Introduction
Gene therapy has been utilized in many clinical
trials for human cancer and has exhibited innovative and attractive
therapeutic potential. Adenovirus-mediated gene delivery continues
to be preferred for the treatment of cancer, as the vectors have a
high transduction efficacy for the therapeutic gene, and due to the
safety of the procedure when it is used for direct local injection
(1,2). A gene expression promoter, such as the
cytomegalovirus (CMV), Rous sarcoma virus (RSV) and CMV early
enhancer/chicken β-actin (CAG)promoters, has been used to increase
the gene expression in a variety of normal and cancer cells,
achieving favorable transduction efficiency (3–6).
However, in some patients, the clinical cancer gene therapy using
even these strong promoters has shown modest therapeutic effects
(1). Some of these unfavorable
results may be due to differences in the accomplishment of the
robust and sufficient expression of the therapeutic gene for the
treatment of the specific cancer type. Thus, particularly for
clinical trials of cancer gene therapy, a more powerful gene
expression system that can be used for a broad range of cancer
types is urgently required.
A variety of enhancer sequences have thus far been
reported and used for the modification and regulation of gene
expression (7–11). In order to develop a novel system
which enables more potent gene expression in comparison to the
conventional systems, we have been modifying the construct sequence
of plasmids by adding and exchanging the multiple known enhancer
sequences. In a series of modifications with multiple enhancers in
the CMV promoter-dependent gene expression cassette, we found that
the insertion of the triple translational enhancer sequences of
hTERT, SV40 and CMV downstream of the BGH polyA sequence led to the
most potent gene expression. The human telomerase reverse
transcriptase (hTERT) promoter/enhancer has been demonstrated to be
available for cancer tissue-specific gene expression in a broad
range of malignant cell types (12–14).
The Simian virus 40 (SV40) and CMV promoter/enhancer have also been
well characterized and have been used to improve the gene
expression in mammalian cells (3,4,7–10).
Since the novel gene expression system using the triple enhancers
significantly enhances the expression of the gene(s) of interest in
comparison to the conventional systems using the strong CMV
promoter, we termed this novel gene expression cassette the super
gene expression (SGE) system. We also call this cassette C-TSC (C,
CMV promoter driving; TSC, enhancer unit composed of triple tandem
enhancer sequences of hTERT, SV40 and CMV).
The reduced expression in immortalized cells (REIC)
gene is a tumor suppressor that is identical to the Dickkopf-3
(Dkk-3) gene (15). The expression
of REIC/Dkk-3 is significantly downregulated in a wide range of
human cancer types, making REIC/Dkk-3 a promising cancer
therapeutic gene (16–23). Expecting that transfection of
REIC/Dkk-3 would provide therapeutic effects as a tumor suppressor
gene, we previously developed an adenoviral vector expressing the
human REIC/Dkk-3 gene (Ad-REIC) and demonstrated that the agent
induced apoptosis in various cancer cell lines (16–18,23).
In order to demonstrate the utility of the SGE system for cancer
gene therapy, we developed an Ad-REIC vector with the SGE system
(Ad-SGE-REIC). We then compared the Ad-SGE-REIC with the
conventional Ad-REIC vectors, and herein report our evaluation and
validation of the SGE system for the enhancement of gene expression
and indicate its potential therapeutic utility.
Materials and methods
Cells and cell culture
Normal human hepatocytes (HC) were purchased from
Applied Cell Biology Research Institute (Kirkland, WA, USA). Normal
human fibroblasts (OUMS24) were established by the Department of
Cell Biology at our university (15). All human cancer cell lines (PC3,
prostate cancer; KPK1, renal cancer; 211H, malignant mesothelioma;
HeLa, cervical cancer), the mouse renal cell carcinoma line (RENCA
cells) and the HEK293 cell line were provided by the American Type
Culture Collection (Rockville, MD, USA), and these cells were
cultivated as previously described (16,21).
Construction of plasmid vectors
In the pShuttle plasmid vector system driven by the
CMV promoter (Clontech Laboratories, Inc., Mountain View, CA, USA),
we inserted the REIC/Dkk-3 gene as the gene of interest and
constructed the pShuttle-REIC plasmid. The full-length cDNA of the
human REIC/Dkk3 gene was amplified by PCR with primers containing
XbaI and KpnI restriction sites. The PCR product was
cloned into the pShuttle vector to generate pShuttle-REIC. To
construct the pShuttle-SGE-REIC plasmid, we inserted the sequences
of the triple translational enhancers of hTERT, SV40 and CMV
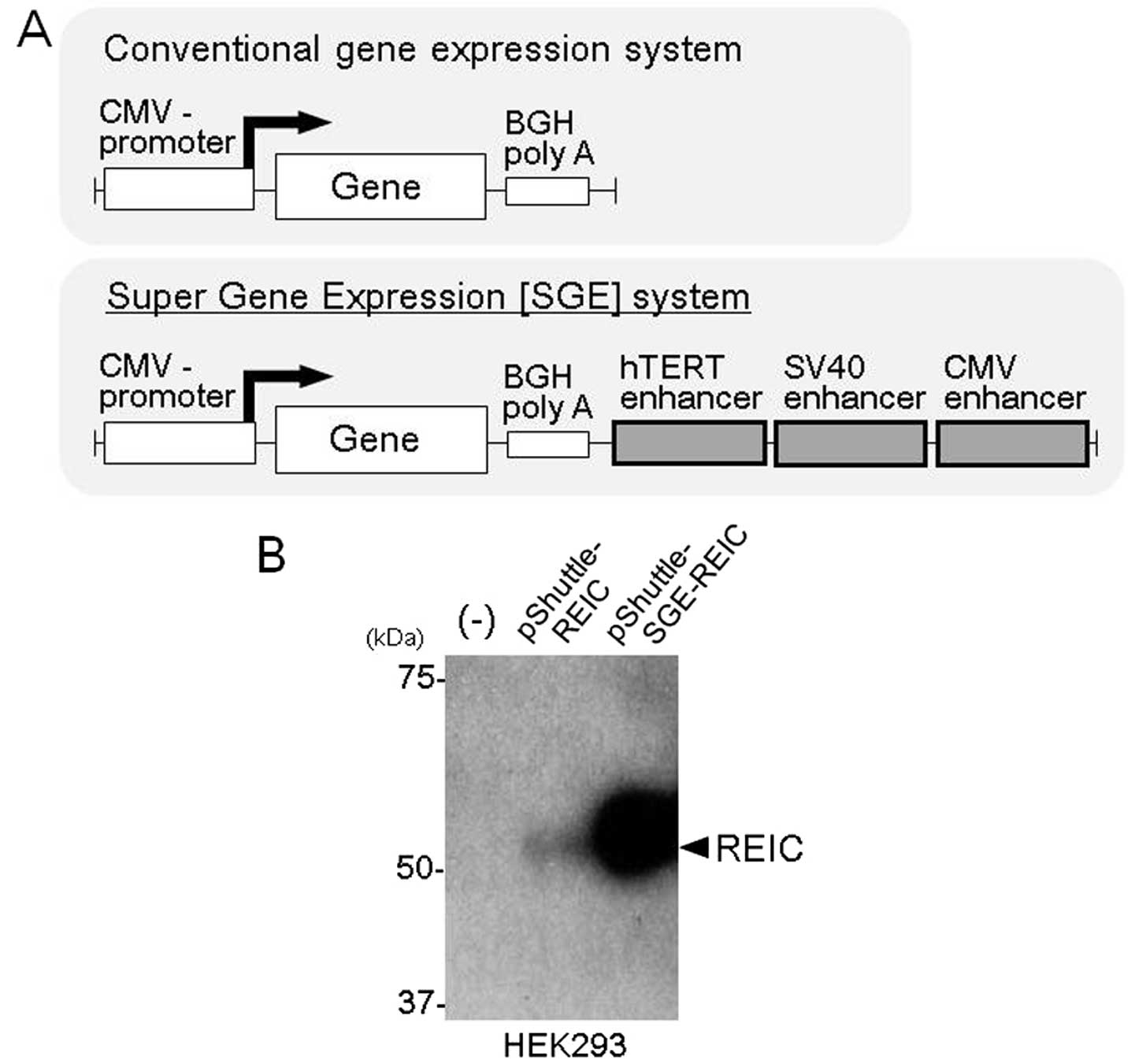
downstream of the BGH polyA sequence (Fig. 1A) of the pShuttle-REIC vector. The
tandem sequences of the hTERT enhancer [189 bp: accession no.
DQ264729 (1618–1806)], SV40 enhancer [319 bp: accession no.
AY864928 (2156–2474)] and CMV enhancer [479 bp: accession no.
AJ318513 (159–637)] were artificially synthesized and cloned into
pShuttle-REIC at the restriction site between KpnI and
EcoRI.
Construction and production of adenoviral
vectors
In order to generate CMV promoter-driven Ad-CMV-REIC
and Ad-SGE-REIC vectors, pShuttle-REIC and pShuttle-SGE-REIC
plasmids were digested with the restriction enzymes I-CeuI
and PI-SceI and inserted into the Adeno-X Viral DNA
(Clontech Laboratories, Inc.). The recombinant adenoviral DNA with
the full-length human REIC/Dkk-3 gene was linearized by PacI
digestion and transfected into HEK293 cells. Seven to ten days
after transfection, the HEK293 cells were harvested, and a viral
solution was obtained by three freeze/thaw cycles. The recovered
virus solution was used to propagate sufficient viruses in HEK293
cells for further studies in other cell lines. All virus particles
were purified by CsCl density gradient ultracentrifugation and
stored at −80°C. For Ad-CAG-REIC under the control of the CAG
promoter (CMV early enhancer/chicken β-actin promoter), the
full-length human REIC/Dkk-3 gene was inserted into the cosmid
vector, pAxCAwt, and then transferred into an adenoviral vector
using the COS-TPC method (Takara Bio, Inc., Shiga, Japan) (16,17).
An adenoviral vector carrying the LacZ gene with a CAG promoter
(Ad-LacZ) was used as the control vector. These adenoviral vectors
were generated using replication-defective adenoviruses of serotype
5.
Western blot analysis
The cells (5.0×105 cells) were plated in
flat-bottom 6-well plates in complete culture medium, and after a
24-h incubation, transient transfections with the plasmid or
adenoviral vectors were performed. During the plasmid transfection,
HEK293 cells were transfected with the FuGENE HD reagent (Promega,
Madison, WI, USA) and the cells were sampled 24 h after
transfection. During the transfection with adenoviral vectors, the
cells were treated with Ad-LacZ and Ad-REIC at the indicated
multiplicity of infection (MOI) in 0.3 ml of complete medium for 1
h. Then, 1.7 ml of fresh medium was added and the cells were
incubated for another 24 h. After the incubation, the attached
cells were sampled, and a western blot analysis was performed as
previously described (18,21). A mouse monoclonal anti-human
REIC/Dkk-3 antibody raised in our laboratory (1:1,000) was used as
the primary antibody.
Apoptosis assay
The cells (5.0×105 cells) were seeded in
6-well plates and incubated in culture medium for 24 h. The cells
were then treated with Ad-LacZ and Ad-REIC at the indicated MOI, as
described above. After 48 or 72 h of incubation, the apoptotic
cells were visualized by Hoechst 33342 staining, and the apoptotic
rate was analyzed as previously described (18,23).
Subcutaneous tumor models and
treatments
Mouse RENCA renal cell carcinoma cells
[2×105 cells/100 μl phosphate-buffered saline (PBS)] and
human PC3 prostate cancer cells (2×106 cells/100 μl PBS)
were subcutaneously injected into the right thigh of female BALB/C
mice and male athymic nude mice, respectively. The tumors were
allowed to reach ~8 mm in diameter, and the mice were then randomly
assigned to treatment groups of four or five animals each. In the
tumor model with RENCA cells, there were five groups treated with a
single intratumoral injection of: i) PBS (100 μl) or ii) Ad-LacZ,
iii) Ad-CMV-REIC, iv) Ad-CAG-REIC or v) Ad-SGE-REIC adenoviral
vectors at a dose of 5×109 viral particles/100 μl
PBS/injection. In the PC3 tumor model, there were four groups
treated with a single injection or three intratumoral injections
of: i) PBS (100 μl) single injection or ii) Ad-CAG-REIC single
injection, iii) Ad-SGE-REIC single injection or iv) three
injections of Ad-SGE-REIC on days 0, 2 and 4 at a dose of
1×1010 viral particles/100 μl PBS/injection. The tumor
volume was calculated as previously described (18,19).
Statistical analysis
The data are shown as the means ± standard error. An
analysis of variance (ANOVA) was performed to determine the
statistical significance of differences in the apoptosis rates.
Unpaired Student’s t-tests were performed for the statistical
analyses of the tumor volumes. Differences were considered to be
statistically significant at P<0.05.
Results
The SGE system significantly enhances the
REIC/Dkk-3 gene expression by the plasmid and adenoviral
vectors
In order to demonstrate the enhanced gene expression
induced by the SGE system, we constructed a pShuttle plasmid vector
with the SGE system encoding the REIC/Dkk-3 gene
(pShuttle-SGE-REIC) and compared the expression with that of the
pShuttle-REIC plasmid vector with the conventional gene expression
system. The REIC/Dkk-3 expression level after transfection with the
pShuttle-SGE-REIC plasmid was significantly upregulated in
comparison to that of the pShuttle-REIC in a western blot analysis
of HEK293 cells (Fig. 1B). To
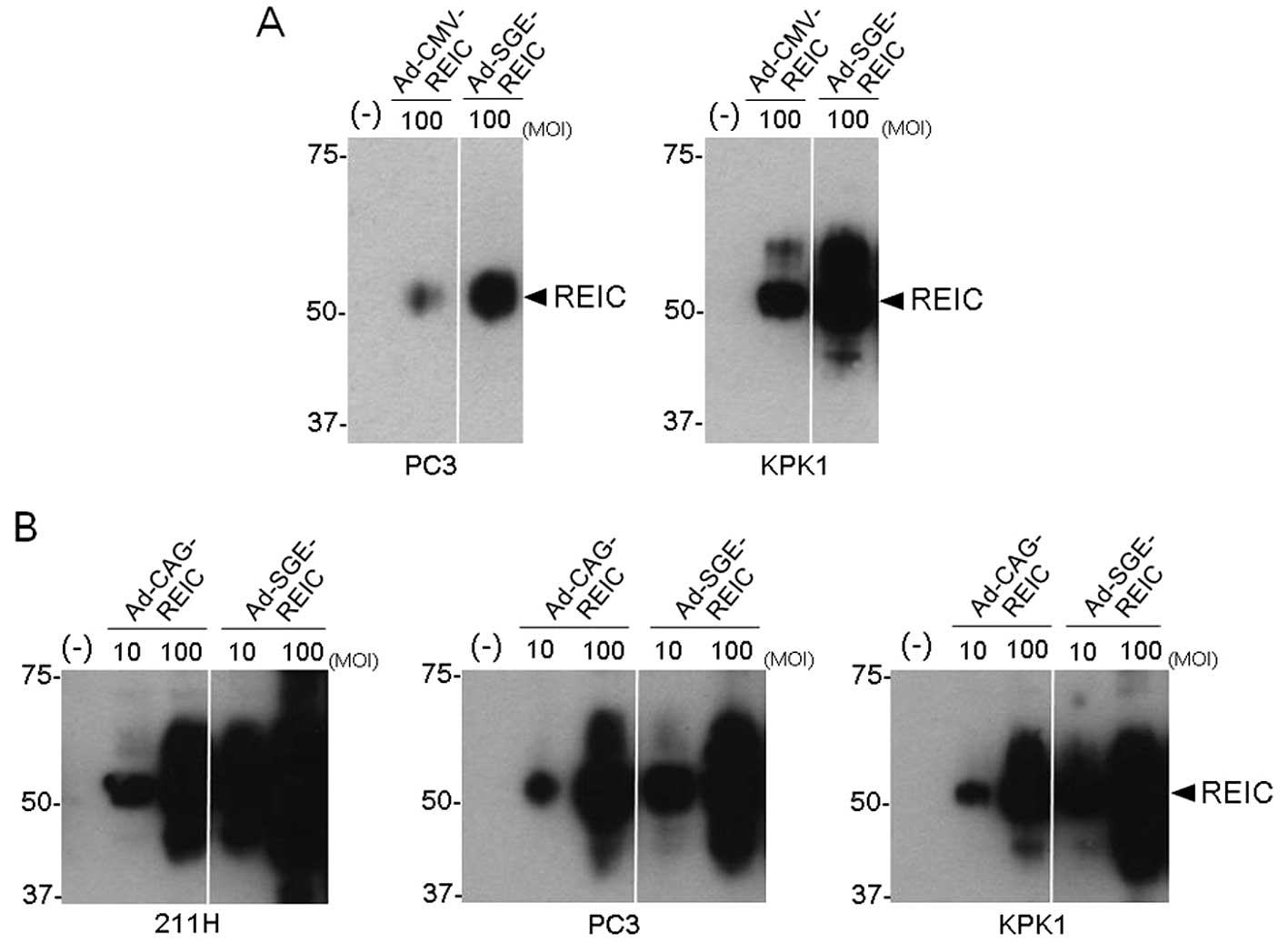
further verify the broad utility of the SGE system, we put the SGE
cassette in adenoviral vectors and developed an adenoviral vector
expressing the REIC/Dkk-3 gene based on the CMV promoter-driven SGE
system (Ad-SGE-REIC). We compared the REIC/Dkk-3 expression levels
between the CMV promoter-driven adenoviral vectors (Ad-CMV-REIC and
Ad-SGE-REIC) and demonstrated significantly enhanced REIC/Dkk-3
gene expression by the SGE system in the human cancer cells
(Fig. 2A). In addition, in the
comparison between the Ad-CAG-REIC and Ad-SGE-REIC vectors by a
western blot analysis in multiple human cancer cell lines, robust
upregulation of REIC/Dkk-3 expression was observed in the
Ad-SGE-REIC-transfected cells, particularly after the transfection
at 10 MOI (Fig. 2B).
Significantly enhanced apoptosis is
induced in various human cancer cell lines by Ad-SGE-REIC
treatment
To examine the therapeutic utility of the SGE system
and Ad-SGE-REIC vector in human cancer cells, we assessed the in
vitro apoptotic effects of Ad-SGE-REIC in comparison to those
of the Ad-CMV-REIC and Ad-CAG-REIC vectors using the conventional
gene expression system. The Ad-SGE-REIC vector significantly
enhanced the in vitro apoptosis induction in the various
human cancer cells in comparison to the control vectors transfected
at 50 MOI for 48 h (Fig. 3A and B).
In the normal HC and fibroblasts (OUMS24), there was no significant
apoptosis induction by any of the Ad-REIC vectors, as was
previously reported for the Ad-CAG-REIC vector (16,18,20).
Under the other experimental condition of 100 MOI for 72 h,
significant apoptosis induction was confirmed following Ad-CMV-REIC
and Ad-CAG-REIC treatment in comparison to the Ad-LacZ treatment
(Fig. 3C). In addition, under these
conditions, the induction of apoptosis was significantly enhanced
in Ad-SGE-REIC-transfected cells compared to the cells transfected
with conventional Ad-REIC vectors.
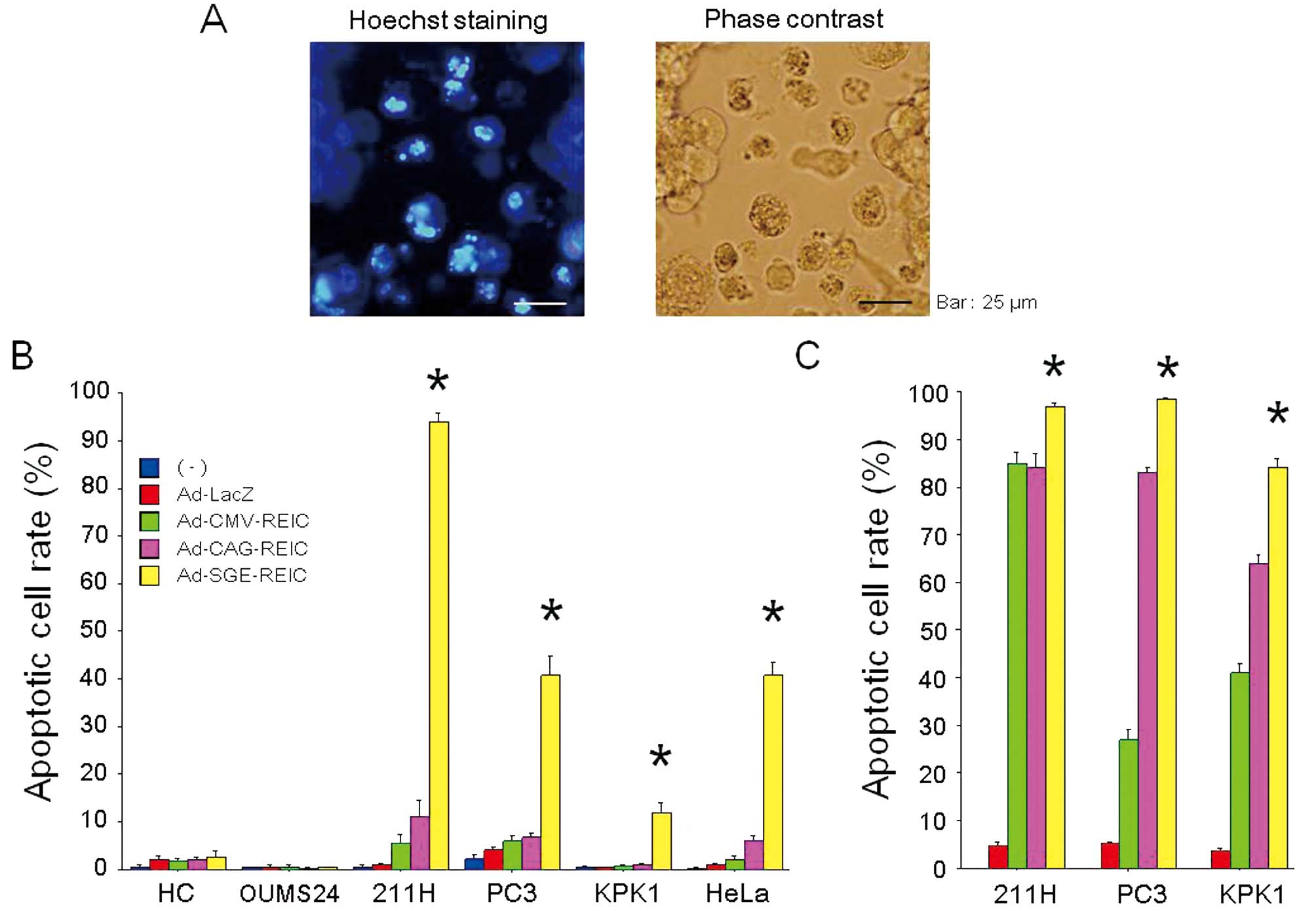 | Figure 3(A) The induction of apoptosis after
Ad-REIC treatment is shown by Hoechst 33342 staining in 211H human
malignant mesothelioma cells. Apoptotic cells can be clearly
observed as bright cells under fluorescence microscopy (left
panel). The appearance of the cells by phase contrast microscopy is
also shown (right panel). (B) The apoptotic cell rate (%) was
examined after the indicated treatments (no treatment, Ad-LacZ,
Ad-CMV-REIC, Ad-CAG-REIC and Ad-SGE-REIC at 50 MOI for 48 h) in
normal human cells (HC, hepatocytes; OUMS24, fibroblasts) and
various human cancer cell lines (211H, PC3, KPK1 and HeLa). The
apoptotic cell rate was determined in five different fields under
microscopic observations. *A significant difference was
observed between the Ad-SGE-REIC group and the other treatment
groups. (C) The apoptotic cell rate (%) was examined under
different experimental conditions (Ad-LacZ, Ad-CMV-REIC,
Ad-CAG-REIC or Ad-SGE-REIC at a 100 MOI for 72 h) in the human
cancer cells (211H, PC3 and KPK1). *A significant
difference was observed between the Ad-SGE-REIC group and the other
treatment groups. SGE, super gene expression; REIC, reduced
expression in immortalized cells; CMV, cytomegalovirus; MOI,
multiplicity of infection. |
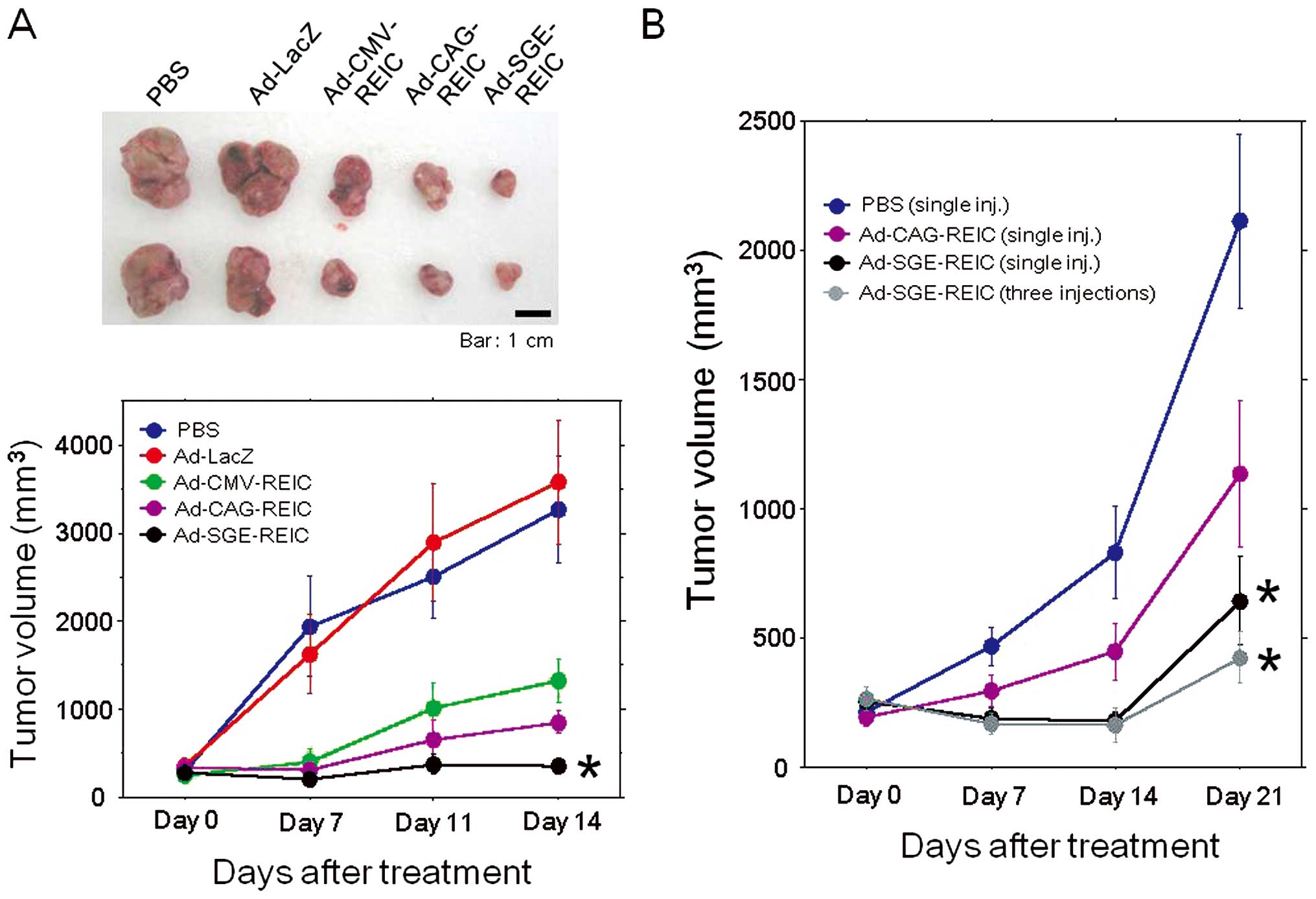
Intratumoral Ad-SGE-REIC administration
strongly inhibits the tumor growth in subcutaneous mouse tumor
models
To further evaluate the therapeutic utility of the
SGE system and Ad-SGE-REIC vector, the inhibitory effects of
Ad-REIC treatment on the subcutaneous tumor growth were analyzed in
mouse tumor models. In both the mouse renal cell carcinoma (RENCA
cells) (Fig. 4A) and human prostate
cancer (PC3 cells) (Fig. 4B)
models, strong suppression of tumor growth was observed in the
Ad-SGE-REIC-treated groups in comparison to the other treatment
groups. These results are consistent with the findings of the in
vitro studies comparing the REIC/Dkk-3 gene expression and
apoptosis induction by the three types of Ad-REIC treatment. Thus,
the novel SGE system significantly enhances the therapeutic
antitumor effects in mouse tumor models, and the Ad-SGE-REIC vector
was superior to the conventional Ad-CMV-REIC and Ad-CAG-REIC
vectors in terms of the efficacy of in vivo intratumoral
gene therapy.
Discussion
Gene therapy-based approaches often require strong
levels of the gene expression and protein products. We have been
developing methods to enhance the gene transcription driven by
various promoters, and herein described a novel strategy to achieve
higher levels of gene expression. We developed the SGE system,
which enables the powerful gene expression, by putting the triple
translational enhancer sequences of hTERT, SV40 and CMV downstream
of the BGH polyA sequence. We herein demonstrated that the plasmid
vector and adenoviral vector systems with a CMV promoter-driven SGE
cassette can significantly enhance the expression of the CMV
promoter, which is one of the strongest promoters reported
(7,8).
The combination of multiple promoter/enhancer
elements has been attempted in previous studies, with several
showing improved gene transcription. The hTERT promoter/enhancer is
well-characterized and has been frequently used for cancer-specific
gene expression (2,12–14).
In one of the studies using hTERT promoter/enhancer-driven plasmid
vectors, the combination of the hTERT promoter with multiple hTERT
enhancer regions significantly enhanced the gene expression in
comparison to that of the parental plasmid (11). With regard to the SV40 enhancer,
several studies demonstrated enhanced gene expression by the
insertion of the SV40 enhancer downstream of the polyA sequences
(7–10). The CMV enhancer is used in the CMV
early enhancer/chicken β-actin promoter (CAG promoter), which is
known to improve the gene expression in various cell types and
tissues (3,4,7,8). The
CMV enhancer is also used to stimulate the CMV promoter, EF-1α
promoter and ubiquitin promoter, thereby further increasing the
expression of the gene of interest (7,8,24).
Therefore, each enhancer (hTERT, SV40 and CMV) can positively
stimulate the promoters in a variety of vector systems and improve
the gene expression levels in mammalian cells. The present study
confirmed the utility of using the combination of the hTERT, SV40
and CMV enhancers in this order in the gene expression construct.
The mechanism underlying the upregulated gene transcription by the
hybrid promoter/enhancer system is not yet fully understood. It is
conceivable that the transcriptional elements of the hTERT, SV40
and CMV triple enhancers interact individually or synergistically
with the CMV promoter that drives the gene transcriptional
complexes and upregulates the capacity of the cells for gene
expression.
The REIC/Dkk-3 gene is a tumor suppressor and
promising cancer therapeutic gene (16–23).
To examine the possible use of REIC/Dkk-3 for targeted gene-based
therapy, we developed a replication-deficient adenoviral vector
encoding the human REIC/Dkk-3 gene (Ad-REIC) (16). The CAG promoter was selected to
drive the REIC/Dkk-3 expression as it enables strong gene
expression (3,4,7,8). The
overexpression of REIC/Dkk-3 with the Ad-CAG-REIC reagent was
previously found to induce apoptosis in a broad range of human
cancer cell lines in vitro (16–18,23).
On the other hand, the ability of Ad-CAG-REIC to induce apoptosis
was lower in the non-malignant cells (16,18,20).
These findings indicate that the Ad-CAG-REIC selectively induces
apoptosis in a cancer cell-specific manner, and this was also
demonstrated in the present study for the other Ad-REIC reagents
(Ad-CMV-REIC and Ad-SGE-REIC). Since REIC/Dkk-3 expression is
significantly downregulated in several human cancer cells, but is
typically expressed in non-malignant or normal cells (15,16,18,21),
the endogenous expression levels of the protein seem to be
correlated with the sensitivity of the cells against the REIC/Dkk-3
overexpression by Ad-REIC.
In addition, the abundant REIC/Dkk-3 protein
expression and secretion after intratumoral Ad-REIC administration
could provide anticancer immunological effects via a mechanism
involving autologous cancer vaccination (19,20).
To further improve the therapeutic outcomes, we developed a new
adenoviral vector expressing the REIC/Dkk-3 gene, based on the CMV
promoter-SGE system (Ad-SGE-REIC), and compared the therapeutic
utility of Ad-SGE-REIC with that of the conventional adenoviral
vectors (Ad-CMV-REIC or Ad-CAG-REIC). In all the cancer cell lines
tested in this study, the Ad-SGE-REIC was superior to conventional
adenoviral systems in terms of the REIC/Dkk-3 protein expression
and in vitro induction of apoptosis. With regard to the
in vivo therapeutic effects, the antitumor effects of the
Ad-SGE-REIC vector were also superior to those of the control
adenoviral vectors, indicating the potential for promising
therapeutic outcomes in future REIC/Dkk-3 gene therapy using
Ad-SGE-REIC. Thus, the SGE system could be successfully applied to
the adenovirus-mediated gene expression system, and might be useful
as gene therapy for a wide range of human malignancies.
We herein demonstrated that the CMV promoter-driven
SGE cassette is superior to the conventional systems with CMV and
CAG promoters in terms of the vector-mediated gene expression. The
novel hybrid promoter/enhancer system is a promising gene
therapeutic cassette that can be used in combination with various
vector systems, and has important practical implications for cancer
gene therapy. We believe that this novel gene expression system
will become an innovative tool in the fields of both gene
expression and gene therapy.
Acknowledgements
This study was supported by scientific research
grants (KAKENHI: 22791473, 23390382 and 25462479) from the Ministry
of Education, Culture, Sports, Science and Technology of Japan. We
thank Ms. Sabina Mahmood and Ms. Fusaka Oonari (Okayama University)
for their valuable assistance. Okayama University and Momotaro-Gene
Inc. are applying for patents on the SGE systems. Drs M.W., M.S.,
N.H.H. and H.K. are the inventors of the patents and own stock in
Momotaro-Gene Inc.
References
|
1
|
Nasu Y, Saika T, Ebara S, Kusaka N, Kaku
H, Abarzua F, Manabe D, Thompson TC and Kumon H: Suicide gene
therapy with adenoviral delivery of HSV-tK gene for patients with
local recurrence of prostate cancer after hormonal therapy. Mol
Ther. 15:834–840. 2007.PubMed/NCBI
|
|
2
|
Fukazawa T, Matsuoka J, Yamatsuji T, Maeda
Y, Durbin ML and Naomoto Y: Adenovirus-mediated cancer gene therapy
and virotherapy (Review). Int J Mol Med. 25:3–10. 2010.PubMed/NCBI
|
|
3
|
Niwa H, Yamamura K and Miyazaki J:
Efficient selection for high-expression transfectants with a novel
eukaryotic vector. Gene. 108:193–199. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Araki K, Araki M, Miyazaki J and Vassalli
P: Site-specific recombination of a transgene in fertilized eggs by
transient expression of Cre recombinase. Proc Natl Acad Sci USA.
92:160–164. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ebara S, Shimura S, Nasu Y, Kaku H, Kumon
H, Yang G, Wang J, Timme TL, Aguilar-Cordova E and Thompson TC:
Gene therapy for prostate cancer: toxicological profile of four
HSV-tk transducing adenoviral vectors regulated by different
promoters. Prostate Cancer Prostatic Dis. 5:316–325. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Watanabe M, Nasu Y, Kashiwakura Y, Kusumi
N, Tamayose K, Nagai A, Sasano T, Shimada T, Daida H and Kumon H:
Adeno-associated virus 2-mediated intratumoral prostate cancer gene
therapy: long-term maspin expression efficiently suppresses tumor
growth. Hum Gene Ther. 16:699–710. 2005. View Article : Google Scholar
|
|
7
|
Xu ZL, Mizuguchi H, Ishii-Watabe A, Uchida
E, Mayumi T and Hayakawa T: Optimization of transcriptional
regulatory elements for constructing plasmid vectors. Gene.
272:149–156. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu ZL, Mizuguchi H, Ishii-Watabe A, Uchida
E, Mayumi T and Hayakawa T: Strength evaluation of transcriptional
regulatory elements for transgene expression by adenovirus vector.
J Control Release. 81:155–163. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Young JL, Benoit JN and Dean DA: Effect of
a DNA nuclear targeting sequence on gene transfer and expression of
plasmids in the intact vasculature. Gene Ther. 10:1465–1470. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Song JS: Enhanced expression of apoptin by
the Myc-Max binding motif and SV40 enhancer for SCLC gene therapy.
Biosci Biotechnol Biochem. 69:51–55. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim SJ, Lee HS, Shin JH, Kim CG, Jeong S,
Park K, Choe H and Lee H: Preferentially enhanced gene expression
from a synthetic human telomerase reverse transcriptase promoter in
human cancer cells. Oncol Rep. 16:975–979. 2006.PubMed/NCBI
|
|
12
|
Kojima T, Hashimoto Y, Watanabe Y, Kagawa
S, Uno F, Kuroda S, Tazawa H, Kyo S, Mizuguchi H, Urata Y, Tanaka N
and Fujiwara T: A simple biological imaging system for detecting
viable human circulating tumor cells. J Clin Invest. 119:3172–3181.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Watanabe M, Ueki H, Ochiai K, Huang P,
Kobayashi Y, Nasu Y, Sasaki K, Kaku H, Kashiwakura Y and Kumon H:
Advanced two-step transcriptional amplification as a novel method
for cancer-specific gene expression and imaging. Oncol Rep.
26:769–775. 2011.PubMed/NCBI
|
|
14
|
Ueki H, Watanabe M, Kaku H, Huang P, Li
SA, Ochiai K, Hirata T, Noguchi H, Yamada H, Takei K, Nasu Y,
Kashiwakura Y and Kumon H: A novel gene expression system for
detecting viable bladder cancer cells. Int J Oncol. 41:135–140.
2012.PubMed/NCBI
|
|
15
|
Tsuji T, Miyazaki M, Sakaguchi M, Inoue Y
and Namba M: A REIC gene shows down-regulation in human
immortalized cells and human tumor-derived cell lines. Biochem
Biophys Res Commun. 268:20–24. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Abarzua F, Sakaguchi M, Takaishi M, Nasu
Y, Kurose K, Ebara S, Miyazaki M, Namba M, Kumon H and Huh NH:
Adenovirus-mediated overexpression of REIC/Dkk-3 selectively
induces apoptosis in human prostate cancer cells through activation
of c-Jun-NH2-kinase. Cancer Res. 65:9617–9622. 2005. View Article : Google Scholar
|
|
17
|
Kashiwakura Y, Ochiai K, Watanabe M,
Abarzua F, Sakaguchi M, Takaoka M, Tanimoto R, Nasu Y, Huh NH and
Kumon H: Down-regulation of inhibition of differentiation-1 via
activation of activating transcription factor 3 and Smad regulates
REIC/Dickkopf-3-induced apoptosis. Cancer Res. 68:8333–8341. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kawasaki K, Watanabe M, Sakaguchi M,
Ogasawara Y, Ochiai K, Nasu Y, Doihara H, Kashiwakura Y, Huh NH,
Kumon H and Date H: REIC/Dkk-3 overexpression downregulates
P-glycoprotein in multidrug-resistant MCF7/ADR cells and induces
apoptosis in breast cancer. Cancer Gene Ther. 16:65–72. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Watanabe M, Kashiwakura Y, Huang P, Ochiai
K, Futami J, Li SA, Takaoka M, Nasu Y, Sakaguchi M, Huh NH and
Kumon H: Immunological aspects of REIC/Dkk-3 in monocyte
differentiation and tumor regression. Int J Oncol. 34:657–663.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sakaguchi M, Kataoka K, Abarzua F,
Tanimoto R, Watanabe M, Murata H, Than SS, Kurose K, Kashiwakura Y,
Ochiai K, Nasu Y, Kumon H and Huh NH: Overexpression of REIC/Dkk-3
in normal fibroblasts suppresses tumor growth via induction of
interleukin-7. J Biol Chem. 284:14236–14244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang K, Watanabe M, Kashiwakura Y, Li SA,
Edamura K, Huang P, Yamaguchi K, Nasu Y, Kobayashi Y, Sakaguchi M,
Ochiai K, Yamada H, Takei K, Ueki H, Huh NH, Li M, Kaku H, Na Y and
Kumon H: Expression pattern of REIC/Dkk-3 in various cell types and
the implications of the soluble form in prostatic acinar
development. Int J Oncol. 37:1495–1501. 2010.PubMed/NCBI
|
|
22
|
Ochiai K, Watanabe M, Ueki H, Huang P,
Fujii Y, Nasu Y, Noguchi H, Hirata T, Sakaguchi M, Huh NH,
Kashiwakura Y, Kaku H and Kumon H: Tumor suppressor REIC/Dkk-3
interacts with the dynein light chain, Tctex-1. Biochem Biophys Res
Commun. 412:391–395. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hirata T, Watanabe M, Kaku H, Kobayashi Y,
Yamada H, Sakaguchi M, Takei K, Huh NH, Nasu Y and Kumon H:
REIC/Dkk-3-encoding adenoviral vector as a potentially effective
therapeutic agent for bladder cancer. Int J Oncol. 41:559–564.
2012.PubMed/NCBI
|
|
24
|
Wang CY, Guo HY, Lim TM, Ng YK, Neo HP,
Hwang PY, Yee WC and Wang S: Improved neuronal transgene expression
from an AAV-2 vector with a hybrid CMV enhancer/PDGF-beta promoter.
J Gene Med. 7:945–955. 2005. View
Article : Google Scholar : PubMed/NCBI
|