both tumors
in vivo, suggesting that Pa-PDT may be an effective
alternative therapy to treat skin cancer.
Materials and methods
Cell culture and reagents
A human epithelial carcinoma cell line (A431) and
malignant melanoma cell line (G361) were maintained in DMEM medium.
Both cell lines were cultured in medium supplemented with 10% fetal
bovine serum (FBS), 100 U/ml of penicillin and 100 μg/ml of
streptomycin and maintained at 37°C in a humidified incubator with
5% CO2. Pa was prepared as a 10 mM stock in dimethyl
sulfoxide (DMSO; BDH Merck, Darmstadt, Germany). Further dilutions
were made in serum-free DMEM.
Photodynamic therapy
The PDT irradiation light source was a
light-emitting diode (LED; 613–645 nm; Philips Luxeon Lumileds, San
Jose, CA, USA). The cells (1×105/well) were
pre-incubated with Pa in complete growth medium in the dark for 2
h. For the following experiments, the cells were irradiated at 1.25
J/cm2. After 24 h of incubation, the cells were rinsed
with phosphate-buffered saline (PBS). For the control group, the
cells were incubated in the same medium without Pa or light.
Cell proliferation assay
The MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl
tetrazolium bromide) assay was used to assess cell proliferation
after Pa-PDT treatment. After the medium was removed, the cells
were incubated with an MTT solution (5 mg/ml in PBS) for 3 h, and
the absorbance was measured using an auto ELISA plate reader at 570
nm. In addition, we performed cell proliferation assays using a
cell proliferation ELISA kit (Roche Applied Science, Indianapolis,
IN, USA).
Western blotting
The cells were treated with Pa-PDT for 24 h. The
cells were then washed with PBS and harvested in lysis buffer.
Samples containing equal amounts of protein were loaded onto each
lane of an SDS-polyacrylamide gel for electrophoresis and
subsequently transferred onto a polyvinylidene difluoride membrane.
The membranes were blocked and then incubated with antibodies.
Antibodies against Beclin-1, Bcl-2, Atg5, LC3B and p-mTOR were
purchased from Cell Signaling Technology (Beverly, MA, USA); p-Akt,
caspase-7, PARP and β-actin were purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA).
Detection and quantification of acidic
vesicular organelles with acridine orange staining using flow
cytometry
Autophagy is characterized by the formation of
acidic vesicular organelles (AVOs) (17). A431 and G361 cells were seeded in
6-cm2 plates and incubated for 24 h. After treatment
with Pa-PDT for 24 h, acridine orange (1 μg/ml) was added to the
living cells for 30 min, and the cells were removed from the plate
with trypsin-EDTA and collected in phenol red-free growth medium.
Green (510–530 nm) and red (650 nm) fluorescence emission from
1×104 cells illuminated with blue (488 nm) excitation
light was measured with a FACSCalibur using CellQuest software
(Becton-Dickinson).
MDC staining
To observe autophagy formation, skin cells were
grown on glass coverslips for 24 h in a humidified incubator in 5%
CO2 and at 37°C. After treatment with Pa-PDT for 24 h,
the cells were treated with 0.05 mM monodansylcadaverine (MDC;
Sigma-Aldrich Chemical) at 37°C in 5% CO2 for 10 min.
The cells were then fixed with 4% paraformaldehyde in PBS for 10
min. Following incubation, the cells were washed three times with
PBS and immediately analyzed under a fluorescence microscope
(IX-71; Olympus, Tokyo, Japan). Fluorescence of MDC was measured at
the excitation wavelength of 380 nm with an emission filter at 530
nm.
Annexin V-FITC/PI double staining
The cells were harvested and fixed with 70% ethanol
for 1 h at 4°C for cell cycle analysis. After washing with cold
PBS, the cells were incubated with DNase-free RNase and propidium
iodide (PI) at 37°C for 30 min. The specific binding of Annexin
V-FITC/PI was performed by incubating the cells for 15 min at room
temperature in a binding buffer (10 mM HEPES, 140 mM NaCl, 2.5 mM
CaCl2, pH 7.4) containing saturating concentrations of
Annexin V-FITC and PI. Following incubation, the cells were
pelleted and analyzed in a FACScan analyzer (Beckman Coulter Inc.,
Fullerton, CA, USA).
Caspase-3 activity assay
Caspase-3 activity was assessed using a caspase-3
colorimetric assay kit (Clontech, Palo Alto, CA, USA) following the
manufacturer’s instructions. The cells seeded in 6-well plates
(2×105 cells/well) were treated with or without Pa-PDT
for 24 h as previously described. The cells were collected and
resuspended in lysis buffer containing 50 mM HEPES, pH 7.4, 0.1%
CHAPS, 1 mM DTT, 0.1 mM EDTA and 0.1% Triton X-100. Cell lysates
were centrifuged at 12,000 × g for 10 min at 4°C. The supernatants
were incubated with the reaction buffer containing 2 mM Ac-DEVD-pNA
for 1 h at 37°C. Caspase activity was determined by measuring the
absorbance at 405 nm.
In vivo chorioallantoic membrane assay
(CAM) assay and immunohistochemistry assay
The CAM assay was used to examine the inhibition of
tumor growth in vivo. The CAM assay was performed as
previously described (18).
Briefly, fertilized chicken eggs were transferred to an egg
incubator maintained at 37°C and 50% humidity and allowed to grow
for 10 days. The fertilized chick eggs were sterilized, and a
1-cm2 window was cut, using the false air sac technique,
on one side of the egg to expose the CAM. Skin cancer cells
(2×106) were placed on the exposed CAM, and the windows
were sealed with transparent tape. The eggs were incubated in a
humidified incubator at 37°C for three days and pre-treated with
normal saline or Pa (0.1 μM) prior to light exposure (1.25
J/cm2). At the next day, the excised tumors were fixed
in 10% formalin and paraffin-embedded and cell staining was
performed with hematoxylin and eosin (H&E).
Statistical analysis
The statistical analyses were performed with data
obtained from three independent experiments. The data are
represented as the mean ± SEM. A P-value <0.05 was considered to
indicate a statistically significant result.
Results
Antitumor effect of Pa-PDT on human skin
cancer cells
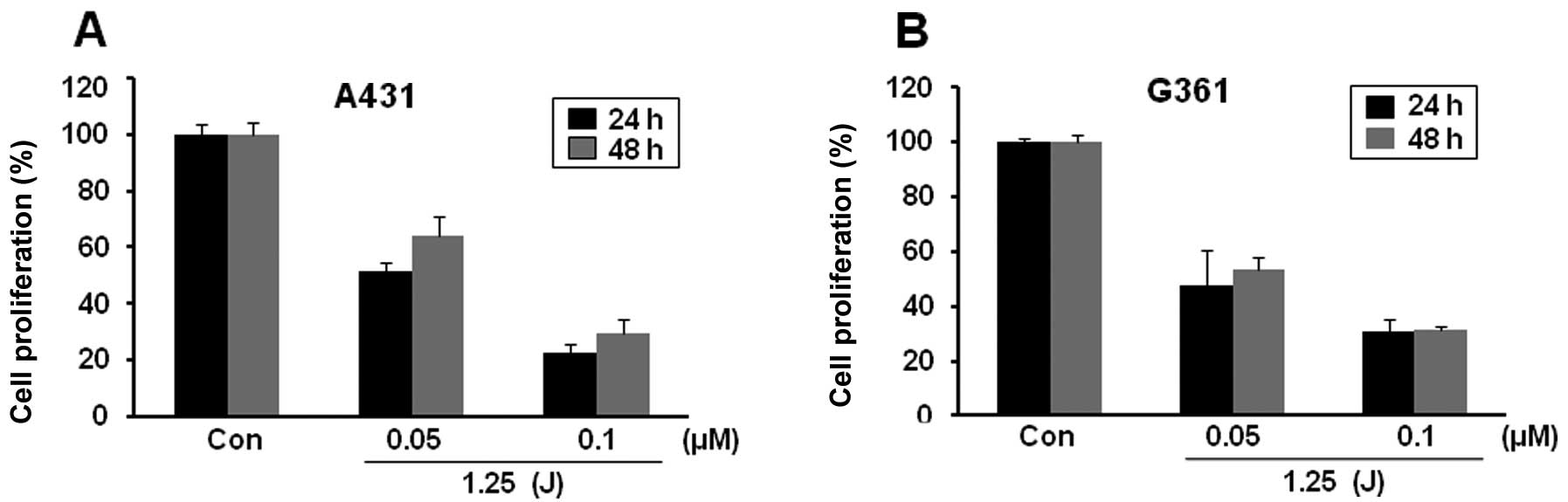
To test the effect of Pa-PDT, A431 and G361 cells
were pre-treated with different concentrations of Pa (0.05 or 0.1
μM) for 2 h in the dark, followed by photoactivation with 1.25
J/cm2 of LED. Twenty-four hours after exposure, the cell
proliferation was determined using an MTT assay. We found that
these cell lines exhibited no significant decrease in cell
proliferation due to Pa or light alone (data not shown). However,
the cell proliferation in both cell lines was severely decreased by
Pa-PDT treatment. In A431 cells, Pa doses of 0.05 and 0.1 μM with
light resulted in cell growth inhibition rates of 48.8 and 77.7% at
24 h, respectively (Fig. 1A).
Similarly, Pa-PDT also induced a significant cytotoxicity in G361
melanoma cells in a Pa-dose-dependent manner (Fig. 1B). After 48 h PDT treatment, the
cell growth slightly recovered in both cells compared with growth
at 24 h (Fig. 1). These results
demonstrate that Pa-PDT exerts an antiproliferative effect on human
skin cancer cells.
Expression of LC3B and Beclin 1 in
Pa-PDT-treated cells
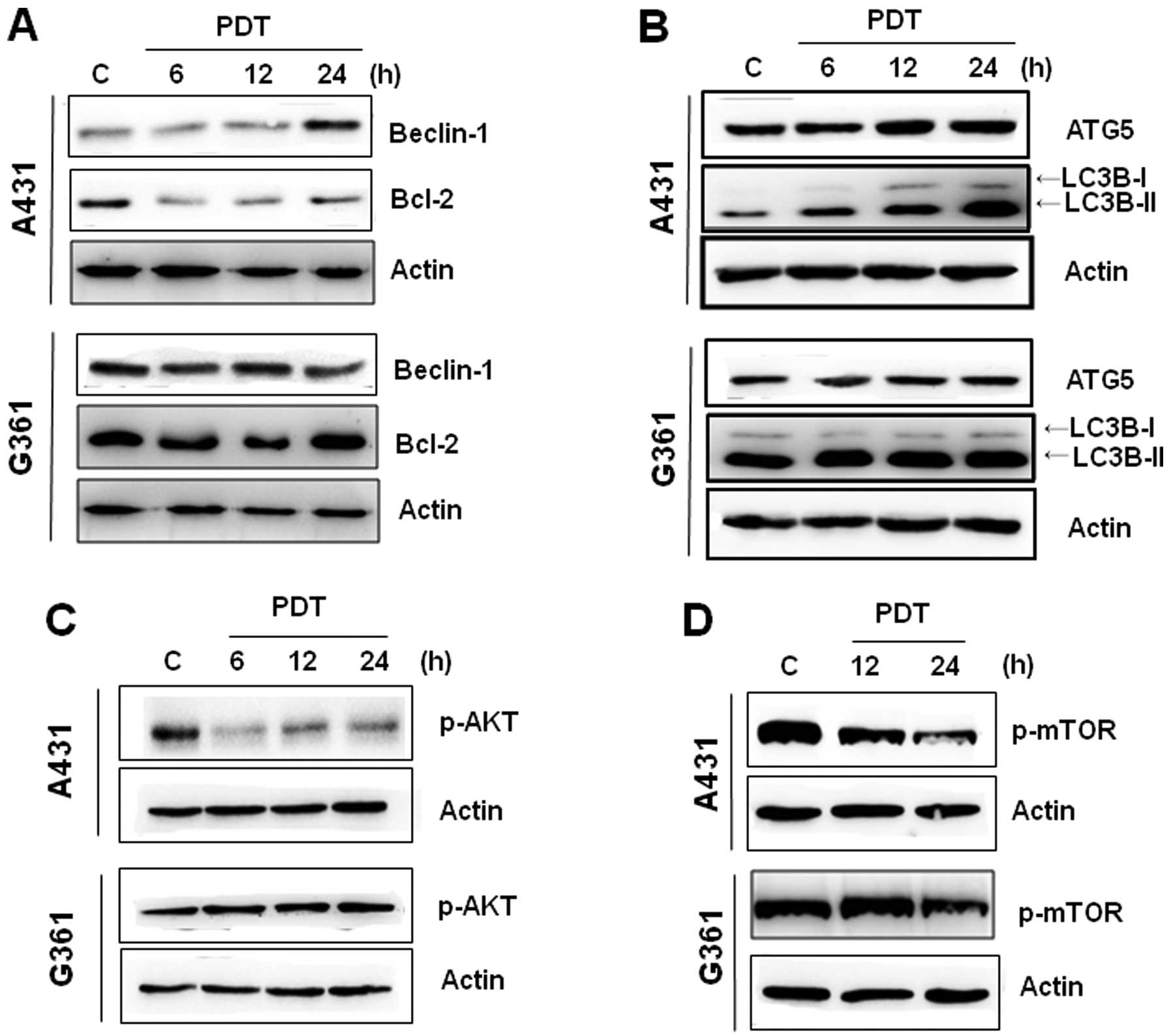
To further elucidate the underlying mechanisms of
Pa-PDT-induced cell growth inhibition, we assessed the levels of
Beclin 1, Bcl-2, ATG5 and LC3B-I/II, which play a crucial role in
autophagy. Western blotting revealed that Pa-PDT induced an
increase in the expressions of Beclin 1 in a time-dependent manner,
whereas the expression of Bcl-2 (Beclin 1 regulatory proteins) was
reduced in A431 cells (Fig. 2A).
The induction of ATG5 and LC3-II expression began to be clearly
observed at 12 h after Pa-PDT treatment (Fig. 2B). However, the G361 cell line
exhibited no significant expression of these proteins after Pa-PDT
treatment. This finding suggested the possibility that the
mechanism of Pa-PDT-mediated cell growth inhibition in A431 cells,
but not in G361 cells, was related to the Beclin 1-dependent
autophagy.
The mTOR/AKT pathway is a major signaling pathway
that regulates autophagy (19). To
confirm whether Pa-PDT regulated autophagy in A431 cells, we
assessed the influence of Pa-PDT on the activation of mTOR and Akt.
In treating A431 cells with Pa-PDT, the levels of p-AKT and p-mTOR
decreased at 6 and 12 h (Fig. 2C and
D). In G361 cells, Pa-PDT did not affect the expression of
p-Akt but slightly decreased the level of phospho-mTOR at 24 h.
These results indicate that Pa-PDT inhibits the Akt/mTOR pathway
and that these changes induce autophagy in A431 cells.
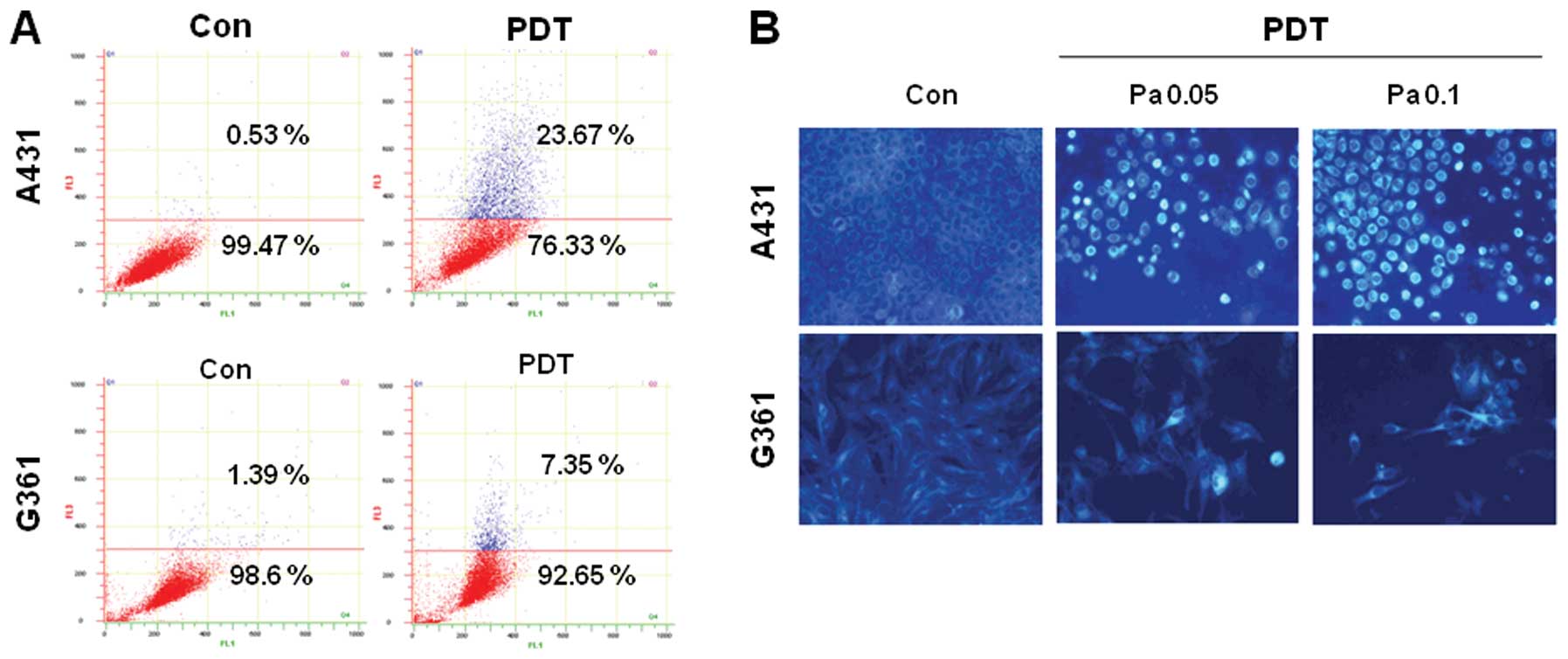
Quantization and detection of autophagic
vacuoles by Pa-PDT
Autophagy is characterized by the formation of
acidic AVOs (20). To confirm the
presence of vacuoles in Pa-PDT-treated A431 cells, we performed
acridine orange staining to obtain acidic AVOs. As shown in
Fig. 3A, the number of AVOs was
increased in A431 cells treated with Pa-PDT compared with the
control cells. In G361 cells, the number of AVOs was slightly
increased. Similar results were also obtained by MDC staining. The
fluorescent compound MDC is a specific marker for autolysosomes and
is commonly used to stain autophagic vesicles. A431 and G361 cells
were treated with PDT for 24 h and analyzed using fluorescence
microscopy. As shown in Fig. 3B, in
the control cells, MDC-labeled vacuoles were not detected. However,
in Pa-PDT-treated A431 cells, MDC-labeled cells were strongly
detected, and the number of these cells increased during treatment
in a concentration-dependent manner. In G361 cells, MDC-labeled
cells were weakly detected. Collectively, these results also
suggest that Pa-PDT induced autophagy in A431 cells but not in G361
cells.
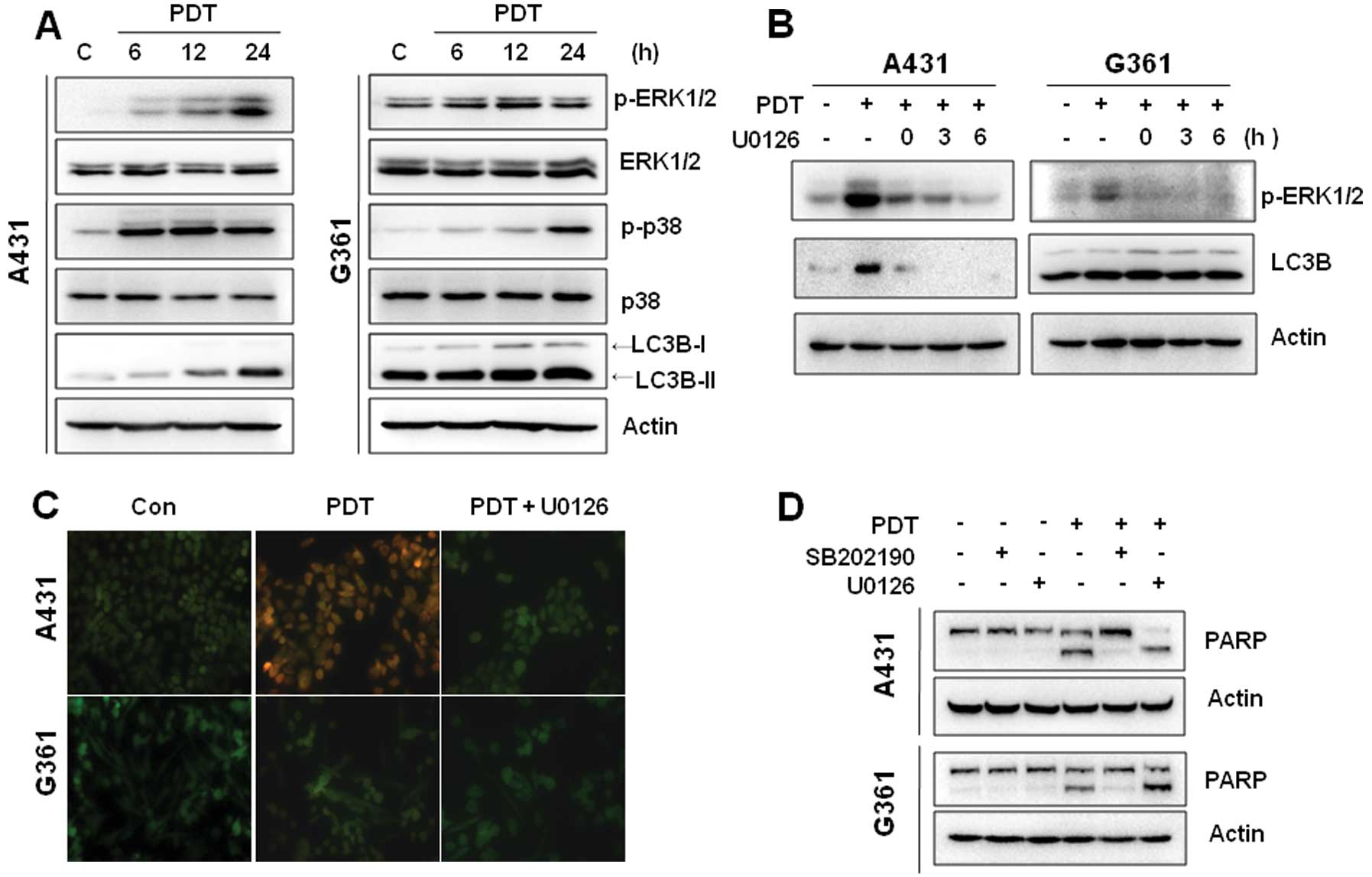
Activation of ERK1/2 and p38 MAPK in
Pa-PDT-treated cells
It has been suggested that PDT can activate the MAPK
pathway and regulate the cell death process (21,22).
The activation of MAPKs in Pa-PDT-treated cells was monitored by
western blotting. As shown in Fig.
4A, the phosphorylation of ERK1/2 and p38 MAPK was induced
after 6 h of Pa-PDT treatment in A431 cells, whereas the activation
of JNK was not detected. Similarly, the LC3B-I/II level was
increased in a time-dependent manner during Pa-PDT treatment. In
G361 cells, the phosphorylation of p38 MAPK was induced at 24 h
after Pa-PDT treatment, whereas the phosphorylation of ERK was
slightly increased at 12 h (Fig.
4A).
We next examined whether MAPK signaling was involved
in Pa-PDT-induced autophagy. A431 cells were treated with an ERK1/2
inhibitor (U0126) in a gradient for 6 h followed by western blot
analysis. As expected, ERK phosphorylation by PDT was inhibited by
treatment with U0126 in a time-dependent manner. Following U0126
treatment, LC3B-II protein levels decreased in A431 cells,
suggesting that ERK1/2 signaling was involved in Pa-PDT-induced
autophagy (Fig. 4B). However, the
inhibition of ERK phosphorylation by U0126 did not affect the
LC3-II levels in G361 cells, suggesting that Pa-PDT induced cell
growth inhibition by other pathways in G361 cells. To confirm the
effect of U0126 on Pa-PDT-induced autophagy, we performed acridine
orange staining to visualize autophagy using fluorescence
microscopy. As shown in Fig. 4C,
the control cells primarily emitted green fluorescence, indicating
a lack of AVO. Pa-PDT strongly increased the intensity of red
fluorescence in A431 cells but not in G361 cells. Alternatively,
treatment of cells with 20 μM U0126 decreased the formation of
Pa-PDT-induced AVO. These results indicate that the ERK1/2 pathway
is involved in Pa-PDT-induced autophagy in A431 cells.
To further examine the role of p38 MAPK in
Pa-PDT-mediated cytotoxicity, the p38 MAPK-specific inhibitor
SB202190 was used. Pa-PDT induced PARP cleavage in A431 and G361
cells. SB202190 inhibited PARP cleavage in both cells. However,
U0126 did not block PARP cleavage by Pa-PDT (Fig. 4D). Collectively, these results
suggest that Pa-PDT also leads to apoptosis through p38 MAPK
activation in A431 and G361 cells.
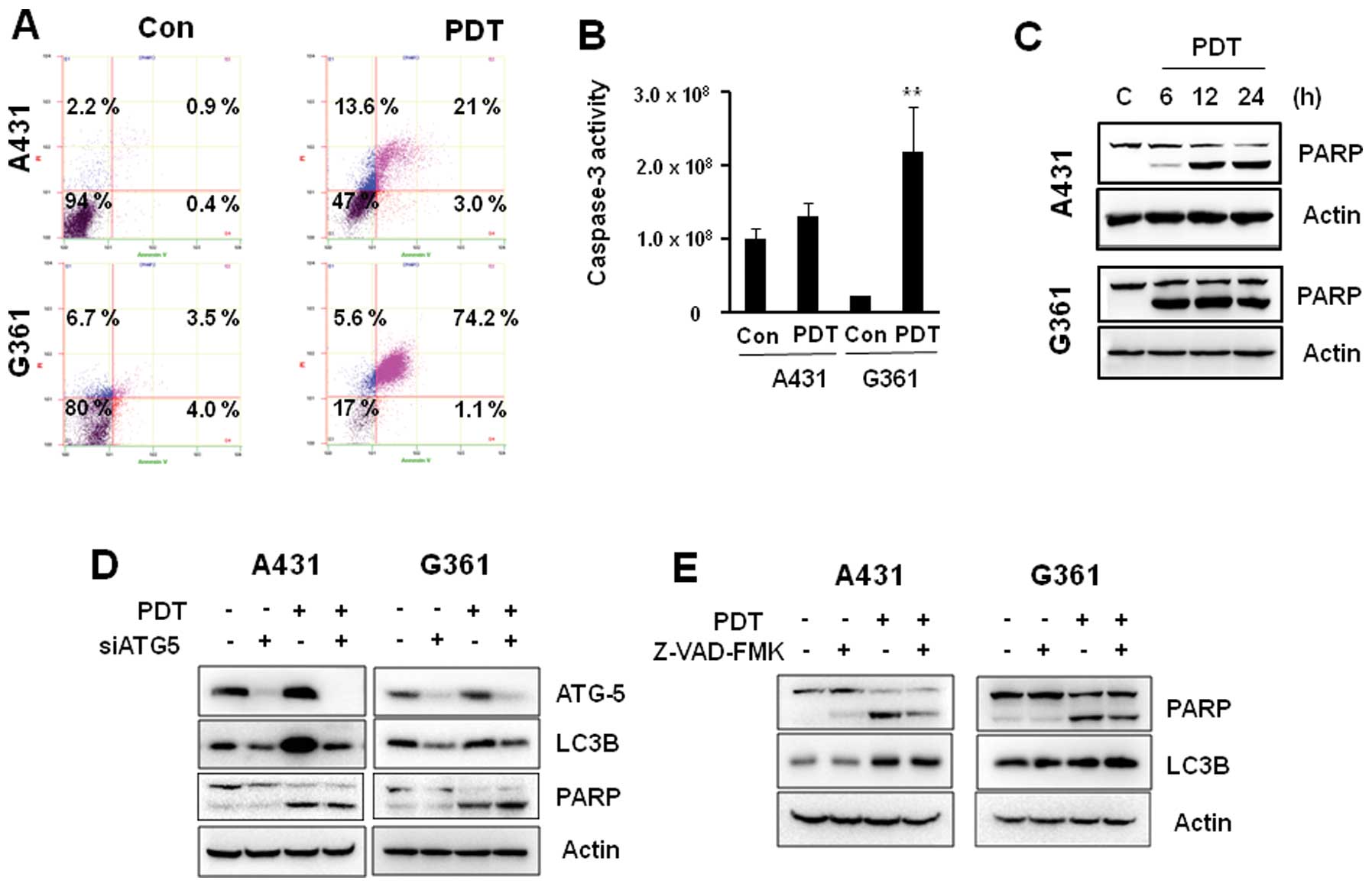
The effects of Pa-PDT on apoptosis
To determine whether the decrease in viability of
A431 and G361 cells was also caused by the induction of apoptosis,
we quantified apoptosis by flow cytometry using the Annexin
V-FITC/PI double staining assay. As shown in Fig. 5A, a significant number of apoptotic
cells (74.2%) was detected in G361 cells after Pa-PDT treatment,
whereas <24% apoptotic-positive cells were detected in A431
cells. Consistent with this observation, caspase-3 activity was
also strongly detected in G361 cells treated with Pa-PDT compared
to A431 cells (Fig. 5B). In
addition, the cleaved form of PARP was increased by Pa-PDT
treatment in a time-dependent manner in both cells. However, PARP
cleavage was stronger in G361 cells than in A431 cells (Fig. 5C).
To investigate potential cross-talk between
autophagy and apoptosis induction in response to Pa-PDT, we
investigated whether Pa-PDT-induced apoptosis could be inhibited by
siRNA against ATG-5. In A431 cells, Pa-PDT-induced LC3B expression
was inhibited by ATG-5 siRNA (Fig.
5D). However, Pa-PDT-induced PARP cleavage was not inhibited by
ATG-5 siRNA in A431 and G361 cells. In addition, 3MA, an early
stage inhibitor of autophagy, did not block Pa-PDT-induced PARP
cleavage (data not shown), suggesting that autophagy is unrelated
to Pa-PDT-mediated apoptosis.
Additionally, to determine whether Pa-PDT-induced
apoptosis affected the autophagy pathway, we examined the
expression levels of key autophagy proteins, LC3B, during treatment
with the caspase inhibitor zVAD-fmk. Pretreatment of cells with the
caspase inhibitor zVAD-fmk did not prevent Pa-PDT-induced
expression of LC3B proteins in A431 cells (Fig. 5E). Our data demonstrated that Pa-PDT
induces autophagy and/or apoptosis in A431 and G361 cells
independently.
The effect of Pa-PDT on tumor growth and
cell death in a skin cancer cell-xenograft CAM model
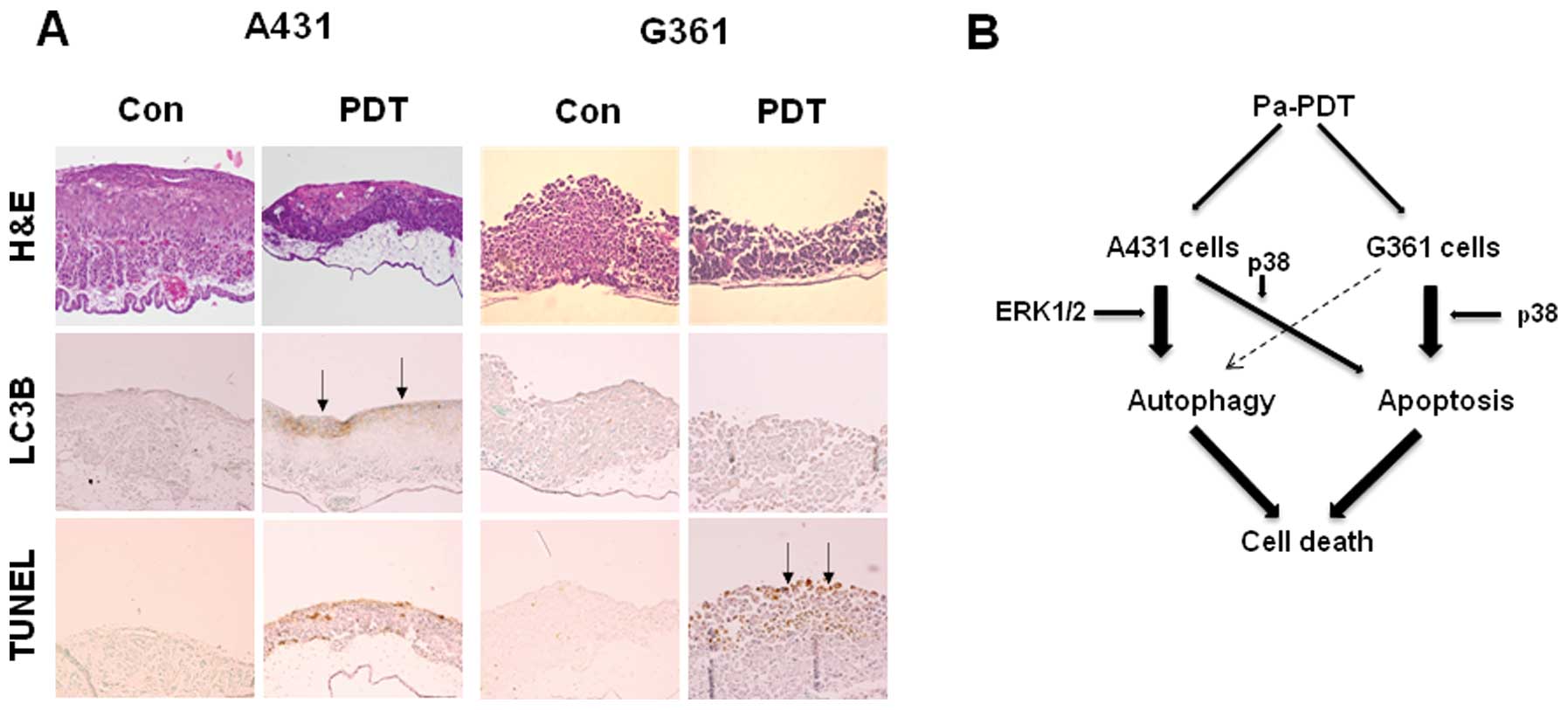
To identify the pathobiological characteristics of
the transplantation tumors in the CAM, hematoxylin-eosin (H&E)
staining and immunohistochemical analysis were performed.
Histological examination demonstrated that Pa-PDT
reduced tumor thickness and increased cell death in both
cell-xenograft CAM. Specifically, the expression of LC3B protein
was more strongly increased by Pa-PDT in A431-implanted tumors.
Additionally, TUNEL staining confirmed that TUNEL-positive cells
were more frequently observed in G361-implanted tumor sections
(Fig. 6A). These results
demonstrate that Pa-PDT leads to selective and effective cell death
of skin cancer cells through the autophagy and apoptosis
pathway.
Discussion
PDT is used worldwide not only in the primary
treatment for malignant and premalignant skin cancer but also for
adjuvant treatment in lung, brain, esophageal, biliary and urinary
tract cancer (2,14–16).
The rapid generation of ROS observed in PDT-treated cells has been
reported to induce damage to mitochondria or the endoplasmic
reticulum (ER) and lead to apoptosis (23,24).
Although apoptosis has been reported as the predominant cell death
modality after photosensitizer-mediated PDT, the mechanism of
PDT-mediated cell death is largely unknown.
The present study was designed to determine whether
Pa-PDT exhibits anticancer properties in non-melanoma A431 and
melanoma G361 skin cancer cells and to further investigate the
underlying mechanisms of Pa-PDT-induced cell death. We observed
that Pa-PDT predominantly induced autophagy in A431 cells, as
indicated by multiple independent approaches that either revealed
the expression of autophagy-specific proteins or the formation of
autophagic vacuoles. Pa-PDT induced Beclin 1-dependent autophagy
through ERK1/2 activation. By contrast, Pa-PDT mainly induced the
apoptotic pathway through p38 MAPK activation in G361 cells.
Previous studies demonstrated that the Akt/mTOR
pathway is the major pathway that regulates autophagy (25). The inhibition of Akt phosphorylation
and downstream mTOR signaling contribute to the initiation of
autophagy (26,27). We clearly demonstrated that Pa-PDT
treatment inhibits the phosphorylation of Akt and mTOR in A431
cells but not in G361 cells. These results demonstrated that
Pa-PDT-induced autophagy is mediated by the downregulation of
Akt/mTOR in A431 cells. In particular, the phosphorylation of
ERK1/2 and p38 MAPK was induced by Pa-PDT in A431 cells. In G361
cells, the activation of p38 MAPK was observed, whereas the
phosphorylation of ERK1/2 was not markedly induced by Pa-PDT. From
these results, we considered Pa-PDT-induced cell death to be
related to the p38 and/or ERK1/2 pathway in skin cancer cells. The
inhibition of ERK activity by the ERK inhibitor U0126 reduced
vesicle formation and LC3B expression in Pa-PDT treated A431 cells.
These results suggest that ERK1/2 activation could mediate
autophagy in Pa-PDT-treated A431 cells.
In addition, we observed PARP cleavage in
Pa-PDT-treated A431 and/or G361 cells. Notably, PARP cleavage was
suppressed by the p38 inhibitor, SB202190. The findings supported
the induction of p38-mediated apoptosis during Pa-PDT treatment.
The reason for the difference between A361 and G361 cell responses
to Pa-PDT treatment is unclear. However, the mechanism of action of
PDT might depend on the subcellular localization and molecular
targets of the photosensitizer, the metabolic potential and the
genotype of the tumor cell type.
It has been suggested that the autophagic response
observed in cells treated with diverse cytotoxic agents is involved
in protecting cells from apoptosis or, alternatively, is associated
with a mechanism contributing to apoptosis (28–30).
Despite these studies, the relationship between autophagy and
apoptosis remains poorly understood.
We investigated whether Pa-PDT-induced autophagy
leads to the activation of the apoptotic pathway. A431 and G361
cells were treated with Pa-PDT in the presence or absence of 3-MA
and chloroquine (CQ), specific inhibitors of the early and
late-stage autophagic process, respectively. 3-MA and CQ did not
inhibit the activation of PARP induced by Pa-PDT in either cell
line (data not shown). Similarly, the suppression of ATG-5 using
siRNA did not block Pa-PDT-induced PARP cleavage. In addition,
caspase inhibition did not inhibit Pa-PDT-induced autophagy-related
protein expression in A431 cells. These findings suggest that the
interplay between Pa-PDT-induced autophagy and apoptosis does not
exist in A431 and G361 cells.
Our previous study demonstrated the therapeutic
potential of Pa-PDT on YD-10B cells as a model of human oral
cancer. Our findings revealed that autophagy contributes to
Pa-PDT-mediated cell growth inhibition. In that study, Pa-PDT
reduced the phosphorylation of ERK, whereas the phosphorylation of
p38 and JNK were unchanged (31).
Therefore, the Pa-PDT-induced MAPK regulation may trigger different
intracellular cell death pathways in a cell type-specific
manner.
In conclusion, we demonstrated for the first time
that Pa-PDT induces autophagy and apoptosis in A431 cells.
Pa-PDT-induced cell death is carried out through ERK1/2-mediated
autophagy as well as apoptosis secondary to p38 MAPK activation.
Alternatively, the potent antitumor effect of Pa-PDT on G361 cells
was induced via p38-mediated caspase-3-dependent apoptotic
pathways. Therefore, Pa-PDT is a potential therapy for human skin
cancer and induces multiple death pathways via MAPK activation.
Acknowledgements
We thank Dr Se-Won Park for insightful discussion.
This study was supported by the National Research Foundation of
Korea (NRF) funded by the Ministry of Science, ICT & Future
Planning (No. R13-2008-010-00000-0). Dr J.-H. Yoon was supported by
a grant of the Korean Health Technology R&D Project, Ministry
for Health, Welfare & Family Affairs, Republic of Korea (No.
A100490).
References
|
1
|
Chen J, Keltner L, Christophersen J, Zheng
F, Krouse M, Singhal A and Wang SS: New technology for deep light
distribution in tissue for phototherapy. Cancer J. 8:154–163. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dolmans DE, Fukumura D and Jain R:
Photodynamic therapy for cancer. Nat Rev Cancer. 3:380–387. 2003.
View Article : Google Scholar
|
|
3
|
Yin X, Zhou J, Jie C, Xing D and Zhang Y:
Anticancer activity and mechanism of Scutellaria barbata
extract on human lung cancer cell line A549. Life Sci.
75:2233–2244. 2004.
|
|
4
|
Chan JY, Tang PM, Hon PM, Au SW, Tsui SK,
Waye MM, Kong SK, Mak TC and Fung KP: Pheophorbide a, a major
antitumor component purified from Scutellaria barbata,
induces apoptosis in human hepatocellular carcinoma cells. Planta
Med. 72:28–33. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakamura Y, Murakami A, Koshimizu K and
Ohigashi H: Inhibitory effect of pheophorbide a, a
chlorophyll-related compound, on skin tumor promotion in ICR mouse.
Cancer Lett. 108:247–255. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee WY, Lim DS, Ko SH, Park YJ, Ryu KS,
Ahn MY, Kim YR, Lee DW and Cho CW: Photoactivation of pheophorbide
a induces a mitochondrial mediated apoptosis in Jurkat leukaemia
cells. J Photochem Photobiol B. 75:119–126. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hajri A, Wack S, Meyer C, Smith MK,
Leberquier C, Kedinger M and Aprahamian M: In vitro and
in vivo efficacy of photofrin and pheophorbide a, a
bacteriochlorin, in photodynamic therapy of colonic cancer cells.
Photochem Photobiol. 75:140–148. 2002. View Article : Google Scholar
|
|
8
|
Jin ZH, Miyoshi N, Ishiguro K, Umemura S,
Kawabata K, Yumita N, Sakata I, Takaoka K, Udagawa T, Nakajima S,
Tajiri H, Ueda K, Fukuda M and Kumakiri M: Combination effect of
photodynamic and sonodynamic therapy on experimental skin squamous
cell carcinoma in C3H/HeN mice. J Dermatol. 27:294–306.
2000.PubMed/NCBI
|
|
9
|
Hajri A, Coffy S, Vallat F, Evrard S,
Marescaux J and Aprahamian M: Human pancreatic carcinoma cells are
sensitive to photodynamic therapy in vitro and in
vivo. Br J Surg. 86:899–906. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang PM, Chan JY, Au SW, Kong SK, Tsui SK,
Waye MM, Mak TC, Fong WP and Fung KP: Pheophorbide a, an active
compound isolated from Scutellaria barbata, possesses
photodynamic activities by inducing apoptosis in human
hepatocellular carcinoma. Cancer Biol Ther. 5:1111–1116.
2006.PubMed/NCBI
|
|
11
|
Tang PM, Zhang DM, Xuan NH, Tsui SK, Waye
MM, Kong SK, Fong WP and Fung KP: Photodynamic therapy inhibits
P-glycoprotein mediated multidrug resistance via JNK activation in
human hepatocellular carcinoma using the photosensitizer
pheophorbide a. Mol Cancer. 8:56–66. 2009. View Article : Google Scholar
|
|
12
|
Tang PM, Liu XZ, Zhang DM, Fong WP and
Fung KP: Pheophorbide a based photodynamic therapy induces
apoptosis via mitochondrial-mediated pathway in human uterine
carcinosarcoma. Cancer Biol Ther. 8:533–539. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ahn MY, Kwon SM, Kim YC, Ahn SG and Yoon
JH: Pheo-phorbide a-mediated photodynamic therapy induces apoptotic
cell death in murine oral squamous cell carcinoma in vitro
and in vivo. Oncol Rep. 27:1772–1778. 2012.PubMed/NCBI
|
|
14
|
Choudhary S, Nouri K and Elsaie ML:
Photodynamic therapy in dermatology. Lasers Med Sci. 24:971–980.
2009. View Article : Google Scholar
|
|
15
|
Brown SB, Brown EA and Walker I: The
present and future role of photodynamic therapy in cancer
treatment. Lancet Oncol. 5:497–508. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baldea I and Filip AG: Photodynamic
therapy in melanoma: an update. J Physiol Pharmacol. 63:109–118.
2012.
|
|
17
|
Calzavara-Pinton PG, Venturini M and Sala
R: Photodynamic therapy: update 2006 Part 1: Photochemistry and
photobiology. J Eur Acad Dermatol Venereol. 21:293–302. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim SA, Kwon SM, Kim JA, Kang KW, Yoon JH
and Ahn SG: 5′-Nitro-indirubinoxime, an indirubin derivative,
suppresses metastatic ability of human head and neck cancer cells
through the inhibition of Integrin β1/FAK/Akt signaling. Cancer
Lett. 306:197–204. 2011.
|
|
19
|
Jung CH, Ro SH, Cao J, Otto NM and Kim DH:
mTOR regulation of autophagy. Cancer Lett. 584:1287–1295.
2010.PubMed/NCBI
|
|
20
|
Paglin S, Hollister T, Delohery T, Hackett
N, McMahill M, Sphicas E, Domingo D and Yahalom J: A novel response
of cancer cells to radiation involves autophagy and formation of
acidic vesicles. Cancer Res. 61:439–444. 2001.PubMed/NCBI
|
|
21
|
Klotz LO, Fritsch C, Briviba K,
Tsacmacidis N, Schliess F and Sies H: Activation of JNK and p38 but
not ERK MAP kinases in human skin cells by
5-aminolevulinate-photodynamic therapy. Cancer Res. 58:4297–4300.
1998.PubMed/NCBI
|
|
22
|
Wu RW, Yow CM, Wong CK and Lam YH:
Photodynamic therapy (PDT): initiation of apoptosis via activation
of stress-activated p38 MAPK and JNK signal pathway in H460 cell
lines. Photodiagnosis Photodyn Ther. 8:254–263. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Buytaert E, Dewaele M and Agostinis P:
Molecular effectors of multiple cell death pathways initiated by
photodynamic therapy. Biochim Biophys Acta. 1776:86–107.
2007.PubMed/NCBI
|
|
24
|
François A, Marchal S, Guillemin F and
Bezdetnaya L: mTHPC-based photodynamic therapy induction of
autophagy and apoptosis in cultured cells in relation to
mitochondria and endoplasmic reticulum stress. Int J Oncol.
39:1537–1543. 2011.PubMed/NCBI
|
|
25
|
Shinojima N, Yokoyama T, Kondo Y and Kondo
S: Roles of the Akt/mTOR/p70S6K and ERK1/2 signaling pathways in
curcumin-induced autophagy. Autophagy. 3:635–637. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Iwamaru A, Kondo Y, Iwado E, Aoki H,
Fujiwara K, Yokoyama T, Mills GB and Kondo S: Silencing mammalian
target of rapamycin signaling by small interfering RNA enhances
rapamycin-induced autophagy in malignant glioma cells. Oncogene.
26:1840–1851. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim KW, Mutter RW, Cao C, Albert JM,
Freeman M, Hallahan DE and Lu B: Autophagy for cancer therapy
through inhibition of pro-apoptotic proteins and mammalian target
of rapamycin signaling. J Biol Chem. 281:36883–36890. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Levy JM and Thorburn A: Targeting
autophagy during cancer therapy to improve clinical outcomes.
Pharmacol Ther. 131:130–141. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen S, Rehman SK, Zhang W, Wen A, Yao L
and Zhang J: Autophagy is a therapeutic target in anticancer drug
resistance. Biochim Biophys Acta. 1806:220–229. 2010.PubMed/NCBI
|
|
30
|
Choi KS: Autophagy and cancer. Exp Mol
Med. 44:109–120. 2012. View Article : Google Scholar
|
|
31
|
Ahn MY, Yoon HE, Kwon SM, Lee J, Min SK,
Kim TC, Ahn SG and Yoon JH: Synthesized Pheophorbide a-mediated
photodynamic therapy induced apoptosis and autophagy in human oral
squamous carcinoma cells. J Oral Pathol Med. 43:17–25. 2013.
View Article : Google Scholar : PubMed/NCBI
|