Introduction
The most common type of brain tumor in adults is
glioma, which is extraordinarily lethal and is associated with
extremely low survival rates (1,2). For
most malignant forms of glioma, the median overall survival is only
12–15 months (2). The most common
as well as the most aggressive type of glioma is glioblastoma
multiforme (1). Glioblastoma
multiforme consists of heterogeneous types of cells which
demonstrate a variety of tumorigenic properties (1).
Gliomas arise from neural stem cells and progenitors
(1). These neural stem cells are a
subtype of astrocytes and are located in the two main regions of
the brain where neurogenesis takes place: the subventricular region
and the subgranular zone of hippocampal formation (2,3).
After cancer cells begin to propagate, they
construct their own microenvironment or niche (1). In this niche there is ‘crosstalk’
between the tumor and the host so that the niche and its
surroundings constantly interact (1,4).
One condition that promotes tumor growth is hypoxia
(5). Rapid growth of a tumor that
outpaces neovascularization results in a hypoxic niche (1). Hypoxia-inducible factor 1 (HIF-1) is
an important regulator of cellular responses to hypoxia (5). HIF-1 promotes glioma growth, mainly by
activating angiogenesis and by upregulating relevant target genes
(5).
Animal models are critical for investigating
malignant tumors. The orthotopic xenograft mouse model of
glioblastoma has been particularly valuable for analyzing the
characteristics of donor human tumor cells, but studies concerning
the role of host tissue and cells are sparse (6). Due to the progress achieved in the
research of cancer stem cells and the tumor microenvironment
(7), it has become obviously
necessary to have a comprehensive understanding of the role of
host-derived tissues and cells in tumor development. In recent
years, green fluorescent protein (GFP) nude mice have been widely
adopted for human cancer xenograft models with the main focus on
the tracing of tumor cells in the xenografts (8,9). The
aims of the present study were to develop a subcutaneous xenograft
GFP nude mouse model by injection of human glioma stem/progenitor
cell line SU3, and investigating the potential roles of host cells
in the repair of necrosis in xenografts. We successfully
established a model that facilitates the observation of how host
cells participate in the repair of tumor necrosis.
Materials and methods
Materials
The human glioma stem/progenitor cell line SU3 was
established according to previously published methods from a
surgical specimen of an adult male patient diagnosed with
glioblastoma multiforme (10). In
the present study, the human glioma stem/progenitor cell line SU3
was successfully obtained from fresh surgical specimens of GBM
patients. These patients underwent surgery at the Neurosurgical
Department, Second Affiliated Hospital of Suzhou University. The
present study was approved by the Medical Review Board of the
Suzhou University Medical School. The patients or their legal
guardians provided informed consent for the use of their surgical
specimen in research prior to sample acquisition. GFP nude
NC-C57BL/6J mice were bred and maintained in a specific
pathogen-free animal care facility. Animal experiments were
approved by the Medical Review Board of Suzhou University Medical
School, and all procedures were conducted in accordance with
Chinese laws governing animal care. The trocars, namely #8
anesthesia epidural puncture needles (Zhejiang Haisheng Medical
Instrument Co., Ltd., Hangzhou, China) were selected for the
passage of tumor tissue in vivo. A fluorescence flashlight
and fluorescence glasses were provided by NightSea Co. (http://www.nightsea.com/fplights.htm).
The frozen sections of the xenografts were observed by fluorescence
microcopy (Carl Zeiss Axio Observer A1; Thornwood, New York, USA).
Immunohistochemical antibodies against TNF-1α (ab1793), HIF-1α
(ab8366), Ki-67 (ab92742), CD68 (ab53444) and CD11b (ab64347) were
purchased from Abcam (Cambridge, UK).
Tumor xenograft model
The human glioma stem/progenitor cell line SU3 was
successfully obtained from fresh surgical specimens of GBM
patients. SU3 cells were cultured in Dulbecco’s modified Eagle’s
medium (DMEM) with 10% fetal bovine serum (FBS) and incubated at
37ºC in 98% humidified air containing 5% CO2. The SU3
cells in logarithmic growth phase in vitro were digested
into a single-cell suspension liquid, and the final cell density
was adjusted to 1×105/ml. A cell suspension, 150 μl per
mouse, was slowly and smoothly injected subcutaneously into the
right forelimb axilla of the GFP nude mice. The implanted tumor
tissue was dissected free of blood clots, washed and minced into
0.5-mm slices for tumor tissue grafting. For the tumor tissue
transplantation, we designed a simple but novel injection system,
which included a 24# trocar and a specially designed propeller
(1). The tumor tissue, 1
mm3 per mouse, was slowly and smoothly injected into the
right forelimb axilla of the GFP nude mice. The animal model
established through tumor tissue injection was regarded as the
first generation in the present study. The implanted tumors were
passed from animal to animal following the same procedure for 10
generations, which was the point at which model stability was
established. The short diameter (a) and long diameter (b) of the
xenografts were measured with calipers every 3 days, and the tumor
volume was calculated according to the equation: V (mm3)
= a2 × b/2. When cachexia occurred, the tumor-bearing
mice were sacrificed for tumor removal and examination.
Molecular pathology and GFP expression of
the tumors
We performed immunohistochemical staining to examine
the expression of tumor necrosis factor (TNF) (1:50),
hypoxia-inducible factor 1α (HIF-1α) (1:200), Ki-67 (1:1,000), CD68
(1:200) and CD11b (1:400) in the xenografts (1). The tumor-bearing mice and the
subcutaneous xenografts were examined with a fluorescence
flashlight and fluorescence glasses. Cells expressing GFP in the
tumor tissue were examined using 5-μm frozen sections by
fluorescence microscopy at a wavelength of 470 nm.
Results
General characteristics of the
xenografts
In the primary xenograft models (SU3 cell suspension
injection), although the tumorigenicity rate was 100% (5/5), there
existed obvious individual differences in incubation period and
tumor volume. In conjunction with the tumor passage through tumor
tissue injection, these differences gradually narrowed, and the
animal model became reliable beginning with the 7th generation.
Subcutaneous seeded tumors can grow quite large; the xenograft
volume of the 9th generation, one month after inoculation, was
~9.96±4.19 cm3 (n=10) in average and the implanted
tumors continued to grow for more than 2 months. When all the
tumor-bearing mice were sacrificed in the late stage (approximately
2 months after tumor inoculation), the tumor weight reached almost
one half of the body weight of the mice, namely, 11.23±3.72 g
(n=10).
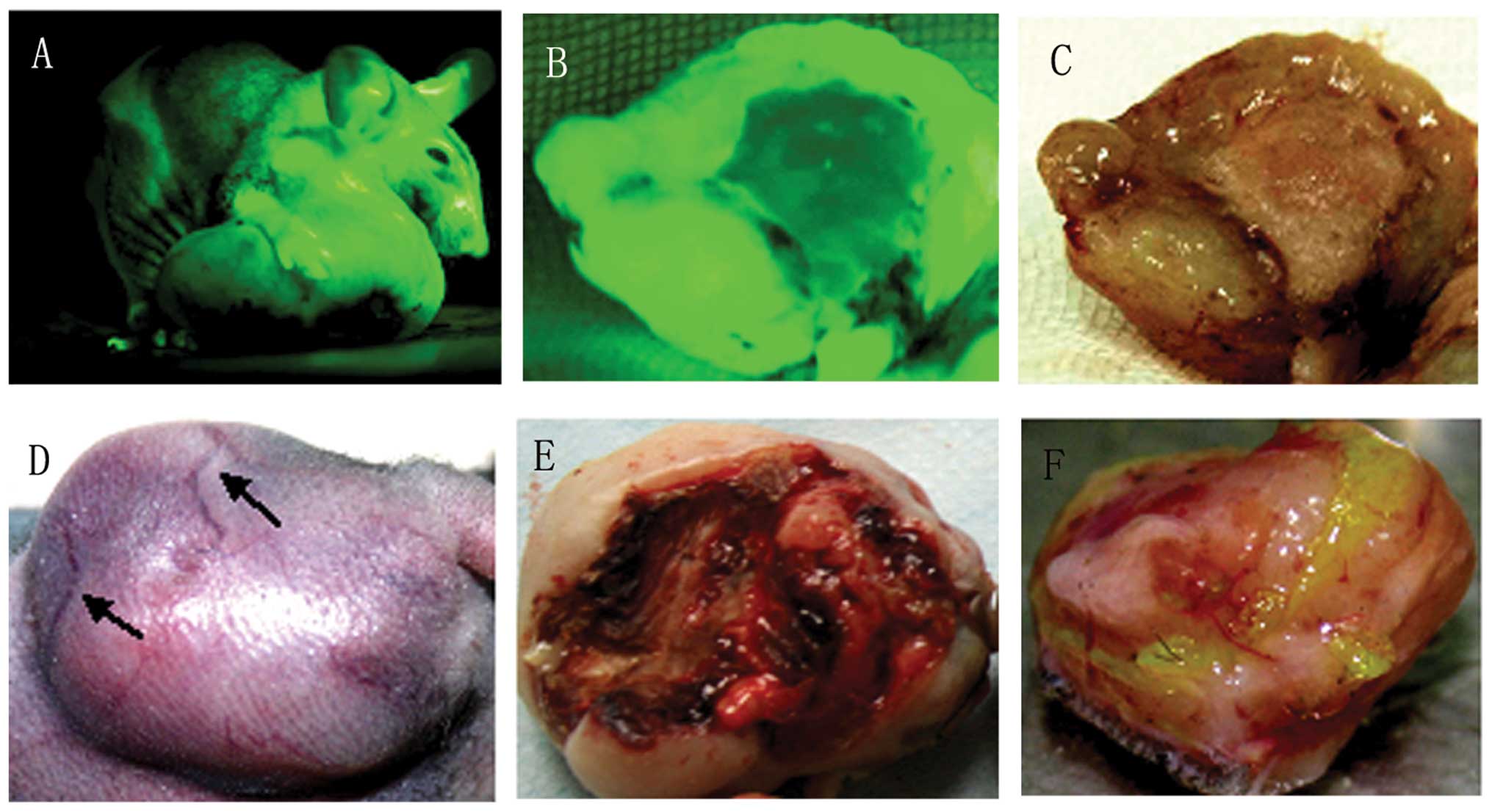
Under a fluorescence flashlight, the tumor-bearing
mice and the xenografts appeared green in color (Fig. 1A and B). Viewed with the naked eye
under natural light, there was enlargement of the xenografts, and
cutaneous vessels of the skin covering tumors were gradually
increased in size, meeting the demand of the blood supply required
by the rapidly growing tumor tissue (Fig. 1C and D). In the middle stage of
xenograft growth, ischemic necrosis, even to the extent of
ulceration, was observed in the local skin, possibly indicating the
insufficiency of the blood supply to the implanted tumors (Fig. 1E); there was still fresh tissue in
the tumor base (Fig. 1F).
Histopathologic characteristics of the
xenografts
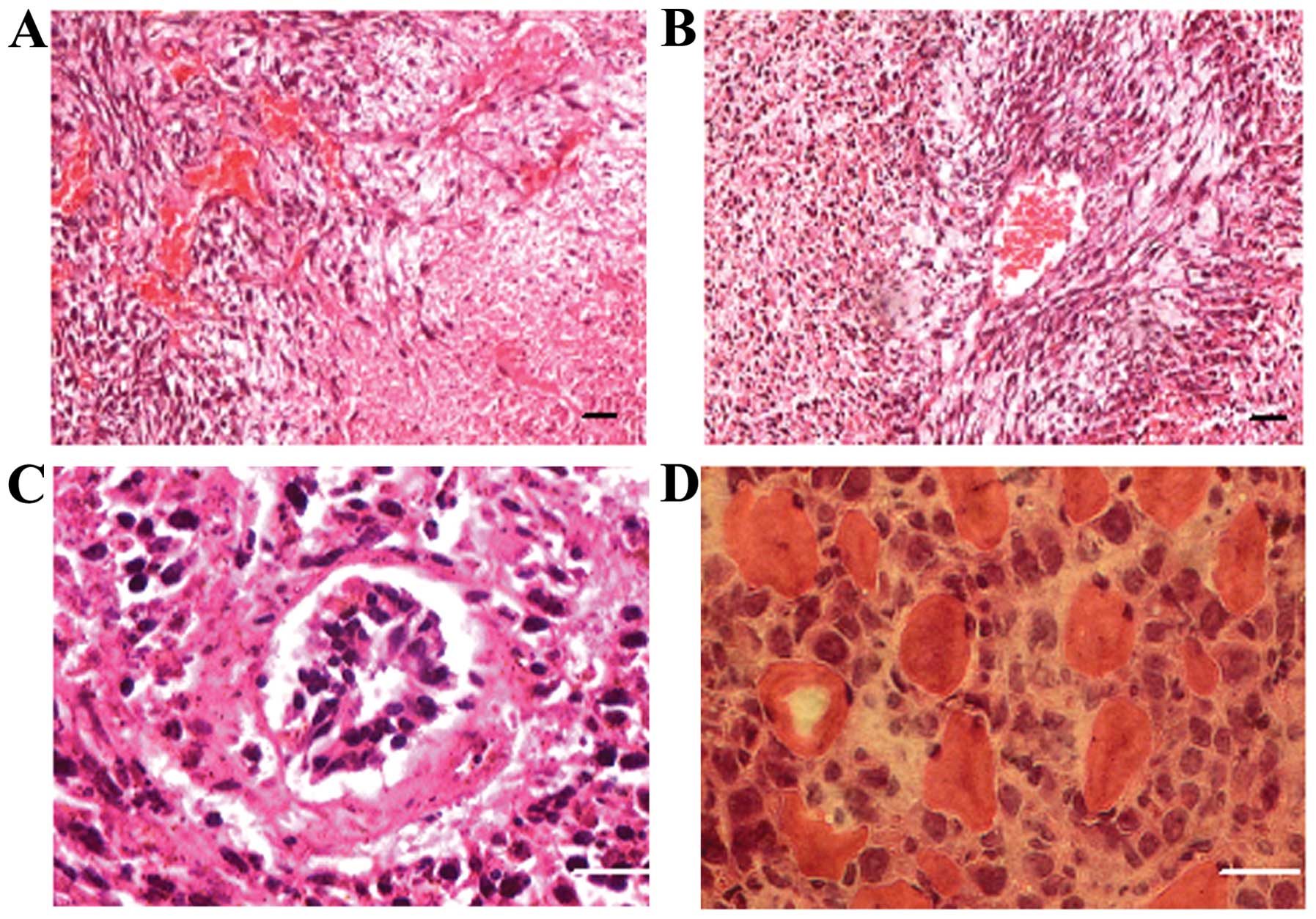
In the xenograft tumor sections stained with
hematoxylin and eosin (H&E), hyperchromatic nuclei, relatively
less cytoplasm, heteromorphic nuclei and regional densely arranged
tumor cells were observed (Fig. 2).
Necrotizing hemorrhage was frequently observed in the xenograft
sections and was divided into fresh hemorrhage and old hemorrhage
(Fig. 2A). In the fresh hemorrhagic
foci, a large number of collapsing red blood cells could still be
observed. In the old hemorrhagic areas, two scenes were observed:
i) red blood cells disappeared while some inflammatory cells were
observed; and ii) there were no inflammatory cells, however, tumor
cells with relatively few mesenchymal components were noted growing
around the vessels. Generally, the xenograft tumors were rich in
blood vessels, but a vast majority of new blood vessels lacked
complete vascular structure. Only a few large vessels possessed a
nearly intact vascular wall. In rare instances, clusters of
nucleated cells were noted in the vascular lumen. Tumor cells could
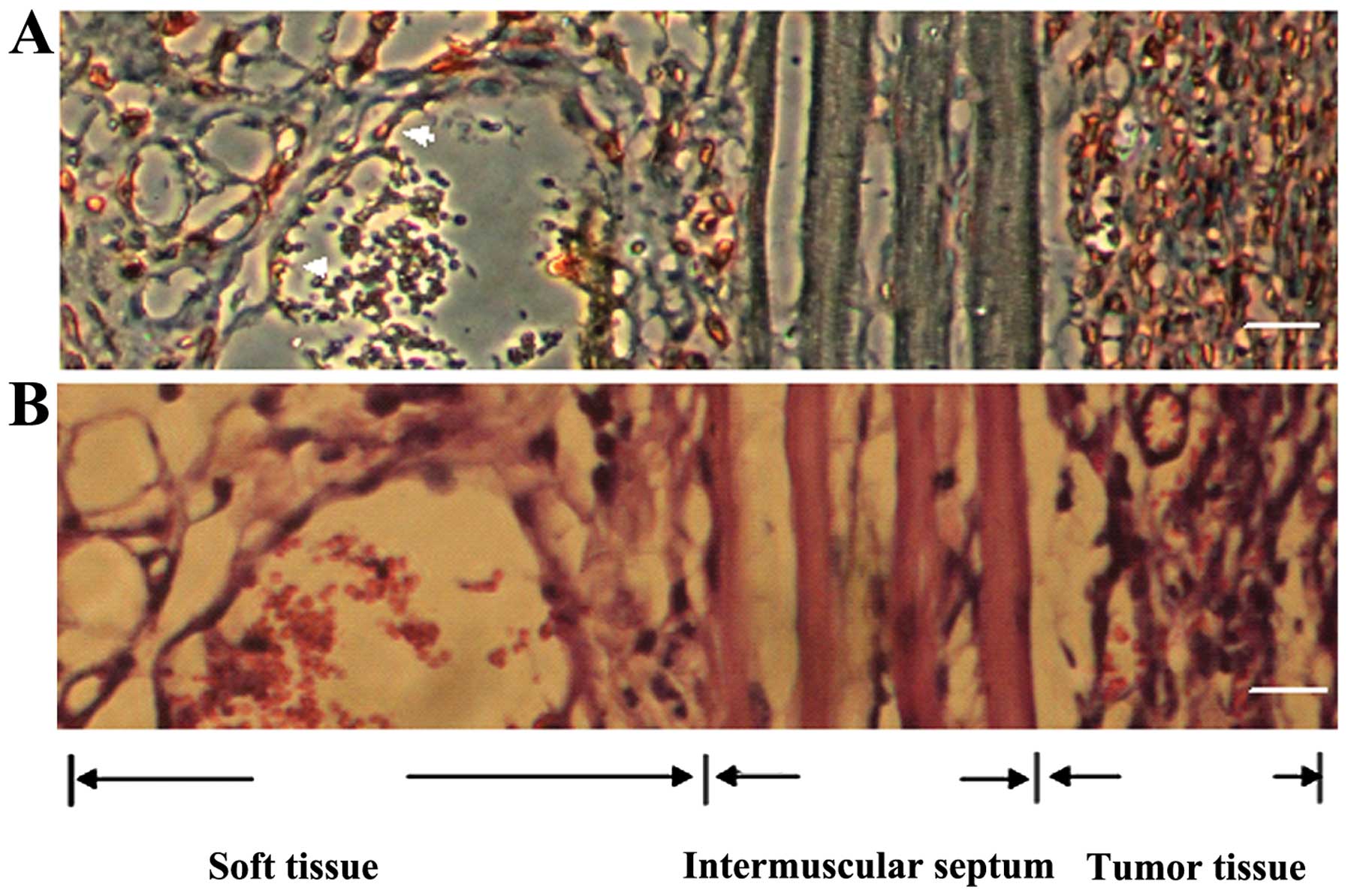
be seen invading soft tissue and muscle fiber (Fig. 2D).
Molecular pathology of the
xenografts
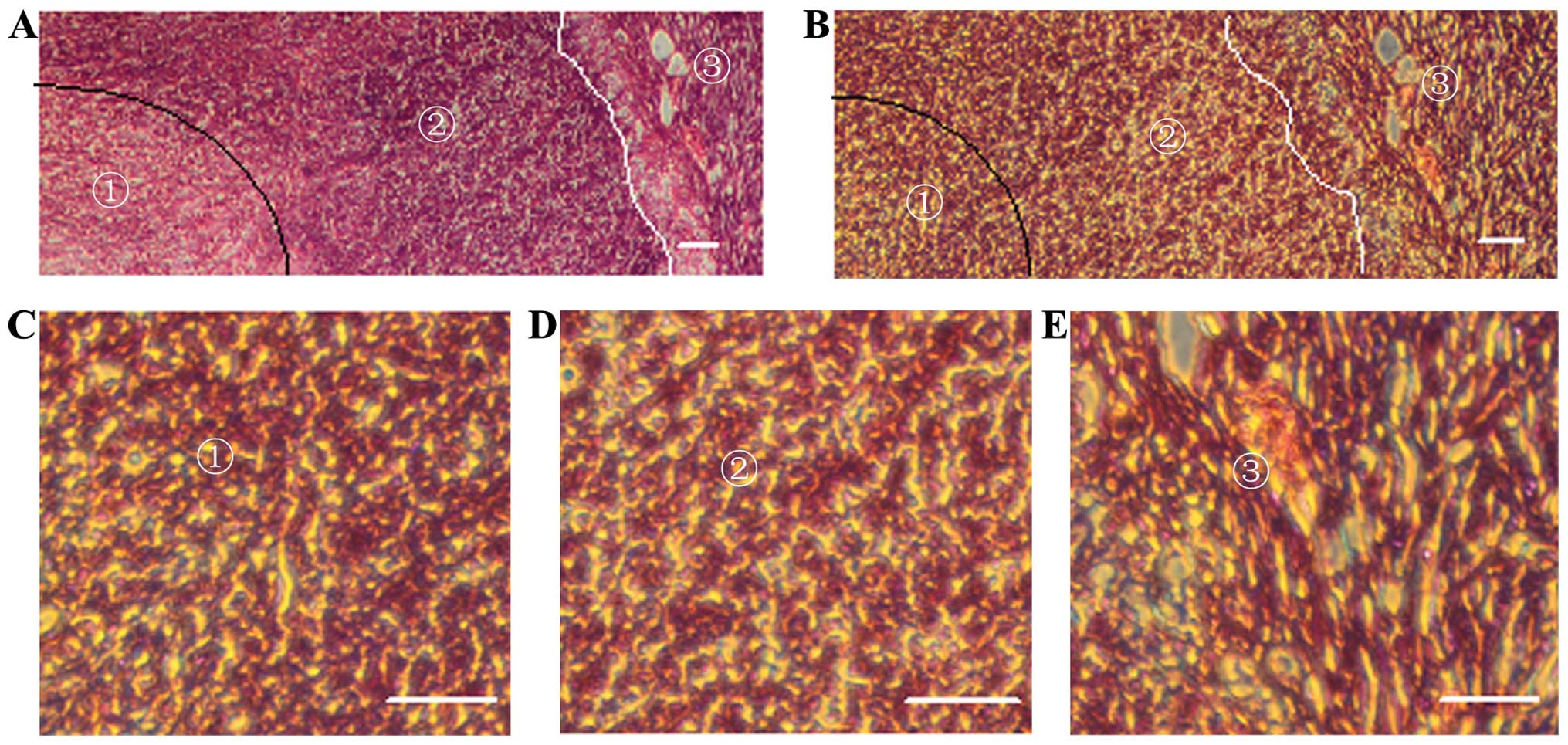
Hypoxia-inducible factor 1α was strongly expressed
in the nuclei of the tumor cells (Fig.
3A and B). Generally, the expression of HIF-1α was high in the
central portion of the tumor and low in the marginal zones
(Fig. 3C and D); however, notably,
invasive tumor cells migrating from the marginal zones also highly
expressed HIF-1α (Fig. 4). In the
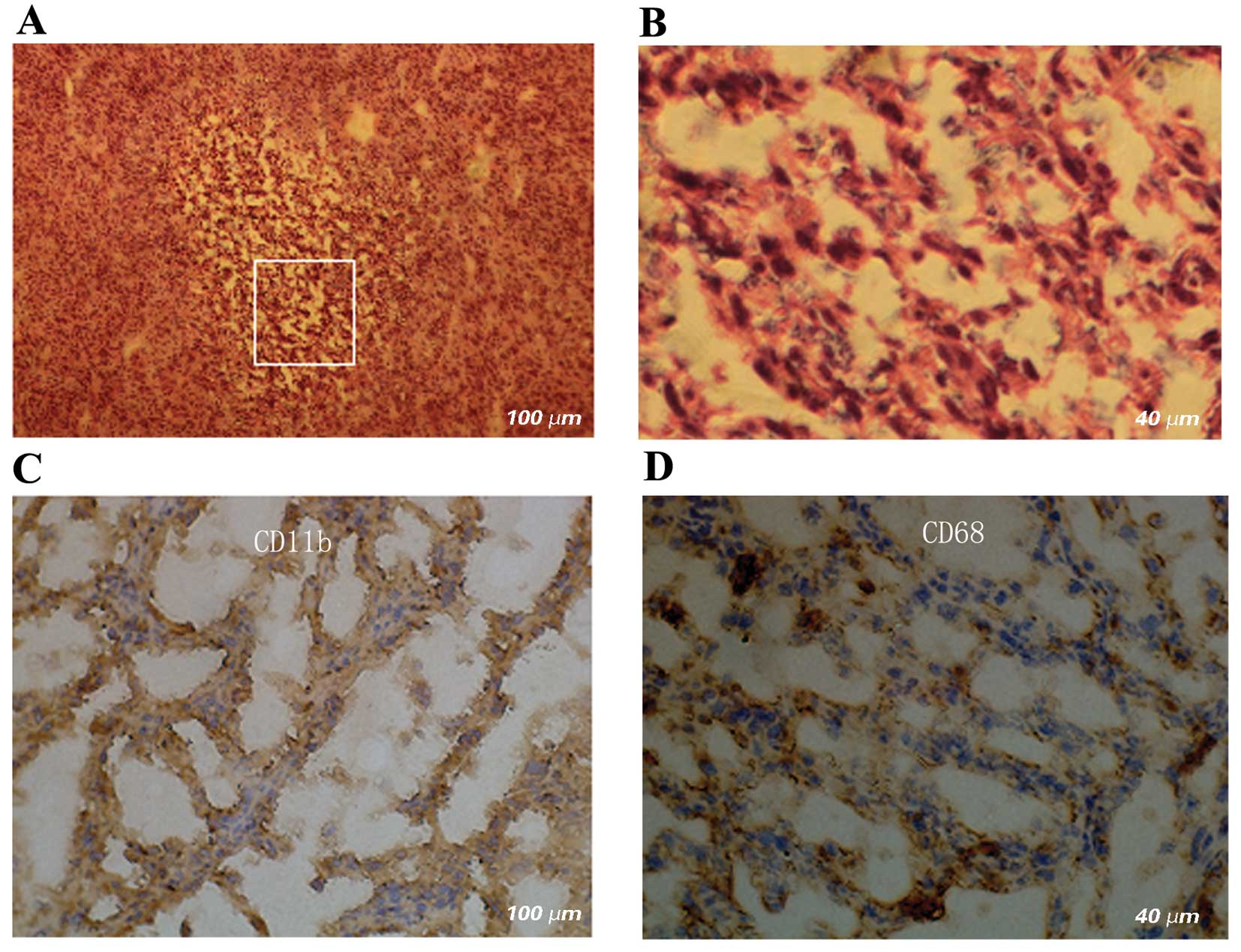
necrotic areas of the tumors, TNF was mainly expressed in shapeless
intercellular fluid; however, in the non-necrotic areas of the
tumors, TNF was diffusedly expressed on the membrane of tumor cells
rather than in the intercellular fluid (Fig. 5). The vast majority of tumor cells,
regardless of cell type, including those in the necrotic areas,
strongly expressed Ki-67 in the nuclei (Fig. 6). In old hemorrhagic areas, the
majority of cells were positively stained for CD11b and CD68, which
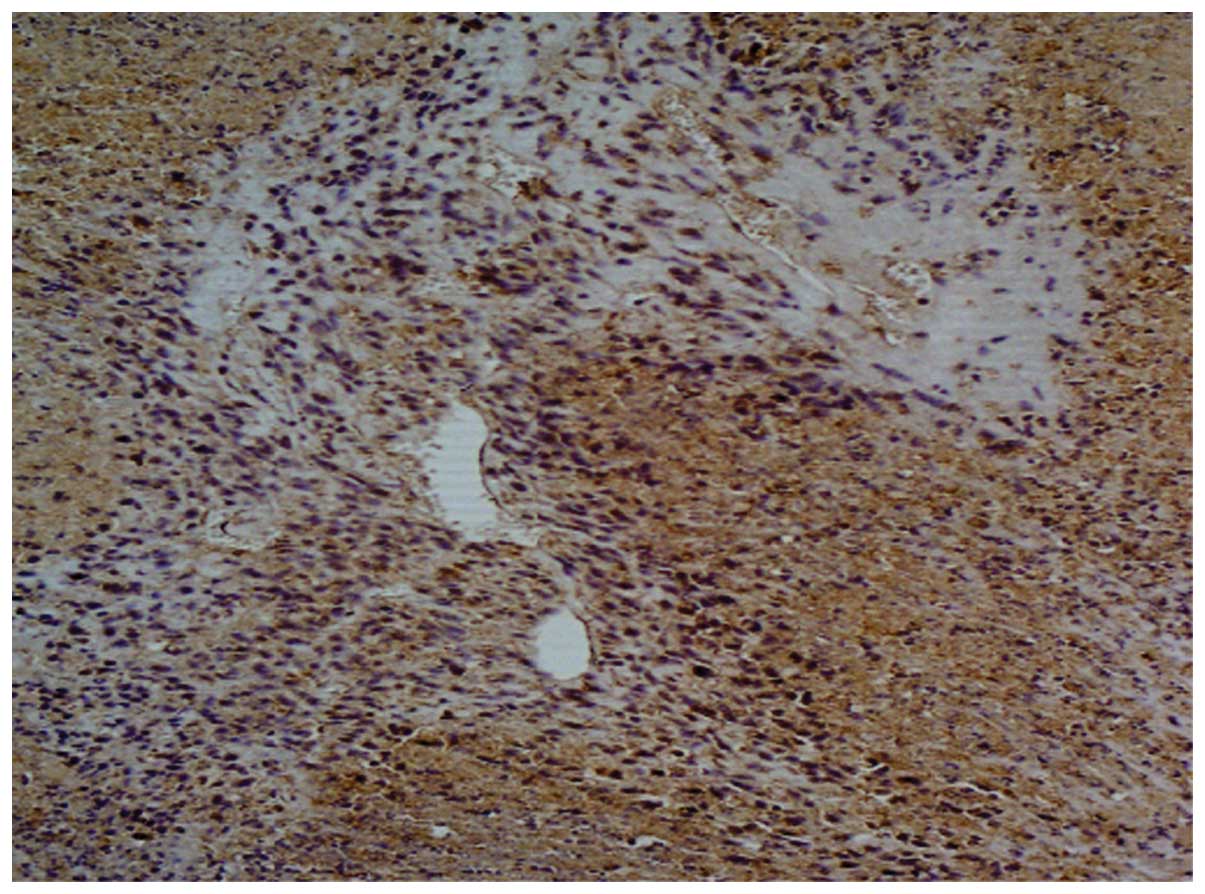
were expressed both in the cytoplasm and on the membrane (Fig. 7).
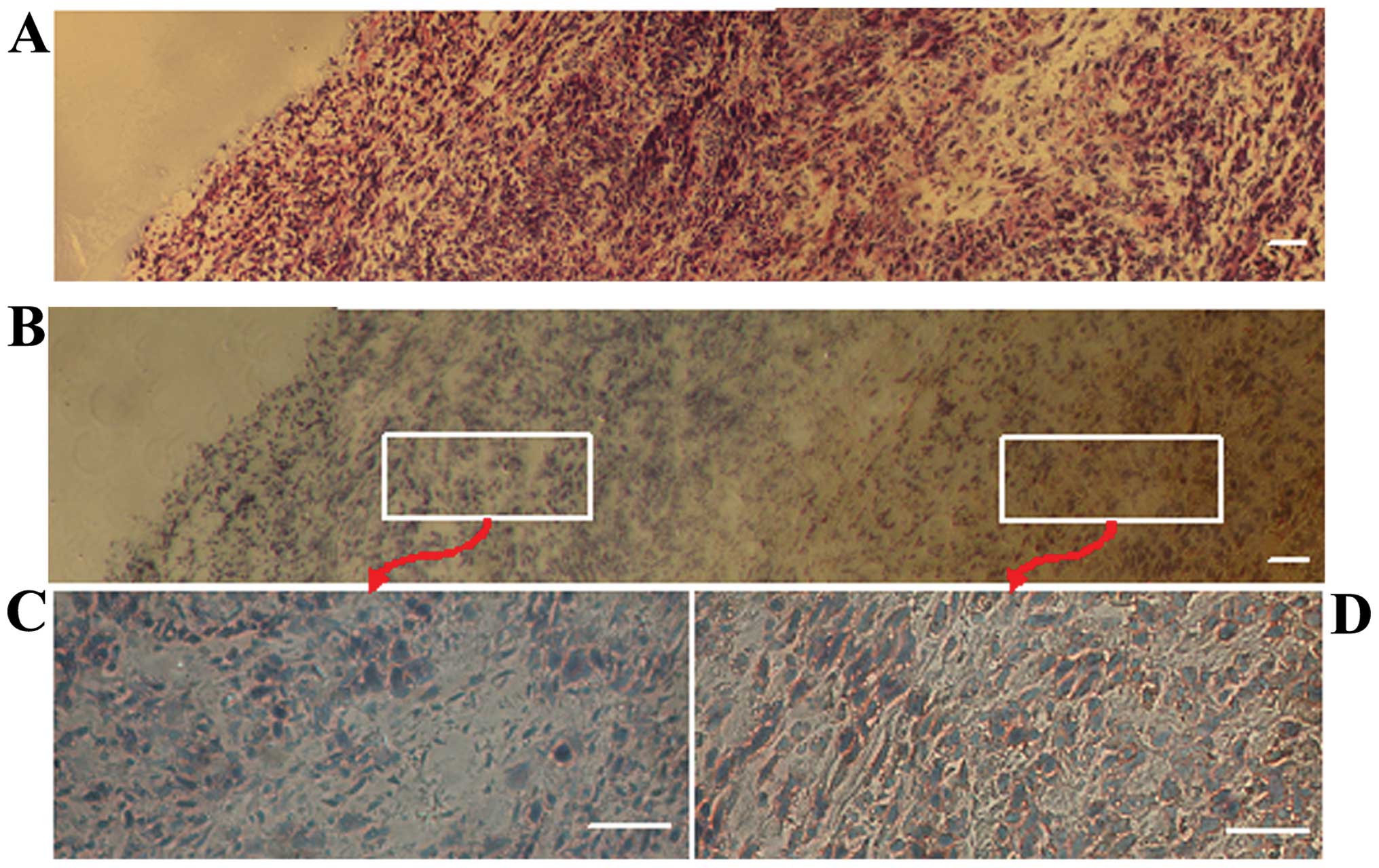
Green fluorescence components in the
xenografts
As the host organs, tissues and cells were
GFP-expressing, all components in the xenografts displayed green
fluorescence under fluorescence microscopy at an excitation
wavelength of 470 nm and were regarded to be derived from the host.
At the same time, tumor cells did not stimulate green fluorescence
under fluorescence microscopy at an excitation wavelength of 470
nm. Based on this concept, it was obvious that the implanted tumors
were covered by host-derived skin, and the green area surrounding
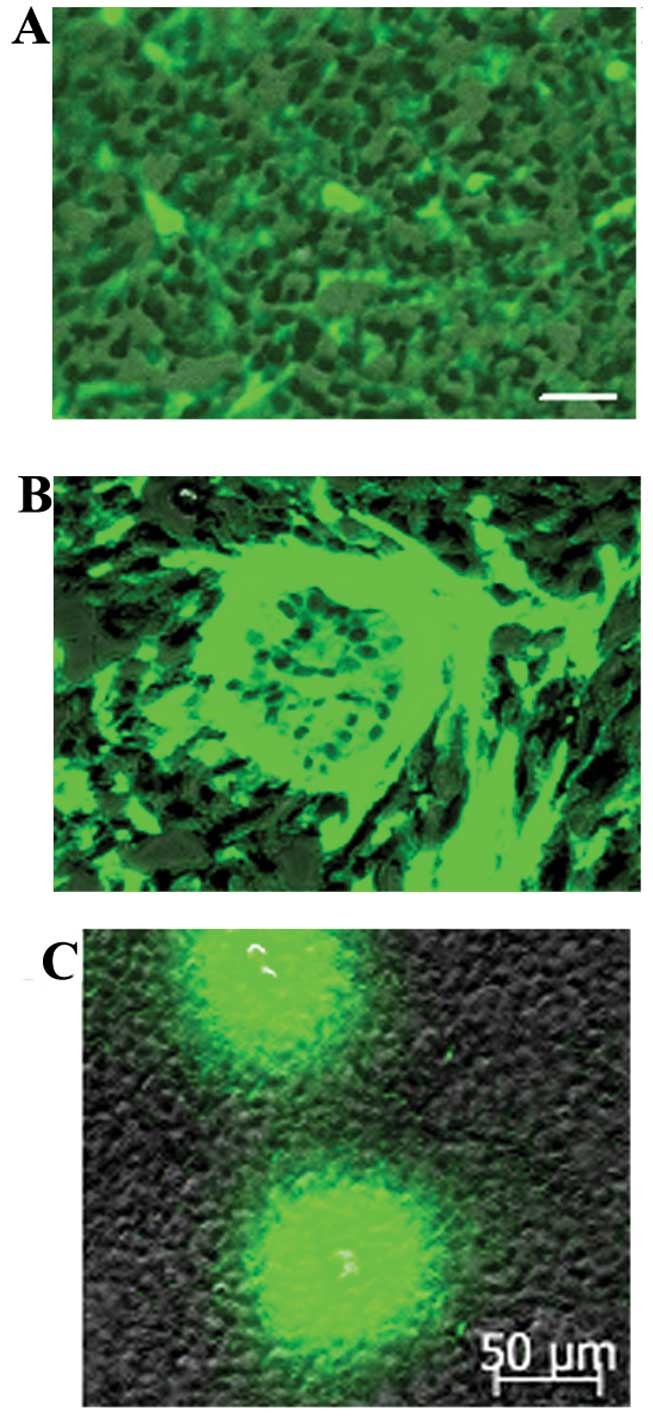
the tumor mass mainly consisted of host components (Fig. 1A and B). Notably, a large number of
GFP-expressing cells were found clustered in the necrotic areas of
the tumors (Fig. 8). GFP-expressing
cells that had a similar appearance to macrophagocytes were also
found scattered among the tumor parenchyma (Fig. 9).
Discussion
In the present study, we developed an animal model
for studying the role of host cells in glioma. We used a
subcutaneous xenograft GFP nude mouse model and passed implanted
tumors from animal to animal for 10 generations. The xenografts,
which came from the human glioma stem/progenitor cell line SU3,
were rich in blood vessels, necrosis and hemorrhagic foci. In the
interstitial tissue of the host, we noted numerous cells expressing
GFP, CD68 and CD11b. Based on our findings we believe this model
has significant potential for application in the future research of
tumor tissue remodeling and the tumor niche.
Hypoxia inducible factor 1α (HIF-1α)
Hypoxia-inducible factor 1 plays an important role
in tumor growth by being a key regulator of cellular responses to
hypoxia (5,11,12).
It consists of α and β subunits (5). In hypoxic conditions, the α subunit is
stable and in normoxia it is rapidly degraded (5). HIF-1 is translocated to the cell
nucleus and induces transcription of its downstream target genes
such as vascular endothelial growth factor and erythropoietin.
Hypoxia-inducible factor 1 has been reported to play
an important role in tumor growth and metastasis by regulating
energy metabolism and inducing angiogenesis to allow survival in
cellular hypoxia. Increased levels of HIF-1 have been noted in
breast carcinogenesis, lung cancer, prostatic cancer, gastric
carcinoma and cervical cancer (13–16).
Therefore, it was necessary to evaluate the expression of HIF-1 in
the assessment of our animal model. HIF-1 expression in our tumor
model showed two features: i) HIF-1α was highly expressed in the
central portion of the tumor and expression was low in the tumor
margin which was mainly due to low oxygen conditions in the central
portion of the tumor (17); and ii)
tumor cells invading the host tissue where oxygen is sufficient
also highly expressed HIF-1. We conjecture that the high expression
of HIF-1 in non-hypoxic host tissue was associated with the
disorder of tumor-suppressor genes (18). With regard to expression of HIF-1,
this tumor animal model meets the required criterion.
Expression of tumor necrosis factor
(TNF)
Tumor necrosis factor was originally discovered in
hemorrhagic necrotic tissue removed from tumor-bearing mice. TNF,
which consists of TNF-α and TNF-β, was found to be secreted by
macrophages and leukomonocytes. TNF-α is a type II membrane protein
and contains soluble TNF-α and membrane-related TNF-α. The effect
of TNF-α is concentration-dependent and type-dependent, including
the induction of tumor cell apoptosis (19) and immune surveillance (20) or the effects are opposite depending
on dose. For example, at high doses, TNF exerts a degenerative
effect on tumor vasculature and induces tumor hemorrhage and
necrosis; at moderate doses, TNF does not have cytotoxic effects on
tumor cells, but stimulates the proliferation and metastasis of
tumor cells by inducing angiogenesis and promoting secretion of
matrix metalloproteinase and endothelial adhesion molecule
(21,22). In tumor necrotic areas of our animal
model, TNF appeared to have the characteristic of sTNF-α; it was
highly expressed in shapeless intercellular fluid, distributed
lamellarly and without specific cell morphology. As there were
numerous immune cells infiltrating into tumor necrotic areas,
sTNF-α was most likely secreted by these immune cells (Fig. 8). Nevertheless, in the non-necrotic
areas of the tumors, TNF appeared to have the characteristic of
mTNF-α; it was diffusely expressed on the membrane of tumor cells.
For this reason, mTNF-α was possibly secreted by tumor cells. In
brief, the expression of TNF-α in this subcutaneous GFP nude mouse
model conformed to the features of malignant tumors.
Tumor microenvironment
The tumor microenvironment, also called the tumor
niche, is composed of interstitial fluid and various types of cells
(inflammatory cells, endothelial cells and fibroblasts). It is
known that in the early stage, the host-derived microenvironment
exerts suppressive effects on the growth of transplanted tumors due
to transplant rejection; however, in conjuction with the growth of
transplanted tumors, the microenvironment changes in favor of tumor
growth. This phenomenon has become a topic of extreme importance in
oncology research during the last few decades. In the early years
of research, Bettinger et al (23) reported that microglia can promote
glioma migration and Markovic et al (24) reported that when microglia were
selectively depleted, the invasiveness of tumors was significantly
decreased. In recent years, Wu et al (25) reported that macrophages/microglia
have the potential to promote the proliferation of tumor cells.
Straussman et al (26)
reported that when co-cultured with stromal cells, tumor cells
become more resistant in vitro to chemotherapy. Taken
together, these studies indicate that the host-derived tumor
microenvironment plays a critical role in tumor development and
progression. In another in vitro study of the tumor
microenvironment, Borovksi et al (27) used a co-culture system comprising
primary brain tumor microvascular endothelial cells and glioma
propagating cells obtained from glioblastoma multiforme biopsies.
They found that glioma propagating cells had higher proliferation
rates when co-cultured with primary brain tumor microvascular
endothelial cells than when cultured alone. However, in
vitro experiments are lacking in the ability to demonstrate the
importance of the tumor microenvironment.
In vivo experiments were performed by
Najbauer et al (4) who used
orthotopic rodent models of human glioma xenografts and reported
that a large number of host cells expressing nestin migrate to
glioma and contribute to the growth of the xenograft by assembling
into the microvasculature of glioma. It was determined that the
host cells originated from the subventricular zone ipsilateral to
the xenografts. The advantage of the GFP nude mouse model
established in the present study is that it is convenient for
tracing host cells and investigating the role of host cells in
tumor growth. For example, a large amount of GFP-expressing cells
were found clustered in old necrotic areas, and the majority of
cells expressed CD11b and CD68, which are inflammatory cells that
can serve as markers for macrophage-specific cells. Thus, we
deduced that host cells are critical component of the tumor
microenvironment that play a role in the repair of tumor necrosis.
As determined by the immunochemical staining, numerous TNF-α immune
complexes were noted in the necrotic areas of the tumors (Fig. 5). This indicated that TNF-α secreted
by macrophages participates in the composition of the cell factor
microenvironment. A large number of cells expressing Ki-67 was
noted in both the tumor necrotic areas and non-necrotic areas
(Fig. 6). Ki-67 is a nuclear
proliferation antigen and its expression reflects the proliferative
ability of cells, and for tumor cells its expression also predicts
malignancy and poor prognosis. Since Ki-67-expressing cells
included GFP cells, this added more evidence to support the
assumption that host-derived cells are involved in the repair of
tumor necrosis. Due to the participation of host cells in the
repair of necrosis of tumor tissue, no cyst was discovered in our
more than 100 tumor specimens.
de Almeida Sassi et al (1) recently reviewed studies on glioma
including the extensive research that has been carried out to
describe the tumor microenvironment. Importantly, they revealed
that there is a reciprocal relationship between glioma stem cells
and their microenvironment; that is, glioma stem cells can modulate
the microenvironment that produces signals that regulate glioma
stem cells. They also pointed out that tumor microvasculature
generates niches that promote the establishment and maintainence of
brain tumor stem cells. In addition, niches were found to have an
important role in protecting these stem cells from environmental
insults (e.g., chemotherapeutic agents). Recently, Gazdzinski and
Neiman (28) injected GL261 and 4C8
glioma cells that were labeled with iron oxide particles or with a
fluorescent probe into the brains of synergeneic mice. Following
tumor development, they used texture analysis to analyze the label
distribution patterns in the two types of gliomas. They reported
that in the GL261 tumors the label was observed mainly in the tumor
core whereas in the 4C8 tumors the label was more randomly
distributed throughout the tumor. They speculated that differences
in the label distribution maps between the two different types of
gliomas were likely affected by differences in the tumor
microenvironment.
In conclusion, a glioma model consisting of GFP nude
mice as the host was successfully established. With this model, we
were easily able to observe how host cells participate in tumor
necrosis repair. Since the host cells constantly express GFP, this
model has great potential for investigating the tumor
microenvironment as a supporting role in tumor growth and
progression and may serve to identify future therapeutic
targets.
Acknowledgements
The present study was funded by the National Natural
Scientific Foundation of China (nos. 81071766; 81172400; 81272799;
and 81272793).
References
|
1
|
de Almeida Sassi F, Brunetto AL,
Schwartsmann G, Roesler R and Abujamra AL: Glioma revisited: from
neurogenesis and cancer stem cells to the epigenetic regulation of
niche. J Oncol. 2012:5378612012. View Article : Google Scholar
|
|
2
|
He H, Li MU and Niu CS: The pathological
characteristics of glioma stem cell niches. J Clin Neurosci.
19:121–127. 2012. View Article : Google Scholar
|
|
3
|
Tavazoie M, Van der Veken L, Silva-Vargas
V, et al: A specialized vascular niche for adult neural stem cells.
Cell Stem Cell. 3:279–288. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Najbauer J, Huszthy PC, Barish ME, et al:
Cellular host responses in gliomas. PLoS One. 7:e351502012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kaur B, Khwaja FW, Severson EA, Matheny
SL, Brat DJ and Van Meir EG: Hypoxia and the
hypoxia-inducible-factor pathway in glioma growth and angiogenesis.
Neuro Oncol. 7:134–153. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fei XF, Zhang QB, Dong J, et al:
Development of a clinically relevant orthotopic xenograft mouse
model of metastatic lung cancer and glioblastoma through surgical
tumor tissue injection with trocar. J Exp Clin Cancer Res.
29:842010. View Article : Google Scholar
|
|
7
|
Charles NA, Holland EC, Gilbertson R,
Glass R and Kettenmann H: The brain tumor microenvironment. Glia.
60:1169–1180. 2011. View Article : Google Scholar
|
|
8
|
Hoffman RM: Green fluorescent protein
imaging of tumor cells in mice. Lab Animal. 31:34–41.
2002.PubMed/NCBI
|
|
9
|
Farin A, Suzuki SO, Weiker M, Goldman JE,
Bruce JN and Canoll P: Transplanted glioma cells migrate and
proliferate on host brain vasculature: a dynamic analysis. Glia.
53:799–808. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wan Y, Fei XF, Wang ZM, et al: Expression
of miRNA-125b in the new highly invasive glioma stem cell and
progenitor cell line SU3. Chin J Cancer. 31:207–214. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gillespie DL, Whang K, Ragel BT, Flynn JR,
Kelly DA and Jensen RL: Silencing of hypoxia inducible factor-1α by
RNA interference attenuates human glioma cell growth in
vivo. Clin Cancer Res. 13:2441–2448. 2007.
|
|
12
|
Ryan HE, Poloni M, McNulty W, Elson D,
Gassmann M, Arbeit JM and Johnson RS: Hypoxia-inducible factor-1α
is a positive factor in solid tumor growth. Cancer Res.
60:4010–4015. 2000.
|
|
13
|
Bos R, Van der Groep P, Greijer AE, et al:
Levels of hypoxia-inducible factor-1α independently predict
prognosis in patients with lymph node negative breast carcinoma.
Cancer. 97:1573–1581. 2003.
|
|
14
|
Zhong H, Semenza GL, Simons JW and De
Marzo AM: Up-regulation of hypoxia-inducible factor 1α is an early
event in prostate carcinogenesis. Cancer Detect Prev. 28:88–93.
2004.
|
|
15
|
Birner P, Schindl M, Obermair A, Plank C,
Breitenecker G and Oberhuber G: Overexpression of hypoxia-inducible
factor 1 α is a marker for an unfavorable prognosis in early-stage
invasive cervical cancer. Cancer Res. 60:4693–4696. 2000.
|
|
16
|
Urano N, Fujiwara Y, Doki Y, et al:
Overexpression of hypoxia-inducible factor-1 alpha in gastric
adenocarcinoma. Gastric Cancer. 9:44–49. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Clottes E: Hypoxia inducible factor 1:
regulation, involvement in carcinogenesis and target for anticancer
therapy. Bull Cancer. 92:119–127. 2005.(In French).
|
|
18
|
Xia Y, Choi HK and Lee K: Recent advances
in hypoxia-inducible factor (HIF)-1 inhibitors. Eur J Med Chem.
49:24–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lyu MA and Rosenblum MG: The
immunocytokine scFv23/TNF sensitizes HER-2/neu-overexpressing
SKBR-3 cells to tumor necrosis factor (TNF) via up-regulation of
TNF receptor-1. Mol Cancer Ther. 4:1205–1213. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ebert LM, Meuter S and Moser B: Homing and
function of human skin γδ T cells and NK cells: relevance for tumor
surveillance. J Immunol. 176:4331–4336. 2006.
|
|
21
|
Warren MA, Shoemaker SF, Shealy DJ, Bshara
W and Ip MM: Tumor necrosis factor deficiency inhibits mammary
tumorigenesis and a tumor necrosis factor neutralizing antibody
decreases mammary tumor growth in neu/erbB2 transgenic mice. Mol
Cancer Ther. 8:2655–2663. 2009. View Article : Google Scholar
|
|
22
|
Yin Y, Chen X and Shu Y: Gene expression
of the invasive phenotype of TNF-α-treated MCF-7 cells. Biomed
Pharmacother. 63:421–428. 2009.
|
|
23
|
Bettinger I, Thanos S and Paulus W:
Microglia promote glioma migration. Acta Neuropathol. 103:351–355.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Markovic DS, Glass R, Synowitz M, Rooijen
Nv and Kettenmann H: Microglia stimulate the invasiveness of glioma
cells by increasing the activity of metalloprotease-2. J
Neuropathol Exp Neurol. 64:754–762. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu A, Wei J, Kong LY, et al: Glioma cancer
stem cells induce immunosuppressive macrophages/microglia. Neuro
Oncol. 12:1113–1125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Straussman R, Morikawa T, Shee K, et al:
Tumour micro-environment elicits innate resistance to RAF
inhibitors through HGF secretion. Nature. 487:500–504. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Borovski T, Verhoeff JJC, ten Cate R, et
al: Tumor microvascular supports proliferation and expansion of
glioma-propagating cells. In J Cancer. 125:1222–1230.
2009.PubMed/NCBI
|
|
28
|
Gazdzinski LM and Nieman BJ: Cellular
imaging and texture analysis distinguish differences in cellular
dynamics in mouse brain tumors. Magn Reson Med. May 9–2013.
View Article : Google Scholar
|