Introduction
Osteosarcoma (OS) is the most common primary
malignant tumor arising from bone in children and young adults
(1). Despite aggressive
chemotherapeutic treatment strategies, the survival of these
patients has limited improvement. The prognosis is very poor, and
the 5-year mortality rate for OS patients remains at 40% (2).
The canonical Wnt signaling pathway β-catenin is
considered the key component of this pathway, which plays important
roles during embryonic development and oncogenesis (3,4). It
was reported that β-catenin has a key role in the Wnt signaling
pathway, which is important in cell growth, mobility,
differentiation and tumor formation (5). Previous studies have demonstrated that
cytoplasmic and nuclear β-catenin protein is frequently elevated in
human OS (6), and a high level of
intracytoplasmic and/or nuclear expression of β-catenin in OS cells
was associated with pulmonary metastasis (7). In the lack of Wnt ligands, cytoplasmic
β-catenin protein is phosphorylated through a complex containing
GSK3β/APC/Axin followed by degradation. In tumor cells, β-catenin
may aberrantly accumulate in the cytosol and further translocates
into the nucleus where it interacts with T-cell factor/lymphoid
enhancing factor (TCF/LEF) that regulate expression of several
downstream target genes including c-myc, cyclin D1, MMP7 (encoding
matrix metalloproteinase 7) and osteoprotegerin (OPG) (8–10),
which play important roles in tumorigenesis and metastasis.
Bone morphogenetic proteins (BMPs) belong to the
transforming growth factor-β (TGF-β) superfamily. BMPs regulate a
wide range of physiologic events, including cell proliferation,
differentiation, apoptosis, migration and invasion (11,12).
BMP9 is probably the most osteogenic differentiation potent
inducer; it was reported that autocrine BMP9 promotes ovarian
cancer and human hepatocellular carcinoma cell proliferation, and
BMP9 overexpression prevents prostate and breast cancer cell growth
(13–16). Although the role of BMP9 in several
types of cancer has been documented, its roles in OS and molecular
mechanisms have yet to be well investigated. Thus, in the present
study, we investigated the role of BMP9 on the growth of the human
OS 143B and MG63 cells following both overexpression and knockout
of BMP9. To increase our understanding of the molecular mechanisms
underlying BMP9 in the development of OS, we investigated the
underlying involvement of β-catenin signaling in human OS.
Materials and methods
Materials
Dulbecco's modified Eagle's medium-high glucose
(DMEM-HG) and fetal bovine serum (FBS) were purchased from HyClone.
The human embryonic kidney cell line HEK293 and the human OS cell
lines 143B and MG63 were obtained from the American Type Culture
Collection (ATCC, Manassas, VA, USA). Recombinant adenovirus
expressing green fluorescent protein (AdGFP), red fluorescent
protein (AdRFP), BMP9 (AdBMP9), β-catenin (Adβ-catenin) were
donated by Professor T.C. He, Chicago University, USA. Recombinant
adenoviruses expressing siRNA targeted β-catenin (Adsiβ-catenin),
BMP9 (AdsiBMP9) with RFP were generated in our laboratory. TRIzol
reagent was purchased from Invitrogen. RT-PCR reagents were
purchased from Takara Biotech (Liaoning, China). Anti-GSK-3β,
anti-p-GSK-3β, anti-β-catenin, anti-β-actin, anti-C-myc, anti-OPG
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). Secondary antibody reagents were obtained
from Zhongshan Golden Bridge, Beijing, China. Western blot
detection reagents were purchased from Beyotime Institute of
Biotechnology (Jiangsu, China). BeyoECL was purchased from
Millipore.
Cell culture, preparation of BMP9
conditioned medium and adenovirus infection
The HEK293 and human OS cell lines 143B and MG63
were maintained in DMEM-HG; HCT116 cells were maintained in MEM-HG
supplemented with 10% FBS and 100 U/ml of penicillin G/streptomycin
at 37°C in 5% CO2. HEK293 cells were used for adenovirus
amplification. Log-phase HCT116 cells were infected with AdBMP9.
After infection for 4–6 h, the culture medium was changed to
serum-free DMEM. After further culture for 24 h, supernatants were
collected and used immediately. OS cells were treated with AdBMP9,
AdsiBMP9, Adβ-catenin, Adsiβ-catenin or the control adenovirus
AdGFP, AdRFP. After infecting for 8–12 h, the medium was replaced
with serum-free or low serum DMEM, Adβ-catenin, Adsiβ-catenin
infected cells replaced with BMP9 conditional medium.
Cell proliferation assay
OS cells were seeded in 96-well culture plates. On
the following day, the cells were treated with AdBMP9, AdsiBMP9 or
Adβ-catenin, Adsiβ-catenin with BMP9 conditioned medium. At the
indicated time, the supernatant was removed and 20 μl of the MTT
reagent (5 mg/ml) was added. The mixture was then incubated for 4
h, the MTT solution was removed and formazan was dissolved in DMSO,
and then absorbance was measured at 490 nm using a Sunrise remote
microplate reader.
Cell cycle and apoptosis analysis
Log-phase cells from each group were harvested by
centrifugation. After washing twice with ice-cold PBS and
resuspending, cell cycle distribution and apoptosis were analyzed
by a FACSVantage SE flow cytometer (Becton-Dickinson, USA). Each
experiment was performed thrice.
RNA extraction and RT-PCR analysis
Total RNA was extracted from OS cells using TRIzol
reagent, according to the manufacturer's instructions. First-strand
DNA was synthesized using the Reverse Transcriptase M-MLV (RNase
H−) kit with random hexamer primers. PCR products were
separated by electrophoresis on a 2% agarose gels. The primers
were: β-catenin (forward, 5′-CTGCAGGGGTCCTCTGTG-3′ and reverse,
5′-TGCATATGTCGCCACACC-3′; 125 bp), C-myc (forward,
5′-TACCCTCTCAACGACAGCAG-3′ and reverse, 5′-TCTTGACATTCTCCTCGGTG-3′;
478 bp), OPG (forward, 5′-AGTGGGAGCAGAAGACAT-3′ and reverse,
5′-TGGA CCTGGTTACCTATC-3′; 264 bp), GAPDH (forward, 5′-CAG
CGACACCCACTCCTC-3′ and reverse, 5′-TGAGGTCCA CCACCCTGT-3′; 122
bp).
Western blot analysis
Briefly, OS cells were lysed with RIPA buffer, then
centrifuged at 13,000 × g for 5 min at 4°C and supernatants were
collected. The protein concentration was determined by the BCA
assay. Proteins were denatured, separated on a 10% SDS-PAGE and
transferred to a PVDF membrane, after blocking with BSA for 2 h,
probed with primary antibody incubation, followed by a secondary
IgG antibody incubation for 1 h. The proteins were detected using
SuperSignal West Pico Chemiluminescent Substrate kit.
Statistical analysis
Data are expressed as the means ± SD. Statistical
analysis was performed using GraphPad Prism 5. p<0.05 was
considered to indicate a statistically significant difference.
Results
In vitro cell viability depends on BMP9
expression
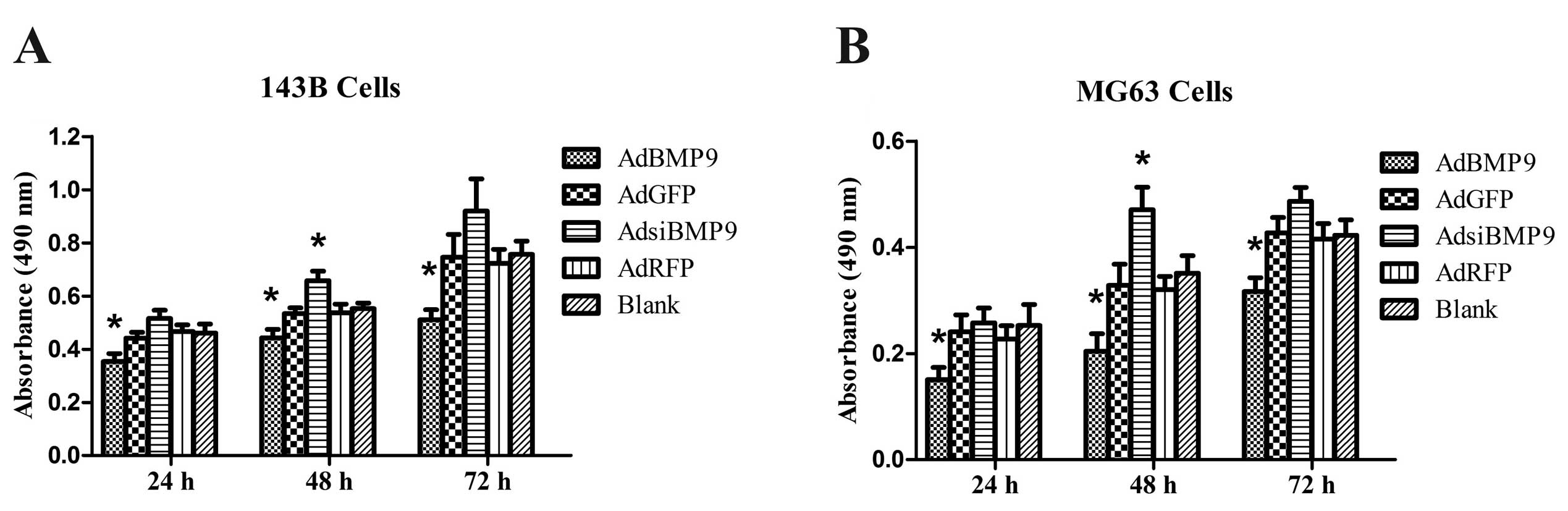
To investigate the effects of BMP9 on cell growth,
OS cells were treated with AdBMP9 and AdsiBMP9 for 24, 48 and 72 h
and cell viability was examined by MTT assay. As shown in Fig. 1, the cell viability of OS cells
treated with AdBMP9 was significantly inhibited at 24 h and tended
to be more significant at 48 and 72 h, whereas the cell viability
of OS cells treated with AdsiBMP9 was significantly promoted only
at 48 h. These results suggest that BMP9 overexpression inhibits
the growth of OS cells.
BMP9 overexpression induces G1 cell cycle
arrest in OS cells
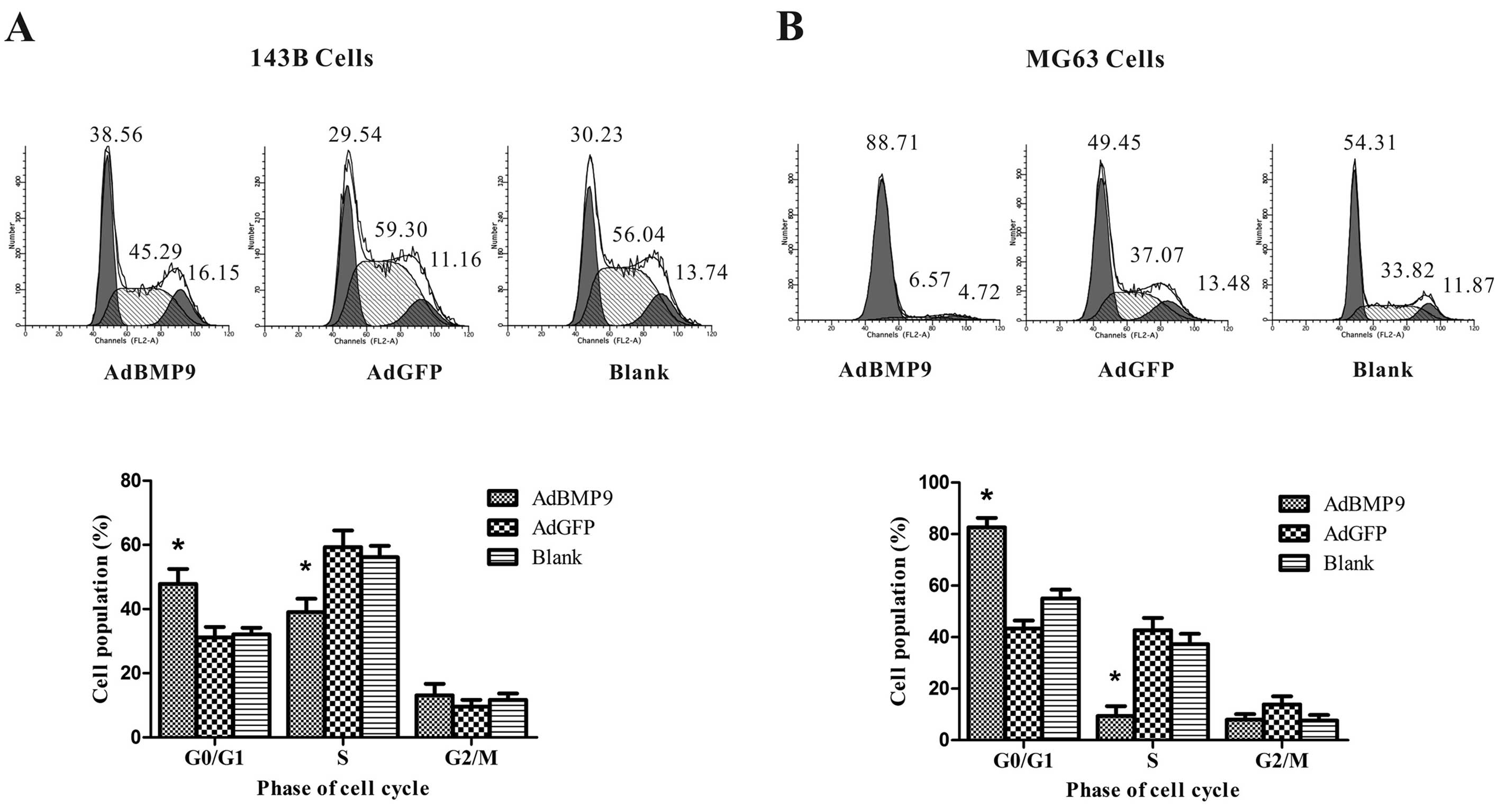
To determine whether the growth inhibition observed
in OS cells is associated with induction of cell cycle arrest, we
next examined the effects of BMP9 overexpression on OS cell cycle
distribution by flow cytometry. The data showed that BMP9
overexpression decreased the percentage of 143B and MG63 cells in
the S phase from 59.32±5.16 to 39.07±4.16% (p<0.05) and
42.75±4.69 to 9.41±3.82% (p<0.05) compared to the AdGFP group,
and increased the percentage of 143B and MG63 cells at the G1 phase
of the cell cycle from 31.14±3.29 to 47.83±4.66% (p<0.05) and
43.36±3.11 to 82.65±3.68% (p<0.05) compared to the AdGFP group,
respectively (Fig. 2).
BMP9 overexpression induces apoptosis in
143B cells
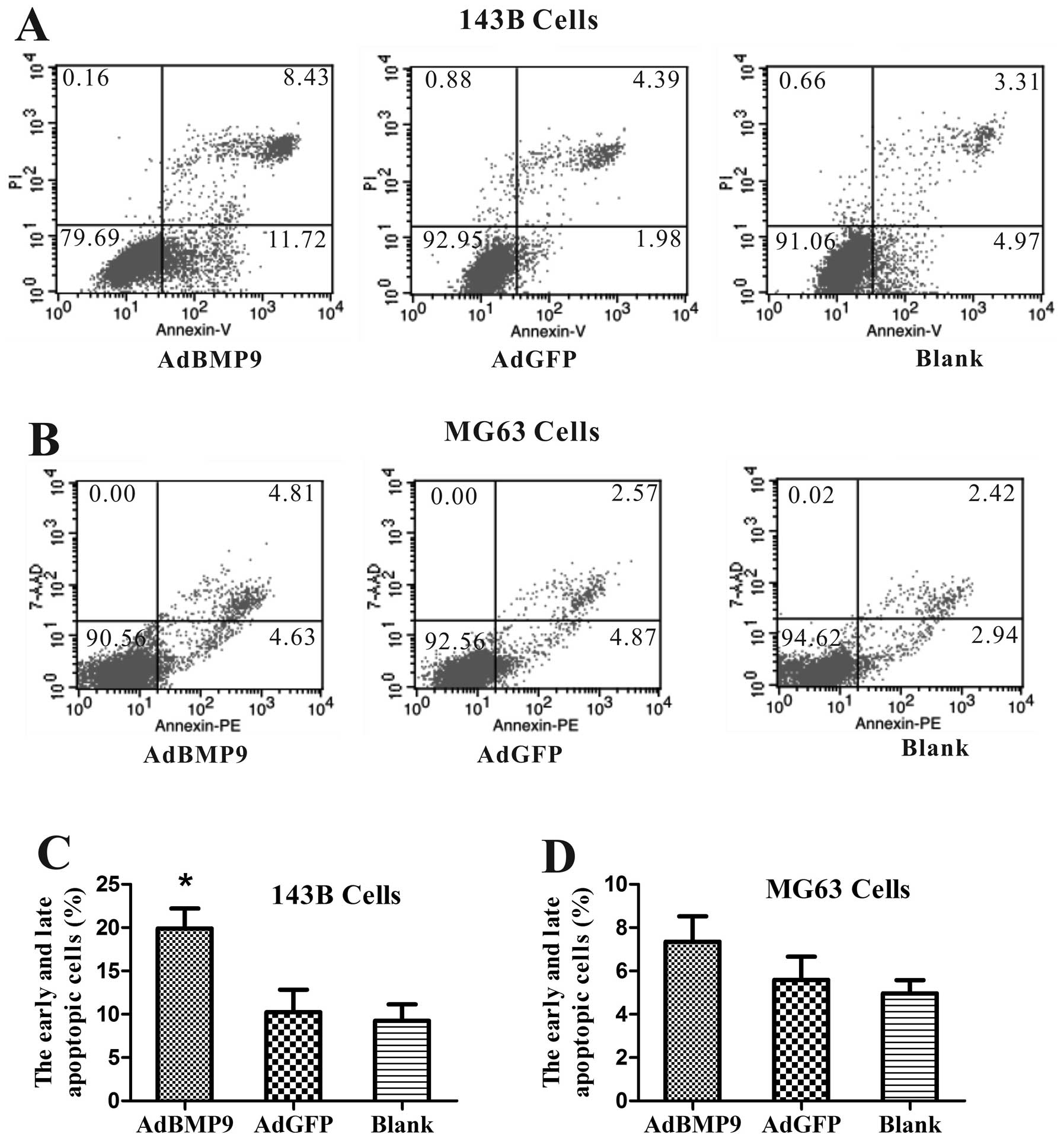
We then examined whether BMP9 overexpression induces
apoptosis in OS cells by flow cytometry. Fig. 3A and C shows that the apoptosis rate
of 143B cells was significantly increased from 10.23±2.61 to
19.91±2.29% (p<0.05) compared to AdGFP groups after BMP9
treatment for 48 h, whereas the apoptosis rate of MG63 cells was
not significantly higher than that of the AdGFP groups after BMP9
treatment for 48 h (Fig. 3B and
D).
Expression of β-catenin and its
downstream target genes depend on BMP9 expression
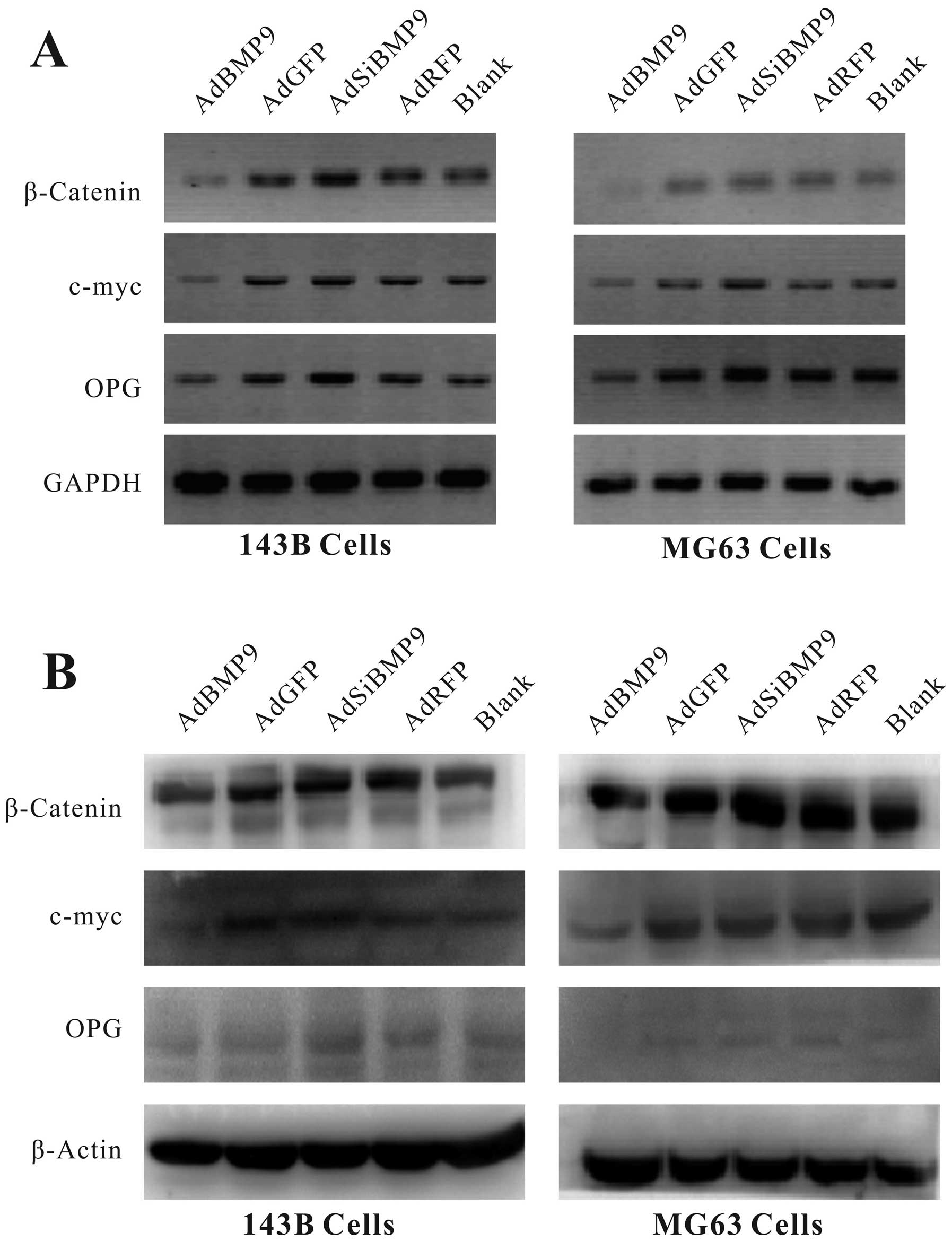
To determine β-catenin and further understand the
regulation of the several target genes by β-catenin, we detected
β-catenin and its downstream target genes c-myc and OPG using
RT-PCR and western blot analysis. As shown in Fig. 4, our results demonstrated that
levels of β-catenin and its target genes c-myc and OPG were
significantly reduced at both the mRNA and protein levels in OS
cells after infection with AdBMP9 for 24 h, whereas they reverted
after infection with AdsiBMP9.
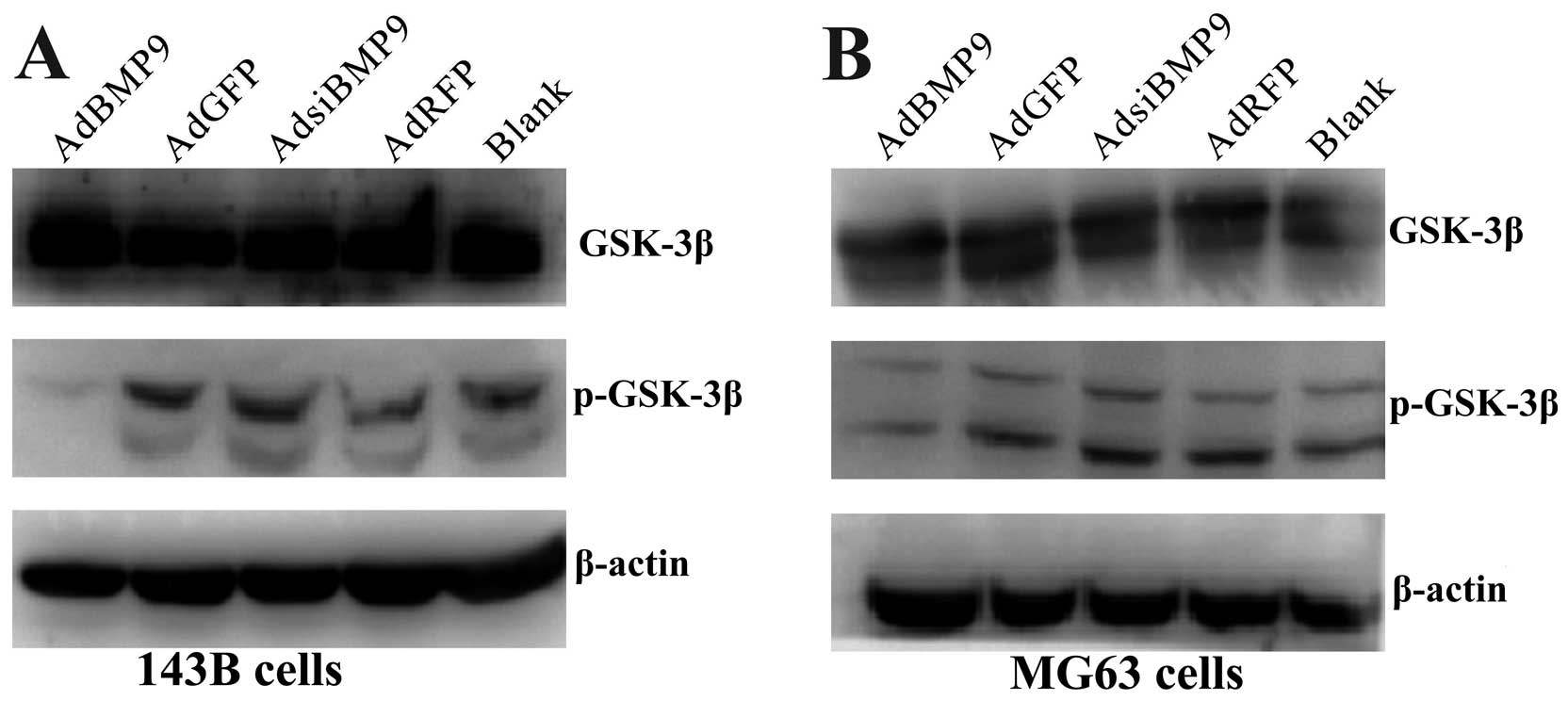
GSK-3β activity depends on BMP9
expression
To evaluate the effects of BMP9 on phosphorylated
GSK-3βSer9, the total GSK-3β and phosphorylated
GSK-3βSer9 were determined by western blotting. Fig. 5 shows that BMP9 overexpression
significantly reduced GSK-3βSer9 phosphorylation in OS
cells and the suppressive effect was reverted by inhibition of
BMP9. This data indicates that BMP9 might downregulate β-catenin by
activating GSK-3β.
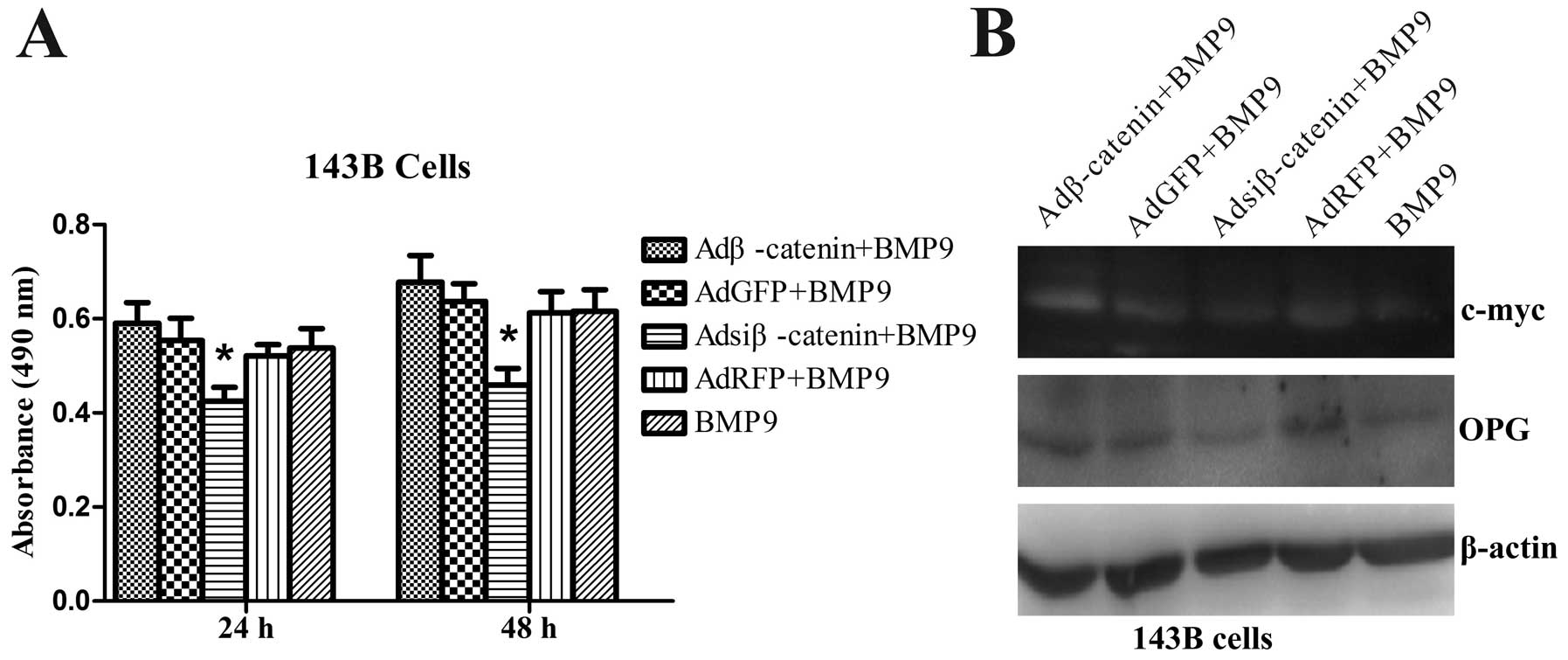
BMP9 inhibits the growth of OS cells
through the Wnt/β-catenin signaling pathways
To determine whether the effect of BMP9 on OS cells
is associated with Wnt/β-catenin signaling, cell viability was
determined by MTT assays. The efficiency of Adβ-catenin and
Adsiβ-catenin was previously confirmed by our group (17); we observed that the proliferation
potency of exogenous β-catenin expression of 143B cells treated
with BMP9 was restored compared with AdGFP infection treated with
BMP9, whereas knockout of β-catenin of 143B cells treated with BMP9
was enhanced compared with AdRFP infection treated with BMP9
(Fig. 6A). Similarly, β-catenin
target genes c-myc and OPG expression were restored after exogenous
β-catenin expression of 143B cells treated with BMP9, whereas they
were reduced after knockout of β-catenin expression of 143B cells
treated with BMP9, compared with AdGFP or AdRFP infection treated
with BMP9, respectively (Fig. 6B).
The data showed that BMP9 overexpression inhibits cell viability of
143B cells possibly through the Wnt/β-catenin pathway.
Discussion
BMPs are a member of the TGF-β superfamily and were
originally identified to play important roles in regulating bone
and cartilage formation (18). More
recently, BMPs were found to express various human cancer cells
including prostate, gastric, ovarian, pancreatic, colon carcinoma,
breast cancer, OS and multiple myeloma (11,19).
Recent studies suggest that BMP2 is involved in OS progression
(20). BMP9 was reported to
probably be the most potent inducer (21) and we previously showed it is
expressed in OS cell lines 143B and MG63 and its overexpression
inhibits OS cell migration and invasion (22); however, the effect of BMP9 on OS has
not yet been fully elucidated.
In the present study, we observed that BMP9
overexpression had a significant inhibitory effect on OS cell
viability in a time-dependent manner, whereas BMP9 silencing
induced opposite effects. A possible explanation for the growth
inhibition of BMP9 on OS cells is either cell cycle arrest or
apoptosis increase. Our results indicated that BMP9 induced cell
cycle arrest in the G1 phase in the two OS cell lines, but only
induced cell apoptosis of 143B cells. This may explain the
different metastatic properties of the two OS cell lines. This
result is consistent with previous studies that have shown an
inhibitory effect of BMP9 overexpression on cancer cell growth
including prostate and breast cancer (15,16).
However, our findings on OS cell lines in vitro differ from
previous observations that BMP9 induces proliferation of several
cells, including ovarian cancer and human hepatocellular carcinoma
(13,14). The inconsistency of the effects of
BMP9 on cancer cells may result from dosage, type of cell or/and
microenvironment differences.
Previous studies have demonstrated that deregulation
of β-catenin signaling is heavily implicated in the development and
progression of various types of cancer including colorectal,
melanoma, gastric, hepatic breast cancer and OS (23,24).
β-catenin signaling is commonly deregulated in human OS, which is
implicated in the pathogenesis of OS (6,7). In
addition, restoration of BMP signaling decreased β-catenin
expression and inhibited Wnt signaling in colon cancer (25). To further investigate the mechanisms
of BMP9 on the regulation of cell viability, the level of β-catenin
was determined by RT-PCR and western blot assay. Our results
revealed that BMP9 overexpression downregulated β-catenin
expression, whereas BMP9 silencing induced opposite effects.
BMPs can regulate OPG expression and bone-derived
OPG was found to protect several cancer cells from TRAIL-induced
apoptosis including breast cancer, prostate cancer and multiple
myeloma cells (10,26–28).
Moreover, OPG expression was found to be negatively correlated with
increasing breast tumor grade (29). Our data showed that BMP9
overexpression decreased expression of c-myc and OPG, whereas
knockout of BMP9 reverted these effects. We also showed that
exogenous β-catenin expression can neutralize these effects of
BMP9, whereas knockout of β-catenin can enhance these effects of
BMP9. Collectively, we hypothesized that BMP9 inhibits the growth
of OS cells possibly through the Wnt/β-catenin pathway.
GSK-3β phosphorylates β-catenin, followed by rapid
degradation of β-catenin via the ubiquitin-proteasome pathway,
which regulates processes including differentiation, growth,
motility and apoptosis (8,30,31).
Our results showed that BMP9 overexpression downregulated
p-GSK-3βSer9 protein levels, whereas BMP9 silencing
induced opposite effects. These findings are compatible with the
hypothesis that the activated GSK-3β resulted in G1 cell-cycle
arrest and apoptosis. In contrast, inactivation of GSK-3β leads to
cell-cycle progression and resistance to apoptosis in human lung
cancer cells (32). However,
whether GSK-3β activation is critical for BMP9-inhibited growth of
OS cells requires further investigation.
In conclusion, our results suggest that BMP9
overexpression inhibits OS cell growth, induces cell G1 phase
arrest and apoptosis, and these suppressive effects may be mediated
by the Wnt/β-catenin pathway. We also demonstrated that BMP9
overexpression increased GSK-3β activity by decreasing its
phosphorylation on ser 9 residue GSK-3βSer9; therefore,
we further propose that inhibiting Wnt/β-catenin may be through
increasing GSK-3β activity. Our results indicate that BMP9
overexpression may prove to be a valuable tool for inhibition of
cancer growth. Further studies to elucidate the relationship
between BMP signaling and Wnt signaling are required.
Acknowledgements
We are grateful to Dr T.C. He of The University of
Chicago Medical Center for providing the adenoviruses. This study
was supported by National Ministry of Education Foundation of China
(20115503110009), grant 31200971 from the National Natural Science
Foundation of China (NSFC31200971), by Program of the Ministry of
Science and Technology of Yu-zhong District, CQ (20130136) and by
the Science and Technology Research Project of Chongqing Education
Commission (KJ120327).
References
|
1
|
Young JL Jr and Miller RW: Incidence of
malignant tumors in U.S. children. J Pediatr. 86:254–258. 1975.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fagioli F, Biasin E, Mereuta OM, et al:
Poor prognosis osteosarcoma: new therapeutic approach. Bone Marrow
Transpl. 41:S131–S134. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barker N: The canonical Wnt/beta-catenin
signalling pathway. Methods Mol Biol. 468:5–15. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Behrens J: Control of beta-catenin
signaling in tumor development. Ann NY Acad Sci. 910:21–35. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Haydon RC, Deyrup A, Ishikawa A, et al:
Cytoplasmic and/or nuclear accumulations of the beta-catenin
protein is a frequent event in human osteosarcoma. Int J Cancer.
102:338–342. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iwaya K, Ogawa H, Kuroda M, Izumi M,
Ishida T and Mukai K: Cytoplasmic and/or nuclear staining of
beta-catenin is associated with lung metastasis. Clin Exp
Metastasis. 20:525–529. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McQueen P, Ghaffar S, Guo Y, Rubin EM, Zi
X and Hoang BH: The Wnt signaling pathway: implications for therapy
in osteosarcoma. Expert Rev Anticancer Ther. 11:1223–1232. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sato MM, Nakashima A, Nashimoto M, Yawaka
Y and Tamura M: Bone morphogenetic protein-2 enhances
Wnt/beta-catenin signaling-induced osteoprotegerin expression.
Genes Cells. 14:141–153. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Singh A and Morris RJ: The Yin and Yang of
bone morphogenetic proteins in cancer. Cytokine Growth Factor Rev.
21:299–313. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim M and Choe S: BMPs and their clinical
potentials. BMB Rep. 44:619–634. 2011. View Article : Google Scholar
|
|
13
|
Herrera B, van Dinther M, ten Dijke P and
Inman GJ: Autocrine bone morphogenetic protein-9 signals through
activin receptor-like kinase-2/Smad1/Smad4 to promote ovarian
cancer cell proliferation. Cancer Res. 69:9254–9262. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Herrera B, García-Álvaro M, Cruz S, et al:
BMP9 is a proliferative and survival factor for human
hepatocellular carcinoma cells. PloS One. 8:e695352013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ye L, Kynaston H and Jiang WG: Bone
morphogenetic protein-9 induces apoptosis in prostate cancer cells,
the role of prostate apoptosis response-4. Mol Cancer Res.
6:1594–1606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang K, Feng H, Ren W, et al: BMP9
inhibits the proliferation and invasiveness of breast cancer cells
MDA-MB-231. J Cancer Res Clin. 137:1687–1696. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu Y, Wang W, Xu J, et al:
Dihydroartemisinin inhibits tumor growth of human osteosarcoma
cells by suppressing Wnt/β-catenin signaling. Oncol Rep.
30:1723–1730. 2013.PubMed/NCBI
|
|
18
|
Chen D, Zhao M and Mundy GR: Bone
morphogenetic proteins. Growth Factors. 22:233–241. 2004.
View Article : Google Scholar
|
|
19
|
Grcevic D, Kusec R, Kovacic N, et al: Bone
morphogenetic proteins and receptors are over-expressed in
bone-marrow cells of multiple myeloma patients and support myeloma
cells by inducing ID genes. Leukemia Res. 34:742–751. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang L, Park P, Zhang H, et al: BMP-2
inhibits the tumorigenicity of cancer stem cells in human
osteosarcoma OS99-1 cell line. Cancer Biol Ther. 11:457–463. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kang Q, Sun MH, Cheng H, et al:
Characterization of the distinct orthotopic bone-forming activity
of 14 BMPs using recombinant adenovirus-mediated gene delivery.
Gene Ther. 11:1312–1320. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lv Z, Yang D, Li J, et al: Bone
morphogenetic protein 9 overexpression reduces osteosarcoma cell
migration and invasion. Mol Cells. 36:119–126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Luo J, Chen J, Deng Z-L, et al: Wnt
signaling and human diseases: what are the therapeutic
implications? Lab Invest. 87:97–103. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Flores RJ, Li Y, Yu A, et al: A systems
biology approach reveals common metastatic pathways in
osteosarcoma. BMC Syst Biol. 6:502012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Freeman TJ, Smith JJ, Chen X, et al:
Smad4-mediated signaling inhibits intestinal neoplasia by
inhibiting expression of β-catenin. Gastroenterology. 142:562–571.
2012.PubMed/NCBI
|
|
26
|
Neville-Webbe HL, Cross NA, Eaton CL, et
al: Osteoprotegerin (OPG) produced by bone marrow stromal cells
protects breast cancer cells from TRAIL-induced apoptosis. Breast
Cancer Res Treat. 86:269–279. 2004.PubMed/NCBI
|
|
27
|
Nyambo R, Cross N, Lippitt J, et al: Human
bone marrow stromal cells protect prostate cancer cells from
TRAIL-induced apoptosis. J Bone Miner Res. 19:1712–1721. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shipman CM and Croucher PI:
Osteoprotegerin is a soluble decoy receptor for tumor necrosis
factor-related apoptosis-inducing ligand/Apo2 ligand and can
function as a paracrine survival factor for human myeloma cells.
Cancer Res. 63:912–916. 2003.
|
|
29
|
Holen I, Cross SS, Neville-Webbe HL, et
al: Osteoprotegerin (OPG) expression by breast cancer cells in
vitro and breast tumours in vivo - a role in tumour cell survival?
Breast Cancer Res Treat. 92:207–215. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fodde R and Brabletz T: Wnt/beta-catenin
signaling in cancer stemness and malignant behavior. Curr Opin Cell
Biol. 19:150–158. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Forde JE and Dale TC: Glycogen synthase
kinase 3: A key regulator of cellular fate. Cell Mol Life Sci.
64:1930–1944. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li J, Xing M, Zhu M, et al: Glycogen
synthase kinase 3 beta induces apoptosis in cancer cells through
increase of survivin nuclear localization. Cancer Lett. 272:91–101.
2008. View Article : Google Scholar : PubMed/NCBI
|