Introduction
Colorectal cancer is one of the most prevalent
cancers and a major cause of mortality worldwide (1). Although adjuvant systemic chemotherapy
or chemoradiation can confer a limited but significant survival
advantage, novel and more effective therapies are needed. To
improve survival rates, new therapeutic agents have been
investigated. Immunotherapy for colorectal cancer is a promising
candidate treatment, and there is evidence that host immune
responses can influence survival (2). Ideal targets for immunotherapy are
gene products overexpressed in cancer cells but silenced in normal
tissues, with the exception of immune-privileged tissues, such as
that of the testis.
We previously reported that heat shock protein 105
(HSP105), identified by SEREX, is overexpressed in a variety of
human cancers, including colorectal, pancreatic and esophageal
cancer, but with little to no expression in normal tissues aside
from the testis (3,4). HSP105 is a stress protein induced by
various stressors and belongs to the HSP105/110 family and plays an
important role as a chaperone under physiological conditions
(5). Using immunohistochemical
analysis, we previously found that HSP105 was specifically
overexpressed in 44 of 53 (83.0%) colorectal cancer patients
(4). It has also been reported that
DNA vaccination with both HSP105 and bone marrow-derived dendritic
cells (BM-DCs) pulsed with HSP105 led to tumor rejection of
colorectal cancer but did not induce an autoimmune reaction in mice
(6–8).
This suggests that HSP105 presents a useful
tumor-specific antigen target for immunotherapy. However,
HSP105-derived epitope peptides of CD8+ T cells have not
been identified. The gene frequency of HLA-A24 (A*24:02) is
relatively high in Asian populations, especially the Japanese, but
low in Caucasians. On the other hand, the gene frequency of HLA-A2
(A*02:01) is high among several ethnic groups, including Asians and
Caucasians (9). Therefore, HLA-A2
or HLA-A24-restricted cytotoxic T cell (CTL) HSP105 epitopes could
be extremely useful for immunotherapy in a large portion of
patients worldwide. In the present study, we identified human
HSP105-derived CTL epitopes restricted by HLA-A2 or HLA-A24 using
HLA-transgenic mice (Tgm) and examined whether these epitope-based
peptides could activate HSP105-reactive CTLs in peripheral blood
mononuclear cells (PBMCs) of patients with colorectal cancer.
Materials and methods
Mice
HLA-A2.1 (HHD) Tgm,
H-2Db−/−β2m−/− double-knockout mice
introduced with the human β2m-HLA-A2.1(α1 α2)-H-2Db (α3
transmembrane cytoplasmic) (HHD) mono-chain gene construct were
generated in the Departmente SIDA-Retrovirus, Unite d’ Immunite
Cellulaire Antivirale, Institut Pasteur, Paris, France (10,11)
and were kindly provided by Dr F.A. Lemonier. HLA-A24.2 (HHD) Tgm
were purchased from Japan SLC, Inc. (Shizuoka, Japan). Female 6- to
8-week-old BALB/c mice (H-2Kd) and BALB/c nude mice,
purchased from Charles River Japan (Yokohama, Japan), were
maintained and handled in accordance with animal care policy.
Cell lines
The human colorectal cancer cell line SW620
(endogenously expressing HSP105 and HLA-A*02:01, 24:02) and human
liver cancer cell line HepG2 (HSP105-low expressing and
HLA-A*02:01, 24:02), were kindly provided by the Cell Resource
Center for Biomedical Research, Institute of Development, Aging and
Cancer (Tohoku University, Sendai, Japan). Murine colorectal cancer
cells, Colon26 (C26) (endogenously expressing HSP105 and
H-2Kd) were kindly provided by Dr Kyoichi Shimomura
(Fujisawa Pharmaceutical Co., Osaka, Japan). T2 cells (a
TAP-deficient and HLA-A*02:01-positive cell line) were provided by
Kyogo Ito of Kurume University. Cells were maintained in
vitro in RPMI-1640 or DMEM supplemented with 10% FCS.
RNA interference
Small interfering RNAs targeting human HSP105 were
chemically synthesized by Dharmacon Research (HSP105-siRNA and
luciferase; Lafayette, CO, USA) as previously described (12), with the following siRNA sequences:
HSP105-siRNA, UUGGCUGCAACUCCGAUU GTT and luciferase,
CGUACGCGGAAUACUUCGATT. The transfection of siRNA oligonucleotides
was carried out using Oligofectamine (Invitrogen, Carlsbad, CA,
USA) according to the manufacturer’s guidelines.
Peptides
Human HSP105-derived peptides, identical in amino
acid sequence with mouse HSP105 and expressing the binding motifs
for HLA-A*02:01- and HLA-A*24:02-encoded molecules, were designed
with BIMAS software (BioInformatics and Molecular Analysis Section;
Center for Information Technology, NIH, MD, USA). We purchased a
total of 16 versions of peptides carrying the HLA-A2
(A*0201)-binding motifs and 9 versions of peptides carrying the
HLA-A24 (A*2402)-binding motifs from Biologica (Tokyo, Japan)
(Table I).
 | Table IHSP105-derived peptides conserved
between human and mouse HSP105 predicted to bind to HLA-A2 or
HLA-A24. |
Table I
HSP105-derived peptides conserved
between human and mouse HSP105 predicted to bind to HLA-A2 or
HLA-A24.
| Peptides | Position | Subsequent residue
listing | HLA-A2 binding
score |
|---|
| HSP105 A2-4 | 120–128 | MLLTKLKET | 107 |
| HSP105 A2-5 | 141–149 | VISVPSFFT | 55 |
| HSP105 A2-6 | 155–163 | SVLDAAQIV | 37 |
| HSP105 A2-7 | 169–177 | RLMNDMTAV | 591 |
| HSP105 A2-9 | 202–210 | DMGHSAFQV | 21 |
| HSP105 A2-10 | 222–230 | VLGTAFDPFL | 759 |
| HSP105 A2-12 | 275–284 | KLMSSNSTDL | 276 |
| HSP105 A2-13 | 276–284 | LMSSNSTDL | 26 |
| HSP105 A2-14 | 300–309 | KMNRSQFEEL | 50 |
| HSP105 A2-15 | 304–313 | SQFEELCAEL | 32 |
| HSP105 A2-16 | 313–321 | LLQKIEVPL | 36 |
| HSP105 A2-19 | 434–442 | FLRRGPFEL | 43 |
| HSP105 A2-20 | 458–467 | KIGRFVVQNT | 76 |
| HSP105 A2-25 | 668–676 | LLTETEDWL | 401 |
| HSP105 A2-26 | 675–684 | WLYEEGEDQA | 146 |
| HSP105 A2-29 | 757–765 | EVMEWMNNV | 15 |
|
| Peptides | Position | Subsequent residue
listing | HLA-A24 binding
score |
|
| HSP105 A24-1 | 180–188 | NYGIYKQDL | 240 |
| HSP105 A24-2 | 214–223 | AFNKGKLKVL | 30 |
| HSP105 A24-3 | 251–260 | KYKLDAKSKI | 110 |
| HSP105 A24-4 | 305–313 | QFEELCAEL | 47 |
| HSP105 A24-5 | 433–442 | TFLRRGPFEL | 33 |
| HSP105 A24-6 | 613–622 | MYIETEGKMI | 90 |
| HSP105 A24-7 | 640–649 | EYVYEFRDKL | 330 |
| HSP105 A24-8 | 725–733 | HYAKIAADF | 140 |
| HSP105 A24-9 | 739–748 | KYNHIDESEM | 82 |
Induction of HSP105-reactive CTLs in
Tgm
Peptide immunizations in mice were performed as
previously described (13). In
brief, bone marrow (BM) cells (2×106) from HLA-A2 or
HLA-A24 Tgm were cultured in RPMI-1640 medium supplemented with 10%
FCS, GM-CSF (5 ng/ml) and 2-mercaptoethanol (0.8 ng/ml) for 7 days
in 10-cm plastic dishes. These BM-DCs were pulsed with the two
HSP105 peptide mixtures (1 μmol/l each peptide) for 2 h at 37°C. We
primed the HLA-A2 or HLA-A24 Tgm with the syngeneic BM-DC vaccine
(5×105/mice) into the peritoneal cavity twice, once per
week. Seven days following the last immunization, the spleens were
collected and CD4− spleen cells were isolated by
negative selection with anti-CD4 microbeads (Miltenyi Biotec,
Bergisch Gladbach, Germany) to exclude any nonspecific IFN-γ
production from the CD4+ spleen cells co-cultured with
the BM-DCs. The CD4− spleen cells
(2×106/well) were stimulated with syngeneic BM-DCs
(2×105/well) that had been pulsed with each peptide
in vitro. After 6 days, the frequency of cells producing
IFN-γ/2×104 CD4− spleen cells upon
stimulation with syngeneic BM-DCs (1×104/well), pulsed
with or without each peptide, was assayed using an enzyme-linked
immunospot (ELISPOT) assay as previously described (13).
Identification of a CTL epitope in BALB/c
mice
The peptide immunizations in mice were performed as
previously described (14).
Splenocytes removed from mice 7 days following the last
immunization were harvested and cultured in 24-well culture plates
(2.5×106/well) in 45% RPMI, 45% AIMV, 10% FCS and
supplemented with recombinant human interleukin 2 (100 U/ml),
2-mercaptoethanol (50 μmol/l) and each peptide (10 μmol/l). After 5
days, the cytotoxicity of these cells against target cells was
assayed using standard 6-h 51Cr release assays (15).
Blood samples
Blood samples from cancer patients were collected
during routine diagnostic procedures after obtaining formal consent
from patients at the Kumamoto University Hospital, from April to
September 2006 and from patients at the National Cancer Center
Hospital East, from December 2006 to March 2007. The study was
approved by the local ethics committee, and informed consent was
obtained from all patients.
Induction of HSP105-reactive human
CTLs
We isolated PBMCs from heparinized blood of
HLA-A24+ and/or HLA-A2+ Japanese patients
with colorectal cancer using Ficoll-Conray density gradient
centrifugation; peripheral monocyte-derived dendritic cells (DCs)
were generated as previously described (16,17).
CD8+ T cells were isolated with CD8 microbeads (Miltenyl
Biotec, Bergisch Gladbach, Germany) from PBMCs of the same donor
and peptide-reactive CD8+ CTLs were generated. Five days
following the last stimulation, the cytotoxic activities of the
CTLs against cancer cell lines were measured by
51Cr-release assay as previously described (15). For these assays, CTLs were
co-cultured with each cancer cell line, as the target cells
(5×103/well), at the indicated effector/target
ratio.
In vivo tumor challenge
Subcutaneous tumors were induced in mice by
injecting 1×104 SW620 cells suspended in 100 μl PBS or
Hanks’ balanced salt solution (Gibco, Grand Island, NY, USA) into
the backs of BALB/c nude mice. Tumor incidence and volumes were
assessed weekly using calipers and tumor areas were measured.
Results are presented as mean tumor areas ± SD.
Ex vivo IFN-γ ELISPOT assay in peripheral
blood in pre-surgical colorectal cancer patients
Ex vivo IFN-γ ELISPOT assays were performed
to determine tumor-specific interferon-γ (IFN-γ)-secreting T cells.
The 96-well plates were coated with anti-human IFN-γ (BD
Biosciences Co., Ltd., USA). After an overnight incubation at 4°C,
the wells were washed and blocked with complete medium for 2 h at
room temperature. A total of 1×106 unfractionated PBMCs
were added in duplicate wells and incubated at 37°C for 18–20 h
with or without peptides at 0.2 μl/well (1–10 μM). The plate was
washed and then incubated with 5 μg/ml biotinylated anti-human
IFN-γ antibody for 2 h at room temperature. After washing away the
antibodies, streptavidin-HRP was added for 1 h. Finally, the plate
was washed and replaced with fresh substrate solution and the
reaction was terminated by washing with distilled water. The
HLA-A2-restricted CMV peptide (NLVPMVATV) and HLA-A24 restricted
CMV peptide (QYDPVAALF), which includes an epitope derived from the
CMV pp65 protein, were used as positive controls.
Histological and immunohistochemical
analysis
To investigate whether CD8+ T cells
infiltrated normal tissues triggered by the HSP105-derived peptide
vaccine, we performed immunohistochemical staining with a
monoclonal antibody against CD8 (1:100; LifeSpan BioSciences, Inc.,
Seattle, WA, USA) in tissue specimens from HLA-A2 Tgm immunized
with the HSP105 peptides, as previously described (7). Immunohistochemical staining with
rabbit polyclonal antibodies against HSP105 (1:200; Santa Cruz
Biotechnology, Inc.; Santa Cruz, CA, USA) was performed according
to the manufacturer’s instructions.
Results
Identification of HLA-A2-or
HLA-A24-restricted CTL epitopes in HLA Tgm
We designed pools of HSP105 peptides possessing
amino acid sequences conserved between humans and mice that have a
highly predicted binding score to HLA-A2 (pool of 16 different
peptides) or HLA-A24 (A*24:02) (pool of 9 different peptides)
(Table I). CD4− spleen
cells were obtained from Tgm immunized twice i.p. with BM-DCs that
had been pulsed with each peptide mixture; the spleen cells were
then stimulated in vitro, again with the BM-DCs pulsed with
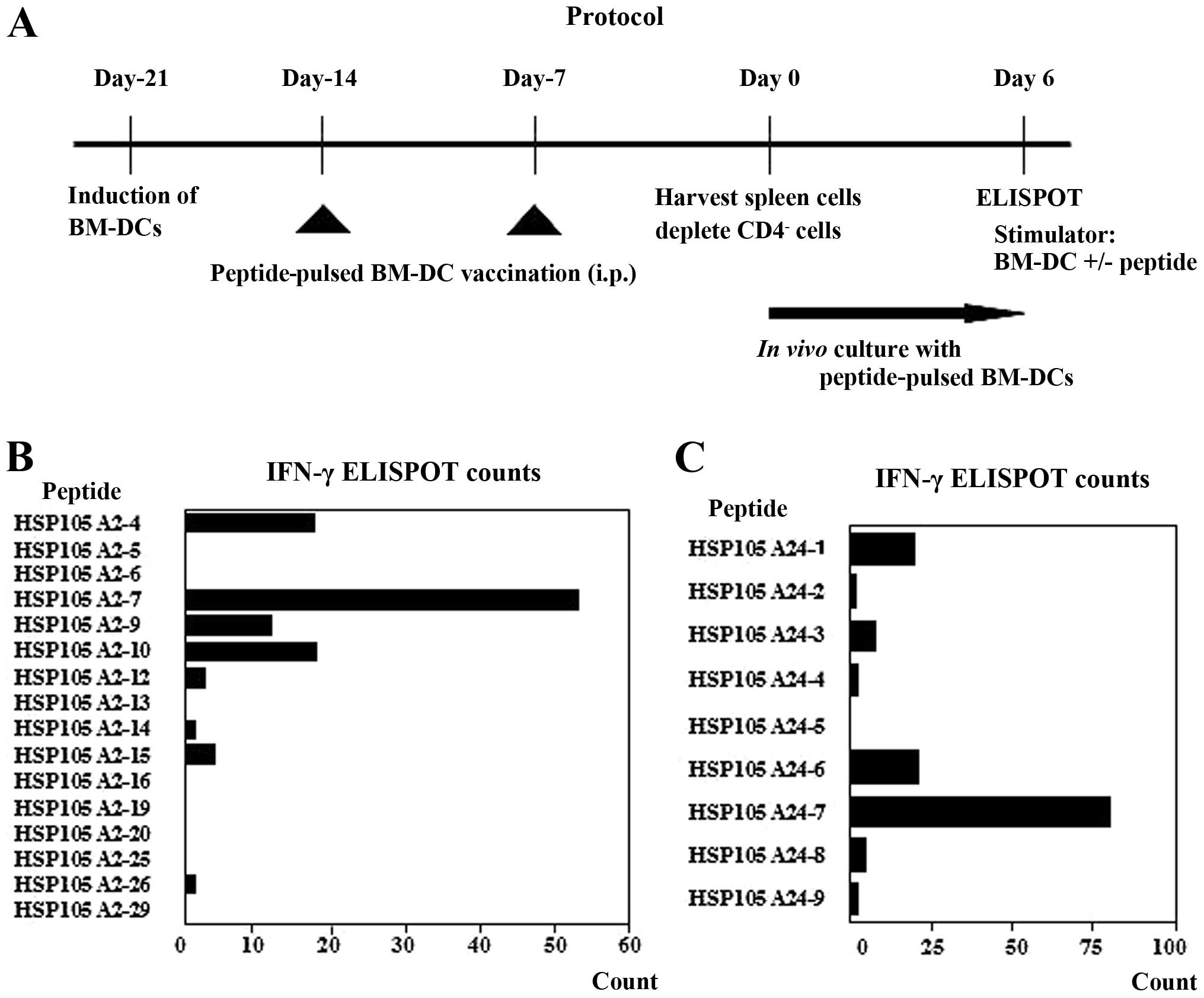
each peptide mixture (Fig. 1A).
The IFN-γ ELISPOT counts, normalized to those of
spleen cells co-cultured with BM-DCs without peptide loading,
clearly indicated a HSP105 A2-7 peptide-specific response in the
CD4− spleen cells (Fig.
1B). These CD4− spleen cells (2×104/well)
showed 55±29.7 spot counts/well in response to the BM-DCs pulsed
with the HSP105 A2-7 peptide, whereas they showed 23±31.1 spot
counts/well in the presence of BM-DCs pulsed with the HSP105 A2-4
peptide. A similarly strong response was observed for the HSP105
A24-7 peptide (Fig. 1C).
CD4− spleen cells (2×104/well) showed
79.5±27.6 spot counts/well in response to the BM-DCs pulsed with
the HSP105 A24-7 peptide, whereas they showed 20.5±14.8 spot
counts/well in the presence of BM-DCs with the HSP A24-6 peptide.
These assays were performed twice with similar results and they
suggest that the HSP105 A2-7 and A24-7 peptides are potential CTL
epitope peptides in both HLA Tgm and humans.
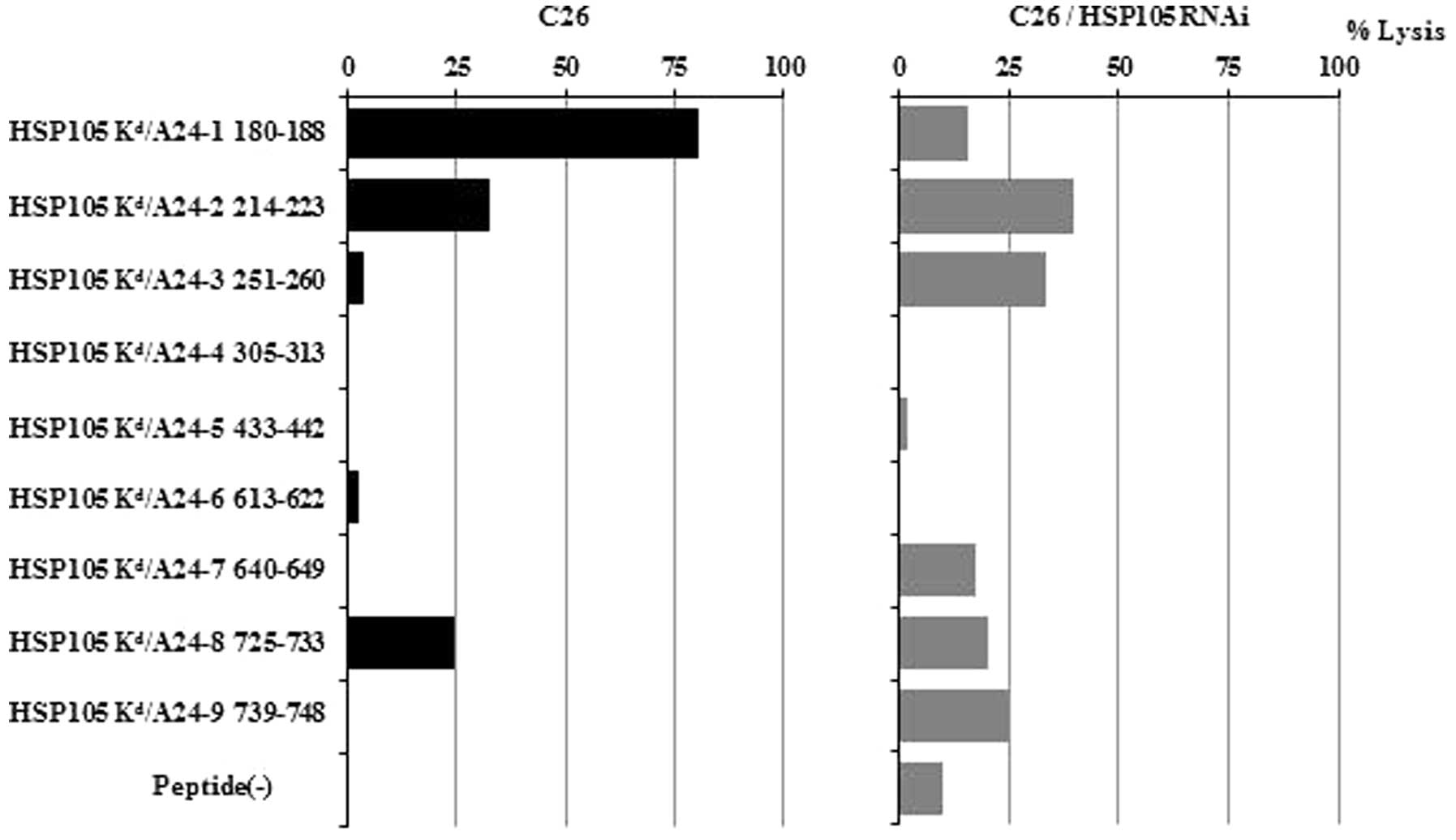
Identification of a CTL epitope in BALB/c
mice and CTLs that are cytotoxic against C26 tumors in mice
There were similar structural motifs within the
peptides that bound to human HLA-A24 and mice Kd. We
selected those peptides with binding motifs for both HLA-A24 and
Kd molecules and prepared 9 different synthetic peptides
(HSP105-1-9). When we tested these peptides for their potential to
induce in vitro tumor reactive CTLs in spleen cells derived
from BALB/c mice immunized with the HSP105 peptides, only the
HSP105 24-1 peptide-induced CTLs showed specific cytotoxicity
against C26 tumors (HSP105+, H-2Kd) (Fig. 2). The cytotoxicity against C26 was
attenuated by HSP105 siRNA. These findings indicate that the HSP105
A24-1 peptide has the capacity to induce tumor reactive CTLs and
that peptide vaccination-primed CTLs are reactive to this peptide
in vivo. We would expect this HSP105 A24-1 (NYGIYKQDL)
peptide to also be an epitope for human CTLs.
HSP105-reactive CTLs from PBMCs of
HLA-A2-positive colorectal cancer patients and CTLs induce
cytotoxicity against HSP105-expressing cancer cells
We generated a CTL line from PBMCs of colorectal
patients by stimulation with the HSP105 A2-12 peptide. As shown by
51Cr release assays, the resulting CTLs showed
HSP105-specific cytotoxicity against SW620 cells
(HSP105+++, HLA-A2) and against T2 cells pulsed with the
HSP105 A2-12 peptide (HSP105−, HLA-A2), but not against
HepG2 cells (HSP105±, HLA-A2) or T2 cells pulsed with an
irrelevant peptide (Fig. 3A).
HSP105 siRNA decreased the cytotoxicity against SW620 cells. We
investigated the effects of the HSP105 A2-12 peptide-reactive CTL
lines on the mice implanted with the SW620 cells. Fourteen days
after inoculation of HSP105 A2-12 peptide-reactive CTLs, there was
an apparent reduction in tumor size in the SW620 compared to that
in untreated mice (Fig. 3B). These
results clearly indicate the efficacy of HSP105 A2-12 (KLMSSNSTDL)
peptide-reactive CTL injection therapy for HSP105+
tumors in mice.
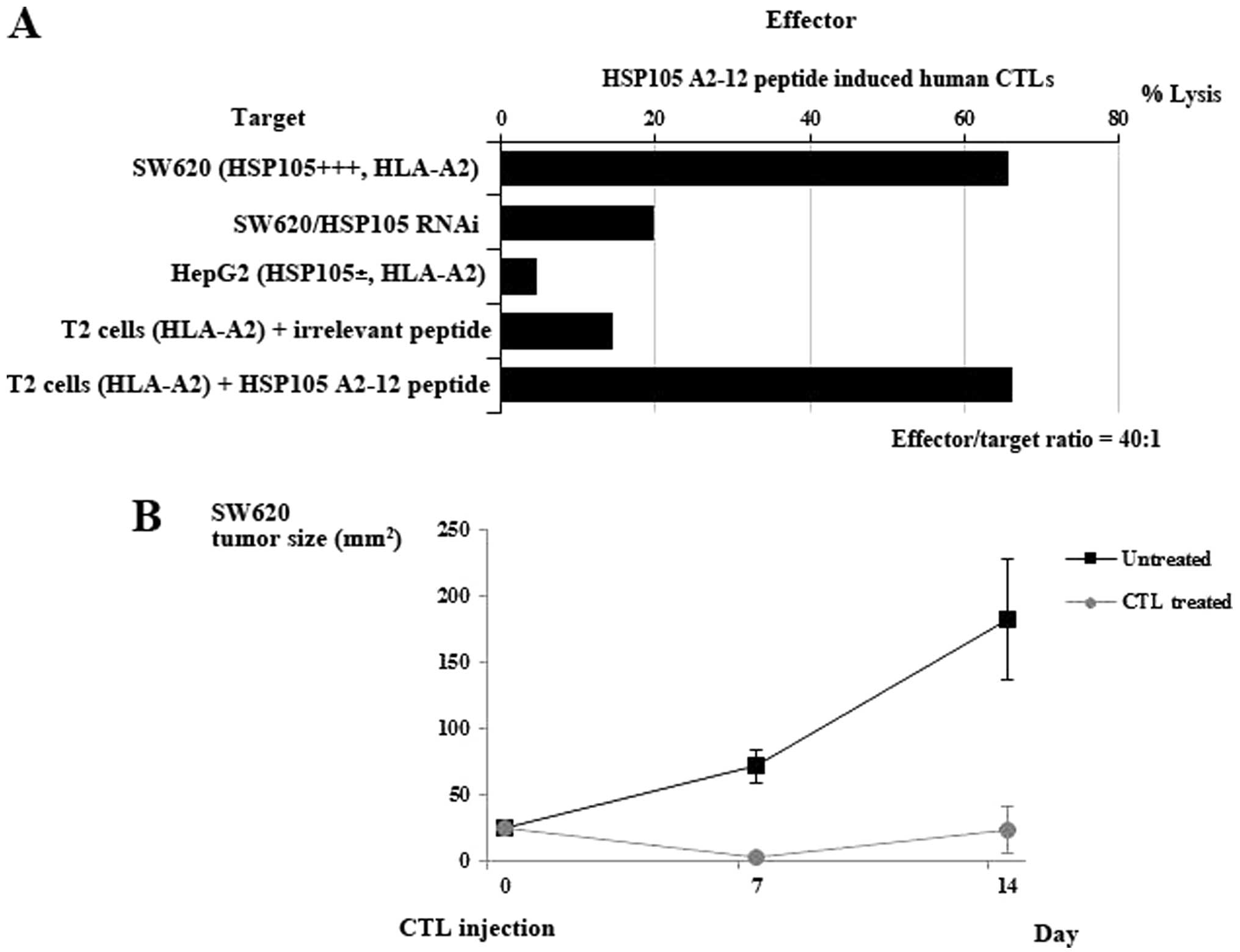 | Figure 3CTL induction from PBMCs of
HLA-A2-positive cancer patients. (A) HSP105 peptide-reactive CTLs
were generated from CD8+ T cells of HLA-A2+
colorectal cancer patients. After three or four stimulations with
autologous monocyte-derived DCs pulsed with the HSP105 A2-12
peptides, the CTLs were subjected to a standard 51Cr
release assay at the indicated effector/target ratio (40/1). Their
cytotoxicity against SW620 cells (HSP105+++, HLA-A2),
SW620 cells transfected with HSP105 siRNA (HSP105−),
HepG2 cells (HSP105±, HLA-A2), T2 cells pulsed with an
irrelevant peptide (HSP105−, HLA-A2) and T2 cells pulsed
with the HSP105 A2-12 epitope peptide were all examined by
51Cr release assay. Values represent the percentage of
specific cell lysis, based on the mean values from triplicate
assays. (B) There was marked growth inhibition of SW620 cells
(HSP105+) engrafted into nude mice after intratumoral
injection of human CTLs induced by the HSP105 peptides. When tumor
size reached 25 mm2 on day 9 after s.c. tumor
implantation, human CTLs (3×106) reactive to the
HLA-A2-restricted HSP105 peptide, generated from an
HLA-A2+ donor, were i.t. inoculated. Tumor sizes in nude
mice administered the HSP105 epitope peptide-induced CTL lines
(n=3), or no treatment (n=3), are shown. The mean tumor size
(mm2) for each group of mice was expressed, and bars
represent SD. |
Detection of HSP105-specific CTLs in
peripheral blood of pre-surgical patients with colorectal
cancer
Our results suggest that the four peptides, HSP105
A2-7 (RLMNDMTAV), HSP105 A2-12 (KLMSSNSTDL), HSP105 A24-1 (NYGIYK
QDL) and HSP105 A24-7 (EYVYEFRDKL), are HSP105-derived, HLA-A2, or
HLA-A24-restricted CTL epitopes. To determine the frequencies of
the HSP105-derived T cells specific for these peptide in
pre-surgical colorectal cancer patients, we analyzed the PBMC
responses for each peptide using the ELISPOT assay. HSP105
expression was detected in 20 of 21 (95%) patients, consistent with
previous studies (4).
HSP105-specific T cells secreting IFN-γ were detected in patients
stimulated with the HSP105 A2-7 (4 patients), HSP105 A2-12 (6
patients), HSP105 A24-1 (2 patients) and HSP105 A24-7 (6 patients)
peptides (Table II). ELISPOT assay
detected positive IFN-γ responses to at least one of the
HSP105-derived peptides in PBMCs in 14 of the 21 patients. In
contrast to the results for colorectal cancer patients, the 4
peptides were not recognized by PBMCs from healthy donors. Both the
ratio of normal donors who showed positive T-cell responses to
CMV-derived peptides and the frequencies of the specific T cells
were identical to those of the colorectal cancer patients (data not
shown).
| Table IIExpression of HSP105 in colorectal
cancer tissue and quantification of HSP-specific CTLs in colorectal
cancer patients. |
Table II
Expression of HSP105 in colorectal
cancer tissue and quantification of HSP-specific CTLs in colorectal
cancer patients.
| | | | | | cSpot number of peptide-specific
CTLs |
|---|
| | | | | |
|
|---|
| HLA-A2-positive
patients | Age (yrs.) | Gender | HLA | Stagea of tumor | HSP105
expressionb | HSP105 A2-7 | HSP105 A2-12 | CMV |
|---|
| 1 | 62 | M | 0201/2601 | IIIB | ++ | 27 | + | 126 | + | 160 |
| 5 | 79 | M | 0207/1101 | IIIB | ++ | 0 | − | 2 | − | 10 |
| 6 | 51 | M | 0201/0206 | I | + | 0 | − | 49 | + | 136 |
| 8 | 55 | M | 0206/2402 | I | ± | 0 | − | 0 | − | 66 |
| 11 | 69 | M | 0206/2402 | IIIC | + | 143 | + | 0 | − | 0 |
| 12 | 61 | M | 0201/3303 | I | ± | 2 | − | 45 | + | 367 |
| 13 | 64 | F | 0201/2601 | IIIC | ± | 0 | − | 2 | − | 254 |
| 14 | 66 | M | 0206/2402 | IIIC | − | 13 | + | 0 | − | 58 |
| 15 | 78 | M | 0201/1101 | IIA | + | 0 | − | 5 | + | 57 |
| 16 | 51 | F | 0206/2601 | IV | ± | 31 | + | 7 | + | 15 |
| 17 | 63 | F | 0206/1101 | IIA | ++ | 0 | − | 25 | + | 96 |
|
| HLA-A2402-positive
patients | | | | | | HSP105 A24-1 | HSP105 A24-7 | CMV |
|
| 2 | 64 | F | 2402 | IV | ++ | 2 | − | 44 | + | 6 |
| 3 | 60 | M | 2402/3101 | IIIC | ++ | 0 | − | 0 | − | 11 |
| 4 | 71 | F | 2402/3101 | IIA | ++ | 25 | + | 51 | + | 12 |
| 7 | 47 | M | 2402/3101 | IIIA | ++ | 4 | − | 6 | + | 3 |
| 9 | 66 | M | 2402 | IV | ++ | 8 | + | 6 | + | 7 |
| 10 | 60 | M | 2402/3101 | I | ++ | 1 | − | 19 | + | 26 |
| 18 | 64 | M | 1101/2402 | IV | + | 0 | − | 2 | − | 40 |
| 20 | 46 | F | 1101/2402 | IIIB | ++ | 4 | − | 7 | + | 5 |
| 21 | 66 | F | 2402 | I | ++ | 3 | − | 0 | − | 38 |
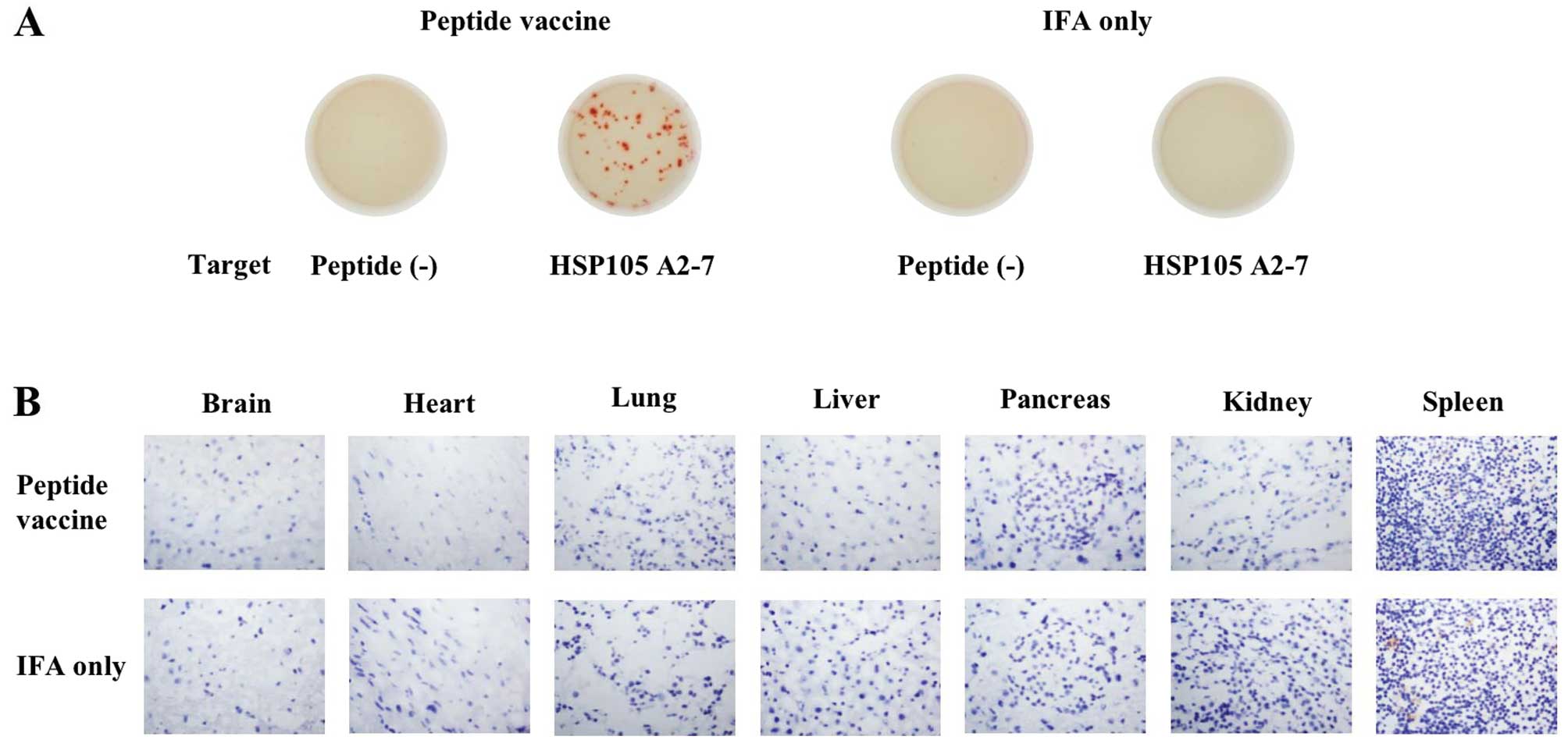
HSP105-derived peptide immunization does
not induce autoimmunity in HLA-A2 Tgm
HSP105 in normal adult mice is expressed in only
certain tissues, and expression in these tissues is less than that
in C26 tumor cells, suggesting a low risk of damage to normal
tissues posed by HSP105 antigen-induced immune responses (6). To investigate whether immunization of
the mice with HSP105-derived peptides causes autoimmunity, HLA-A2
Tgm were immunized with the HSP105 A2-7 and A2-12 peptides
emulsified in incomplete Freund’s adjuvant at 7-day intervals and
then sacrificed 7 days after the second vaccination. Using the
IFN-γ ELISPOT assay, we confirmed the induction of HSP105
peptide-specific CTLs in the spleen cells of immunized mice
(Fig. 4A). We did not detect any
pathological changes, such as CD8+ lymphocyte
infiltration or tissue destruction/repair, in the brain, heart,
lung, liver, pancreas, or kidney of HLA-A2 Tgm (Fig. 4B). These results indicate that the
HSP105 peptide-reactive CD8+ CTLs did not attack the
healthy tissue specimens that we evaluated.
Discussion
Heat shock proteins (HSPs) have essential functions
in the regulation of protein folding, conformation, assembly and
sorting. They function as molecular chaperones to maintain the
native conformational states of proteins, preventing protein
aggregation (18). HSPs are
classified into several families based on their molecular weight,
including HSP105/110, HSP90, HSP70, HSP60, HSP40 and HSP27
(19). HSP105 is a stress protein
within the HSP105/110 family that we previously reported to be
overexpressed in a variety of human cancers but with little to no
expression in normal tissues, aside from the testis. Thus, HSP105
presents a promising candidate for a target antigen in cancer
immunotherapy (3–7). In particular, HSP105 is specifically
overexpressed in colorectal cancer (83%) (4). Furthermore, HSP105 is expressed in
highly metastatic colon cancer cell lines and its expression is
correlated with advanced clinical cancer stages and positive lymph
node involvement (20). When
considering immunogenic target molecules for cancer immunotherapy,
it is important to select a tumor antigen that does not run the
risk of becoming lost during immunoediting (21). We reported previously that
siRNA-mediated suppression of HSP105 protein expression induced
apoptosis in various types of cancer cells, but not in fibroblasts
(12). Therefore, it is possible
that tumor cells do not lose HSP105 expression, allowing for
continued growth.
Advances in molecular biology and tumor immunology
have paved the way for identification of a large number of
tumor-associated antigens (TAAs) and antigenic peptides recognized
by tumor reactive CTLs; hence, peptide-based cancer immunotherapy
has become an intensely studied field (22,23).
Several HSPs, including HSP70, HSP90 and gp96, bind and deliver
(through receptor-mediated endocytosis of HSP) antigenic peptides
to the antigen-processing pathway of antigen-presenting cells
(APCs) and these peptides are then presented on major
histocompatibility complex (MHC) class I molecules. This
HSP-mediated pathway has been demonstrated to evoke potent
antiviral and antitumor immune responses (24). On the other hand, many researchers
have identified MHC class I-presenting peptide epitopes derived
from HSP (25). Furthermore, HSP105
itself may induce CD8+ T cells to become reactive
towards tumor cells that express HSP105, using HSP105-DNA and
HSP105-pulsed DC vaccines in mice (6–8).
We found 4 peptides [HSP105 A2-7 (RLMNDMTAV), HSP105
A2-12 (KLMSSNSTDL), HSP105 A24-1 (NYGI YKQDL) and HSP105 A24-7
(EYVYEFRDKL)] to be potential HSP105-derived, HLA-A2 or
A24-restricted CTL epitopes. There was a discrepancy between the
expected HSP105 CTL epitopes in Tgm and in PBMCs of colorectal
cancer patient. To identify the HSP105-derived CTL epitope
peptides, we analyzed the PBMC responses to each of the 4 peptides
in colorectal cancer patients using the ex vivo IFN-γ
ELISPOT assay.
In this study, we used an ex vivo assay to
detect HSP105-specific IFN-γ-secreting T cells in PBMCs from 14 of
21 pre-surgical patients with colorectal cancer. Generally, CTLs
specific for tumor antigens cannot be detected directly ex
vivo; rather only after expansion by repeated in vitro
stimulation with the antigenic peptide in the appropriate
antigen-presenting cells. This is attributed to assay sensitivity
and the low frequency of tumor antigen-specific CTLs (26). HSP105-specific CTLs in PBMCs, which
can be detected directly ex vivo without in vitro
stimulation, provide strong immunological evidence of
HSP105-derived CTL epitopes, which we were able to identify in this
study. However, because the prognosis of the pre-surgical patients
was affected by various factors, it was difficult to evaluate the
correlation between a positive CTL response before surgery and
clinical improvement at the present stage; an increase in the
number of patients at each stage and further analyses of this
relationship are necessary.
Although the SEREX method facilitated the
identification of tumor antigens that could be recognized by
antibodies and CD4+ T cells, few of their T-cell
epitopes have been determined (27). We previously reported in mice that
HSP105-DNA and HSP105-pulsed DC vaccines induced a reaction in
CD4+ T cells and CD8+ T cells towards tumor
cells expressing HSP105 (6–8). HSP105 was identified by SEREX
(3) and thus, HSP105-specific
CD4+ T cell reactions may be induced by HSP105
immunization. It was shown that antigen-specific CD4+ T
cells are required to activate memory CD8+ T cells into
fully functional effector killer cells (28). We are now preparing a clinical trial
to investigate HSP105-based immunotherapy for HSP105-expressing
tumors, including those from colorectal cancer. We plan to use the
HSP105 epitope peptides identified in this study as an initial
attempt. We expect that HSP105-based immunotherapy will be a novel
treatment strategy for colorectal cancer patients.
Acknowledgements
This study was supported by MEXT KAKENHI grant
numbers 12213111, 17015035 and the National Cancer Center Research
and Development Fund (25-A-7), as well as Health and Labor Science
Research Grants for Research on Hepatitis and Third Term
Comprehensive Control Research for Cancer from the Ministry of
Health, Labor and Welfare, Japan. Y.S. would like to thank the
Foundation for Promotion of Cancer Research (Japan) for the
Third-Term Comprehensive Control Research for Cancer for the award
of a research resident fellowship. T.N. is supported by funding
from MEDINET Co., Ltd.
References
|
1
|
Weitz J, Koch M, Debus J, Höhler T, Galle
PR and Büchler MW: Colorectal cancer. Lancet. 365:153–165. 2005.
View Article : Google Scholar
|
|
2
|
Mlecnik B, Tosolini M, Kirilovsky A, et
al: Histopathologic-based prognostic factors of colorectal cancers
are associated with the state of the local immune reaction. J Clin
Oncol. 29:610–618. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nakatsura T, Senju S, Yamada K, Jotsuka T,
Ogawa M and Nishimura Y: Gene cloning of immunogenic antigens
overexpressed in pancreatic cancer. Biochem Biophys Res Commun.
281:936–944. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kai M, Nakatsura T, Egami H, Senju S,
Nishimura Y and Ogawa M: Heat shock protein 105 is overexpressed in
a variety of human tumors. Oncol Rep. 10:1777–1782. 2003.PubMed/NCBI
|
|
5
|
Hatayama T, Yasuda K and Nishiyama E:
Characterization of high-molecular-mass heat shock proteins and 42
degrees C-specific heat shock proteins of murine cells. Biochem
Biophys Res Commun. 204:357–365. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miyazaki M, Nakatsura T, Yokomine K, Senju
S, Monji M, Hosaka S, et al: DNA vaccination of HSP105 leads to
tumor rejection of colorectal cancer and melanoma in mice through
activation of both CD4 T-cells and CD8 T-cells. Cancer Sci.
96:695–705. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yokomine K, Nakatsura T, Minohara M, et
al: Immunization with heat shock protein 105-pulsed dendritic cells
leads to tumor rejection in mice. Biochem Biophys Res Commun.
343:269–278. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yokomine K, Nakatsura T, Senju S, Nakagata
N and Minohara M: Regression of intestinal adenomas by vaccination
with heat shock protein 105-pulsed bone marrow-derived dendritic
cells in Apc(Min/+) mice. Cancer Sci. 98:1930–1935. 2007.PubMed/NCBI
|
|
9
|
Browning M and Krausa P: Genetic diversity
of HLA-A2: evolutionary and functional significance. Immunol Today.
17:165–170. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pascolo S, Bervas N, Ure JM, Smith AG,
Lemonnier FA and Pérarnau B: HLA-A2.1-restricted education and
cytolytic activity of CD8(+) T lymphocytes from beta2 microglobulin
(beta2m) HLA-A2.1 monochain transgenic H-2Db beta2m double knockout
mice. J Exp Med. 185:2043–2051. 1997.PubMed/NCBI
|
|
11
|
Firat H, Garcia-Pons F, Tourdot S, et al:
H-2 class I knockout, HLA-A2.1-transgenic mice: a versatile animal
model for preclinical evaluation of antitumor immunotherapeutic
strategies. Eur J Immunol. 29:3112–3121. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hosaka S, Nakatsura T, Tsukamoto H,
Hatayama T, Baba H and Nishimura Y: Synthetic small interfering RNA
targeting heat shock protein 105 induces apoptosis of various
cancer cells both in vitro and in vivo. Cancer Sci. 97:623–632.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Komori H, Nakatsura T, Senju S, et al:
Identification of HLA-A2- or HLA-A24-restricted CTL epitopes
possibly useful for glypican-3-specific immunotherapy of
hepatocellular carcinoma. Clin Cancer Res. 12:2689–2697. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakatsura T, Komori H, Kubo T, et al:
Mouse homologue of a novel human oncofetal antigen, glypican-3,
evokes T-cell-mediated tumor rejection without autoimmune reactions
in mice. Clin Cancer Res. 10:8630–8640. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakatsura T, Senju S, Ito M, Nishimura Y
and Itoh K: Cellular and humoral immune responses to a human
pancreatic cancer antigen, coactosin-like protein, originally
defined by the SEREX method. Eur J Immunol. 32:826–836. 2002.
View Article : Google Scholar
|
|
16
|
Yoshitake Y, Nakatsura T, Monji M, et al:
Proliferation potential-related protein, an ideal esophageal cancer
antigen for immunotherapy, identified using complementary DNA
microarray analysis. Clin Cancer Res. 10:6437–6448. 2004.
View Article : Google Scholar
|
|
17
|
Monji M, Nakatsura T, Senju S, et al:
Identification of a novel human cancer/testis antigen, KM-HN-1,
recognized by cellular and humoral immune responses. Clin Cancer
Res. 10:6047–6057. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feder ME and Hofmann GE: Heat-shock
proteins, molecular chaperones and the stress response:
evolutionary and ecological physiology. Annu Rev Physiol.
61:243–282. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Craig EA, Weissman JS and Horwich AL: Heat
shock proteins and molecular chaperones: mediators of protein
conformation and turnover in the cell. Cell. 78:365–372. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hwang TS, Han HS, Choi HK, et al:
Differential, stage-dependent expression of Hsp70, Hsp110 and Bcl-2
in colorectal cancer. J Gastroenterol Hepatol. 18:690–700. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kawakami Y and Rosenberg SA: Human tumor
antigens recognized by T-cells. Immunol Res. 16:313–339. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
van der Bruggen P, Traversari C, Chomez P,
et al: A gene encoding an antigen recognized by cytolytic T
lymphocytes on a human melanoma. Science. 254:1643–1647. 1991.
|
|
23
|
Kawakami Y, Eliyahu S, Delgado CH, et al:
Identification of a human melanoma antigen recognized by
tumor-infiltrating lymphocytes associated with in vivo tumor
rejection. Proc Natl Acad Sci USA. 91:6458–6462. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Srivastava P: Interaction of heat shock
proteins with peptides and antigen presenting cells: chaperoning of
the innate and adaptive immune responses. Annu Rev Immunol.
20:395–425. 2002. View Article : Google Scholar
|
|
25
|
Hickman-Miller HD and Hildebrand WH: The
immune response under stress: the role of HSP-derived peptides.
Trends Immunol. 25:427–433. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Romero P, Cerottini JC and Speiser DE:
Monitoring tumor antigen specific T-cell responses in cancer
patients and phase I clinical trials of peptide-based vaccination.
Cancer Immunol Immunother. 53:249–255. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jäger E, Chen YT, Drijfhout JW, et al:
Simultaneous humoral and cellular immune response against
cancer-testis antigen NY-ESO-1: definition of human
histocompatibility leukocyte antigen (HLA)-A2-binding peptide
epitopes. J Exp Med. 187:265–270. 1998.
|
|
28
|
Gao FG, Khammanivong V, Liu WJ, Leggatt
GR, Frazer IH and Fernando GJ: Antigen-specific
CD4+T-cell help is required to activate a memory
CD8+T-cell to a fully functional tumor killer cell.
Cancer Res. 62:6438–6441. 2002.PubMed/NCBI
|