Introduction
Certain types of cancer have a special proclivity
for perineural invasion (PNI), and these cancers are known as
‘neurotropic cancers’ (1). PNI is
defined as the presence of tumor cells in the perinerium space of
local peripheral nerves. Clinically, PNI has been recognized as a
distinct route of tumor cell dissemination and is associated with
the poor prognosis of these neurotropic cancers (1). Studies have found that PNI is more
common than previously suspected and is associated with tumor
recurrence and the poor prognosis of salivary adenoid cystic
carcinoma (SACC) (2,3), colon cancer (4), prostate cancer (5,6), and
pancreatic cancer (7,8). Although a series of molecules has been
found to play critical roles in the progression of PNI of
neurotropic cancers, the molecular mechanisms of PNI have not yet
been fully understood. Therefore, a better understanding of the
molecular mechanisms of PNI would provide new strategies for the
control of tumor dissemination in patients with neurotropic
cancers.
Chemokines coupled with their receptors on cancer
cells play important roles in tumor cell migration, invasion and
metastasis (9–11). CCL5 is an inflammatory chemokine
that is associated with the chemotactic activity of T cells,
monocytes, dendritic cells, eosinophils and basophils (12). Previous studies have found that CCL5
is also secreted by Schwann and dorsal root ganglia cells (13). CCR5, as the chemokine receptor for
CCL5, is predominantly expressed in tumor cells and is correlated
with invasion and metastasis of various types of tumors (14,15).
Recent studies have found that exogenous CCL5 may significantly
promote the migration and invasion of human breast cancer cells
in vitro (16–18). Further studies showed that the CCR5
antagonist may obviously block the invasion and metastasis of
breast cancer cells (19). These
results suggest that the CCL5/CCR5 axis is associated with the
invasion and dissemination of tumor cells. Interaction of tumor
cells with nerves is currently recognized to have a
fate-determining role in the progression and outcome of neurotropic
cancers (20). However, whether the
CCL5/CCR5 axis participates in tumor-nerve interaction and mediates
the PNI of neurotropic cancers has not been previously
reported.
SACC is a common neurotropic cancer and accounts for
~20% of all salivary gland malignancies and 1% of all head and neck
cancers (21). In the present
study, we investigated the expression of the CCR5/CCL5 axis in SACC
cases and its association with the PNI of SACC by
immunohistochemistry assay. We further investigated the effect of
the CCR5/CCL5 axis on the PNI activity of SACC cells by a series of
in vitro assays using CCL5 stimulation and/or CCR5 blockage.
Our results demonstrated for the first time that the CCL5/CCR5 axis
may participate in the tumor-nerve interaction and may function in
the progression of PNI of SACC.
Materials and methods
Tumor sample collection and
processing
The present study was approved by the Ethics
Committee of the Fourth Military Medical University, Xi’an, China.
Written informed consent was provided by all patients. Sixty-four
primary SACC samples and 30 normal human peripheral nerve tissues
were collected from the School of Stomatology of the Fourth
Military Medical University, Xi’an, China. All tissues were
immediately fixed with 4% paraformaldehyde in PBS, embedded in
paraffin and sectioned (4-μm) for use. All SACC samples were
pathologically diagnosed as SACC and evaluated by H&E staining
for conventional histological assessment.
Cell line and culture conditions
The human SACC cell line, SACC-83, was kindly
provided by Dr Shenglin Li (Peking University, China). SACC-83
cells were cultured in RPMI-1640 (HyClone, USA) with 10% fetal
bovine serum, 100 units/ml penicillin and 0.1 mg/ml streptomycin,
and were maintained in media at 37°C in a humid atmosphere of 5%
CO2/95% air.
Immunohistochemistry
To evaluate regional distribution and cellular
expression of CCR5 and CCL5 in the clinical tissues,
immunohistochemical staining was performed as previously described
(22). Primary antibodies,
polyclonal rabbit anti-human CCR5 (1:100) and polyclonal rabbit
anti-human CCL5 (both from PeproTech, USA; 1:100), were used.
Negative controls were performed by omitting the primary
antibodies.
All immunostained sections were evaluated in a
blinded manner by two pathologists (Yuan Liu and Jun Zhou). Results
of the staining for CCR5 and CCL5 were classified into the
following 3 categories: negative staining (−), <25% of cells
stained; moderate staining (+), 25–50% of cells stained; and strong
staining (++), >50% of cells stained.
Immunofluorocytochemistry
Cells were cultured on 12-mm coverslips for 24 h at
37°C in a humid atmosphere of 5% CO2/95% air. Cells were
then fixed with 4% paraformaldehyde for 15 min and permeabilized in
0.1% Triton X-100 for 10 min. The cells were subsequently incubated
with polyclonal rabbit anti-human CCR5 (1:100) or polyclonal rabbit
anti-human CCL5 (1:100) overnight at 4°C, and then incubated in
goat anti-rabbit secondary antibody (PeproTech; 1:200) for 1 h at
room temperature. After washing with PBS for 3 times, the cells
were examined using a fluorescence microscope (Leica, Germany).
Analysis of CCR5 expression by flow
cytometry
Harvested cells (5×106) were resuspended
with 1% FCS-PBS and were incubated with anti-CCR5-FITC (1:100) or
anti-CCL5-FITC (both from PeproTech; 1:100) in the dark at 37°C for
1 h. Cells were then washed, centrifuged and resuspended in 500 μl
PBS. The fluorescence intensity was analyzed by a flow cytometer
(BD Biosciences, USA).
Analysis of Ca2+ mobilization
by flow cytometry
Ca2+ mobilization was performed with the
fluorescent Ca2+ indicator Fluo-3 as reported (23). Harvested cells (1×107)
were loaded with Fluo-3AM (5 μM; Sigma, USA) in the dark at 37°C
for 45 min, and then washed 2 times with 1% FCS-PBS. Then, cells
were diluted to 5×106 cells/ml and kept in the dark at
37°C for 30 min before the beginning of the Ca2+
mobilization measurements. After adding 100 ng/ml CCL5, the
fluorescence intensity of the cells was recorded by a flow
cytometer. Maximal Ca2+ release was measured by
lonomycin (Sigma) as previously reported (23). In the other group, cells were
pretreated with 200 μg/ml maraviroc for 30 min at 37°C before CCL5
treatment.
Confocal microscopy for the actin
polymerization assay
Cells were cultured on 12-mm coverslips for 6 h, and
incubated with 100 ng/ml CCL5 for 30 min at 37°C. In the other
group, cells were pretreated with 200 μg/ml maraviroc for 30 min at
37°C before CCL5 treatment. Cells were then fixed with 4%
paraformaldehyde for 15 min and incubated with rhodamine-labeled
phalloidin (Sigma) at 37°C for 30 min. After washing with PBS for 3
times, cells were observed by confocal microscopy (Olympus
Corporation, Japan). Negative controls were performed by incubating
cells in serum-free RPMI-1640 without CCL5.
In vitro migration and invasion
assays
Migration and invasion assays were performed using
12-mm-diameter inserts with a 8-μm pore size in 24-well dishes
(Corning, USA). For the migration assays, 1×105 cells in
200 μl of serum-free RPMI-1640 were placed in the upper chamber.
Serum-free (600 μl) RPMI-1640 containing various concentrations of
CCL5 were placed in the lower chamber. Cells were incubated at 37°C
for 12 h. Then, cells on the upper surface of the filters were
scraped with a cotton swab. Cells that had migrated and reached the
lower surface of the filters were fixed in methanol, stained with
H&E and counted as previously described (24). In another set of migration assays,
cells were pretreated for 30 min with different concentrations of
maraviroc (Sigma). For invasion assays, the Transwell inserts were
covered with Matrigel (BD Biosciences; 100 μg/cm2) and
the cells were incubated on the inserts for 24 h. The other steps
were identical to those of the migration assays.
In vitro PNI assays
To study the effects of the CCR5/CCL5 axis on the
PNI activity of SACC cells, we modified the in vitro PNI
model established in our previous study (25). Briefly, the Transwell inserts were
covered with Matrigel (100 μg/cm2). Cells
(1×105) in 200 μl of serum-free RPMI-1640 were placed in
the upper chamber. Then 600 μl of conditioned medium (incubating
1×106 neural cells in 600 μl serum-free RPMI-1640 medium
for 24 h) was placed in the lower chamber to simulate the
perineural surrounding environment. Cells were incubated at 37°C
for 24 h. Then, cells on the upper surface of the filters were
removed. Cells that had invaded the Matrigel and reached the lower
surface of the filters were fixed, stained and counted as
previously described (25).
Statistical analysis
Statistical analysis was performed by SPSS 17.0
software package (USA). The differences in expression of CCR5 and
CCL5 were compared by the Fisher’s exact test. Correlations were
evaluated by the Spearman’s rank correlation coefficient test. All
in vitro assays were performed in triplicate, and the data
are presented as means ± SD. Results were analyzed with the
Student’s t-test and the one-way ANOVA tests. P-values <0.05
were considered to indicate statistically significant results in
all tests.
Results
Expression of the CCR5/CCL5 axis in
SACC
To determine the distribution of CCR5 and CCL5 in
SACC and nerve tissues, we performed immunohistochemical analysis.
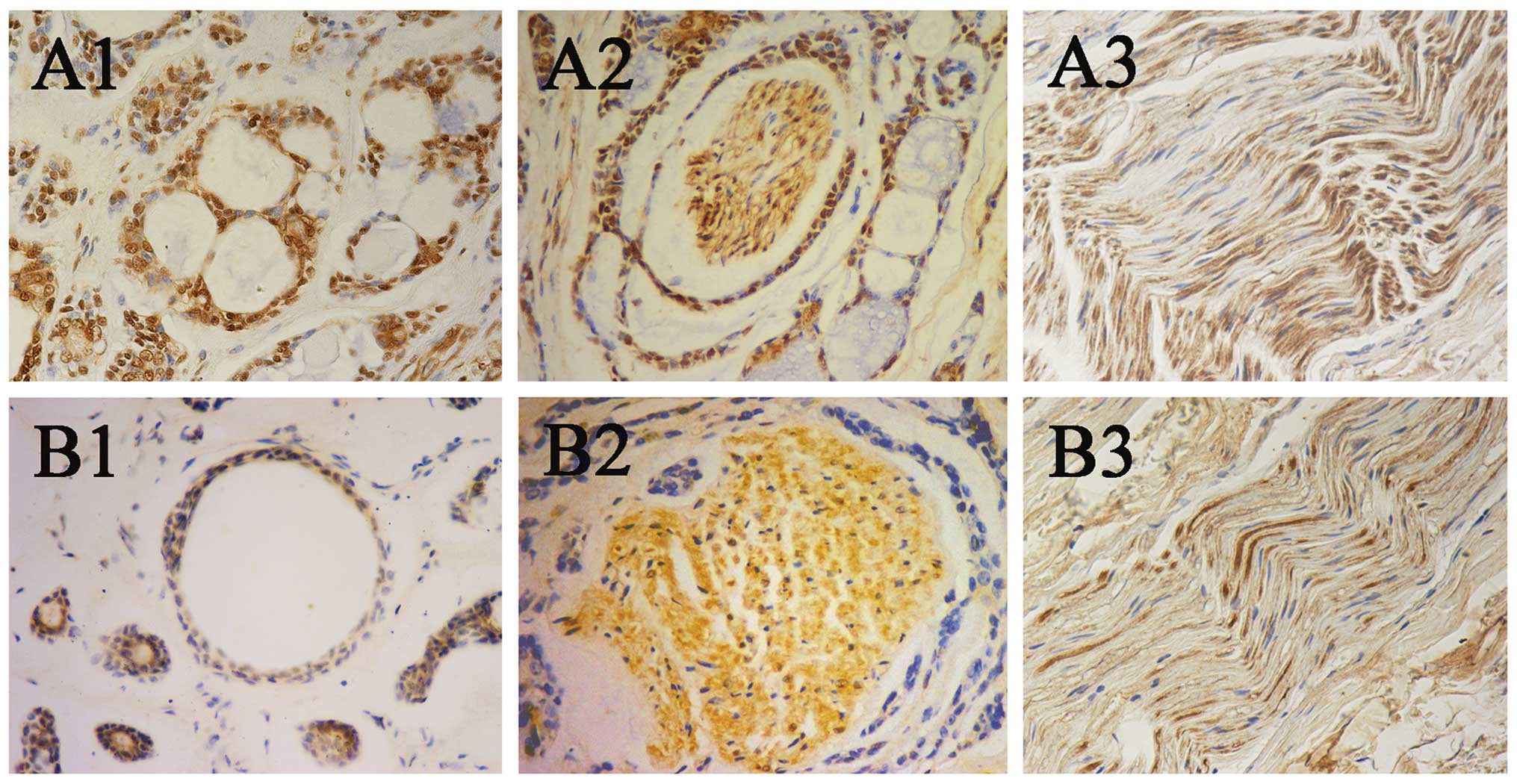
The results showed that CCR5 was highly expressed in the cytoplasm
and nuclei of the tumor cells and the neural cells (Fig. 1A1–A3). CCL5 was weakly expressed in
the cytoplasm and the nuclei of the tumor cells, and was highly
expressed in the cytoplasm and the nuclei of the neural cells
(Fig. 1B1–B3). Notably, the
expression of CCL5 in the nerve tissues with PNI was stronger than
that in the normal nerve tissues (Fig.
1B2 and B3). Of the 64 SACC samples, CCR5 was detected in 45
cases (70.3%), and CCL5 was observed in 23 cases (35.9%). The
statistical analysis showed that the expression of CCR5 was
significantly corrrelated the PNI of SACC (P<0.05; Table I), while the expression of CCL5 was
negatively associated with the PNI of SACC (P>0.05; Table I).
 | Table ICorrelation between CCR5 and CCL5
expression and the perineural invasion of SACC. |
Table I
Correlation between CCR5 and CCL5
expression and the perineural invasion of SACC.
| | CCR5 | | CCL5 | |
|---|
| |
| |
| |
|---|
| Variables | n | − | + | ++ | P-value | − | + | ++ | P-value |
|---|
| Perineural
invasion | | | | | 0.027a | | | | 0.587 |
| Positive | 34 | 6 | 10 | 18 | | 20 | 8 | 6 | |
| Negative | 30 | 13 | 10 | 7 | | 21 | 6 | 3 | |
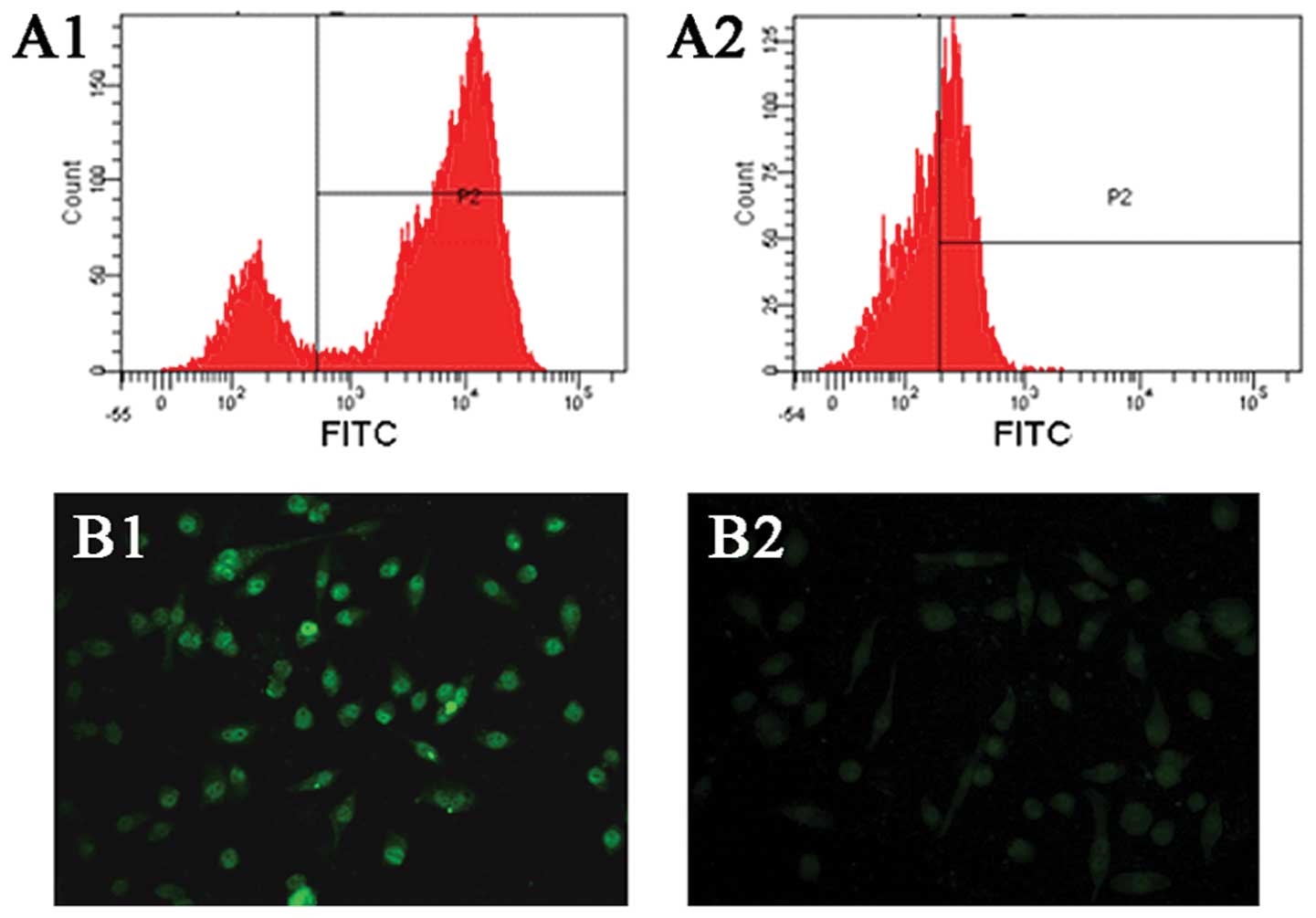
To examine the expression of CCR5 and CCL5 in
SACC-83 cells, flow cytometric analysis and immunofluorescence
analysis were performed. As shown in Fig. 2A1 and A2, flow cytometric analysis
showed that SACC-83 cells highly expressed CCR5 and weakly
expressed CCL5. In addition, immunofluorescence analysis also
demonstrated the stronger expression of CCR5 and weaker expression
of CCL5 in SACC-83 cells (Fig. 2B1 and
B2).
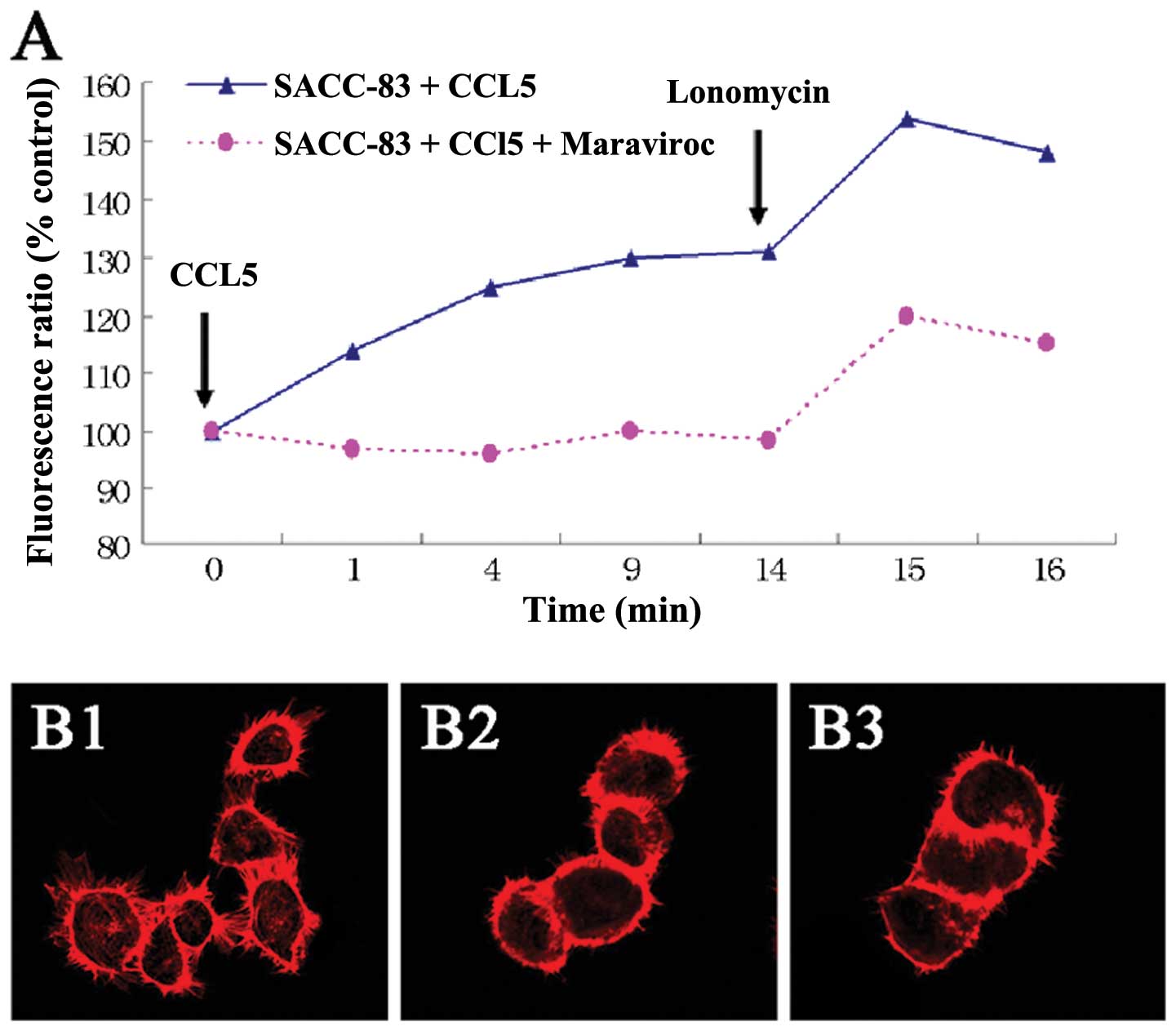
Effect of CCL5/CCR5 on calcium influx and
F-actin polymerization of SACC-83 cells
To verify that the CCL5/CCR5 axis was functional,
the cytoplasmic free Ca2+ concentration was measured by
flow cytometric analysis. Fig. 3A
shows a rapid and transient Ca2+ elevation in SACC-83
cells preloaded with Fluo-3AM after the addition of CCL5. However,
Ca2+ fluctuation in SACC-83 cells pretreated with 200
ng/ml maraviroc (CCR5 inhibitor) was not obviously changed after
the addition of CCL5.
To investigate whether F-actin polymerization is
induced by the CCL5/CCR5 axis in SACC-83 cells, we observed changes
in F-actin in SACC-83 cells by confocal microscopy. As a result,
after treatment with CCL5 at 100 ng/ml for 30 min, SACC-83 cells
showed intense F-actin staining in the periphery of the cells and
distinct pseudopodium formation (Fig.
3B1). However, the cells in the control group or the cells
pre-blocked for CCR5 by 200 ng/ml maraviroc showed no marked
F-actin redistribution and pseudopodium formation (Fig. 3B2 and B3).
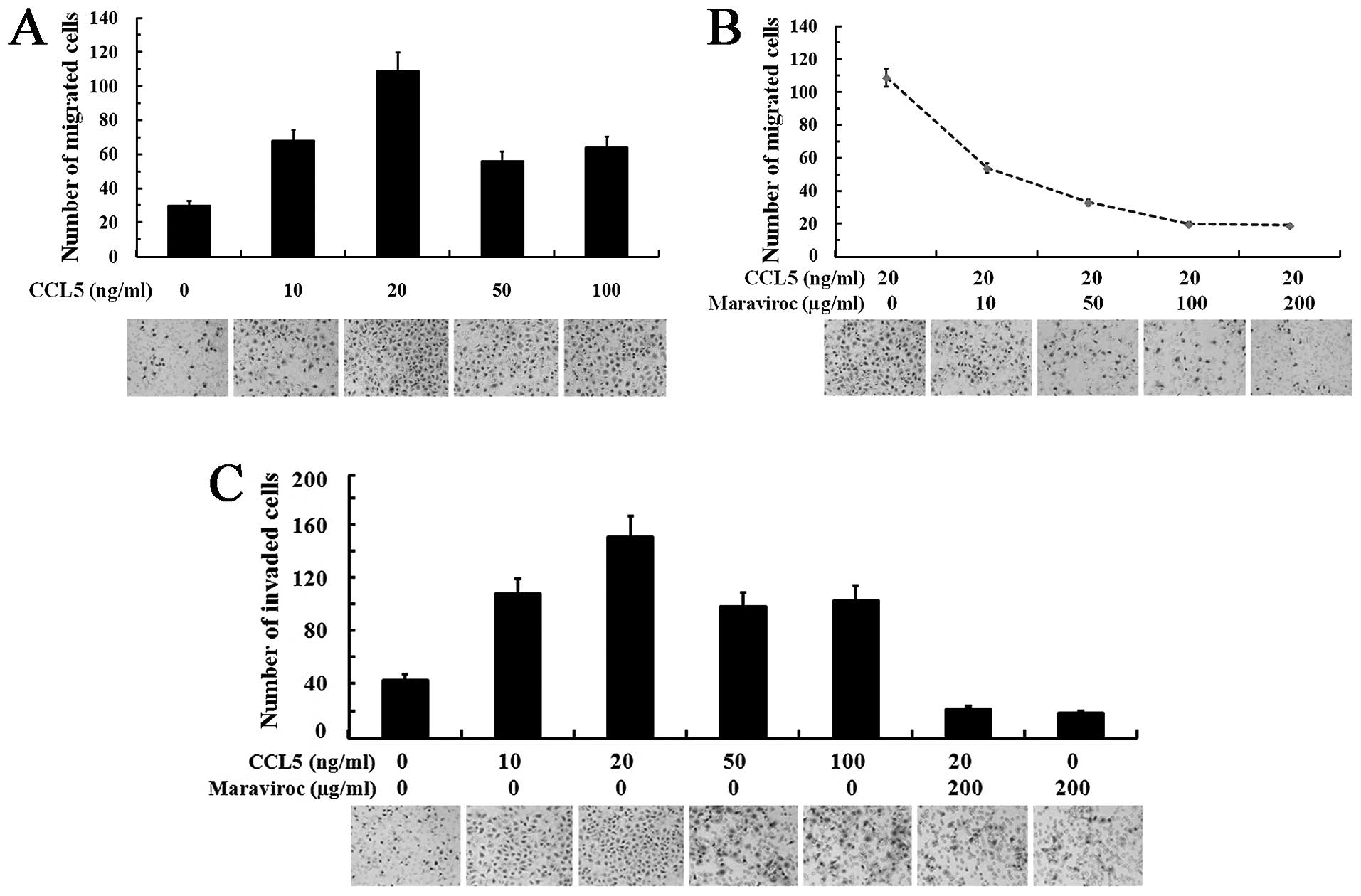
Effects of the CCR5/CCL5 axis on the
migration and invasion of SACC-83 cells
To assess the effects of the CCL5/CCR5 axis on the
migration and invasion activities of SACC-83 cells, we performed
Transwell assays under the condition of CCL5 stimulation and/or
CCR5 blockage. Fig. 4A shows that
various concentrations of CCL5 promoted the migration of SACC-83
cells. A Maximal promotive effect of CCL5 on the migration of
SACC-83 cells was observed at a concentration of 20 ng/ml
(P<0.05). In addition, the migration of SACC-83 cells induced by
CCL5 at 20 ng/ml was maximally inhibited by maraviroc at
concentrations from 100 to 200 μg/ml (P<0.05; Fig. 4B).
The effects of CCL5 and the CCR5 inhibitor on the
invasive activity of SACC-83 cells are shown in Fig. 4C. CCL5 (20 ng/ml) obviously promoted
the invasion of SACC-83 cells (P<0.05), while 200 μg/ml
maraviroc significantly inhibited the CCL5-induced invasiveness of
SACC-83 cells (P<0.05). In addition, SACC-83 cells pretreated by
maraviroc showed no significantly changes in invasion activity when
stimulated with 20 ng/ml CCL5 or without (P>0.05).
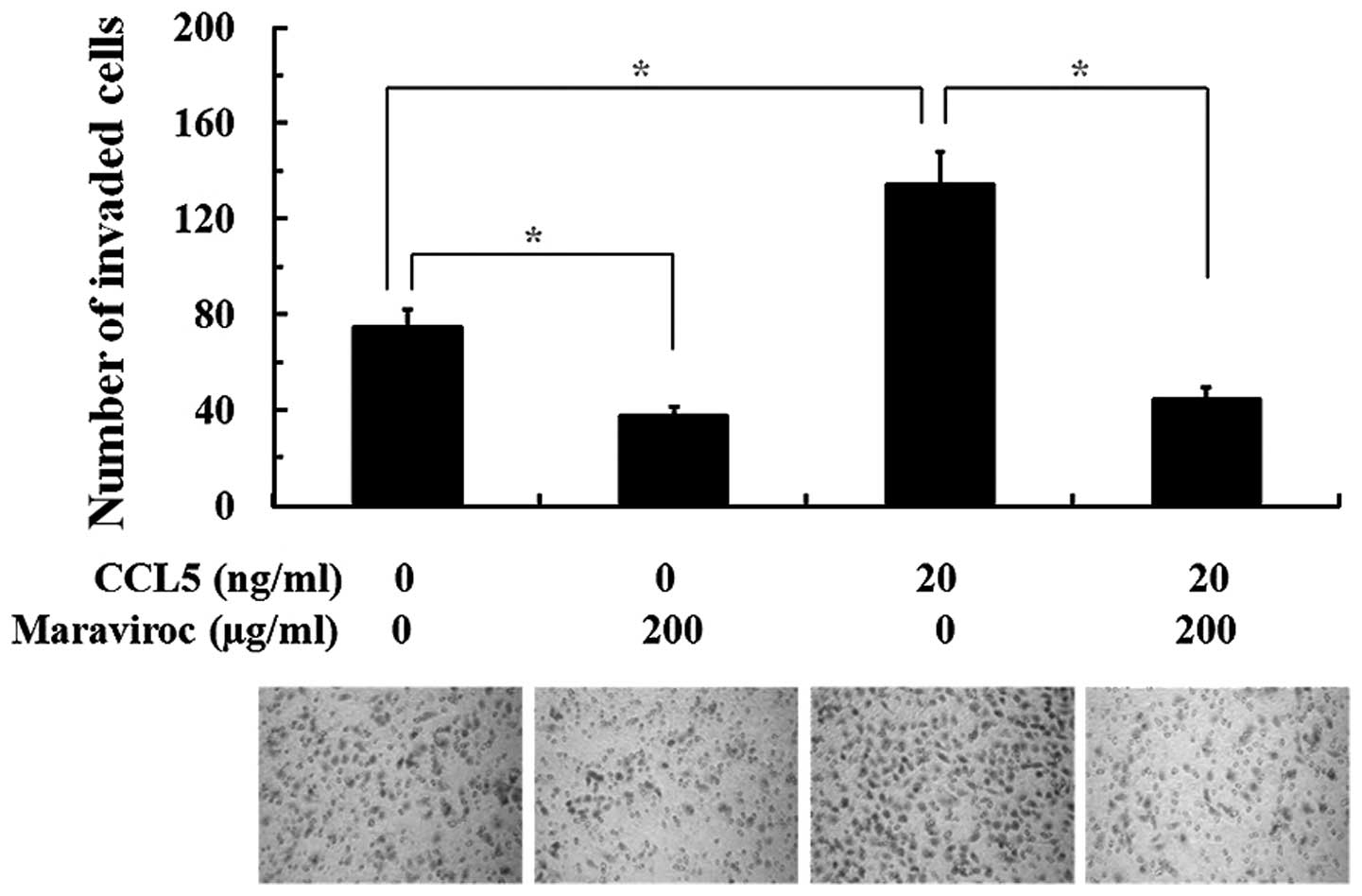
Effects of the CCL5/CCR5 axis on the PNI
activity of SACC-83 cells
The effects of the CCR5/CCL5 axis on the PNI
activity of SACC cells were studied by modified in vitro PNI
models. As shown in Fig. 5, after
24 h of incubation, abundant SACC-83 cells were observed on the
lower surface of the filter when the lower chamber was supplemented
with neural cell conditioned medium to simulate the perineural
surrounding environment. The PNI activity of SACC-83 cells was
significantly increased under the condition of 20 ng/ml CCL5 when
compared with the control group (P<0.05). Compared with the
control group, the PNI activity of SACC-83 cells both with or
without CCL5 stimulation was obviously inhibited when cells were
pretreated with 200 μg/ml maraviroc (P<0.05).
Discussion
Salivary adenoid cystic carcinoma (SACC) has a
special proclivity for perineural invasion (PNI), which leads to
tumor recurrence and poor prognosis (2,3,21).
Elucidation of the molecules involved in the progression of PNI
would provide new insight for the understanding of the mechanisms
of PNI and would be beneficial for the control of SACC with PNI.
Recent studies have shown that the chemokine CCL5 and its receptor
CCR5 play important roles in tumor invasion and metastasis
(14–19). However, the role of the CCL5/CCR5
axis in the PNI of SACC has not been studied to date. In the
present study, we found that SACC tissues and cells highly
expressed CCR5, while the nerves invaded by SACC expressed elevated
levels of CCL5. Immunohistochemical analysis showed that high
expression of CCR5 was significantly associated with the PNI of
SACC. In the in vitro assays, we found that exogenous CCL5
significantly promoted the migration, invasion and PNI activity of
SACC-83 cells, while the CCR5 inhibitor effectively inhibited the
migration, invasion and PNI activity of SACC-83 cells both with or
without CCL5 stimulation. These findings suggest that the CCL5/CCR5
axis may contribute to the progression of PNI of SACC.
To verify whether the interaction of CCR5 and its
ligand was functional, we detected the cytoplasmic free
Ca2+ concentration and the reorganization of actin
cytoskeleton after stimulation with CCL5. Numerous studies have
shown that a rapid and transient Ca2+ elevation was
induced by chemokine in a series of tumors and was significantly
correlated with the mobility of tumor cells (19,23).
Our results showed an obvious Ca2+ elevation after
stimulation by CCL5 in SACC-83 cells. We also found that SACC-83
cells pretreated with the CCR5 inhibitor showed no marked
intracellular calcium flow both with or without CCL5 stimulation.
In tumor cells, high levels of actin polymerization are needed for
the formation of pseudopodia, which is the foundation and early
event of efficient migration and invasion of tumor cells (26,27).
Invading cells require an alteration in the cell-cell adhesion
properties and reorganize their cytoskeletons to facilitate cell
motility (26,27). In the present study, we observed
that, following treatment of SACC-83 cells with CCL5, marked actin
polymerization and pseudopodium formation were noted in the
periphery of cells. These results indicate that the CCR5 expressed
in SACC cells was the functional receptor that responds to CCL5.
Take together, the motility of SACC cells may be associated with
the cytoplasmic free Ca2+ concentration and the
reorganization of actin cytoskeleton induced by CCL5/CCR5 axis
activation.
During the process of invasion, invading cells
require a change in cell-cell adhesion properties, reorganization
of their cytoskeleton and reconstitution of the extracellular
microenvironment (28). To confirm
the functionality of the CCL5/CCR5 axis in SACC cell migratory and
invasive activity, we evaluated the migration and invasion
capabilities of SACC cells by adding CCL5 or by blocking CCR5 in
vitro. We found that exogenous CCL5 facilitated the migration
and invasion of SACC cells. We also found that neutralizing CCR5 by
the CCR5 inhibitor efficiently impaired the migration and invasion
of SACC cells. In addition, we found that the CCL5/CCR5 axis was
associated with the PNI of SACC cells in vitro. The PNI
ability of SACC cells was improved after adding exogenous CCL5, and
was obviously suppressed by the CCR5 inhibitor. These results
indicate that the CCL5/CCR5 axis plays a critical role in the
migration, invasion and PNI activity of SACC cells.
Traditionally, studies of PNI have focused on the
neurotropism of tumor cells. Whereas recent findings underline the
interaction of tropism between tumor and nerve cells, nerve tissues
provide a prosperous microenvironment for tumor invasion and this
interaction has beneficial effects on the growth and motility of
both tumor and nerve cells (1,20). One
study previously found that obvious reciprocity between neurite
outgrowth from mouse dorsal root ganglia (DRG) and pancreatic
cancer (PCa) cell invasive growth was observed when human PCa cells
were co-cultured with mouse DRG (29). Swanson et al also discovered
tumor-nerve interactions by means of the transmembrane mucin MUC1
expressed by tumor cells binding to myelin-associated glycoprotein
expressed on Schwann cells of peripheral nerves (30). In the present study, we observed the
role of the CCL5/CCR5 axis in the PNI of SACC cells. We found that
CCR5 was highly expressed in SACC cells, while CCL5 was obviously
observed in nerves invaded by SACC. Moreover, exogenous CCL5
functionally activated the CCR5 and thus promoted the motility and
PNI activity of SACC cells in vitro. The CCR5 inhibitor
significantly inhibited the motility and PNI activity of SACC
cells. All of these results suggest that the activation of the
CCR5/CCL5 axis may be a potent mechanism for the PNI of SACC.
However, whether the CCL5/CCR5 axis is involved in the reciprocal
growth interaction between nerves and SACC in the progression of
PNI needs to be further explored.
The tumor-nerve interaction is an important research
topic, and not only their abundant resounding potential, but also
the major therapeutic potential deserves considerable attention.
Maraviroc is a new CCR5 antagonist (31). It has received full US Food and Drug
Administration (FDA) approval for use in treatment-naive adults
with CCR5-trophic HIV (32). Recent
studies have shown that maraviroc inhibits breast cancer cell
invasion in vitro and effectively reduces migration of
breast cancer cells to target organs in vivo (19). Therefore, maraviroc is considered to
be an antineoplastic agent for breast tumors and other tumor types
that highly express CCR5 (9,19). Our
study found that exogenous CCL5 promoted the in vitro PNI
activity of SACC cells that highly express CCR5. Our study further
found that maraviroc effectively inhibited the PNI activity of SACC
cells. These results suggest that maraviroc may interrupt the
tumor-nerve interaction through blocking activation of the
CCL5/CCR5 axis and thus may inhibit the PNI progression of SACC
cells.
In conclusion, this is the first study to show that
the CCL5/CCR5 axis acts as a crucial mediator of the PNI of SACC.
Moreover, the CCR5 inhibitor maraviroc may effectively suppress the
PNI activity of SACC cells in vitro. Our study suggests that
small-molecule antagonists of CCR5 may be a potent anti-PNI agent
for SACC.
Acknowledgements
We thank Professor Zhou Jun and Dr Liu Yuan for
their excellent technical assistance. This work was supported by
the National Natural Science Foundation of China for Dr Moyi Sun
(grant nos. 81072230, 30772428) and Dr Xinjie Yang (grant no.
81302352).
References
|
1
|
Liebig C, Ayala G, Wilks JA, Berger DH and
Albo D: Perineural invasion in cancer: a review of the literature.
Cancer. 115:3379–3391. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barrett AW and Speight PM: Perineural
invasion in adenoid cystic carcinoma of the salivary glands: a
valid prognostic indicator? Oral Oncol. 45:936–940. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnston M, Yu E and Kim J: Perineural
invasion and spread in head and neck cancer. Expert Rev Anticancer
Ther. 12:359–371. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Poeschl EM, Pollheimer MJ, Kornprat P, et
al: Perineural invasion: correlation with aggressive phenotype and
independent prognostic variable in both colon and rectum cancer. J
Clin Oncol. 28:e358–e360. 2010. View Article : Google Scholar
|
|
5
|
Fromont G, Godet J, Pires C, Yacoub M,
Dore B and Irani J: Biological significance of perineural invasion
(PNI) in prostate cancer. Prostate. 72:542–548. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sakurai J, Fujii Y and Shirotani M:
Contraction induced by Clostridium perfringens alpha toxin
in the isolated rat ileum. Toxicon. 28:411–418. 1990.
|
|
7
|
Cavel O, Shomron O, Shabtay A, et al:
Endoneurial macrophages induce perineural invasion of pancreatic
cancer cells by secretion of GDNF and activation of RET tyrosine
kinase receptor. Cancer Res. 72:5733–5743. 2012. View Article : Google Scholar
|
|
8
|
Bapat AA, Hostetter G, von Hoff DD and Han
H: Perineural invasion and associated pain in pancreatic cancer.
Nat Rev Cancer. 11:695–707. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang SW, Wu HH, Liu SC, et al: CCL5 and
CCR5 interaction promotes cell motility in human osteosarcoma. PLoS
One. 7:e351012012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
McIver SC, Loveland KL, Roman SD, Nixon B,
Kitazawa R and McLaughlin EA: The chemokine CXCL12 and its receptor
CXCR4 are implicated in human seminoma metastasis. Andrology.
1:517–529. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu Q, Han X, Peng J, Qin H and Wang Y:
The role of CXC chemokines and their receptors in the progression
and treatment of tumors. J Mol Histol. 43:699–713. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lv D, Zhang Y, Kim HJ, Zhang L and Ma X:
CCL5 as a potential immunotherapeutic target in triple-negative
breast cancer. Cell Mol Immunol. 10:303–310. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bolin LM, Murray R, Lukacs NW, et al:
Primary sensory neurons migrate in response to the chemokine
RANTES. J Neuroimmunol. 81:49–57. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vaday GG, Peehl DM, Kadam PA and Lawrence
DM: Expression of CCL5 (RANTES) and CCR5 in prostate cancer.
Prostate. 66:124–134. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Borczuk AC, Papanikolaou N, Toonkel RL, et
al: Lung adenocarcinoma invasion in TGFβRII-deficient cells
is mediated by CCL5/RANTES. Oncogene. 27:557–564. 2008.PubMed/NCBI
|
|
16
|
Zhang Y, Yao F, Yao X, et al: Role of CCL5
in invasion, proliferation and proportion of
CD44+/CD24− phenotype of MCF-7 cells and
correlation of CCL5 and CCR5 expression with breast cancer
progression. Oncol Rep. 21:1113–1121. 2009.PubMed/NCBI
|
|
17
|
Swamydas M, Ricci K, Rego SL and Dréau D:
Mesenchymal stem cell-derived CCL-9 and CCL-5 promote mammary tumor
cell invasion and the activation of matrix metalloproteinases. Cell
Adh Migr. 7:315–324. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pinilla S, Alt E, Abdul Khalek FJ, et al:
Tissue resident stem cells produce CCL5 under the influence of
cancer cells and thereby promote breast cancer cell invasion.
Cancer Lett. 284:80–85. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Velasco-Velázquez M, Jiao X, De La Fuente
M, et al: CCR5 antagonist blocks metastasis of basal breast cancer
cells. Cancer Res. 72:3839–3850. 2012.PubMed/NCBI
|
|
20
|
Demir IE, Friess H and Ceyhan GO:
Nerve-cancer interactions in the stromal biology of pancreatic
cancer. Front Physiol. 3:972012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li LJ, Li Y, Wen YM, Liu H and Zhao HW:
Clinical analysis of salivary gland tumor cases in West China in
past 50 years. Oral Oncol. 44:187–192. 2008.PubMed/NCBI
|
|
22
|
Yang X, Dai J, Li T, et al: Expression of
EMMPRIN in adenoid cystic carcinoma of salivary glands: correlation
with tumor progression and patients’ prognosis. Oral Oncol.
46:755–760. 2010.PubMed/NCBI
|
|
23
|
Sukhanova KY, Harhun MI, Bouryi VA and
Gordienko DV: Mechanisms of [Ca2+]i elevation following P2X
receptor activation in the guinea-pig small mesenteric artery
myocytes. Pharmacol Rep. 65:152–163. 2013.
|
|
24
|
Yang X, Zhang P, Ma Q, et al: EMMPRIN
contributes to the in vitro invasion of human salivary
adenoid cystic carcinoma cells. Oncol Rep. 27:1123–1127. 2012.
|
|
25
|
Yang X, Zhang P, Ma Q, et al: EMMPRIN
silencing inhibits proliferation and perineural invasion of human
salivary adenoid cystic carcinoma cells in vitro and in vivo.
Cancer Biol Ther. 13:85–91. 2012. View Article : Google Scholar
|
|
26
|
Bravo-Cordero JJ, Hodgson L and Condeelis
J: Directed cell invasion and migration during metastasis. Curr
Opin Cell Biol. 24:277–283. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Olson MF and Sahai E: The actin
cytoskeleton in cancer cell motility. Clin Exp Metastasis.
26:273–287. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Detchokul S, Williams ED, Parker MW and
Frauman AG: Tetraspanins as regulators of the tumour
microenvironment: implications for metastasis and therapeutic
strategies. Br J Pharmacol. Jun 3–2013.(Epub ahead of print).
View Article : Google Scholar
|
|
29
|
Dai H, Li R, Wheeler T, Ozen M, et al:
Enhanced survival in perineural invasion of pancreatic cancer: an
in vitro approach. Hum Pathol. 38:299–307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Swanson BJ, McDermott KM, Singh PK, Eggers
JP, Crocker PR and Hollingsworth MA: MUC1 is a counter-receptor for
myelin-associated glycoprotein (Siglec-4a) and their interaction
contributes to adhesion in pancreatic cancer perineural invasion.
Cancer Res. 67:10222–10229. 2007. View Article : Google Scholar
|
|
31
|
Sayana S and Khanlou H: Maraviroc: a new
CCR5 antagonist. Expert Rev Anti Infect Ther. 7:9–19. 2009.
View Article : Google Scholar
|
|
32
|
Kast RE: Glioblastoma: synergy of growth
promotion between CCL5 and NK-1R can be thwarted by blocking CCL5
with miraviroc, an FDA approved anti-HIV drug and blocking NK-1R
with aprepitant, an FDA approved anti-nausea drug. J Clin Pharm
Ther. 35:657–663. 2010. View Article : Google Scholar : PubMed/NCBI
|